Dietary Sources and Bioactivities of Melatonin




Abstract
:1. Introduction
2. Dietary Sources of Melatonin
2.1. Animal Foods
2.2. Plant Foods
2.2.1. Cereals
2.2.2. Fruits
2.2.3. Vegetables
2.2.4. Legumes and Seeds (Raw and Germinated)
2.2.5. Nuts
2.2.6. Juices and Beverages
2.2.7. Medical Herbs
2.2.8. Edible Oils
2.2.9. Yeast
2.3. Other Issues about Melatonin Intake
2.3.1. The Alternation of Endogenous Melatonin along Life Cycle
2.3.2. Bioavailability of Exogenous Melatonin
2.3.3. Benefits of Consuming Melatonin-Containing Foods
2.3.4. Guidance on Regulating Dietary Supplement of Melatonin
3. Bioactivities of Melatonin
3.1. Antioxidant Activities
3.2. Anti-Inflammatory Activities
3.2.1. NF-κB Signaling Pathway Involved Mechanisms
3.2.2. SIRT1 Pathway Involved Mechanisms
3.3. Enhancing Immune Activities
3.4. Improving Circadian Rhythm and Sleep
3.5. Anticancer Activities
3.5.1. Effects on Tumor Cell Cycle, in Terms of Growth, Proliferation, Metabolism and Apoptosis
3.5.2. Effects on Invasion and Metastasis of Tumor Cells
3.5.3. Therapy Adjunct in Tumor Treatment
3.6. Cardiovascular Protection
3.7. Anti-Diabetic Activities
3.8. Anti-Obese Activities
3.9. Neuroprotective Activities
3.10. Other Bioactivities
3.11. Adverse Effects
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| 2-OHM | 2-hydroxymelatonin |
| 4-OHM | 4-hydroxymelatonin |
| 6-OHM | 6-hydroxymelatonin |
| AAAD | aromatic L-amino acid decarboxylase |
| Aβ | amyloid-beta |
| ACC | acetyl-CoA carboxylase |
| AD | Alzheimer’s disease |
| AD-MSCs | adipose-derived mesenchymal stem cells |
| AFMK | N1-acetyl-N2-formyl-5-methoxykynuramine |
| AMK | N1-acetyl-5-methoxykynuramine |
| AMPK | adenosine monophosphate-activated protein kinase |
| AP-2β | activator protein 2β |
| ARE | antioxidant responsive element |
| ATF4 | activating transcription factor 4 |
| BACE1 | β-APP-cleaving enzyme 1 |
| Bax | Bcl-2-associated X protein |
| BBB | blood-brain barrier |
| Bcl-2 | B-cell lymphoma 2 |
| BDNF | brain derived neurotrophic factor |
| bFGF | basic fibroblast growth factor |
| BMMNCs | bone marrow mononuclear cells |
| BM-MSCs | bone marrow mesenchymal stem cells |
| C3-OHM | cyclic 3-hydroxymelatonin |
| CaMKIIα | calmodulin dependent protein kinase II alpha |
| cAMP | cyclic AMP |
| CAT | catalase |
| CCL20 | chemokine C-C motif ligand 20 |
| cGMP | cyclic GMP |
| CNS | central nerve system |
| COX-2 | cyclo-oxygenase 2 |
| CVDs | cardiovascular diseases |
| CXCL1 | chemokine C-X-C motif ligand 1 |
| cyt c | cytochrome c |
| DAG | diacylglycerol |
| DS | diclofenac sodium |
| DW | dry weight |
| EBPβ | enhancer-binding protein beta |
| eIF2α | eukaryotic initiation factor 2α |
| EMF | electromagnetic fields |
| EMT | epithelial mesenchymal transition |
| eNOS | endothelial nitric oxide synthase |
| ER | endoplasmic reticulum |
| ERK | extracellular signal-regulated kinase |
| ERR-α | estrogen-related receptor alpha |
| FRAP | ferric reducing antioxidant power |
| FSS | flow shear stress |
| FW | fresh weight |
| G6PD | glucose-6-phosphate dehydrogenase |
| GAS | gamma-activated sequence |
| γ-GCS | gamma-glutamylcysteine synthetase |
| G-CSF | granulocyte colony-stimulating factor |
| GFAP | glia and fibrillary acidic protein |
| GM-CSF | granulocyte–macrophage colony-stimulating factor |
| GPx | glutathione peroxidase |
| GSH-Rd | glutathione reductase |
| GSK3β | glycogen synthase kinase 3β |
| H2-Ab1 | histocompatibility class II antigen A, beta 1 |
| HDAC | histone deacetylase |
| Hes1 | hairy and enhancer of split 1 |
| HIOMT | hydroxyindole O-methyltransferase |
| HOMA-IR index | homeostasis model assessment of insulin resistance index |
| hTERT | human telomerase reserve transcriptase |
| i.v. | intravenously |
| i.p. | intraperitoneally |
| IFN-γ | interferon gamma |
| IGF-1 | insulin-like growth factor 1 |
| IKK | inhibitor of nuclear factor kappa-B kinase |
| IL-6 | interleukin 6 |
| iNOS | inducible nitric oxide synthase |
| IRI | ischemia/reperfusion injury |
| JAK2 | Janus kinase 2 |
| JNK | Jun N-terminal kinase |
| LAN | light at night |
| LPS | lipopolysaccharide |
| MAPK | mitogen-activated protein kinase |
| MDH | malondialdehyde |
| MI | myocardial infarction |
| MLHb | medial lateral habenula |
| MMP-9 | matrix metalloproteinase-9 |
| Mst1 | mammalian Ste20-like kinase 1 |
| MT1, MT2 | melatonin receptors |
| NAD | Nicotinamide adenine dinucleotide |
| NAT | N-acetyltransferase |
| NDRG2 | N-myc downstream-regulated gene 2 |
| NF-κB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| NK | natural killer |
| NLRP3 | NOD-like receptor P3 |
| NO | nitric oxide |
| NOMela | N-nitrosomelatonin |
| Nrf2 | nuclear factor-erythroid 2-related factor 2 |
| OA | osteoarthritis |
| ON | optic neuritis |
| ORAC | oxygen radical antioxidant capacity |
| Oxa | oxaliplatin |
| p-Akt | phosphorylated protein kinase B |
| PARP | poly-ADP-ribose polymerase |
| pCREB | phosphorylated cAMP response element-binding protein |
| PD | Parkinson’s disease |
| PDGF | platelet-derived growth factor |
| PGC-1α | peroxisome proliferator-activated receptor gamma coactivator-1 alpha |
| PI3K | phosphatidyl inositol 3-kinase |
| PKB | protein kinase B |
| PLR | pupil light reflex |
| PPAR-γ | peroxisome proliferator-activated receptor gamma |
| PRDX1 | peroxiredoxin 1 |
| PS1 | presenilin 1 |
| PUMA | p53 upregulated modulator of apoptosis |
| RNS | reactive nitrogen species |
| ROCK-1 | Rho-associated kinase protein |
| ROS | reactive oxygen species |
| S100B | S100 calcium-binding protein B |
| SAH | subarachnoid hemorrhage |
| SCN | suprachiasmatic nucleus |
| SE | standard error |
| SEEMs | selective estrogen enzyme modulators |
| SIRT1 | sirtuin 1 |
| SOD | superoxide dismutase |
| STAT | signal transducer and activator of transcription |
| TBI | traumatic brain injury |
| TGF-β | transforming growth factor β |
| TIMP-1 | tissue inhibitor of metalloproteinases 1 |
| TNF-α | tumor necrosis factor α |
| TPH | tryptophan hydroxylase |
| UCP1 | uncoupling protein 1 |
| UPS | ubiquitin/proteasome system |
| VEGF | vascular endothelial growth factor |
| VEP | visual evoked potentials |
References
- Lerner, A.B.; Case, J.D.; Takahashi, Y.; Lee, T.H.; Mori, W. Isolation of melatonin, a pineal factor that lightens melanocytes. J. Am. Chem. Soc. 1958, 80, 2587. [Google Scholar] [CrossRef]
- Brown, G.M.; Pandi-Perumal, S.R.; Trakht, I.; Cardinali, D.P. Melatonin and its relevance to jet lag. Travel Med. Infect. Dis. 2009, 7, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Pandi-Perumal, S.R.; Srinivasan, V.; Poeggeler, B.; Hardeland, R.; Cardinali, D.P. Drug Insight: The use of melatonergic agonists for the treatment of insomnia-focus on ramelteon. Nat. Clin. Pract. Neurol. 2007, 3, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Luo, X.; Li, L.; Peng, Q.; Yang, Y.; Zhao, L.; Ma, M.; Hou, Z. The protective effects of melatonin against oxidative stress and inflammation induced by acute cadmium exposure in mice testis. Biol. Trace Elem. Res. 2016, 170, 152–164. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.J.; Huang, S.H.; Chen, J.W.; Wang, K.C.; Yang, Y.R.; Liu, P.F.; Lin, G.J.; Sytwu, H.K. Melatonin enhances interleukin-10 expression and suppresses chemotaxis to inhibit inflammation in situ and reduce the severity of experimental autoimmune encephalomyelitis. Int. Immunopharmacol. 2016, 31, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Oxenkrug, G.; Requintina, P.; Bachurin, S. Antioxidant and antiaging activity of N-acetylserotonin and melatonin in the in vivo models. Ann. N. Y. Acad. Sci. 2001, 939, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Li, S.; Li, H.B.; Deng, G.F.; Ling, W.H.; Wu, S.; Xu, X.R.; Chen, F. Antiproliferative activity of peels, pulps and seeds of 61 fruits. J. Funct. Foods 2013, 5, 1298–1309. [Google Scholar] [CrossRef]
- Anisimov, V.N.; Popovich, I.G.; Zabezhinski, M.A.; Anisimov, S.V.; Vesnushkin, G.M.; Vinogradova, I.A. Melatonin as antioxidant, geroprotector and anticarcinogen. Biochim. Biophys. Acta 2006, 1757, 573–589. [Google Scholar] [CrossRef] [PubMed]
- Pandi-Perumal, S.R.; BaHammam, A.S.; Brown, G.M.; Spence, D.W.; Bharti, V.K.; Kaur, C.; Hardeland, R.; Cardinali, D.P. Melatonin antioxidative defense: Therapeutical implications for aging and neurodegenerative processes. Neurotox. Res. 2013, 23, 267–300. [Google Scholar] [CrossRef] [PubMed]
- Pandi-Perumal, S.R.; Zisapel, N.; Srinivasan, V.; Cardinali, D.P. Melatonin and sleep in aging population. Exp. Gerontol. 2005, 40, 911–925. [Google Scholar] [CrossRef] [PubMed]
- Agil, A.; Elmahallawy, E.K.; Rodriguez-Ferrer, J.M.; Adem, A.; Bastaki, S.M.; Al-Abbadi, I.; Fino Solano, Y.A.; Navarro-Alarcon, M. Melatonin increases intracellular calcium in the liver, muscle, white adipose tissues and pancreas of diabetic obese rats. Food Funct. 2015, 6, 2671–2678. [Google Scholar] [CrossRef] [PubMed]
- Agil, A.; El-Hammadi, M.; Jimenez-Aranda, A.; Tassi, M.; Abdo, W.; Fernandez-Vazquez, G.; Reiter, R.J. Melatonin reduces hepatic mitochondrial dysfunction in diabetic obese rats. J. Pineal Res. 2015, 59, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Chenevard, R.; Suter, Y.; Erne, P. Effects of the heart-lung machine on melatonin metabolism and mood disturbances. Eur. J. Cardiothorac. Surg. 2008, 34, 338–343. [Google Scholar] [CrossRef] [PubMed]
- Esquifino, A.I.; Villanua, M.A.; Agrasal, C. Effect of neonatal melatonin administration on sexual development in the rat. J. Steroid Biochem. 1987, 27, 1089–1093. [Google Scholar] [CrossRef]
- Agilli, M.; Aydin, F.N.; Cayci, T. The effect of body temperature, melatonin and cortisol on obesity in women: A biochemical evaluation? Clin. Nutr. 2015, 34, 332. [Google Scholar] [CrossRef] [PubMed]
- Najeeb, S.; Khurshid, Z.; Zohaib, S.; Zafar, M.S. Therapeutic potential of melatonin in oral medicine and periodontology. Kaohsiung J. Med. Sci. 2016, 32, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Setyaningsih, W.; Saputro, I.E.; Barbero, G.F.; Palma, M.; Garcia Barroso, C. Determination of melatonin in rice (Oryza sativa) grains by pressurized liquid extraction. J. Agric. Food Chem. 2015, 63, 1107–1115. [Google Scholar] [CrossRef] [PubMed]
- Escriva, L.; Manyes, L.; Barbera, M.; Martinez-Torres, D.; Meca, G. Determination of melatonin in Acyrthosiphon pisum aphids by liquid chromatography-tandem mass spectrometry. J. Insect Physiol. 2016, 86, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Muszynska, B.; Sulkowska-Ziaja, K. Analysis of indole compounds in edible Basidiomycota species after thermal processing. Food Chem. 2012, 132, 455–459. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, C.; Kocadagli, T.; Gokmen, V. Formation of melatonin and its isomer during bread dough fermentation and effect of baking. J. Agric. Food Chem. 2014, 62, 2900–2905. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X. Melatonin: An antioxidant in edible plants. Ann. N. Y. Acad. Sci. 2002, 957, 341–344. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Zanghi, B.M.; Manchester, L.C.; Reiter, R.J. Melatonin identified in meats and other food stuffs: Potentially nutritional impact. J. Pineal Res. 2014, 57, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, Y.; Herrera, T.; Benitez, V.; Arribas, S.M.; Lopez De Pablo, A.L.; Esteban, R.M.; Martin-Cabrejas, M.A. Estimation of scavenging capacity of melatonin and other antioxidants: Contribution and evaluation in germinated seeds. Food Chem. 2015, 170, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Sae-Teaw, M.; Johns, J.; Johns, N.P.; Subongkot, S. Serum melatonin levels and antioxidant capacities after consumption of pineapple, orange, or banana by healthy male volunteers. J. Pineal Res. 2013, 55, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Delgado, J.; Terron, M.P.; Garrido, M.; Pariente, J.A.; Barriga, C.; Rodriguez, A.B.; Paredes, S.D. Diets enriched with a Jerte Valley cherry-based nutraceutical product reinforce nocturnal behaviour in young and old animals of nocturnal (Rattus norvegicus) and diurnal (Streptopelia risoria) chronotypes. J. Anim. Physiol. Anim. Nutr. 2013, 97, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Iriti, M.; Faoro, F. Grape phytochemicals: A bouquet of old and new nutraceuticals for human health. Med. Hypotheses 2006, 67, 833–838. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, C.K. Functional foods, herbs and nutraceuticals: Towards biochemical mechanisms of healthy aging. Biogerontology 2004, 5, 275–289. [Google Scholar] [CrossRef] [PubMed]
- Oladi, E.; Mohamadi, M.; Shamspur, T.; Mostafavi, A. Spectrofluorimetric determination of melatonin in kernels of four different Pistacia varieties after ultrasound-assisted solid-liquid extraction. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2014, 132, 326–329. [Google Scholar] [CrossRef] [PubMed]
- Murch, S.J.; Simmons, C.B.; Saxena, P.K. Melatonin in feverfew and other medicinal plants. Lancet 1997, 350, 1598–1599. [Google Scholar] [CrossRef]
- González-Gómez, D.; Lozano, M.; Fernández-León, M.F.; Ayuso, M.C.; Bernalte, M.J.; Rodríguez, A.B. Detection and quantification of melatonin and serotonin in eight Sweet Cherry cultivars (Prunus avium L.). Eur. Food Res. Technol. 2009, 229, 223–229. [Google Scholar] [CrossRef]
- Wang, J.; Liang, C.; Li, S.; Zheng, J. Study on analysis method of melatonin and melatonin content in corn & rice seeds. Chin. Agric. Sci. Bull. 2009, 25, 20–24. [Google Scholar]
- Hernandez-Ruiz, J.; Arnao, M.B. Distribution of melatonin in different zones of lupin and barley plants at different ages in the presence and absence of light. J. Agric. Food Chem. 2008, 56, 10567–10573. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Burkhardt, S.; Manchester, L.C. Melatonin in plants. Nutr. Rev. 2001, 59, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R.; Pandi-Perumal, S.R.; Cardinali, D.P. Melatonin. Int. J. Biochem. Cell Biol. 2006, 38, 313–316. [Google Scholar] [CrossRef] [PubMed]
- Iriti, M.; Rossoni, M.; Faoro, F. Melatonin content in grape: Myth or panacea. J. Sci. Food Agric. 2006, 86, 1432–1438. [Google Scholar] [CrossRef]
- Wang, C.; Yin, L.Y.; Shi, X.Y.; Xiao, H.; Kang, K.; Liu, X.Y.; Zhan, J.C.; Huang, W.D. Effect of cultivar, temperature, and environmental conditions on the dynamic change of melatonin in mulberry fruit development and wine fermentation. J. Food Sci. 2016, 81, M958–M967. [Google Scholar] [CrossRef] [PubMed]
- Karunanithi, D.; Radhakrishna, A.; Sivaraman, K.P.; Biju, V.M. Quantitative determination of melatonin in milk by LC-MS/MS. J. Food Sci. Technol. 2014, 51, 805–812. [Google Scholar] [CrossRef] [PubMed]
- Milagres, M.P.; Minim, V.P.; Minim, L.A.; Simiqueli, A.A.; Moraes, L.E.; Martino, H.S. Night milking adds value to cow’s milk. J. Sci. Food Agric. 2014, 94, 1688–1692. [Google Scholar] [CrossRef] [PubMed]
- Berthelot, X.; Laurentie, M.; Ravault, J.P.; Ferney, J.; Toutain, P.L. Circadian profile and production rate of melatonin in the cow. Domest. Anim. Endocrinol. 1990, 7, 315–322. [Google Scholar] [CrossRef]
- Kennaway, D.J.; Stamp, G.E.; Goble, F.C. Development of melatonin production in infants and the impact of prematurity. J. Clin. Endocrinol. Metab. 1992, 75, 367–369. [Google Scholar] [CrossRef] [PubMed]
- Cohen Engler, A.; Hadash, A.; Shehadeh, N.; Pillar, G. Breastfeeding may improve nocturnal sleep and reduce infantile colic: Potential role of breast milk melatonin. Eur. J. Pediatr. 2012, 171, 729–732. [Google Scholar] [CrossRef] [PubMed]
- Kocadagli, T.; Yilmaz, C.; Gokmen, V. Determination of melatonin and its isomer in foods by liquid chromatography tandem mass spectrometry. Food Chem. 2014, 153, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Mercolini, L.; Mandrioli, R.; Raggi, M.A. Content of melatonin and other antioxidants in grape-related foodstuffs: Measurement using a MEPS-HPLC-F method. J. Pineal Res. 2012, 53, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Sturtz, M.; Cerezo, A.B.; Cantos-Villar, E.; Garcia-Parrilla, M.C. Determination of the melatonin content of different varieties of tomatoes (Lycopersicon esculentum) and strawberries (Fragariaananassa). Food Chem. 2011, 127, 1329–1334. [Google Scholar] [CrossRef] [PubMed]
- Boccalandro, H.E.; Gonzalez, C.V.; Wunderlin, D.A.; Silva, M.F. Melatonin levels, determined by LC-ESI-MS/MS, fluctuate during the day/night cycle in Vitis vinifera cv. Malbec: Evidence of its antioxidant role in fruits. J. Pineal Res. 2011, 51, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Burkhardt, S.; Tan, D.X.; Manchester, L.C.; Hardeland, R.; Reiter, R.J. Detection and quantification of the antioxidant melatonin in Montmorency and Balaton tart cherries (Prunus cerasus). J. Agric. Food Chem. 2001, 49, 4898–4902. [Google Scholar] [CrossRef] [PubMed]
- Badria, F.A. Melatonin, serotonin, and tryptamine in some Egyptian food and medicinal plants. J. Med. Food 2002, 5, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Dubbels, R.; Reiter, R.J.; Klenke, E.; Goebel, A.; Schnakenberg, E.; Ehlers, C.; Schiwara, H.W.; Schloot, W. Melatonin in edible plants identified by radioimmunoassay and by high performance liquid chromatography-mass spectrometry. J. Pineal Res. 1995, 18, 28–31. [Google Scholar] [CrossRef] [PubMed]
- Riga, P.; Medina, S.; Garcia-Flores, L.A.; Gil-Izquierdo, A. Melatonin content of pepper and tomato fruits: Effects of cultivar and solar radiation. Food Chem. 2014, 156, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Muszynska, B.; Kala, K.; Sulkowska-Ziaja, K.; Krakowska, A.; Opoka, W. Agaricus bisporus and its in vitro culture as a source of indole compounds released into artificial digestive juices. Food Chem. 2016, 199, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Manchester, L.C.; Tan, D.X.; Reiter, R.J.; Park, W.; Monis, K.; Qi, W. High levels of melatonin in the seeds of edible plants: Possible function in germ tissue protection. Life Sci. 2000, 67, 3023–3029. [Google Scholar] [CrossRef]
- Zielin Ski, H.; Lewczuk, B.; Przybylska-Gornowicz, B.; Kozlowska, H. Melatonin in germinated legume seeds as a potentially significant agent for health. In Biologically-Active phytochemicals in Food: Analysis, Metabolism, Bioavailability and Function, Norwich, UK, 2001; Pfannhauser, W., Fenwick, G.R., Khokhar, S.R., Eds.; Royal Society of Chemistry: Cambridge, UK, 2001. [Google Scholar]
- Aguilera, Y.; Herrera, T.; Liebana, R.; Rebollo-Hernanz, M.; Sanchez-Puelles, C.; Martin-Cabrejas, M.A. Impact of melatonin enrichment during germination of legumes on bioactive compounds and antioxidant activity. J. Agric. Food Chem. 2015, 63, 7967–7974. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Moreno, H.; Calvo, J.R.; Maldonado, M.D. High levels of melatonin generated during the brewing process. J. Pineal Res. 2013, 55, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Naranjo, M.I.; Gil-Izquierdo, A.; Troncoso, A.M.; Cantos, E.; Garcia-Parrilla, M.C. Melatonin: A new bioactive compound in wine. J. Food Compos. Anal. 2011, 24, 603–608. [Google Scholar] [CrossRef]
- Stege, P.W.; Sombra, L.L.; Messina, G.; Martinez, L.D.; Silva, M.F. Determination of melatonin in wine and plant extracts by capillary electrochromatography with immobilized carboxylic multi-walled carbon nanotubes as stationary phase. Electrophoresis 2010, 31, 2242–2248. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Naranjo, M.I.; Gil-Izquierdo, A.; Troncoso, A.M.; Cantos-Villar, E.; Garcia-Parrilla, M.C. Melatonin is synthesised by yeast during alcoholic fermentation in wines. Food Chem. 2011, 126, 1608–1613. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishna, A.; Giridhar, P.; Sankar, K.U.; Ravishankar, G.A. Melatonin and serotonin profiles in beans of Coffea species. J. Pineal Res. 2012, 52, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Vitalini, S.; Gardana, C.; Simonetti, P.; Fico, G.; Iriti, M. Melatonin, melatonin isomers and stilbenes in Italian traditional grape products and their antiradical capacity. J. Pineal Res. 2013, 54, 322–333. [Google Scholar] [CrossRef] [PubMed]
- Kirakosyan, A.; Seymour, E.M.; Llanes, D.E.U.; Kaufman, P.B.; Bolling, S.F. Chemical profile and antioxidant capacities of tart cherry products. Food Chem. 2009, 115, 20–25. [Google Scholar] [CrossRef]
- Chen, G.; Huo, Y.; Tan, D.X.; Liang, Z.; Zhang, W.; Zhang, Y. Melatonin in Chinese medicinal herbs. Life Sci. 2003, 73, 19–26. [Google Scholar] [CrossRef]
- Padumanonda, T.; Johns, J.; Sangkasat, A.; Tiyaworanant, S. Determination of melatonin content in traditional Thai herbal remedies used as sleeping aids. Daru 2014, 22, 6. [Google Scholar] [CrossRef] [PubMed]
- de la Puerta, C.; Carrascosa-Salmoral, M.P.; García-Luna, P.P.; Lardone, P.J.; Herrera, J.L.; Fernández-Montesinos, R.; Guerrero, J.M.; Pozo, D. Melatonin is a phytochemical in olive oil. Food Chem. 2007, 104, 609–612. [Google Scholar] [CrossRef]
- Venegas, C.; Cabrera-Vique, C.; Garcia-Corzo, L.; Escames, G.; Acuna-Castroviejo, D.; Lopez, L.C. Determination of coenzyme Q10, coenzyme Q9, and melatonin contents in virgin argan oils: Comparison with other edible vegetable oils. J. Agric. Food Chem. 2011, 59, 12102–12108. [Google Scholar] [CrossRef] [PubMed]
- Reinholds, I.; Pugajeva, I.; Radenkovs, V.; Rjabova, J.; Bartkevics, V. Development and validation of new ultra-high-performance liquid chromatography-hybrid quadrupole-orbitrap mass spectrometry method for determination of melatonin in fruits. J. Chromatogr. Sci. 2016, 54, 977–984. [Google Scholar] [CrossRef] [PubMed]
- Vakkuri, O.; Kivela, A.; Leppaluoto, J.; Valtonen, M.; Kauppila, A. Decrease in melatonin precedes follicle-stimulating hormone increase during perimenopause. Eur. J. Endocrinol. 1996, 135, 188–192. [Google Scholar] [CrossRef] [PubMed]
- Waldhauser, F.; Weiszenbacher, G.; Tatzer, E.; Gisinger, B.; Waldhauser, M.; Schemper, M.; Frisch, H. Alterations in nocturnal serum melatonin levels in humans with growth and aging. J. Clin. Endocrinol. Metab. 1988, 66, 648–652. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Palacios-Bois, J.; Schwartz, G.; Iskandar, H.; Thakur, M.; Quirion, R.; Nair, N.P. Circadian rhythms of melatonin and cortisol in aging. Biol. Psychiatry 1989, 25, 305–319. [Google Scholar] [CrossRef]
- Singh, M.; Jadhav, H.R. Melatonin: Functions and ligands. Drug Discov. Today 2014, 19, 1410–1418. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Xiao, X.; Zhang, C.; Yu, W.; Guo, W.; Zhang, Z.; Li, Z.; Feng, X.; Hao, J.; Zhang, K.; et al. Melatonin synergizes the chemotherapeutic effect of 5-fluorouracil in colon cancer by suppressing PI3K/AKT and NF-kappaB/iNOS signaling pathways. J. Pineal Res. 2017, 62. [Google Scholar] [CrossRef] [PubMed]
- DeMuro, R.L.; Nafziger, A.N.; Blask, D.E.; Menhinick, A.M.; Bertino, J.J. The absolute bioavailability of oral melatonin. J. Clin. Pharmacol. 2000, 40, 781–784. [Google Scholar] [CrossRef] [PubMed]
- Fourtillan, J.B.; Brisson, A.M.; Gobin, P.; Ingrand, I.; Decourt, J.P.; Girault, J. Bioavailability of melatonin in humans after day-time administration of D7 melatonin. Biopharm. Drug Dispos. 2000, 21, 15–22. [Google Scholar] [CrossRef]
- Shirakawa, S.; Tsuchiya, S.; Tsutsumi, Y.; Kotorii, T.; Uchimura, N.; Sakamoto, T.; Yamada, S. Time course of saliva and serum melatonin levels after ingestion of melatonin. Psychiatry Clin. Neurosci. 1998, 52, 266–267. [Google Scholar] [CrossRef] [PubMed]
- Oba, S.; Nakamura, K.; Sahashi, Y.; Hattori, A.; Nagata, C. Consumption of vegetables alters morning urinary 6-sulfatoxymelatonin concentration. J. Pineal Res. 2008, 45, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, M.D.; Moreno, H.; Calvo, J.R. Melatonin present in beer contributes to increase the levels of melatonin and antioxidant capacity of the human serum. Clin. Nutr. 2009, 28, 188–191. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Flores, D.; Gamero, E.; Garrido, M.; Ramirez, R.; Moreno, D.; Delgado, J.; Valdes, E.; Barriga, C.; Rodriguez, A.B.; Paredes, S.D. Urinary 6-sulfatoxymelatonin and total antioxidant capacity increase after the intake of a grape juice cv. Tempranillo stabilized with HHP. Food Funct. 2012, 3, 34–39. [Google Scholar] [CrossRef] [PubMed]
- González-Flores, D.; Velardo, B.; Garrido, M.; González-Gómez, D.; Lozano, M.; Ayuso, M.C.; Barriga, C.; Paredes, S.D.; Rodríguez, A.B. Ingestion of Japanese plums (Prunus salicina Lindl. cv. Crimson Globe) increases the urinary 6-sulfatoxymelatonin and total antioxidant capacity levels in young, middle-aged and elderly humans: nutritional and functional characterization of their content. J. Food Nutr. Res. 2011, 50, 229. [Google Scholar]
- Reiter, R.J.; Manchester, L.C.; Tan, D.X. Melatonin in walnuts: Influence on levels of melatonin and total antioxidant capacity of blood. Nutrition 2005, 21, 920–924. [Google Scholar] [CrossRef] [PubMed]
- Schernhammer, E.S.; Feskanich, D.; Niu, C.; Dopfel, R.; Holmes, M.D.; Hankinson, S.E. Dietary correlates of urinary 6-sulfatoxymelatonin concentrations in the Nurses’ Health Study cohorts. Am. J. Clin. Nutr. 2009, 90, 975–985. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, Y.; Rebollo-Hernanz, M.; Herrera, T.; Cayuelas, L.T.; Rodriguez-Rodriguez, P.; de Pablo, A.L.; Arribas, S.M.; Martin-Cabrejas, M.A. Intake of bean sprouts influences melatonin and antioxidant capacity biomarker levels in rats. Food Funct. 2016, 7, 1438–1445. [Google Scholar] [CrossRef] [PubMed]
- Tetens, I. Scientific Opinion on the Substantiation of Health Claims Related to Melatonin and Alleviation of Subjective Feelings of Jet Lag (ID 1953), and Reduction of Sleep onset Latency, and Improvement of Sleep Quality (ID 1953) Pursuant to Article 13(1) of Regulation (EC) No 1924/2006; European Food Safety Authority: Parma, Itlay, 2010. [Google Scholar]
- Food Standards Australia New Zealand (FSANZ). Supporting document 9: Consideration of EU Approved Health Claims, P293—Nutrition, Health & Related Claim; Food Standards Australia New Zealand: Kingston, Australia, Wellington, Newzealand, 2013. [Google Scholar]
- Hattori, A.; Migitaka, H.; Iigo, M.; Itoh, M.; Yamamoto, K.; Ohtani-Kaneko, R.; Hara, M.; Suzuki, T.; Reiter, R.J. Identification of melatonin in plants and its effects on plasma melatonin levels and binding to melatonin receptors in vertebrates. Biochem. Mol. Biol. Int. 1995, 35, 627–634. [Google Scholar] [PubMed]
- Byeon, Y.; Back, K. An increase in melatonin in transgenic rice causes pleiotropic phenotypes, including enhanced seedling growth, delayed flowering, and low grain yield. J. Pineal Res. 2014, 56, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Ruiz, J.; Cano, A.; Arnao, M.B. Melatonin acts as a growth-stimulating compound in some monocot species. J. Pineal Res. 2005, 39, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Fardet, A. New hypotheses for the health-protective mechanisms of whole-grain cereals: What is beyond fibre? Nutr. Res. Rev. 2010, 23, 65–134. [Google Scholar] [CrossRef] [PubMed]
- Hosseinian, F.S.; Li, W.; Beta, T. Measurement of anthocyanins and other phytochemicals in purple wheat. Food Chem. 2008, 109, 916–924. [Google Scholar] [CrossRef] [PubMed]
- Lei, Q.; Wang, L.; Tan, D.X.; Zhao, Y.; Zheng, X.D.; Chen, H.; Li, Q.T.; Zuo, B.X.; Kong, J. Identification of genes for melatonin synthetic enzymes in ‘Red Fuji’ apple (Malus domestica Borkh. cv. Red) and their expression and melatonin production during fruit development. J. Pineal Res. 2013, 55, 443–451. [Google Scholar] [PubMed]
- Zhao, Y.; Tan, D.X.; Lei, Q.; Chen, H.; Wang, L.; Li, Q.T.; Gao, Y.; Kong, J. Melatonin and its potential biological functions in the fruits of sweet cherry. J. Pineal Res. 2013, 55, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Vitalini, S.; Gardana, C.; Zanzotto, A.; Simonetti, P.; Faoro, F.; Fico, G.; Iriti, M. The presence of melatonin in grapevine (Vitis vinifera L.) berry tissues. J. Pineal Res. 2011, 51, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Brown, P.N.; Turi, C.E.; Shipley, P.R.; Murch, S.J. Comparisons of large (Vaccinium macrocarpon Ait.) and small (Vaccinium oxycoccos L., Vaccinium vitis-idaea L.) cranberry in British Columbia by phytochemical determination, antioxidant potential, and metabolomic profiling with chemometric analysis. Planta Med. 2012, 78, 630–640. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P.; Tan, D.X.; Manchester, L.C.; Reiter, R.J. Purslane: A plant source of omega-3 fatty acids and melatonin. J. Pineal Res. 2005, 39, 331–332. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, M.; Ezura, H. Profiling of melatonin in the model tomato (Solanum lycopersicum L.) cultivar Micro-Tom. J. Pineal Res. 2009, 46, 338–343. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhao, Y.; Reiter, R.J.; He, C.; Liu, G.; Lei, Q.; Zuo, B.; Zheng, X.D.; Li, Q.; Kong, J. Changes in melatonin levels in transgenic ‘Micro-Tom’ tomato overexpressing ovine AANAT and ovine HIOMT genes. J. Pineal Res. 2014, 56, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.J.; Zhang, N.; Yang, R.C.; Wang, L.; Sun, Q.Q.; Li, D.B.; Cao, Y.Y.; Weeda, S.; Zhao, B.; Ren, S.; Guo, Y.D. Melatonin promotes seed germination under high salinity by regulating antioxidant systems, ABA and GA(4) interaction in cucumber (Cucumis sativus L.). J. Pineal Res. 2014, 57, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Byeon, Y.; Park, S.; Lee, H.Y.; Kim, Y.S.; Back, K. Elevated production of melatonin in transgenic rice seeds expressing rice tryptophan decarboxylase. J. Pineal Res. 2014, 56, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Tapia, M.I.; Sánchez-Morgado, J.R.; García-Parra, J.; Ramírez, R.; Hernández, T.; González-Gómez, D. Comparative study of the nutritional and bioactive compounds content of four walnut (Juglans regia L.) cultivars. J. Food Compos. Anal. 2013, 31, 232–237. [Google Scholar] [CrossRef]
- Fernandez-Pachon, M.S.; Medina, S.; Herrero-Martin, G.; Cerrillo, I.; Berna, G.; Escudero-Lopez, B.; Ferreres, F.; Martin, F.; Garcia-Parrilla, M.C.; Gil-Izquierdo, A. Alcoholic fermentation induces melatonin synthesis in orange juice. J. Pineal Res. 2014, 56, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Pothinuch, P.; Tongchitpakdee, S. Melatonin contents in mulberry (Morus spp.) leaves: Effects of sample preparation, cultivar, leaf age and tea processing. Food Chem. 2011, 128, 415–419. [Google Scholar] [CrossRef] [PubMed]
- Wehr, T.A.; Duncan, W.C., Jr.; Sher, L.; Aeschbach, D.; Schwartz, P.J.; Turner, E.H.; Postolache, T.T.; Rosenthal, N.E. A circadian signal of change of season in patients with seasonal affective disorder. Arch. Gen. Psychiatry 2001, 58, 1108–1114. [Google Scholar] [CrossRef] [PubMed]
- Rexhaj, E.; Pireva, A.; Paoloni-Giacobino, A.; Allemann, Y.; Cerny, D.; Dessen, P.; Sartori, C.; Scherrer, U.; Rimoldi, S.F. Prevention of vascular dysfunction and arterial hypertension in mice generated by assisted reproductive technologies by addition of melatonin to culture media. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H1151–H1156. [Google Scholar] [CrossRef] [PubMed]
- Michurina, S.V.; Ishchenko, I.Y.; Arkhipov, S.A.; Klimontov, V.V.; Rachkovskaya, L.N.; Konenkov, V.I.; Zavyalov, E.L. Effects of melatonin, aluminum oxide, and polymethylsiloxane complex on the expression of LYVE-1 in the liver of mice with obesity and type 2 diabetes mellitus. Bull. Exp. Biol. Med. 2016, 162, 269–272. [Google Scholar] [CrossRef] [PubMed]
- Bielli, A.; Scioli, M.G.; Mazzaglia, D.; Doldo, E.; Orlandi, A. Antioxidants and vascular health. Life Sci. 2015, 143, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Barton, S.K.; Tolcos, M.; Miller, S.L.; Christoph-Roehr, C.; Schmolzer, G.M.; Moss, T.J.; Hooper, S.B.; Wallace, E.M.; Polglase, G.R. Ventilation-induced brain injury in preterm neonates: A review of potential therapies. Neonatology 2016, 110, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Droge, W. Free radicals in the physiological control of cell function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Haldar, C. Age dependent nitro-oxidative load and melatonin receptor expression in the spleen and immunity of goat Capra hircus. Exp. Gerontol. 2014, 60, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Aktoz, T.; Aydogdu, N.; Alagol, B.; Yalcin, O.; Huseyinova, G.; Atakan, I.H. The protective effects of melatonin and vitamin E against renal ischemia-reperfusion injury in rats. Ren. Fail. 2007, 29, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Lissoni, P.; Chilelli, M.; Villa, S.; Cerizza, L.; Tancini, G. Five years survival in metastatic non-small cell lung cancer patients treated with chemotherapy alone or chemotherapy and melatonin: A randomized trial. J. Pineal Res. 2003, 35, 12–15. [Google Scholar] [CrossRef] [PubMed]
- Paredes, S.D.; Forman, K.A.; Garcia, C.; Vara, E.; Escames, G.; Tresguerres, J.A. Protective actions of melatonin and growth hormone on the aged cardiovascular system. Horm. Mol. Biol. Clin. Investig. 2014, 18, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Zephy, D.; Ahmad, J. Type 2 diabetes mellitus: Role of melatonin and oxidative stress. Diabetes Metab. Syndr. 2015, 9, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.H.; Lin, K.C.; Wallace, C.G.; Chen, Y.T.; Yang, C.C.; Leu, S.; Chen, Y.C.; Sun, C.K.; Tsai, T.H.; Chen, Y.L.; et al. Additional benefit of combined therapy with melatonin and apoptotic adipose-derived mesenchymal stem cell against sepsis-induced kidney injury. J. Pineal Res. 2014, 57, 16–32. [Google Scholar] [CrossRef] [PubMed]
- Deng, G.F.; Lin, X.; Xu, X.R.; Gao, L.L.; Xie, J.F.; Li, H.B. Antioxidant capacities and total phenolic contents of 56 vegetables. J. Funct. Foods 2013, 5, 260–266. [Google Scholar] [CrossRef]
- Fu, L.; Xu, B.T.; Xu, X.R.; Qin, X.S.; Gan, R.Y.; Li, H.B. Antioxidant capacities and total phenolic contents of 56 wild fruits from South China. Molecules 2010, 15, 8602–8617. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.J.; Deng, G.F.; Xu, X.R.; Wu, S.; Li, S.; Xia, E.Q.; Li, F.; Chen, F.; Ling, W.H.; Li, H.B. Antioxidant capacities, phenolic compounds and polysaccharide contents of 49 edible macro-fungi. Food Funct. 2012, 3, 1195–1205. [Google Scholar] [CrossRef] [PubMed]
- Li, A.N.; Li, S.; Li, H.B.; Xu, D.P.; Xu, X.R.; Chen, F. Total phenolic contents and antioxidant capacities of 51 edible and wild flowers. J. Funct. Foods 2014, 6, 319–330. [Google Scholar] [CrossRef]
- Li, S.; Li, S.K.; Gan, R.Y.; Song, F.L.; Kuang, L.; Li, H.B. Antioxidant capacities and total phenolic contents of infusions from 223 medicinal plants. Ind. Crops Prod. 2013, 51, 289–298. [Google Scholar] [CrossRef]
- Deng, G.F.; Xu, X.R.; Zhang, Y.; Li, D.; Gan, R.Y.; Li, H.B. Phenolic compounds and bioactivities of pigmented rice. Crit. Rev. Food Sci. 2013, 53, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Li, A.N.; Li, S.; Zhang, Y.J.; Xu, X.R.; Chen, Y.M.; Li, H.B. Resources and biological activities of natural polyphenols. Nutrients 2014, 6, 6020–6047. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, J.J.; Xu, D.P.; Zhou, T.; Zhou, Y.; Li, S.; Li, H.B. Bioactivities and health benefits of wild fruits. Int. J. Mol. Sci. 2016, 17, 1258. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.J.; Li, Y.; Zhou, T.; Xu, D.P.; Zhang, P.; Li, S.; Li, H.B. Bioactivities and health benefits of mushrooms mainly from China. Molecules 2016, 21, 938. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.J.; Gan, R.Y.; Li, S.; Zhou, Y.; Li, A.N.; Xu, D.P.; Li, H.B. Antioxidant phytochemicals for the prevention and treatment of chronic diseases. Molecules 2015, 20, 21138–21156. [Google Scholar] [CrossRef] [PubMed]
- Okatani, Y.; Wakatsuki, A.; Reiter, R.J.; Miyahara, Y. Acutely administered melatonin restores hepatic mitochondrial physiology in old mice. Int. J. Biochem. Cell Biol. 2003, 35, 367–375. [Google Scholar] [CrossRef]
- Khaldy, H.; Escames, G.; Leon, J.; Vives, F.; Luna, J.D.; Acuna-Castroviejo, D. Comparative effects of melatonin, L-deprenyl, Trolox and ascorbate in the suppression of hydroxyl radical formation during dopamine autoxidation in vitro. J. Pineal Res. 2000, 29, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Urata, Y.; Honma, S.; Goto, S.; Todoroki, S.; Iida, T.; Cho, S.; Honma, K.; Kondo, T. Melatonin induces gamma-glutamylcysteine synthetase mediated by activator protein-1 in human vascular endothelial cells. Free Radic. Biol. Med. 1999, 27, 838–847. [Google Scholar] [CrossRef]
- Montilla, P.; Cruz, A.; Padillo, F.J.; Tunez, I.; Gascon, F.; Munoz, M.C.; Gomez, M.; Pera, C. Melatonin versus vitamin E as protective treatment against oxidative stress after extra-hepatic bile duct ligation in rats. J. Pineal Res. 2001, 31, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Poeggeler, B.; Reiter, R.J.; Tan, D.X.; Chen, L.D.; Manchester, L.C. Melatonin: A potent, endogenous hydroxyl radical scavenger. J. Pineal Res. 1993, 14, 57–60. [Google Scholar]
- Okatani, Y.; Watanabe, K.; Hayashi, K.; Wakatsuki, A.; Sagara, Y. Melatonin inhibits vasospastic action of hydrogen peroxide in human umbilical artery. J. Pineal Res. 1997, 22, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.; Han, B.; Liu, M.; Yeh, C.; Casida, J.E. Phosphine-induced oxidative damage in rats: Attenuation by melatonin. Free Radic. Biol. Med. 2000, 28, 636–642. [Google Scholar] [CrossRef]
- Tan, D.X.; Hardeland, R.; Manchester, L.C.; Poeggeler, B.; Lopez-Burillo, S.; Mayo, J.C.; Sainz, R.M.; Reiter, R.J. Mechanistic and comparative studies of melatonin and classic antioxidants in terms of their interactions with the ABTS cation radical. J. Pineal Res. 2003, 34, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Ressmeyer, A.R.; Mayo, J.C.; Zelosko, V.; Sainz, R.M.; Tan, D.X.; Poeggeler, B.; Antolin, I.; Zsizsik, B.K.; Reiter, R.J.; Hardeland, R. Antioxidant properties of the melatonin metabolite N1-acetyl-5-methoxykynuramine (AMK): Scavenging of free radicals and prevention of protein destruction. Redox Rep. 2003, 8, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Manchester, L.C.; Terron, M.P.; Flores, L.J.; Reiter, R.J. One molecule, many derivatives: A never-ending interaction of melatonin with reactive oxygen and nitrogen species? J. Pineal Res. 2007, 42, 28–42. [Google Scholar] [CrossRef] [PubMed]
- Escames, G.; Lopez, L.C.; Ortiz, F.; Ros, E.; Acuna-Castroviejo, D. Age-dependent lipopolysaccharide-induced iNOS expression and multiorgan failure in rats: Effects of melatonin treatment. Exp. Gerontol. 2006, 41, 1165–1173. [Google Scholar] [CrossRef] [PubMed]
- Sonmez, M.F.; Narin, F.; Akkus, D.; Ozdamar, S. Effect of melatonin and vitamin C on expression of endothelial NOS in heart of chronic alcoholic rats. Toxicol. Ind. Health 2009, 25, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Buldak, R.J.; Pilc-Gumula, K.; Buldak, L.; Witkowska, D.; Kukla, M.; Polaniak, R.; Zwirska-Korczala, K. Effects of ghrelin, leptin and melatonin on the levels of reactive oxygen species, antioxidant enzyme activity and viability of the HCT 116 human colorectal carcinoma cell line. Mol. Med. Rep. 2015, 12, 2275–2282. [Google Scholar] [PubMed]
- Tesoriere, L.; Allegra, M.; D’Arpa, D.; Butera, D.; Livrea, M.A. Reaction of melatonin with hemoglobin-derived oxoferryl radicals and inhibition of the hydroperoxide-induced hemoglobin denaturation in red blood cells. J. Pineal Res. 2001, 31, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Pi, H.; Yang, Z.; Reiter, R.J.; Xu, S.; Chen, X.; Chen, C.; Zhang, L.; Yang, M.; Li, Y.; et al. Melatonin antagonizes cadmium-induced neurotoxicity by activating the transcription factor EB-dependent autophagy-lysosome machinery in mouse neuroblastoma cells. J. Pineal Res. 2016, 61, 353–369. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.V.; Chhunchha, B. Protective role of melatonin against the mercury induced oxidative stress in the rat thyroid. Food Chem. Toxicol. 2010, 48, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wei, Z.; Liu, W.; Wang, J.; He, X.; Huang, H.; Zhang, J.; Yang, Z. Melatonin protects against arsenic trioxide-induced liver injury by the upregulation of Nrf2 expression through the activation of PI3K/AKT pathway. Oncotarget 2017, 8, 3773–3780. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Plata, E.; Quiroz-Compean, F.; Ramirez-Garcia, G.; Barrientos, E.Y.; Rodriguez-Morales, N.M.; Flores, A.; Wrobel, K.; Wrobel, K.; Mendez, I.; Diaz-Munoz, M.; et al. Melatonin reduces lead levels in blood, brain and bone and increases lead excretion in rats subjected to subacute lead treatment. Toxicol. Lett. 2015, 233, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Karabulut-Bulan, O.; Bayrak, B.B.; Arda-Pirincci, P.; Sarikaya-Unal, G.; Us, H.; Yanardag, R. Role of exogenous melatonin on cell proliferation and oxidant/antioxidant system in aluminum-induced renal toxicity. Biol. Trace Elem. Res. 2015, 168, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Alarcon, M.; Ruiz-Ojeda, F.J.; Blanca-Herrera, R.M.; Kaki, A.; Adem, A.; Agil, A. Melatonin administration in diabetes: Regulation of plasma Cr, V, and Mg in young male Zucker diabetic fatty rats. Food Funct. 2014, 5, 512–516. [Google Scholar] [CrossRef] [PubMed]
- Leon, J.; Escames, G.; Rodriguez, M.I.; Lopez, L.C.; Tapias, V.; Entrena, A.; Camacho, E.; Carrion, M.D.; Gallo, M.A.; Espinosa, A.; et al. Inhibition of neuronal nitric oxide synthase activity by N1-acetyl-5-methoxykynuramine, a brain metabolite of melatonin. J. Neurochem. 2006, 98, 2023–2033. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Campa, C.; Gonzalez, A.; Mediavilla, M.D.; Alonso-Gonzalez, C.; Alvarez-Garcia, V.; Sanchez-Barcelo, E.J.; Cos, S. Melatonin inhibits aromatase promoter expression by regulating cyclooxygenases expression and activity in breast cancer cells. Br. J. Cancer 2009, 101, 1613–1619. [Google Scholar] [CrossRef] [PubMed]
- Mayo, J.C.; Sainz, R.M.; Antoli, I.; Herrera, F.; Martin, V.; Rodriguez, C. Melatonin regulation of antioxidant enzyme gene expression. Cell. Mol. Life Sci. 2002, 59, 1706–1713. [Google Scholar] [CrossRef] [PubMed]
- Mehrzadi, S.; Safa, M.; Kamrava, S.K.; Darabi, R.; Hayat, P.; Motevalian, M. Protective mechanisms of melatonin against hydrogen peroxide induced toxicity in human bone-marrow derived mesenchymal stem cells. Can. J. Physiol. Pharmacol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Bilici, D.; Suleyman, H.; Banoglu, Z.N.; Kiziltunc, A.; Avci, B.; Ciftcioglu, A.; Bilici, S. Melatonin prevents ethanol-induced gastric mucosal damage possibly due to its antioxidant effect. Dig. Dis. Sci. 2002, 47, 856–861. [Google Scholar] [CrossRef] [PubMed]
- Ciftci, M.; Bilici, D.; Kufrevioglu, O.I. Effects of melatonin on enzyme activities of glucose-6-phosphate dehydrogenase from human erythrocytes in vitro and from rat erythrocytes in vivo. Pharmacol. Res. 2001, 44, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Gitto, E.; Tan, D.X.; Reiter, R.J.; Karbownik, M.; Manchester, L.C.; Cuzzocrea, S.; Fulia, F.; Barberi, I. Individual and synergistic antioxidative actions of melatonin: Studies with vitamin E, vitamin C, glutathione and desferrioxamine (desferoxamine) in rat liver homogenates. J. Pharm. Pharmacol. 2001, 53, 1393–1401. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Zhang, F.; Ge, X.; Yan, T.; Chen, X.; Shi, X.; Zhai, Q. SIRT1 improves insulin sensitivity under insulin-resistant conditions by repressing PTP1B. Cell Metab. 2007, 6, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Chahbouni, M.; Escames, G.; Venegas, C.; Sevilla, B.; Garcia, J.A.; Lopez, L.C.; Munoz-Hoyos, A.; Molina-Carballo, A.; Acuna-Castroviejo, D. Melatonin treatment normalizes plasma pro-inflammatory cytokines and nitrosative/oxidative stress in patients suffering from Duchenne muscular dystrophy. J. Pineal Res. 2010, 48, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Kepka, M.; Szwejser, E.; Pijanowski, L.; Verburg-van Kemenade, B.M.; Chadzinska, M. A role for melatonin in maintaining the pro- and anti-inflammatory balance by influencing leukocyte migration and apoptosis in carp. Dev. Comp. Immunol. 2015, 53, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Perkins, N.D. Integrating cell-signalling pathways with NF-kappaB and IKK function. Nat. Rev. Mol. Cell Biol. 2007, 8, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Gilmore, T.D. Introduction to NF-kappaB: Players, pathways, perspectives. Oncogene 2006, 25, 6680–6684. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.A.; Volt, H.; Venegas, C.; Doerrier, C.; Escames, G.; Lopez, L.C.; Acuna-Castroviejo, D. Disruption of the NF-kappaB/NLRP3 connection by melatonin requires retinoid-related orphan receptor-alpha and blocks the septic response in mice. FASEB J. 2015, 29, 3863–3875. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.P.; Fang, X.L.; Fang, N.; Wang, X.B.; Qian, H.Y.; Cao, Z.; Cheng, Y.; Wang, B.N.; Wang, Y. Melatonin ameliorates vascular endothelial dysfunction, inflammation, and atherosclerosis by suppressing the TLR4/NF-kappaB system in high-fat-fed rabbits. J. Pineal Res. 2013, 55, 388–398. [Google Scholar] [PubMed]
- Yi, C.; Zhang, Y.; Yu, Z.; Xiao, Y.; Wang, J.; Qiu, H.; Yu, W.; Tang, R.; Yuan, Y.; Guo, W.; et al. Melatonin enhances the anti-tumor effect of fisetin by inhibiting COX-2/iNOS and NF-kappaB/p300 signaling pathways. PLoS ONE 2014, 9, e99943. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.J.; Fu, L.; Tang, Z.; Zhang, C.; Qin, L.; Wang, J.; Yu, Z.; Shi, D.; Xiao, X.; Xie, F.; et al. Melatonin inhibits AP-2beta/hTERT, NF-kappaB/COX-2 and Akt/ERK and activates caspase/Cyto C signaling to enhance the antitumor activity of berberine in lung cancer cells. Oncotarget 2016, 7, 2985–3001. [Google Scholar] [PubMed]
- Mayo, J.C.; Sainz, R.M.; Tan, D.X.; Hardeland, R.; Leon, J.; Rodriguez, C.; Reiter, R.J. Anti-inflammatory actions of melatonin and its metabolites, N1-acetyl-N2-formyl-5-methoxykynuramine (AFMK) and N1-acetyl-5-methoxykynuramine (AMK), in macrophages. J. Neuroimmunol. 2005, 165, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Permpoonputtana, K.; Govitrapong, P. The anti-inflammatory effect of melatonin on methamphetamine-induced proinflammatory mediators in human neuroblastoma dopamine SH-SY5Y cell lines. Neurotox. Res. 2013, 23, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Niranjan, R.; Nath, C.; Shukla, R. Melatonin attenuated mediators of neuroinflammation and alpha-7 nicotinic acetylcholine receptor mRNA expression in lipopolysaccharide (LPS) stimulated rat astrocytoma cells, C6. Free Radic. Res. 2012, 46, 1167–1177. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.D.; Kim, Y.S.; Ko, S.H.; Yoon, I.J.; Cho, S.G.; Chun, Y.H.; Choi, B.J.; Kim, E.C. Cytoprotective and anti-inflammatory effects of melatonin in hydrogen peroxide-stimulated CHON-001 human chondrocyte cell line and rabbit model of osteoarthritis via the SIRT1 pathway. J. Pineal Res. 2012, 53, 225–237. [Google Scholar] [CrossRef] [PubMed]
- Min, K.J.; Jang, J.H.; Kwon, T.K. Inhibitory effects of melatonin on the lipopolysaccharide-induced CC chemokine expression in BV2 murine microglial cells are mediated by suppression of Akt-induced NF-kappaB and STAT/GAS activity. J. Pineal Res. 2012, 52, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.H.; Li, Y.C.; Wang, Z.Q.; Liu, B.; Ye, W.; Ni, L.; Zeng, R.; Miao, S.Y.; Wang, L.F.; Liu, C.W. Protective effect of melatonin on cigarette smoke-induced restenosis in rat carotid arteries after balloon injury. J. Pineal Res. 2014, 57, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Shao, G.; Tian, Y.; Wang, H.; Liu, F.; Xie, G. Protective effects of melatonin on lipopolysaccharide-induced mastitis in mice. Int. Immunopharmacol. 2015, 29, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, M.D.; Garcia-Moreno, H.; Gonzalez-Yanes, C.; Calvo, J.R. Possible involvement of the inhibition of NF-kappaB factor in anti-inflammatory actions that melatonin exerts on mast cells. J. Cell Biochem. 2016, 117, 1926–1933. [Google Scholar] [CrossRef] [PubMed]
- Wilson, B.J.; Tremblay, A.M.; Deblois, G.; Sylvain-Drolet, G.; Giguere, V. An acetylation switch modulates the transcriptional activity of estrogen-related receptor alpha. Mol. Endocrinol. 2010, 24, 1349–1358. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, J.T.; Lerin, C.; Haas, W.; Gygi, S.P.; Spiegelman, B.M.; Puigserver, P. Nutrient control of glucose homeostasis through a complex of PGC-1alpha and SIRT1. Nature 2005, 434, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.A.; Khan, M.; Jo, M.H.; Jo, M.G.; Amin, F.U.; Kim, M.O. Melatonin stimulates the SIRT1/Nrf2 signaling pathway counteracting lipopolysaccharide (LPS)-induced oxidative stress to rescue postnatal rat brain. CNS Neurosci. Ther. 2016. [Google Scholar] [CrossRef] [PubMed]
- Ge, D.; Dauchy, R.T.; Liu, S.; Zhang, Q.; Mao, L.; Dauchy, E.M.; Blask, D.E.; Hill, S.M.; Rowan, B.G.; Brainard, G.C.; et al. Insulin and IGF1 enhance IL-17-induced chemokine expression through a GSK3B-dependent mechanism: A new target for melatonin’s anti-inflammatory action. J. Pineal Res. 2013, 55, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Koyama, F.C.; Azevedo, M.F.; Budu, A.; Chakrabarti, D.; Garcia, C.R. Melatonin-induced temporal up-regulation of gene expression related to ubiquitin/proteasome system (UPS) in the human malaria parasite Plasmodium falciparum. Int. J. Mol. Sci. 2014, 15, 22320–22330. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.H.; Park, Y.S.; Kim, O.S.; Kim, J.H.; Baik, H.W.; Hong, Y.O.; Kim, S.S.; Shin, J.H.; Jun, J.H.; Jo, Y.; et al. Melatonin attenuates dextran sodium sulfate induced colitis with sleep deprivation: Possible mechanism by microarray analysis. Dig. Dis. Sci. 2014, 59, 1134–1141. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Ni, Y.F.; Wang, W.C.; Du, H.Y.; Zhang, H.; Zhang, L.; Zhang, W.D.; Jiang, T. Melatonin attenuates intestinal ischemia—Reperfusion-induced lung injury in rats by upregulating N-myc downstream-regulated gene 2. J. Surg. Res. 2015, 194, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Ghosh, S.; Basu, P.; Haldar, C. Daily variation in melatonin level, antioxidant activity and general immune response of peripheral blood mononuclear cells and lymphoid tissues of Indian goat Capra hircus during summer and winter. Indian J. Exp. Biol. 2014, 52, 467–477. [Google Scholar] [PubMed]
- Zhang, M.; Wang, T.; Chen, H.M.; Chen, Y.Q.; Deng, Y.C.; Li, Y.T. Serum levels of interleukin-1 beta, interleukin-6 and melatonin over summer and winter in kidney deficiency syndrome in Bizheng rats. Chin. Med. Sci. J. 2014, 29, 107–111. [Google Scholar] [CrossRef]
- Provinciali, M.; Di Stefano, G.; Bulian, D.; Tibaldi, A.; Fabris, N. Effect of melatonin and pineal grafting on thymocyte apoptosis in aging mice. Mech. Ageing Dev. 1996, 90, 1–19. [Google Scholar] [CrossRef]
- Fernandes, P.A.; Cecon, E.; Markus, R.P.; Ferreira, Z.S. Effect of TNF-alpha on the melatonin synthetic pathway in the rat pineal gland: Basis for a ‘feedback’ of the immune response on circadian timing. J. Pineal Res. 2006, 41, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Lissoni, P.; Rovelli, F.; Giani, L.; Fumagalli, L.; Mandala, M. Immunomodulatory effects of IL-12 in relation to the pineal endocrine function in metastatic cancer patients. Nat. Immun. 1998, 16, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Withyachumnarnkul, B.; Nonaka, K.O.; Santana, C.; Attia, A.M.; Reiter, R.J. Interferon-gamma modulates melatonin production in rat pineal glands in organ culture. J. Interferon Res. 1990, 10, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Manchester, L.C.; Reiter, R.J. CSF generation by pineal gland results in a robust melatonin circadian rhythm in the third ventricle as an unique light/dark signal. Med. Hypotheses 2016, 86, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Markus, R.P.; Cecon, E.; Pires-Lapa, M.A. Immune-pineal axis: Nuclear factor kappaB (NF-êB) mediates the shift in the melatonin source from pinealocytes to immune competent cells. Int. J. Mol. Sci. 2013, 14, 10979–10997. [Google Scholar] [CrossRef] [PubMed]
- Pontes, G.N.; Cardoso, E.C.; Carneiro-Sampaio, M.M.; Markus, R.P. Injury switches melatonin production source from endocrine (pineal) to paracrine (phagocytes)—Melatonin in human colostrum and colostrum phagocytes. J. Pineal Res. 2006, 41, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, R.; Haldar, C. Photoperiodic regulation of MT1 and MT2 melatonin receptor expression in spleen and thymus of a tropical rodent Funambulus pennanti during reproductively active and inactive phases. Chronobiol. Int. 2010, 27, 446–462. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Perganeda, A.; Pozo, D.; Guerrero, J.M.; Calvo, J.R. Signal transduction for melatonin in human lymphocytes: Involvement of a pertussis toxin-sensitive G protein. J. Immunol. 1997, 159, 3774–3781. [Google Scholar] [PubMed]
- Vishwas, D.K.; Haldar, C. Photoperiodic induced melatonin regulates immunity and expression pattern of melatonin receptor MT1 in spleen and bone marrow mononuclear cells of male golden hamster. J. Photochem. Photobiol. B 2013, 128, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Lardone, P.J.; Rubio, A.; Cerrillo, I.; Gomez-Corvera, A.; Carrillo-Vico, A.; Sanchez-Hidalgo, M.; Guerrero, J.M.; Fernandez-Riejos, P.; Sanchez-Margalet, V.; Molinero, P. Blocking of melatonin synthesis and MT(1) receptor impairs the activation of Jurkat T cells. Cell Mol. Life Sci. 2010, 67, 3163–3172. [Google Scholar] [CrossRef] [PubMed]
- Mohseni, M.; Mihandoost, E.; Shirazi, A.; Sepehrizadeh, Z.; Bazzaz, J.T.; Ghazi-khansari, M. Melatonin may play a role in modulation of bax and bcl-2 expression levels to protect rat peripheral blood lymphocytes from gamma irradiation-induced apoptosis. Mutat. Res. 2012, 738–739, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Dong, Y.; Cao, J.; Wang, Z.; Zhang, Z.; Chen, Y. Developmental changes of melatonin receptor expression in the spleen of the chicken, Gallus domesticus. Acta Histochem. 2015, 117, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, M.K.; Hubbard, G.B.; Champney, T.H.; Vaughan, G.M.; Little, J.C.; Reiter, R.J. Splenic hypertrophy and extramedullary hematopoiesis induced in male Syrian hamsters by short photoperiod or melatonin injections and reversed by melatonin pellets or pinealectomy. Am. J. Anat. 1987, 179, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Lopez Gonzalez, M.A.; Guerrero, J.M.; Ceballo Pedraja, J.M.; Delgado Moreno, F. Melatonin in palate tonsils with recurrent acute tonsillitis and tonsillar hypertrophy. Acta Otorrinolaringol. Esp. 1998, 49, 625–628. [Google Scholar] [PubMed]
- Barjavel, M.J.; Mamdouh, Z.; Raghbate, N.; Bakouche, O. Differential expression of the melatonin receptor in human monocytes. J. Immunol. 1998, 160, 1191–1197. [Google Scholar] [PubMed]
- Kim, Y.O.; Ahn, Y.K.; Kim, J.H. Influence of melatonin on immunotoxicity of cadmium. Int. J. Immunopharmacol. 2000, 22, 275–284. [Google Scholar] [CrossRef]
- Silva, S.O.; Rodrigues, M.R.; Ximenes, V.F.; Bueno-da-Silva, A.E.; Amarante-Mendes, G.P.; Campa, A. Neutrophils as a specific target for melatonin and kynuramines: Effects on cytokine release. J. Neuroimmunol. 2004, 156, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Suke, S.G.; Pathak, R.; Ahmed, R.S.; Tripathi, A.K.; Banerjee, B.D. Melatonin treatment prevents modulation of cell-mediated immune response induced by propoxur in rats. Indian J. Biochem. Biophys. 2008, 45, 278–281. [Google Scholar] [PubMed]
- Ghosh, S.; Singh, A.K.; Haldar, C. Seasonal modulation of immunity by melatonin and gonadal steroids in a short day breeder goat Capra hircus. Theriogenology 2014, 82, 1121–1130. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Maurino, S.; Pozo, D.; Carrillo-Vico, A.; Calvo, J.R.; Guerrero, J.M. Melatonin activates Th1 lymphocytes by increasing IL-12 production. Life Sci. 1999, 65, 2143–2150. [Google Scholar] [CrossRef]
- Xia, M.Z.; Liang, Y.L.; Wang, H.; Chen, X.; Huang, Y.Y.; Zhang, Z.H.; Chen, Y.H.; Zhang, C.; Zhao, M.; Xu, D.X.; et al. Melatonin modulates TLR4-mediated inflammatory genes through MyD88- and TRIF-dependent signaling pathways in lipopolysaccharide-stimulated RAW264.7 cells. J. Pineal Res. 2012, 53, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Carrillo-Vico, A.; Reiter, R.J.; Lardone, P.J.; Herrera, J.L.; Fernandez-Montesinos, R.; Guerrero, J.M.; Pozo, D. The modulatory role of melatonin on immune responsiveness. Curr. Opin. Investig. Drugs 2006, 7, 423–431. [Google Scholar] [PubMed]
- Carrillo-Vico, A.; Lardone, P.J.; Fernandez-Santos, J.M.; Martin-Lacave, I.; Calvo, J.R.; Karasek, M.; Guerrero, J.M. Human lymphocyte-synthesized melatonin is involved in the regulation of the interleukin-2/interleukin-2 receptor system. J. Clin. Endocrinol. Metab. 2005, 90, 992–1000. [Google Scholar] [CrossRef] [PubMed]
- Morrey, K.M.; McLachlan, J.A.; Serkin, C.D.; Bakouche, O. Activation of human monocytes by the pineal hormone melatonin. J. Immunol. 1994, 153, 2671–2680. [Google Scholar] [PubMed]
- Bouhafs, R.K.; Jarstrand, C. Effects of antioxidants on surfactant peroxidation by stimulated human polymorphonuclear leukocytes. Free Radic. Res. 2002, 36, 727–734. [Google Scholar] [CrossRef] [PubMed]
- Vishwas, D.K.; Mukherjee, A.; Haldar, C.; Dash, D.; Nayak, M.K. Improvement of oxidative stress and immunity by melatonin: An age dependent study in golden hamster. Exp. Gerontol. 2013, 48, 168–182. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.K.; Haldar, C. Experimentally induced stress, oxidative load and changes in immunity in a tropical wild bird, Perdicula asiatica: Involvement of melatonin and glucocorticoid receptors. Zoology 2014, 117, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, M. Pharmacology of ramelteon, a selective MT1/MT2 receptor agonist: A novel therapeutic drug for sleep disorders. CNS Neurosci. Ther. 2009, 15, 32–51. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.S.; Green, E.W.; Zhao, Y.; van Ooijen, G.; Olmedo, M.; Qin, X.; Xu, Y.; Pan, M.; Valekunja, U.K.; Feeney, K.A.; et al. Peroxiredoxins are conserved markers of circadian rhythms. Nature 2012, 485, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Sturgeon, S.R.; Luisi, N.; Balasubramanian, R.; Reeves, K.W. Sleep duration and endometrial cancer risk. Cancer Causes Control 2012, 23, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Gan, L.; Luo, D.; Sun, C. Melatonin promotes circadian rhythm-induced proliferation through interaction of Clock/HDAC3/c-Myc in mice adipose tissue. J. Pineal Res. 2016. [Google Scholar] [CrossRef]
- Tamura, H.; Takasaki, A.; Taketani, T.; Tanabe, M.; Lee, L.; Tamura, I.; Maekawa, R.; Aasada, H.; Yamagata, Y.; Sugino, N. Melatonin and female reproduction. J. Obstet. Gynaecol. Res. 2014, 40, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lundmark, P.O.; Pandi-Perumal, S.R.; Srinivasan, V.; Cardinali, D.P.; Rosenstein, R.E. Melatonin in the eye: Implications for glaucoma. Exp. Eye Res. 2007, 84, 1021–1030. [Google Scholar] [CrossRef] [PubMed]
- Espino, J.; Pariente, J.A.; Rodriguez, A.B. Role of melatonin on diabetes-related metabolic disorders. World J. Diabetes 2011, 2, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R. Neurobiology, pathophysiology, and treatment of melatonin deficiency and dysfunction. Sci. World J. 2012, 2012, 640389. [Google Scholar] [CrossRef] [PubMed]
- Torres-Farfan, C.; Seron-Ferre, M.; Dinet, V.; Korf, H.W. Immunocytochemical demonstration of day/night changes of clock gene protein levels in the murine adrenal gland: Differences between melatonin-proficient (C3H) and melatonin-deficient (C57BL) mice. J. Pineal Res. 2006, 40, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Johnston, J.D.; Tournier, B.B.; Andersson, H.; Masson-Pevet, M.; Lincoln, G.A.; Hazlerigg, D.G. Multiple effects of melatonin on rhythmic clock gene expression in the mammalian pars tuberalis. Endocrinology 2006, 147, 959–965. [Google Scholar] [CrossRef] [PubMed]
- Dinet, V.; Ansari, N.; Torres-Farfan, C.; Korf, H.W. Clock gene expression in the retina of melatonin-proficient (C3H) and melatonin-deficient (C57BL) mice. J. Pineal Res. 2007, 42, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Chellappa, S.L.; Viola, A.U.; Schmidt, C.; Bachmann, V.; Gabel, V.; Maire, M.; Reichert, C.F.; Valomon, A.; Gotz, T.; Landolt, H.P.; et al. Human melatonin and alerting response to blue-enriched light depend on a polymorphism in the clock gene PER3. J. Clin. Endocrinol. Metab. 2012, 97, E433–E437. [Google Scholar] [CrossRef] [PubMed]
- West, A.; Dupre, S.M.; Yu, L.; Paton, I.R.; Miedzinska, K.; McNeilly, A.S.; Davis, J.R.; Burt, D.W.; Loudon, A.S. Npas4 is activated by melatonin, and drives the clock gene Cry1 in the ovine pars tuberalis. Mol. Endocrinol. 2013, 27, 979–989. [Google Scholar] [CrossRef] [PubMed]
- Bracci, M.; Manzella, N.; Copertaro, A.; Staffolani, S.; Strafella, E.; Barbaresi, M.; Copertaro, B.; Rapisarda, V.; Valentino, M.; Santarelli, L. Rotating-shift nurses after a day off: Peripheral clock gene expression, urinary melatonin, and serum 17-beta-estradiol levels. Scand. J. Work Environ. Health 2014, 40, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Ikeno, T.; Nelson, R.J. Acute melatonin treatment alters dendritic morphology and circadian clock gene expression in the hippocampus of Siberian hamsters. Hippocampus 2015, 25, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Nagy, A.D.; Iwamoto, A.; Kawai, M.; Goda, R.; Matsuo, H.; Otsuka, T.; Nagasawa, M.; Furuse, M.; Yasuo, S. Melatonin adjusts the expression pattern of clock genes in the suprachiasmatic nucleus and induces antidepressant-like effect in a mouse model of seasonal affective disorder. Chronobiol. Int. 2015, 32, 447–457. [Google Scholar] [CrossRef] [PubMed]
- Rodenbeck, A.; Hajak, G. Neuroendocrine dysregulation in primary insomnia. Rev. Neurol. 2001, 157, S57–S61. [Google Scholar] [PubMed]
- Takaesu, Y.; Futenma, K.; Kobayashi, M.; Komada, Y.; Tanaka, N.; Yamashina, A.; Inoue, Y. A preliminary study on the relationships between diurnal melatonin secretion profile and sleep variables in patients emergently admitted to the coronary care unit. Chronobiol. Int. 2015, 32, 875–879. [Google Scholar] [CrossRef] [PubMed]
- Evely, K.M.; Hudson, R.L.; Dubocovich, M.L.; Haj-Dahmane, S. Melatonin receptor activation increases glutamatergic synaptic transmission in the rat medial lateral habenula. Synapse 2016, 70, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Waldhauser, F.; Kovacs, J.; Reiter, E. Age-related changes in melatonin levels in humans and its potential consequences for sleep disorders. Exp. Gerontol. 1998, 33, 759–772. [Google Scholar] [CrossRef]
- Carpentieri, A.; Diaz De Barboza, G.; Areco, V.; Peralta Lopez, M.; Tolosa De Talamoni, N. New perspectives in melatonin uses. Pharmacol. Res. 2012, 65, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Lemoine, P.; Zisapel, N. Prolonged-release formulation of melatonin (Circadin) for the treatment of insomnia. Expert Opin. Pharmacother. 2012, 13, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, V.; Pandi-Perumal, S.R.; Trahkt, I.; Spence, D.W.; Poeggeler, B.; Hardeland, R.; Cardinali, D.P. Melatonin and melatonergic drugs on sleep: Possible mechanisms of action. Int. J. Neurosci. 2009, 119, 821–846. [Google Scholar] [CrossRef] [PubMed]
- Vimala, P.V.; Bhutada, P.S.; Patel, F.R. Therapeutic potential of agomelatine in epilepsy and epileptic complications. Med. Hypotheses 2014, 82, 105–110. [Google Scholar] [CrossRef] [PubMed]
- WHO (World Health Organization). Cancer Fact Sheet No. 297. Available online: https://www.google.com.hk/url?sa=t&rct=j&q=&esrc=s&source=web&cd=2&ved=0ahUKEwjT_cjhzYfTAhVIqJQKHYFWCRgQFggjMAE&url=http%3A%2F%2Fwww.afro.who.int%2Findex.php%3Foption%3Dcom_docman%26task%3Ddoc_download%26gid%3D6139&usg=AFQjCNE1HISbfHk2ijLPCzy3VxfgMAbK3Q (accessed on 20 November 2016).
- Li, F.; Li, S.; Li, H.B.; Deng, G.F.; Ling, W.H.; Xu, X.R. Antiproliferative activities of tea and herbal infusions. Food Funct. 2013, 4, 530–538. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Zhou, Y.; Li, Y.; Xu, D.P.; Li, S.; Li, H.B. Spices for prevention and treatment of cancers. Nutrients 2016, 8, 495. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Li, Y.; Zhou, T.; Zheng, J.; Li, S.; Li, H.B. Dietary natural products for prevention and treatment of liver cancer. Nutrients 2016, 8, 156. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zheng, J.; Li, Y.; Xu, D.P.; Li, S.; Chen, Y.M.; Li, H.B. Natural polyphenols for prevention and treatment of cancer. Nutrients 2016, 8, 515. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.M.; Lin, W.Y.; Shen, C.C.; Pan, H.C.; Keh-Bin, W.; Chen, Y.C.; Jan, Y.J.; Lai, D.W.; Tang, S.C.; Tien, H.R.; et al. Melatonin set out to ER stress signaling thwarts epithelial mesenchymal transition and peritoneal dissemination via calpain-mediated C/EBPbeta and NFkappaB cleavage. J. Pineal Res. 2016, 60, 142–154. [Google Scholar] [CrossRef] [PubMed]
- Hevia, D.; Gonzalez-Menendez, P.; Quiros-Gonzalez, I.; Miar, A.; Rodriguez-Garcia, A.; Tan, D.X.; Reiter, R.J.; Mayo, J.C.; Sainz, R.M. Melatonin uptake through glucose transporters: A new target for melatonin inhibition of cancer. J. Pineal Res. 2015, 58, 234–250. [Google Scholar] [CrossRef] [PubMed]
- Yun, S.M.; Woo, S.H.; Oh, S.T.; Hong, S.E.; Choe, T.B.; Ye, S.K.; Kim, E.K.; Seong, M.K.; Kim, H.A.; Noh, W.C.; et al. Melatonin enhances arsenic trioxide-induced cell death via sustained upregulation of Redd1 expression in breast cancer cells. Mol. Cell Endocrinol. 2016, 422, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Borin, T.F.; Arbab, A.S.; Gelaleti, G.B.; Ferreira, L.C.; Moschetta, M.G.; Jardim-Perassi, B.V.; Iskander, A.S.; Varma, N.R.; Shankar, A.; Coimbra, V.B.; et al. Melatonin decreases breast cancer metastasis by modulating Rho-associated kinase protein-1 expression. J. Pineal Res. 2016, 60, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Ziolko, E.; Kokot, T.; Skubis, A.; Sikora, B.; Szota-Czyz, J.; Kruszniewska-Rajs, C.; Wierzgon, J.; Mazurek, U.; Grochowska-Niedworok, E.; Muc-Wierzgon, M. The profile of melatonin receptors gene expression and genes associated with their activity in colorectal cancer: A preliminary report. J. Biol. Regul. Homeost. Agents 2015, 29, 823–828. [Google Scholar] [PubMed]
- Watanabe, M.; Kobayashi, Y.; Takahashi, N.; Kiguchi, K.; Ishizuka, B. Expression of melatonin receptor (MT1) and interaction between melatonin and estrogen in endometrial cancer cell line. J. Obstet. Gynaecol. Res. 2008, 34, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Chottanapund, S.; Van Duursen, M.B.; Navasumrit, P.; Hunsonti, P.; Timtavorn, S.; Ruchirawat, M.; Van den Berg, M. Anti-aromatase effect of resveratrol and melatonin on hormonal positive breast cancer cells co-cultured with breast adipose fibroblasts. Toxicol. In Vitro 2014, 28, 1215–1221. [Google Scholar] [CrossRef] [PubMed]
- Sohn, E.J.; Won, G.; Lee, J.; Lee, S.; Kim, S.H. Upregulation of miRNA3195 and miRNA374b Mediates the anti-angiogenic properties of melatonin in Hypoxic PC-3 prostate cancer cells. J. Cancer 2015, 6, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.Y.; Li, W.M.; Zhou, L.L.; Lu, Q.N.; He, W. Melatonin induces apoptosis of colorectal cancer cells through HDAC4 nuclear import mediated by CaMKII inactivation. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 58, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Ordonez, R.; Carbajo-Pescador, S.; Prieto-Dominguez, N.; Garcia-Palomo, A.; Gonzalez-Gallego, J.; Mauriz, J.L. Inhibition of matrix metalloproteinase-9 and nuclear factor kappa B contribute to melatonin prevention of motility and invasiveness in HepG2 liver cancer cells. J. Pineal Res. 2014, 56, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.W.; Lee, L.M.; Lee, W.J.; Chu, C.Y.; Tan, P.; Yang, Y.C.; Chen, W.Y.; Yang, S.F.; Hsiao, M.; Chien, M.H. Melatonin inhibits MMP-9 transactivation and renal cell carcinoma metastasis by suppressing Akt-MAPKs pathway and NF-kappaB DNA-binding activity. J. Pineal Res. 2016, 60, 277–290. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Gonzalez, C.; Gonzalez, A.; Martinez-Campa, C.; Menendez-Menendez, J.; Gomez-Arozamena, J.; Garcia-Vidal, A.; Cos, S. Melatonin enhancement of the radiosensitivity of human breast cancer cells is associated with the modulation of proteins involved in estrogen biosynthesis. Cancer Lett. 2016, 370, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Ben-David, M.A.; Elkayam, R.; Gelernter, I.; Pfeffer, R.M. Melatonin for prevention of breast radiation dermatitis: A phase II, prospective, double-blind rendomized trial. Isr. Med. Assoc. J. 2016, 18, 188–192. [Google Scholar] [PubMed]
- Madhu, P.; Reddy, K.P.; Reddy, P.S. Role of melatonin in mitigating chemotherapy-induced testicular dysfunction in Wistar rats. Drug Chem. Toxicol. 2016, 39, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.M.; Jin, B.Z.; Ai, F.; Duan, C.H.; Lu, Y.Z.; Dong, T.F.; Fu, Q.L. The efficacy and safety of melatonin in concurrent chemotherapy or radiotherapy for solid tumors: A meta-analysis of randomized controlled trials. Cancer Chemother. Pharmacol. 2012, 69, 1213–1220. [Google Scholar] [CrossRef] [PubMed]
- WHO. Cardiovascular Diseases. Available online: http://www.who.int/mediacentre/factsheets/fs317/en/ (accessed on 8 December 2016).
- Wolf, K.; Braun, A.; Haining, E.J.; Tseng, Y.L.; Kraft, P.; Schuhmann, M.K.; Gotru, S.K.; Chen, W.; Hermanns, H.M.; Stoll, G.; et al. Partially defective store operated calcium entry and Hem(ITAM) signaling in platelets of serotonin transporter deficient Mice. PLoS ONE 2016, 11, e147664. [Google Scholar] [CrossRef] [PubMed]
- Grossman, E.; Laudon, M.; Yalcin, R.; Zengil, H.; Peleg, E.; Sharabi, Y.; Kamari, Y.; Shen-Orr, Z.; Zisapel, N. Melatonin reduces night blood pressure in patients with nocturnal hypertension. Am. J. Med. 2006, 119, 898–902. [Google Scholar] [CrossRef] [PubMed]
- Arangino, S.; Cagnacci, A.; Angiolucci, M.; Vacca, A.M.; Longu, G.; Volpe, A.; Melis, G.B. Effects of melatonin on vascular reactivity, catecholamine levels, and blood pressure in healthy men. Am. J. Cardiol. 1999, 83, 1417–1419. [Google Scholar] [CrossRef]
- Wu, Q.; Jing, Y.; Yuan, X.; Zhang, X.; Li, B.; Liu, M.; Wang, B.; Li, H.; Liu, S.; Xiu, R. Melatonin treatment protects against acute spinal cord injury-induced disruption of blood spinal cord barrier in mice. J. Mol. Neurosci. 2014, 54, 714–722. [Google Scholar] [CrossRef] [PubMed]
- Girouard, H.; Denault, C.; Chulak, C.; de Champlain, J. Treatment by N-acetylcysteine and melatonin increases cardiac baroreflex and improves antioxidant reserve. Am. J. Hypertens. 2004, 17, 947–954. [Google Scholar] [CrossRef] [PubMed]
- Girouard, H.; Chulak, C.; LeJossec, M.; Lamontagne, D.; de Champlain, J. Chronic antioxidant treatment improves sympathetic functions and beta-adrenergic pathway in the spontaneously hypertensive rats. J. Hypertens. 2003, 21, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Gurses, I.; Ozeren, M.; Serin, M.; Yucel, N.; Erkal, H.S. Histopathological evaluation of melatonin as a protective agent in heart injury induced by radiation in a rat model. Pathol. Res. Pract. 2014, 210, 863–871. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Fan, C.; Deng, C.; Zhao, L.; Hu, W.; Di, S.; Ma, Z.; Zhang, Y.; Qin, Z.; Jin, Z.; et al. Melatonin reverses flow shear stress-induced injury in bone marrow mesenchymal stem cells via activation of AMP-activated protein kinase signaling. J. Pineal Res. 2016, 60, 228–241. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Huang, W.; Li, X.; Gao, L.; Su, T.; Li, X.; Ma, S.; Liu, T.; Li, C.; Chen, J.; et al. Melatonin facilitates adipose-derived mesenchymal stem cells to repair the murine infarcted heart via the SIRT1 signaling pathway. J. Pineal Res. 2016, 60, 178–192. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Duan, W.; Jin, Z.; Yi, W.; Yan, J.; Zhang, S.; Wang, N.; Liang, Z.; Li, Y.; Chen, W.; et al. JAK2/STAT3 activation by melatonin attenuates the mitochondrial oxidative damage induced by myocardial ischemia/reperfusion injury. J. Pineal Res. 2013, 55, 275–286. [Google Scholar] [CrossRef] [PubMed]
- An, R.; Zhao, L.; Xi, C.; Li, H.; Shen, G.; Liu, H.; Zhang, S.; Sun, L. Melatonin attenuates sepsis-induced cardiac dysfunction via a PI3K/Akt-dependent mechanism. Basic Res. Cardiol. 2016, 111, 8. [Google Scholar] [CrossRef] [PubMed]
- VanKirk, T.; Powers, E.; Dowse, H.B. Melatonin increases the regularity of cardiac rhythmicity in the Drosophila heart in both wild-type and strains bearing pathogenic mutations. J. Comp. Physiol. B 2017, 187, 63–78. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Rodriguez, A.; Abreu-Gonzalez, P.; Piccolo, R.; Galasso, G.; Reiter, R.J. Melatonin is associated with reverse remodeling after cardiac resynchronization therapy in patients with heart failure and ventricular dyssynchrony. Int. J. Cardiol. 2016, 221, 359–363. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Zhang, L.; Yang, Y.; Guo, Y.; Fan, Y.; Zhang, M.; Man, W.; Gao, E.; Hu, W.; Reiter, R.J.; et al. Melatonin alleviates postinfarction cardiac remodeling and dysfunction by inhibiting Mst1. J. Pineal Res. 2017. [Google Scholar] [CrossRef] [PubMed]
- WHO. Global Status Report on Noncommunicable Diseases 2014. Available online: http://www.who.int/nmh/publications/ncd-status-report-2014/en/ (accessed on 9 December 2016).
- Simko, F.; Paulis, L. Melatonin as a potential antihypertensive treatment. J. Pineal Res. 2007, 42, 319–322. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Leon, J.; Kilic, U.; Kilic, E. When melatonin gets on your nerves: Its beneficial actions in experimental models of stroke. Exp. Biol. Med. 2005, 230, 104–117. [Google Scholar]
- Hung, M.W.; Kravtsov, G.M.; Lau, C.F.; Poon, A.M.; Tipoe, G.L.; Fung, M.L. Melatonin ameliorates endothelial dysfunction, vascular inflammation, and systemic hypertension in rats with chronic intermittent hypoxia. J. Pineal Res. 2013, 55, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Y.F.; Guo, W.J.; Li, L.; Shao, S.; Qiao, X.; Shao, J.J.; Zhang, Q.; Li, R.S.; Wang, L.H. Melatonin attenuates hypertension-induced renal injury partially through inhibiting oxidative stress in rats. Mol. Med. Rep. 2016, 13, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Pechanova, O.; Paulis, L.; Simko, F. Peripheral and central effects of melatonin on blood pressure regulation. Int. J. Mol. Sci. 2014, 15, 17920–17937. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.Y.; Shen, F.C.; Xie, D.; Han, Q.P.; Fang, M.; Chen, C.B.; Zeng, H.K. Progress in drug treatment of cerebral edema. Mini Rev. Med. Chem. 2016, 16, 917–925. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Lv, J.; Li, W.; Zhang, P.; Mao, C.; Xu, Z. Exogenous melatonin reduced blood pressure in late-term ovine fetus via MT1/MT2 receptor pathways. Reprod. Biol. 2016, 16, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Butun, I.; Ekmekci, H.; Ciftci, O.; Sonmez, H.; Caner, M.; Altug, T.; Kokoglu, E. The effects of different doses of melatonin on lipid peroxidation in diet-induced hypercholesterolemic rats. Bratisl. Lek. Listy 2013, 114, 129–132. [Google Scholar] [CrossRef] [PubMed]
- Salmanoglu, D.S.; Gurpinar, T.; Vural, K.; Ekerbicer, N.; Dariverenli, E.; Var, A. Melatonin and L-carnitin improves endothelial disfunction and oxidative stress in Type 2 diabetic rats. Redox Biol. 2016, 8, 199–204. [Google Scholar] [CrossRef] [PubMed]
- She, M.; Hu, X.; Su, Z.; Zhang, C.; Yang, S.; Ding, L.; Laudon, M.; Yin, W. Piromelatine, a novel melatonin receptor agonist, stabilizes metabolic profiles and ameliorates insulin resistance in chronic sleep restricted rats. Eur. J. Pharmacol. 2014, 727, 60–65. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Global Report on Diabetes. Available online: http://www.who.int/mediacentre/factsheets/fs312/en/ (accessed on 18 December 2016).
- Zavodnik, I.B.; Lapshina, E.A.; Cheshchevik, V.T.; Dremza, I.K.; Kujawa, J.; Zabrodskaya, S.V.; Reiter, R.J. Melatonin and succinate reduce rat liver mitochondrial dysfunction in diabetes. J. Physiol. Pharmacol. 2011, 62, 421–427. [Google Scholar] [PubMed]
- Faria, J.A.; Kinote, A.; Ignacio-Souza, L.M.; de Araujo, T.M.; Razolli, D.S.; Doneda, D.L.; Paschoal, L.B.; Lellis-Santos, C.; Bertolini, G.L.; Velloso, L.A.; et al. Melatonin acts through MT1/MT2 receptors to activate hypothalamic Akt and suppress hepatic gluconeogenesis in rats. Am. J. Physiol. Endocrinol. Metab. 2013, 305, E230–E242. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Liang, H.; Dong, X.; Zhao, G.; Jin, Z.; Zhai, M.; Yang, Y.; Chen, W.; Liu, J.; Yi, W.; et al. Reduced silent information regulator 1 signaling exacerbates myocardial ischemia-reperfusion injury in type 2 diabetic rats and the protective effect of melatonin. J. Pineal Res. 2015, 59, 376–390. [Google Scholar] [CrossRef] [PubMed]
- Costes, S.; Boss, M.; Thomas, A.P.; Matveyenko, A.V. Activation of melatonin signaling promotes beta-cell survival and function. Mol. Endocrinol. 2015, 29, 682–692. [Google Scholar] [CrossRef] [PubMed]
- Wongchitrat, P.; Lansubsakul, N.; Kamsrijai, U.; Sae-Ung, K.; Mukda, S.; Govitrapong, P. Melatonin attenuates the high-fat diet and streptozotocin-induced reduction in rat hippocampal neurogenesis. Neurochem. Int. 2016, 100, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Yildirimturk, S.; Batu, S.; Alatli, C.; Olgac, V.; Firat, D.; Sirin, Y. The effects of supplemental melatonin administration on the healing of bone defects in streptozotocin-induced diabetic rats. J. Appl. Oral Sci. 2016, 24, 239–249. [Google Scholar] [CrossRef] [PubMed]
- WHO. Obesity and Overweight. Available online: http://www.who.int/mediacentre/factsheets/fs311/en/ (accessed on 18 December 2016).
- Szewczyk-Golec, K.; Wozniak, A.; Reiter, R.J. Inter-relationships of the chronobiotic, melatonin, with leptin and adiponectin: Implications for obesity. J. Pineal Res. 2015, 59, 277–291. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Aranda, A.; Fernandez-Vazquez, G.; Campos, D.; Tassi, M.; Velasco-Perez, L.; Tan, D.X.; Reiter, R.J.; Agil, A. Melatonin induces browning of inguinal white adipose tissue in Zucker diabetic fatty rats. J. Pineal Res. 2013, 55, 416–423. [Google Scholar] [CrossRef] [PubMed]
- Winiarska, K.; Focht, D.; Sierakowski, B.; Lewandowski, K.; Orlowska, M.; Usarek, M. NADPH oxidase inhibitor, apocynin, improves renal glutathione status in Zucker diabetic fatty rats: A comparison with melatonin. Chem. Biol. Interact. 2014, 218, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Sharabiani, M.T.; Vermeulen, R.; Scoccianti, C.; Hosnijeh, F.S.; Minelli, L.; Sacerdote, C.; Palli, D.; Krogh, V.; Tumino, R.; Chiodini, P.; et al. Immunologic profile of excessive body weight. Biomarkers 2011, 16, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Favero, G.; Stacchiotti, A.; Castrezzati, S.; Bonomini, F.; Albanese, M.; Rezzani, R.; Rodella, L.F. Melatonin reduces obesity and restores adipokine patterns and metabolism in obese (ob/ob) mice. Nutr. Res. 2015, 35, 891–900. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.; Chatterjee, S.; Li, L.; Kim, J.M.; Lee, J.; Yechoor, V.K.; Minze, L.J.; Hsueh, W.; Ma, K. The clock gene, brain and muscle Arnt-like 1, regulates adipogenesis via Wnt signaling pathway. FASEB J. 2012, 26, 3453–3463. [Google Scholar] [CrossRef] [PubMed]
- Otway, D.T.; Mantele, S.; Bretschneider, S.; Wright, J.; Trayhurn, P.; Skene, D.J.; Robertson, M.D.; Johnston, J.D. Rhythmic diurnal gene expression in human adipose tissue from individuals who are lean, overweight, and type 2 diabetic. Diabetes 2011, 60, 1577–1581. [Google Scholar] [CrossRef] [PubMed]
- McFadden, E.; Jones, M.E.; Schoemaker, M.J.; Ashworth, A.; Swerdlow, A.J. The relationship between obesity and exposure to light at night: Cross-sectional analyses of over 100,000 women in the Breakthrough Generations Study. Am. J. Epidemiol. 2014, 180, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Nduhirabandi, F.; Huisamen, B.; Strijdom, H.; Blackhurst, D.; Lochner, A. Short-term melatonin consumption protects the heart of obese rats independent of body weight change and visceral adiposity. J. Pineal Res. 2014, 57, 317–332. [Google Scholar] [CrossRef] [PubMed]
- Stacchiotti, A.; Favero, G.; Giugno, L.; Lavazza, A.; Reiter, R.J.; Rodella, L.F.; Rezzani, R. Mitochondrial and metabolic dysfunction in renal convoluted tubules of obese mice: Protective role of melatonin. PLoS ONE 2014, 9, e111141. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Wang, X.; Chen, J.; Song, K.; Gusdon, A.M.; Li, L.; Bu, L.; Qu, S. Melatonin improves non-alcoholic fatty liver disease via MAPK-JNK/P38 signaling in high-fat-diet-induced obese mice. Lipids Health Dis. 2016, 15, 202. [Google Scholar] [CrossRef] [PubMed]
- Sagrillo-Fagundes, L.; Assuncao Salustiano, E.M.; Yen, P.W.; Soliman, A.; Vaillancourt, C. Melatonin in pregnancy: Effects on brain development and CNS programming disorders. Curr. Pharm. Des. 2016, 22, 978–986. [Google Scholar] [CrossRef] [PubMed]
- Ding, K.; Wang, H.; Xu, J.; Li, T.; Zhang, L.; Ding, Y.; Zhu, L.; He, J.; Zhou, M. Melatonin stimulates antioxidant enzymes and reduces oxidative stress in experimental traumatic brain injury: The Nrf2-ARE signaling pathway as a potential mechanism. Free Radic. Biol. Med. 2014, 73, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Jiang, S.; Dong, Y.; Fan, C.; Zhao, L.; Yang, X.; Li, J.; Di, S.; Yue, L.; Liang, G.; et al. Melatonin prevents cell death and mitochondrial dysfunction via a SIRT1-dependent mechanism during ischemic-stroke in mice. J. Pineal Res. 2015, 58, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Chen, G.; Li, J.; Qian, C.; Mo, H.; Gu, C.; Yan, F.; Yan, W.; Wang, L. Melatonin attenuates inflammatory response-induced brain edema in early brain injury following a subarachnoid hemorrhage: A possible role for the regulation of pro-inflammatory cytokines. J. Pineal Res. 2014, 57, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.J.; Lee, M.Y.; Chen, H.Y.; Hsu, Y.S.; Wu, T.S.; Chen, S.T.; Chang, G.L. Melatonin attenuates gray and white matter damage in a mouse model of transient focal cerebral ischemia. J. Pineal Res. 2005, 38, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Pazar, A.; Kolgazi, M.; Memisoglu, A.; Bahadir, E.; Sirvanci, S.; Yaman, A.; Yegen, B.C.; Ozek, E. The neuroprotective and anti-apoptotic effects of melatonin on hemolytic hyperbilirubinemia-induced oxidative brain damage. J. Pineal Res. 2016, 60, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Keskin, I.; Kaplan, S.; Kalkan, S.; Sutcu, M.; Ulkay, M.B.; Esener, O.B. Evaluation of neuroprotection by melatonin against adverse effects of prenatal exposure to a nonsteroidal anti-inflammatory drug during peripheral nerve development. Int. J. Dev. Neurosci. 2015, 41, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Aranda, M.L.; Fleitas, M.F.G.; De Laurentiis, A.; Sarmiento, M.I.K.; Chianelli, M.; Sande, P.H.; Dorfman, D.; Rosenstein, R.E. Neuroprotective effect of melatonin in experimental optic neuritis in rats. J. Pineal Res. 2016, 60, 360–372. [Google Scholar] [CrossRef] [PubMed]
- Willis, G.L. Parkinson’s disease as a neuroendocrine disorder of circadian function: Dopamine-melatonin imbalance and the visual system in the genesis and progression of the degenerative process. Rev. Neurosci. 2008, 19, 245–316. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wang, P.; Ren, L.; Hu, C.; Bi, J. Protective effect of melatonin on soluble Abeta1–42-induced memory impairment, astrogliosis, and synaptic dysfunction via the Musashi1/Notch1/Hes1 signaling pathway in the rat hippocampus. Alzheimers Res. Ther. 2016, 8, 40. [Google Scholar] [CrossRef] [PubMed]
- Breen, D.P.; Nombela, C.; Vuono, R.; Jones, P.S.; Fisher, K.; Burn, D.J.; Brooks, D.J.; Reddy, A.B.; Rowe, J.B.; Barker, R.A. Hypothalamic volume loss is associated with reduced melatonin output in Parkinson’s disease. Mov. Disord. 2016, 31, 1062–1066. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Ran, D.; Zhao, X.; Huang, Y.; Long, S.; Liang, F.; Guo, W.; Nucifora, F.C., Jr.; Gu, H.; Lu, X.; et al. Melatonin attenuates hLRRK2-induced sleep disturbances and synaptic dysfunction in a Drosophila model of Parkinson’s disease. Mol. Med. Rep. 2016, 13, 3936–3944. [Google Scholar] [CrossRef] [PubMed]
- Cardinali, D.P.; Vigo, D.E.; Olivar, N.; Vidal, M.F.; Furio, A.M.; Brusco, L.I. Therapeutic application of melatonin in mild cognitive impairment. Am. J. Neurodegener. Dis. 2012, 1, 280–291. [Google Scholar] [PubMed]
- Mukda, S.; Panmanee, J.; Boontem, P.; Govitrapong, P. Melatonin administration reverses the alteration of amyloid precursor protein-cleaving secretases expression in aged mouse hippocampus. Neurosci. Lett. 2016, 621, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Waseem, M.; Tabassum, H.; Parvez, S. Neuroprotective effects of melatonin as evidenced by abrogation of oxaliplatin induced behavioral alterations, mitochondrial dysfunction and neurotoxicity in rat brain. Mitochondrion 2016, 30, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Li, J.G.; Zhang, G.W.; Meng, Z.Z.; Wang, L.Z.; Liu, H.Y.; Liu, Q.; Buren, B. Neuroprotective effect of acute melatonin treatment on hippocampal neurons against irradiation by inhibition of caspase-3. Exp. Ther. Med. 2016, 11, 2385–2390. [Google Scholar] [CrossRef] [PubMed]
- Koc, G.E.; Kaplan, S.; Altun, G.; Gumus, H.; Deniz, O.G.; Aydin, I.; Onger, M.E.; Altunkaynak, Z. Neuroprotective effects of melatonin and omega-3 on hippocampal cells prenatally exposed to 900 MHz electromagnetic fields. Int. J. Radiat. Biol. 2016, 92, 590–595. [Google Scholar]
- Letra-Vilela, R.; Sanchez-Sanchez, A.M.; Rocha, A.M.; Martin, V.; Branco-Santos, J.; Puente-Moncada, N.; Santa-Marta, M.; Outeiro, T.F.; Antolin, I.; Rodriguez, C.; et al. Distinct roles of N-acetyl and 5-methoxy groups in the antiproliferative and neuroprotective effects of melatonin. Mol. Cell Endocrinol. 2016, 434, 238–249. [Google Scholar] [CrossRef] [PubMed]
- Acuna-Castroviejo, D.; Escames, G.; Venegas, C.; Diaz-Casado, M.E.; Lima-Cabello, E.; Lopez, L.C.; Rosales-Corral, S.; Tan, D.X.; Reiter, R.J. Extrapineal melatonin: Sources, regulation, and potential functions. Cell Mol. Life Sci. 2014, 71, 2997–3025. [Google Scholar] [CrossRef] [PubMed]
- Tresguerres, I.F.; Tamimi, F.; Eimar, H.; Barralet, J.E.; Prieto, S.; Torres, J.; Calvo-Guirado, J.L.; Tresguerres, J.A. Melatonin dietary supplement as an anti-aging therapy for age-related bone loss. Rejuvenation Res. 2014, 17, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Hibaoui, Y.; Reutenauer-Patte, J.; Patthey-Vuadens, O.; Ruegg, U.T.; Dorchies, O.M. Melatonin improves muscle function of the dystrophic mdx5Cv mouse, a model for Duchenne muscular dystrophy. J. Pineal Res. 2011, 51, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Shin, I.S.; Park, J.W.; Shin, N.R.; Jeon, C.M.; Kwon, O.K.; Lee, M.Y.; Kim, H.S.; Kim, J.C.; Oh, S.R.; Ahn, K.S. Melatonin inhibits MUC5AC production via suppression of MAPK signaling in human airway epithelial cells. J. Pineal Res. 2014, 56, 398–407. [Google Scholar] [CrossRef] [PubMed]
- Sehajpal, J.; Kaur, T.; Bhatti, R.; Singh, A.P. Role of progesterone in melatonin-mediated protection against acute kidney injury. J. Surg. Res. 2014, 191, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Elbe, H.; Vardi, N.; Esrefoglu, M.; Ates, B.; Yologlu, S.; Taskapan, C. Amelioration of streptozotocin-induced diabetic nephropathy by melatonin, quercetin, and resveratrol in rats. Hum. Exp. Toxicol. 2015, 34, 100–113. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.Z.; He, T.; Gao, J.X.; Liu, Y.; Liu, J.Q.; Han, S.C.; Li, Y.; Shi, J.H.; Han, J.T.; Tao, K.; et al. Melatonin prevents acute kidney injury in severely burned rats via the activation of SIRT1. Sci. Rep. 2016, 6, 32199. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Young, Y.K.; Fradette, J.; Eliopoulos, N. Melatonin pretreatment of human adipose tissue-derived mesenchymal stromal cells enhances their prosurvival and protective effects on human kidney cells. Am. J. Physiol. Ren. Physiol. 2015, 308, F1474–F1483. [Google Scholar] [CrossRef] [PubMed]
- Wierrani, F.; Grin, W.; Hlawka, B.; Kroiss, A.; Grunberger, W. Elevated serum melatonin levels during human late pregnancy and labour. J. Obstet. Gynaecol. 1997, 17, 449–451. [Google Scholar] [CrossRef] [PubMed]
- Takayama, H.; Nakamura, Y.; Tamura, H.; Yamagata, Y.; Harada, A.; Nakata, M.; Sugino, N.; Kato, H. Pineal gland (melatonin) affects the parturition time, but not luteal function and fetal growth, in pregnant rats. Endocr. J. 2003, 50, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Tola, E.N.; Mungan, M.T.; Uguz, A.C.; Naziroglu, M. Intracellular Ca2+ and antioxidant values induced positive effect on fertilisation ratio and oocyte quality of granulosa cells in patients undergoing in vitro fertilisation. Reprod. Fertil. Dev. 2013, 25, 746–752. [Google Scholar] [CrossRef] [PubMed]
- Bromfield, E.G.; Aitken, R.J.; Anderson, A.L.; McLaughlin, E.A.; Nixon, B. The impact of oxidative stress on chaperone-mediated human sperm-egg interaction. Hum. Reprod. 2015, 30, 2597–2613. [Google Scholar] [CrossRef] [PubMed]
- Kose, O.; Arabaci, T.; Kara, A.; Yemenoglu, H.; Kermen, E.; Kizildag, A.; Gedikli, S.; Ozkanlar, S. Effects of melatonin on oxidative stress index and Alveolar bone loss in diabetic rats with periodontitis. J. Periodontol. 2016, 87, e82–e90. [Google Scholar] [CrossRef] [PubMed]
- Arabaci, T.; Kermen, E.; Ozkanlar, S.; Kose, O.; Kara, A.; Kizildag, A.; Duman, S.B.; Ibisoglu, E. Therapeutic effects of melatonin on alveolar bone resorption after experimental periodontitis in rats: A biochemical and immunohistochemical study. J. Periodontol. 2015, 86, 874–881. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R. Melatonin and the theories of aging: A critical appraisal of melatonin’s role in antiaging mechanisms. J. Pineal Res. 2013, 55, 325–356. [Google Scholar] [CrossRef] [PubMed]
- Wahab, M.H.; Akoul, E.S.; Abdel-Aziz, A.A. Modulatory effects of melatonin and vitamin E on doxorubicin-induced cardiotoxicity in Ehrlich ascites carcinoma-bearing mice. Tumori 2000, 86, 157–162. [Google Scholar] [PubMed]
- Andersen, L.P.; Rosenberg, J.; Gogenur, I. Perioperative melatonin: Not ready for prime time. Br. J. Anaesth. 2014, 112, 7–8. [Google Scholar] [CrossRef] [PubMed]
- Seabra, M.L.; Bignotto, M.; Pinto, L.R., Jr.; Tufik, S. Randomized, double-blind clinical trial, controlled with placebo, of the toxicology of chronic melatonin treatment. J. Pineal Res. 2000, 29, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Jahnke, G.; Marr, M.; Myers, C.; Wilson, R.; Travlos, G.; Price, C. Maternal and developmental toxicity evaluation of melatonin administered orally to pregnant Sprague-Dawley rats. Toxicol. Sci. 1999, 50, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Wright, B.; Sims, D.; Smart, S.; Alwazeer, A.; Alderson-Day, B.; Allgar, V.; Whitton, C.; Tomlinson, H.; Bennett, S.; Jardine, J.; et al. Melatonin versus placebo in children with autism spectrum conditions and severe sleep problems not amenable to behaviour management strategies: A randomised controlled crossover trial. J. Autism Dev. Disord. 2011, 41, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Gringras, P.; Gamble, C.; Jones, A.P.; Wiggs, L.; Williamson, P.R.; Sutcliffe, A.; Montgomery, P.; Whitehouse, W.P.; Choonara, I.; Allport, T.; et al. Melatonin for sleep problems in children with neurodevelopmental disorders: Randomised double masked placebo controlled trial. BMJ 2012, 345, e6664. [Google Scholar] [CrossRef] [PubMed]
- Holliman, B.J.; Chyka, P.A. Problems in assessment of acute melatonin overdose. South. Med. J. 1997, 90, 451–453. [Google Scholar] [CrossRef] [PubMed]
- Gitto, E.; Aversa, S.; Salpietro, C.D.; Barberi, I.; Arrigo, T.; Trimarchi, G.; Reiter, R.J.; Pellegrino, S. Pain in neonatal intensive care: Role of melatonin as an analgesic antioxidant. J. Pineal Res. 2012, 52, 291–295. [Google Scholar] [CrossRef] [PubMed]
- Gitto, E.; Karbownik, M.; Reiter, R.J.; Tan, D.X.; Cuzzocrea, S.; Chiurazzi, P.; Cordaro, S.; Corona, G.; Trimarchi, G.; Barberi, I. Effects of melatonin treatment in septic newborns. Pediatr. Res. 2001, 50, 756–760. [Google Scholar] [CrossRef] [PubMed]
- Gitto, E.; Romeo, C.; Reiter, R.J.; Impellizzeri, P.; Pesce, S.; Basile, M.; Antonuccio, P.; Trimarchi, G.; Gentile, C.; Barberi, I.; et al. Melatonin reduces oxidative stress in surgical neonates. J. Pediatr. Surg. 2004, 39, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Gitto, E.; Reiter, R.J.; Amodio, A.; Romeo, C.; Cuzzocrea, E.; Sabatino, G.; Buonocore, G.; Cordaro, V.; Trimarchi, G.; Barberi, I. Early indicators of chronic lung disease in preterm infants with respiratory distress syndrome and their inhibition by melatonin. J. Pineal Res. 2004, 36, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Acil, M.; Basgul, E.; Celiker, V.; Karagoz, A.H.; Demir, B.; Aypar, U. Perioperative effects of melatonin and midazolam premedication on sedation, orientation, anxiety scores and psychomotor performance. Eur. J. Anaesthesiol. 2004, 21, 553–557. [Google Scholar] [CrossRef] [PubMed]
- Naguib, M.; Samarkandi, A.H. The comparative dose-response effects of melatonin and midazolam for premedication of adult patients: A double-blinded, placebo-controlled study. Anesth. Analg. 2000, 91, 473–479. [Google Scholar] [PubMed]
- Edmonds, K.E.; Stetson, M.H. Pineal gland and melatonin affect testicular status in the adult marsh rice rat (Oryzomys palustris). Gen. Comp. Endocrinol. 1995, 99, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Kennaway, D.J.; Rowe, S.A. Controlled-release melatonin implants delay puberty in rats without altering melatonin rhythmicity. J. Pineal Res. 1997, 22, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Kelestimur, H.; Ozcan, M.; Kacar, E.; Alcin, E.; Yilmaz, B.; Ayar, A. Melatonin elicits protein kinase C-mediated calcium response in immortalized GT1–7 GnRH neurons. Brain Res. 2012, 1435, 24–28. [Google Scholar] [CrossRef] [PubMed]
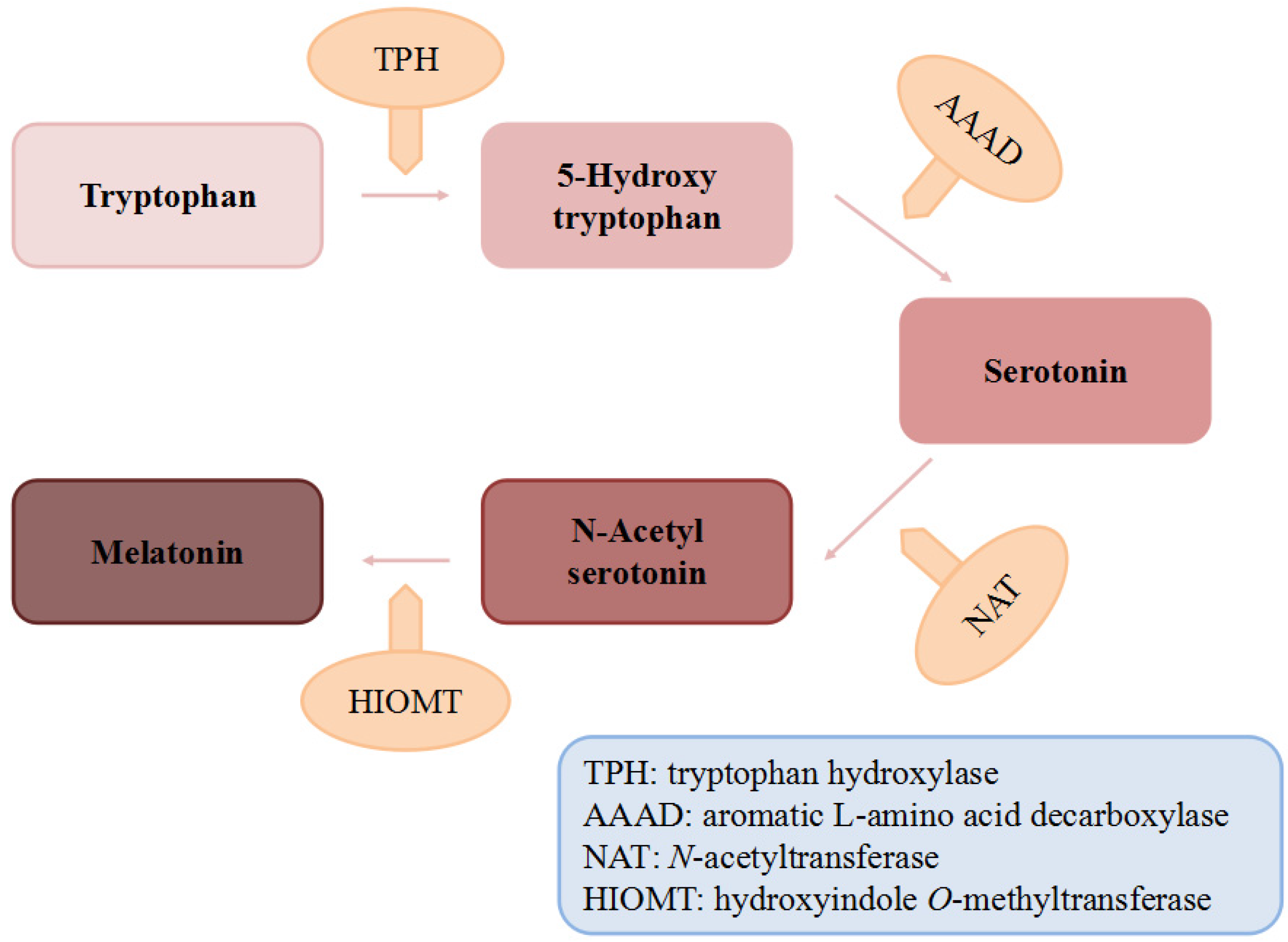
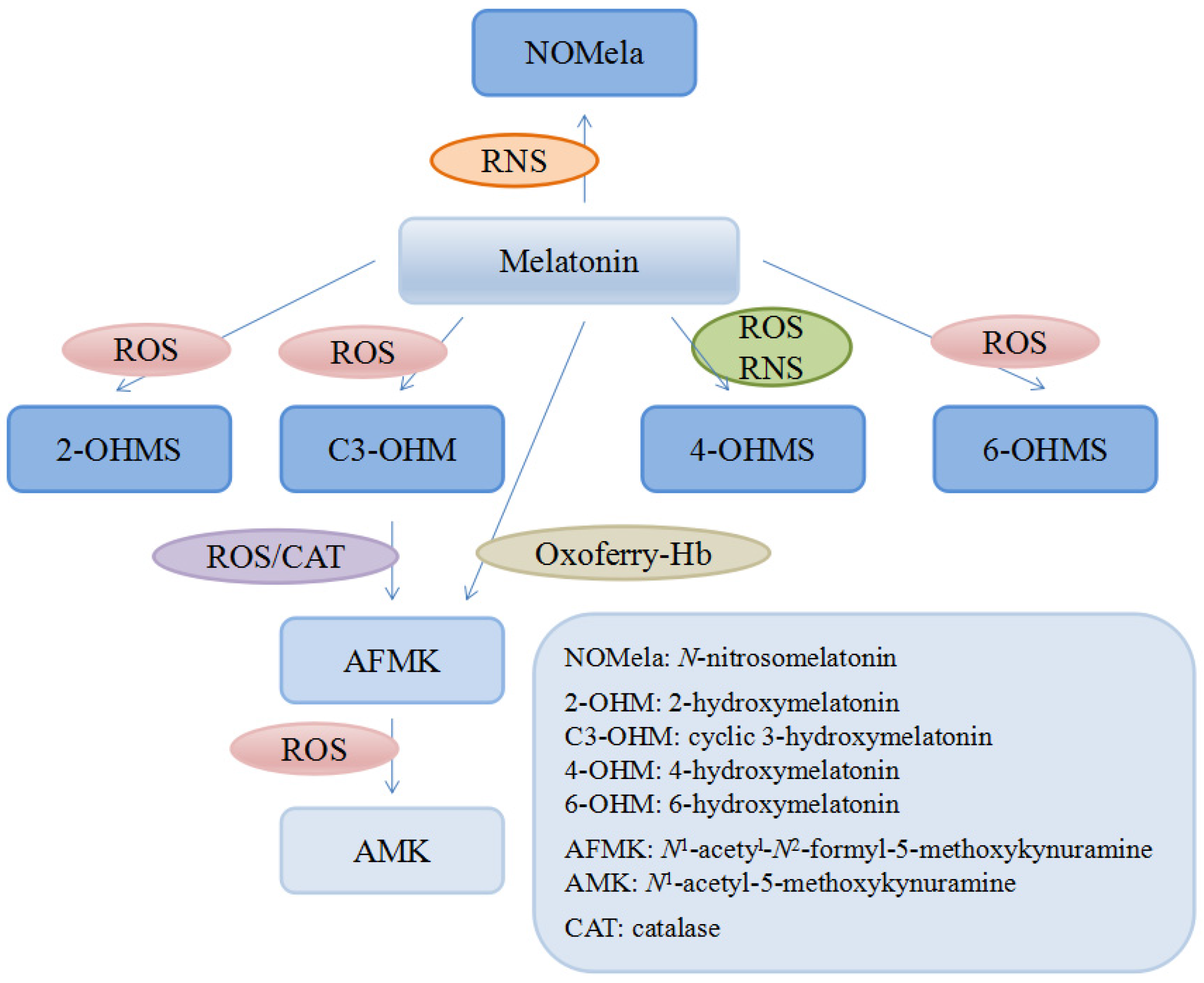
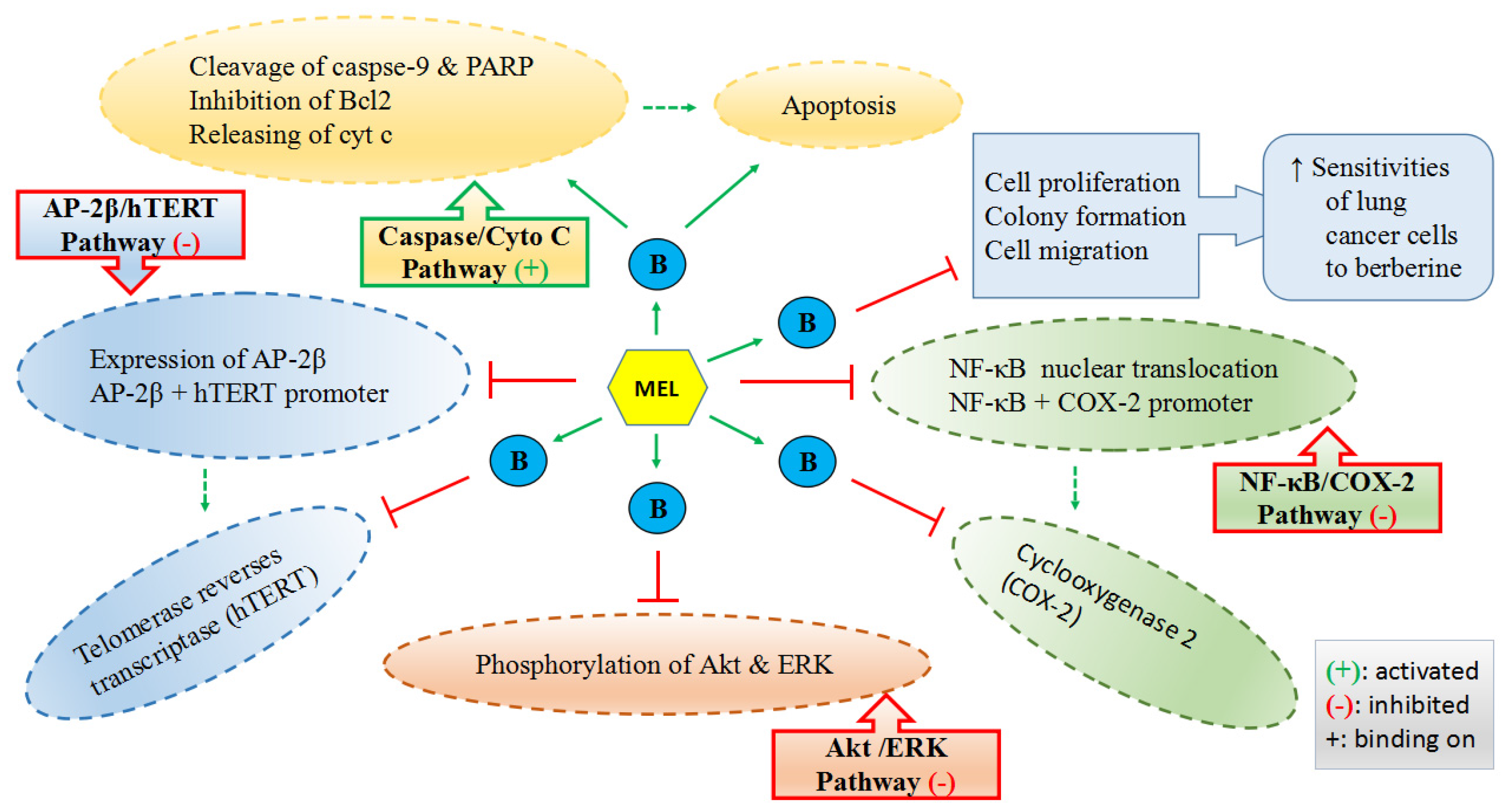

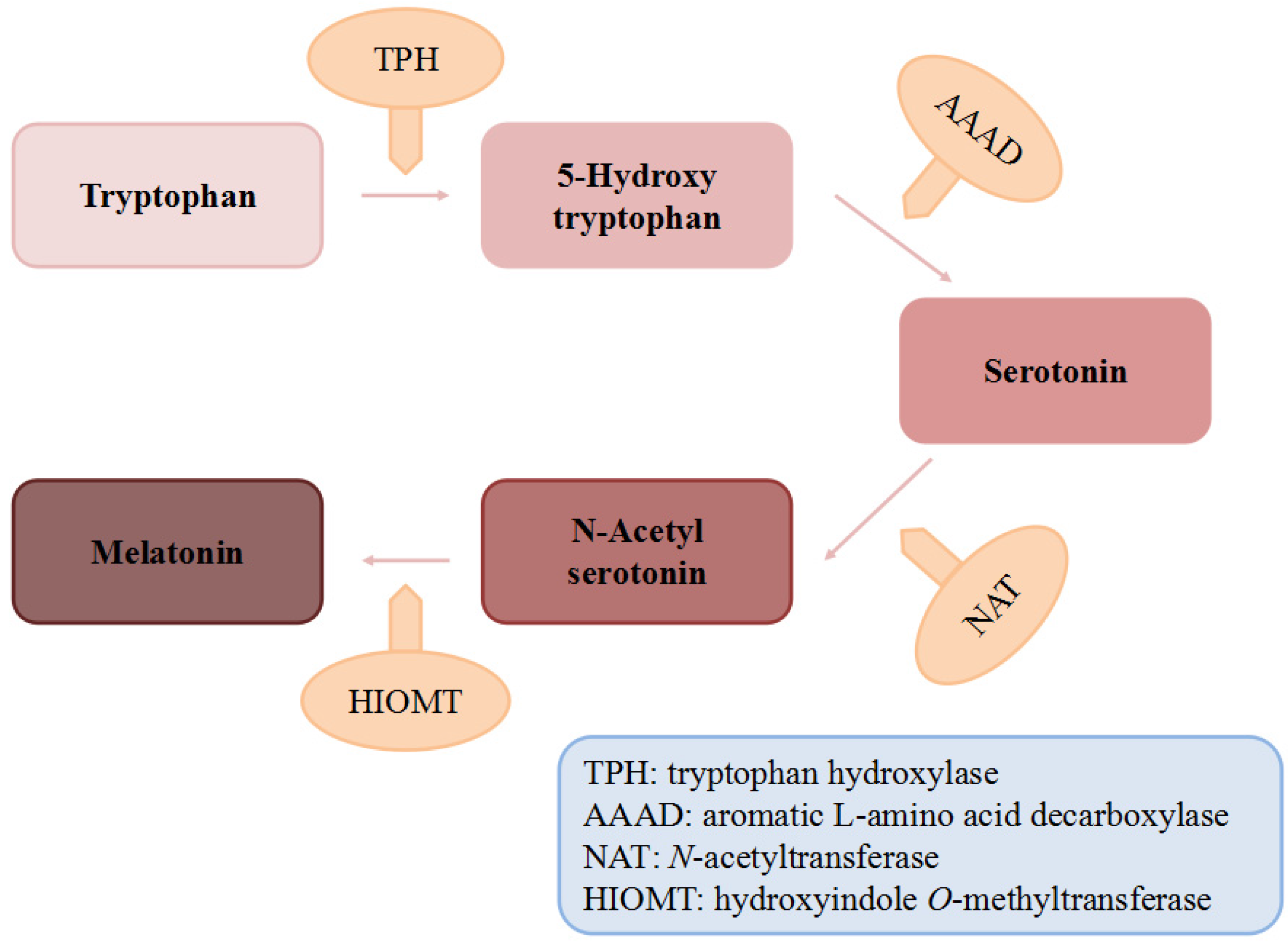{kind=link}
{kind=link}
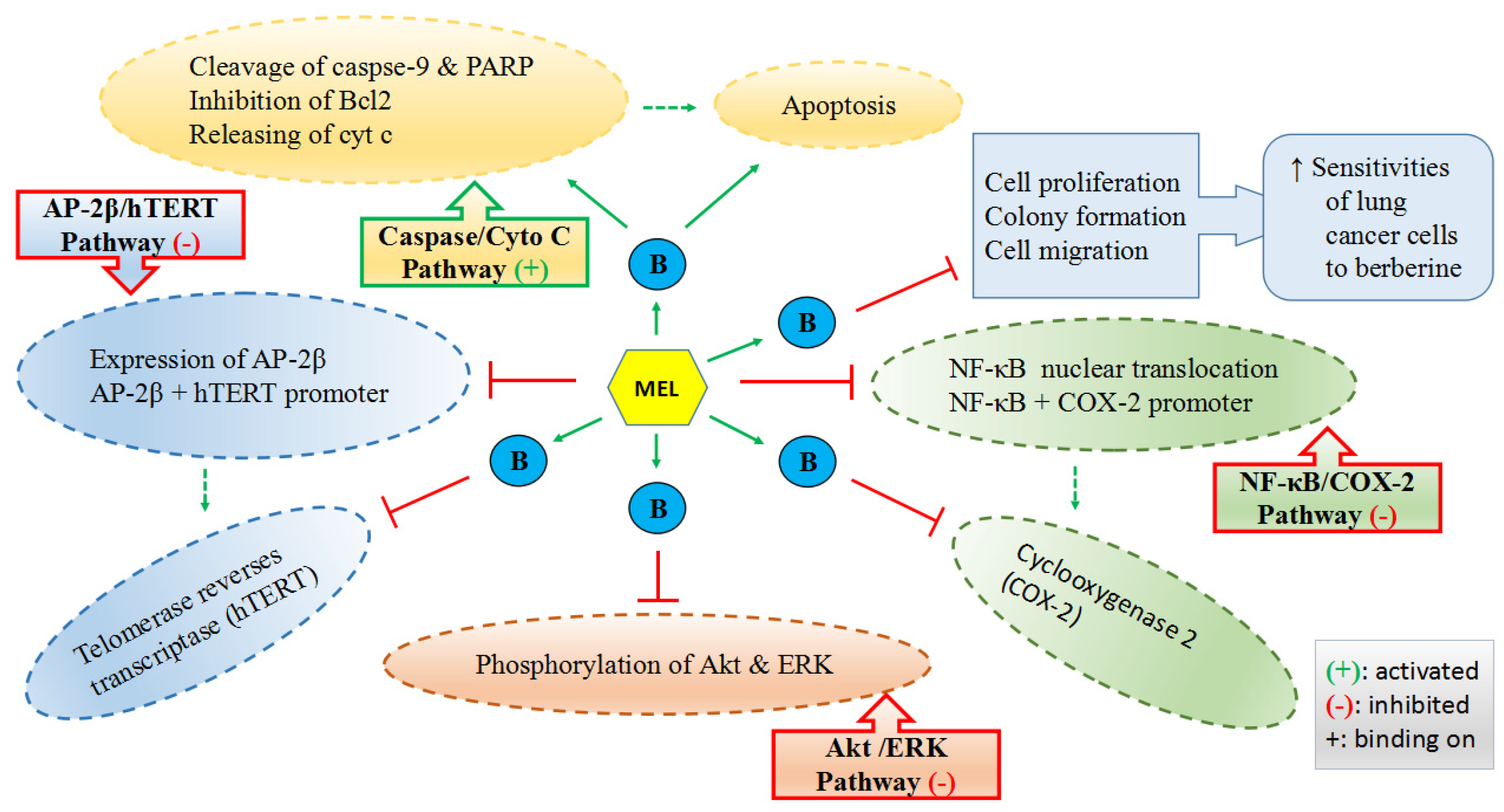{kind=link}
| Name | Scientific Name/Variety/Origin | MT Value or Range ng/g or pg/mL | No. of Samples | Quantified by | Reference |
|---|---|---|---|---|---|
| Animal Foods | |||||
| Meat | |||||
| Lamb | Not specified | 1.6 ± 0.14 ng/g | 5 | HPLC | [22] |
| Beef | Not specified | 2.1 ± 0.13 ng/g | 5 | HPLC | [22] |
| Pork | Not specified | 2.5 ± 0.18 ng/g | 5 | HPLC | [22] |
| Fish | |||||
| Salmon | Not specified | 3.7 ± 0.21 ng/g | 5 | HPLC | [22] |
| Chicken | |||||
| Meat and skin | Not specified | 2.3 ± 0.23 ng/g | 5 | HPLC | [22] |
| Liver and heart | Not specified | 1.1 ± 0.01 ng/g | 5 | HPLC | [22] |
| Egg | |||||
| Dried solids | Not specified | 6.1 ± 0.95 ng/g | 5 | HPLC | [22] |
| Raw, whole | Not specified | 1.54 ng/g | 5 | HPLC | [22] |
| Milk and dairy products | |||||
| Human milk | |||||
| Indian human milk | Not specified | 15.92 ± 1.02 pg/mL | 6 | LC-MS/MS | [37] |
| Breast milk | Not specified | 0–42 pg/mL | 5 | ELISA | [41] |
| Bovine milk | |||||
| Fresh colostrum, | Not specified | 0.06 ng/g | 5 | HPLC | [22] |
| Cow milk | Not specified | 14.45 ± 0.12 pg/mL | 6 | LC-MS/MS | [37] |
| Cow Milk | Holstein cows | 4.03–39.43 pg/mL | 3 | EIA | [38] |
| Colostrum powder | Not specified | 0.6 ± 0.06 ng/g | 5 | HPLC | [22] |
| Toned milk | Not specified | 18.41 ± 0.62 pg/mL | 6 | LC-MS/MS | [37] |
| Yoghurt | Not specified | 0.13 ± 0.01 ng/mL | 5 | LC-MS/MS | [42] |
| Artificial formulas | Not specified | nd | 15 | ELISA | [41] |
| Fermented milk drink | Kefir | nd | 5 | LC-MS/MS | [42] |
| Plant foods | |||||
| Cereals | |||||
| Corn | |||||
| Corn (whole, yellow) | Not specified | 1.3 ± 0.28 ng/g | 5 | HPLC | [22] |
| Corn (germ meal) | Not specified | 1.0 ± 0.10 ng/g | 5 | HPLC | [22] |
| Corn (YM001-) | 58 cultivars | 10–2034 ng/g DW | N/A | HPLC | [31] |
| Corn | Not specified | 1.88 ng/g FW | N/A | GC/MS | [47] |
| Sweet corn | Not specified | 1.37 ng/g FW | N/A | HPLC-FD | [83] |
| Rice | |||||
| Rice | Oryza sativum L. ssp. japonica | 1.50 ng/g FW | N/A | GC/MS | [47] |
| Rice (SD001-) | 25 cultivars | 0–264 ng/g DW | N/A | HPLC | [31] |
| Black glutinous | Long grain, waxy, Thailand (Bran) | 73.81 ± 0.07 ng/g DW | 9 | PLE HPLC-FD | [17] |
| Black | Not specified | 182.04 ± 1.62 ng/g DW | 9 | PLE HPLC-FD | [17] |
| Red | Not specified | 212.01 ± 1.37 ng/g DW | 9 | PLE HPLC-FD | [17] |
| Whole short grain | Not specified | 47.83 ± 0.12 ng/g DW | 9 | PLE HPLC-FD | [17] |
| Whole semi-long grain | Not specified | 42.95 ± 0.64 ng/g DW | 9 | PLE HPLC-FD | [17] |
| Polished short grain | Not specified | 31.99 ±0.31 ng/g DW | 9 | PLE HPLC-FD | [17] |
| Polished long grain | Not specified | 27.61 ± 1.16 ng/g DW | 9 | PLE HPLC-FD | [17] |
| Basmati | Not specified | 38.46 ± 0.07 ng/g DW | 9 | PLE HPLC-FD | [17] |
| Parboiled rice | Not specified | 28.33 ± 0.61 ng/g DW | 9 | PLE HPLC-FD | [17] |
| Rice | Oryza sativa cv. Dongjin | 0.04 ng/g DW | 20 | HPLC | [84] |
| Rice (transgenic) | Oryza sativa cv. Dongjin | 0.07–1.25 ng/g DW | 20 | HPLC | [84] |
| Rice | Oryza sativum L. ssp. japonica | 1.01 ng/g FW | N/A | HPLC-FD | [83] |
| Wheat | |||||
| Wheat | Triticum aestivum L. | 124.7 ± 14.9 ng/g FW | N/A | HPLC-ECD | [85] |
| Whole grain | Not specified | 2–4 ng/g | N/A | Not specified | [86] |
| Purple wheat | Not specified | 4 ng/g DW | 3 | HPLC-UV | [87] |
| Purple (heat stressed) | Not specified | 2 ng/g DW | 3 | HPLC-UV | [87] |
| Barley | |||||
| Barley | Hordeum vulgare L. | 0.87 ng/g FW | N/A | GC/MS | [47] |
| Barley | Hordeum vulgare L. | 82.3 ± 6.0 ng/g FW | N/A | HPLC-ECD | [85] |
| Barley | Hordeum vulgare L. | 0.38 ng/g FW | N/A | HPLC-FD | [83] |
| Oats | |||||
| Oats | Avena sativa L. | 90.6 ± 7.7 ng/g FW | N/A | HPLC-ECD | [85] |
| Oat | Avena sativa L. | 1.80 ng/g FW | N/A | HPLC-FD | [83] |
| Bread | |||||
| Crumb | Specific ingredients | 0.19–0.63 ng/g DW | 3 | LC-ESI-MS/MS | [20] |
| Crumb | Not specified | 0.34 ± 0.03 ng/g DW | 5 | LC-MS/MS | [42] |
| Crust | Specific ingredients | 0.14–0.82 ng/g DW | 3 | LC-ESI-MS/MS | [20] |
| Crust | Not specified | 0.14 ± 0.02 ng/g DW | 5 | LC-MS/MS | [42] |
| Fruits | |||||
| Pineapple | |||||
| Pineapple | Ananas comosus L. | 0.28 ng/g FW | N/A | GC/MS | [47] |
| Pineapple | Ananas comosus L. | 0.04 ng/g FW | N/A | HPLC-FD | [83] |
| Kiwi fruit | Actinidia chinensis L. | 0.02 ng/g FW | N/A | HPLC-FD | [83] |
| Strawberry | |||||
| Strawberry | Fragaria magna L. | 0.14 ng/g FW | N/A | GC/MS | [47] |
| Strawberry | Fragaria ananassa L. cv. Camarosa | 5.58 ± 0.01 ng/g FW | 3 | LC-MS/LC-FD | [44] |
| Strawberry | Fragaria ananassa L. cv. Candonga | 5.5 ± 0.6 ng/g FW | 3 | LC-MS/LC-FD | [44] |
| Strawberry | Fragaria ananassa L. cv. Festival | 11.26 ± 0.13 ng/g FW | 3 | LC-MS/LC-FD | [44] |
| Strawberry | Fragaria ananassa L. cv. Primoris | 8.5 ± 0.6 ng/g FW | 3 | LC-MS/LC-FD | [44] |
| Strawberry | Fragaria magna L. | 0.01 ng/g FW | N/A | HPLC-FD | [83] |
| Banana | Musa ensete | 0.66 ng/g FW | N/A | GC/MS | [47] |
| Apple | |||||
| Apple | Malus domestica | nd | N/A | HPLC-FD | [83] |
| Apple | Malus domestica Borkh. cv. Red Fuji | 5 ng/g FW | 3 | HPLC | [88] |
| Apple | Malus domestica | 0.16 ng/g FW | N/A | GC/MS | [47] |
| Pomegranata | Punica granatum | 0.17 ng/g FW | N/A | GC/MS | [47] |
| Mulberry | |||||
| Mulberry | Hongguo2 Morusnigra, black | 1.41 ng/g FW | 3 | HPLC-ESI-MS/MS | [36] |
| Mulberry | Baiyuwang Morus alba, white | 0.58 ng/g FW | 3 | HPLC-ESI-MS/MS | [36] |
| Cherry | |||||
| Tart cherries (frozen) | Prunus cerasus L. cv. Balaton | 2.9 ± 0.6 ng/g DW | 3 | HPLC-EMS | [60] |
| Tart cherries (dry) | Prunus cerasus L. cv. Balaton | nd | 3 | HPLC-EMS | [60] |
| Tart cherries | Prunus cerasus L. cv. Balaton | 13.46 ± 1.10 ng/g FW | 3 | HPLC-ECD | [46] |
| Tart cherries (frozen) | Prunus cerasus L. cv. Montmorency | 12.3 ± 2 ng/g DW | 3 | HPLC-EMS | [60] |
| Tart cherries (dry) | Prunus cerasus L. cv. Montmorency | nd | 3 | HPLC-EMS | [60] |
| Tart cherries | Prunus cerasus L. cv. Montmorency | 2.06 ± 0.17 ng/g FW | 3 | HPLC-ECD | [46] |
| Cherry | Prunus avium L. cv. Hongdeng | 10–20 ng/g FW | 3 | SPE HPLC | [89] |
| Cherry | Prunus avium L. cv. Rainier | 10–20 ng/g FW | 3 | SPE HPLC | [89] |
| Cherry | Prunus avium L. cv. Burlat | 0.22 ng/g FW | 3 | HPLC-MS | [30] |
| Cherry | Prunus avium L. cv. Navalinda | 0.03 ng/g FW | 3 | HPLC-MS | [30] |
| Cherry | Prunus avium L. cv. Van | 0.01 ng/g FW | 3 | HPLC-MS | [30] |
| Cherry | Prunus avium L. cv. Pico Limón Negro | 0.01 ng/g FW | 3 | HPLC-MS | [30] |
| Cherry | Prunus avium L. cv. Sweetheart | 0.06 ng/g FW | 3 | HPLC-MS | [30] |
| Cherry | Prunus avium L. cv. Pico Negro | 0.12 ng/g FW | 3 | HPLC-MS | [30] |
| Cherry | Prunus avium L. cv. Ambrunés | nd | 3 | HPLC-MS | [30] |
| Cherry | Prunus avium L. cv. Pico Colorado | 0.05 ng/g FW | 3 | HPLC-MS | [30] |
| Grape | |||||
| Grape | Albana, white | 1.2 ng/g | 3 | HPLC-FD | [43] |
| Grape (skin) | Vitis vinifera L. cv. Merlot | 9.3 ± 0.14 ng/g grapes | 3 | UPLC-MS/MS | [90] |
| Grape (fresh) | Vitis vinifera L. cv. Merlot | 3.9 ± 0.06 ng/g grapes | 3 | UPLC-MS/MS | [90] |
| Grape (skin) | Vitis vinifera L. cv Malbec | 8.9–158.9 ng/g DW | 3 | HPLC-ESI-MS/MS | [45] |
| Grape (skin) | Vitis vinifera L. cv. Nebbiolo | 0.97 ng/g | 3 | HPLC-ELISA | [35] |
| Grape (skin) | Vitis vinifera L. cv. Croatina, | 0.87 ng/g | 3 | HPLC-ELISA | [35] |
| Grape (skin) | Vitis vinifera L. cv. Barbera | 0.63 ng/g | 3 | HPLC-ELISA | [35] |
| Grape (skin) | Vitis vinifera L. cv. Cabernet Sauvignon | 0.42 ng/g | 3 | HPLC-ELISA | [35] |
| Grape (skin) | Vitis vinifera L. cv. Cabernet Franc | 0.01 ng/g | 3 | HPLC-ELISA | [35] |
| Grape (skin) | Vitis vinifera L. cv. Marzemino, | 0.03 ng/g | 3 | HPLC-ELISA | [35] |
| Grape (skin) | Vitis vinifera L. cv. Sangiovese | 0.33 ng/g | 3 | HPLC-ELISA | [35] |
| Grape (skin) | Vitis vinifera L. cv. Merlot, | 0.26 ng/g | 3 | HPLC-ELISA | [35] |
| Grape (skin) | Vitis vinifera L. cv. Malbec, | 1.2 ng/g | 5 | CEC | [56] |
| Grape (skin) | Vitis vinifera L. cv. Cabernet Sauvignon | 0.8 ng/g | 5 | CEC | [56] |
| Grape (skin) | Chardonnay | 0.6 ng/g | 5 | CEC | [56] |
| Cranberry | |||||
| Cranberry | Vaccinium oxycoccos L. | 40 ± 10 ug/g DW ** | 5 | UPLC-MS | [91] |
| Cranberry | Vaccinium vitis-idaea L. | 25 ± 3 ug/g DW ** | 5 | UPLC-MS | [91] |
| Cranberry | Vaccinium macrocarpon Ait. | 96 ± 26 ug/g DW ** | 5 | UPLC-MS | [91] |
| Vegetables | |||||
| Onion | Allium cepa L. | 0.30 ng/g FW | N/A | GC/MS | [47] |
| Onion | Allium fistulosum L., Welsh | 0.09 ng/g FW | N/A | HPLC-FD | [83] |
| Onion | Allium cepa L. | 0.03 ng/g FW | N/A | HPLC-FD | [83] |
| Garlic | Allium sativum L. | 0.59 ng/g FW | N/A | GC/MS | [47] |
| Cabbage | Brassica oleraceae L. var. capitata | 0.31 ng/g FW | N/A | GC/MS | [47] |
| Cauliflower | Brassica oleraceae L. var. botrytis | 0.82 ng/g FW | N/A | GC/MS | [47] |
| Turnip | Brassica rapa L. | 0.50 ng/g FW | N/A | GC/MS | [47] |
| Cucumber | Cucumis sativus L. | 0.59 ng/g FW | N/A | GC/MS | [47] |
| Cucumber | Cucumis sativus L. | 0.03 ng/g FW | N/A | HPLC-FD | [83] |
| Cucumber | Not specified | 0.01 ng/g | 1 | GC/MS | [48] |
| Carrot | Daucus carota L. | 0.49 ng/g FW | N/A | GC/MS | [47] |
| Carrot | Paucus carota L. | 0.06 ng/g FW | N/A | HPLC-FD | [83] |
| Radish | Raphnus sativus L. | 0.76 ng/g FW | N/A | GC/MS | [47] |
| Japanese radish | Bassica campestris L. | 0.66 ng/g FW | N/A | HPLC-FD | [83] |
| Potato | Solanum tuberosum L. | nd | N/A | GC/MS | [47] |
| Potato | Not specified | nd | 1 | GC/MS | [48] |
| Ginger | Zingiber officinale Rosc. | 1.42 ng/g FW | N/A | GC/MS | [47] |
| Black olive | Not specified | 0.01 ng/g DW | 5 | LC-MS/MS | [42] |
| Beetroot | Beta vulgaris | 0.002 ng/g | 1 | GC/MS | [48] |
| Purslane | Portulaca oleracea L. | 19 ng/g WW | N/A | GC/MS | [92] |
| Spinach | Not specified | 0.04 ng/g WW | N/A | GC/MS | [92] |
| Indian spinach | Basella alba L. | 0.04 ng/g FW | N/A | HPLC-FD | [83] |
| Asparagus | Asparagus officinalis L. | 0.01 ng/g FW | N/A | HPLC-FD | [83] |
| Pepper | Capsicum annuum L. cv. Barranca | 4.48 ng/g FW/31.01 ng/g DW | 4 | UHPLC-MS/MS | [49] |
| Pepper | Capsicum annuum L. cv. F26 | 11.9 ng/g FW/93.4 ng/g DW | 4 | UHPLC-MS/MS | [49] |
| Tomato | Solanum lycopersicum L. cv. Ciliegia | 0.64 ng/g FW/7.47 ng/g DW | 4 | UHPLC-MS/MS | [49] |
| Tomato | Solanum lycopersicum L. cv. Optima | 14.77 ng/g FW/249.98 ng/g DW | 4 | UHPLC-MS/MS | [49] |
| Tomato | Solanum lycopersicum L. cv. Micro-Tom | 1.5–66.6 ng/g FW | N/A | EIA | [93] |
| Tomato | Lycopersicon pimpinellifolium | 0.11 ng/g | 1 | GC/MS | [48] |
| Tomato | Lycopersicon esculentum Mill. cv. Sweet 100 | 0.51 ng/g | 1 | GC/MS | [48] |
| Tomato | Lycopersicon esculentum Mill. cv. Rutgers California Supreme | 0.17 ng/g | 1 | GC/MS | [48] |
| Tomatoes | Lycopersicon pimpinellifolium | 0.30 ng/g FW | N/A | GC/MS | [47] |
| Tomatoes | Lycopersicon esculentum cv. Bonda | 23.87 ± 2.02 ng/g FW | 3 | LC-MS/LC-FD | [44] |
| Tomatoes | Lycopersicon esculentum cv. Borsalinaa | 8.2 ± 0.6 ng/g FW | 3 | LC-MS/LC-FD | [44] |
| Tomatoes | Lycopersicon esculentum cv. Catalinaa | 4.1 ± 0.9 ng/g FW | 3 | LC-MS/LC-FD | [44] |
| Tomatoes | Lycopersicon esculentum cv. Gordala | 17.10 ± 1.21 ng/g FW | 3 | LC-MS/LC-FD | [44] |
| Tomatoes | Lycopersicon esculentum cv. Lucindaa | 4.45 ± 0.05 ng/g FW | 3 | LC-MS/LC-FD | [44] |
| Tomatoes | Lycopersicon esculentum cv. Marbonea | 18.13 ± 2.24 ng/g FW | 3 | LC-MS/LC-FD | [44] |
| Tomatoes | Lycopersicon esculentum cv. Myriadea | 8.0 ± 1.3 ng/g FW | 3 | LC-MS/LC-FD | [44] |
| Tomatoes | Lycopersicon esculentum cv. Pitenzaa | 14.0 ± 2.5 ng/g FW | 3 | LC-MS/LC-FD | [44] |
| Tomatoes | Lycopersicon esculentum cv. Santonioa | 7.73 ± 1.22 ng/g FW | 3 | LC-MS/LC-FD | [44] |
| Tomatoes (wild type) | Solanumlycopersicum L. cv. Micro-Tom | 6.58 ng/g FW | 3 | HPLC | [94] |
| Tomatoes (transgenic) | Solanumlycopersicum L. cv. Micro-Tom | 7.39–10.34 ng/g FW | 3 | HPLC | [94] |
| Tomatoes | Not specified | 0.03 ± 0.01 ng/g DW | 5 | LC-MS/MS | [42] |
| Tomato | Lycopersicon esculentum L. | 0.03 ng/g FW | N/A | HPLC-FD | [83] |
| Japanese butterbur | Patasites japonicus 50 | nd | N/A | HPLC-FD | [83] |
| Taro | Colocasis escutenta | 0.06 ng/g FW | N/A | HPLC-FD | [83] |
| Cabbage | Brassica oleracea | 0.11 ng/g FW | N/A | HPLC-FD | [83] |
| Chinese cabbage | Raphamus sativas | 0.11 ng/g FW | N/A | HPLC-FD | [83] |
| Chungitsu | Chrysanthemum cororarum | 0.42 ng/g FW | N/A | HPLC-FD | [83] |
| Ginger | Zinigiber officinale | 0.58 ng/g FW | N/A | HPLC-FD | [83] |
| Japanese ashitaba | Angelica keiskei | 0.62 ng/g FW | N/A | HPLC-FD | [83] |
| Mushrooms | |||||
| Mushroom | Agaricus bisporus* | 4300–6400 ng/g DW | 3 | RP-HPLC | [50] |
| Basidiomycota | Armillaria mellea | <10 ng/g DW | 3 | HPLC | [19] |
| Basidiomycota | Boletus badius | <10 ng/g DW | 3 | HPLC | [19] |
| Basidiomycota | Boletus edulis | 6800 ± 60 ng/g DW | 3 | HPLC | [19] |
| Basidiomycota | Cantharellus cibarius | 1400 ± 110 ng/g DW | 3 | HPLC | [19] |
| Basidiomycota | Lactarius deliciosus | 12,900 ± 770 ng/g DW | 3 | HPLC | [19] |
| Basidiomycota | Pleurotus ostreatus | <10 ng/g DW | 3 | HPLC | [19] |
| Legumes and seeds (raw) | |||||
| Legumes | |||||
| Lentils | Lens culinaris L. | 0.5 ng/g DW | 3 | HPLC-MS/MS | [53] |
| Kidney beans | Phaseolus vulgaris L | 1.0 ng/g DW | 3 | HPLC-MS/MS | [53] |
| Soybean | Glycine max | 0.45 ± 0.03 ng/g DW | N/A | RIA | [52] |
| Seeds | |||||
| Lupin (seed-cotyledons) | Lupinus albus L. | 3.83 ± 0.21 ng/g FW | 5 | HPLC-FD | [32] |
| Lupin (seed-coat) | Lupinus albus L. | 37.50 ± 2.3 ng/g FW | 5 | HPLC-FD | [32] |
| Lupin (seed-flour) | Lupinus albus L. | 0.53 ± 0.04 ng/g DW | 5 | HPLC-FD | [32] |
| Grape (seed) | Vitis vinifera L. cv. Merlot | 10.04 ± 0.49 ng/g grapes | 3 | UPLC-MS/MS | [90] |
| Barley (seed) | Hordeum vulgare L. | 0.58 ± 0.05 ng/g FW | 5 | HPLC-FD | [32] |
| Barley(seed-flour) | Hordeum vulgare L. | 0.09 ± 0.01 ng/g DW | 5 | HPLC-FD | [32] |
| Black mustard | Brassica nigra | 129 ng/g DW | 2 | HPLC-ECD | [51] |
| White mustard | Brassica hirta | 189 ng/g DW | 2 | HPLC-ECD | [51] |
| Fenugreek | Trigonella faena-graecum | 43 ng/g DW | 2 | HPLC-ECD | [51] |
| Milk thistle | Silybum marianum | 2 ng/g DW | 2 | HPLC-ECD | [51] |
| Celery | Apium gravolens | 7 ng/g DW | 2 | HPLC-ECD | [51] |
| Alfalfa | Medicago sativa | 16 ng/g DW | 2 | HPLC-ECD | [51] |
| Coriander | Coriandrum sativum | 7 ng/g DW | 2 | HPLC-ECD | [51] |
| Green cardamom | Elettaria cardamomum | 15 ng/g DW | 2 | HPLC-ECD | [51] |
| Fennel | Foeniculum vulgare | 28 ng/g DW | 2 | HPLC-ECD | [51] |
| Poppy | Popaver somniferum | 6 ng/g DW | 2 | HPLC-ECD | [51] |
| Anise | Pimpinella anisum | 7 ng/g DW | 2 | HPLC-ECD | [51] |
| Sunflower | Helianthus annuus | 29 ng/g DW | 2 | HPLC-ECD | [51] |
| Flax | Linum usitatissimum | 12 ng/g DW | 2 | HPLC-ECD | [51] |
| Almond | Prunus amydalus | 39 ng/g DW | 2 | HPLC-ECD | [51] |
| Chinese wolfberry | Lycium barbarum | 103 ng/g DW | 2 | HPLC-ECD | [51] |
| Cucumber | Cucumissativus L. cv. Jingyu-1 | 5.1 ng/g FW | 3 | UPLC-ESI-MS/MS | [95] |
| Alfalfa | Medicago sativa L. | 0.05 ± 0.00 ng/g DW | 3 | ELISA | [23] |
| Lentil | Lens sculenta L. | 0.07 ± 0.01 ng/g DW | 3 | ELISA | [23] |
| Mung bean | Vignaradiata L. | 0.01 ± 0.0 ng/g DW | 3 | ELISA | [23] |
| Onion | Allium cepa L. | 0.22 ± 0.01 ng/g DW | 3 | ELISA | [23] |
| Broccoli | Brassica oleraceae L. | 0.41 ± 0.04 ng/g DW | 3 | ELISA | [23] |
| Red cabbage | Brassica oleraceae capitate rubra L. | 0.34 ± 0.04 ng/g DW | 3 | ELISA | [23] |
| Radish (mixed) | Raphanus sativus japonicum L. | 0.28 ± 0.01 ng/g DW | 3 | ELISA | [23] |
| Raphanus sativus rambo L. | |||||
| Raphanus sativus sinicum rosae L. | |||||
| Legumes and seeds (germination) | |||||
| Legumes sprouts | |||||
| Lentils | Lens culinaris L. | 1089.8 ng/g DW | 3 | HPLC-MS/MS | [53] |
| Lentil | Not specified | 0.92 ± 0.06 ng/g DW | N/A | RIA | [52] |
| Kidney beans | Phaseolus vulgaris L. | 529.1 ng/g DW | 3 | HPLC-MS/MS | [53] |
| Soya bean | Glycine max L. | 1.89 ± 0.11 ng/g DW | N/A | RIA | [52] |
| Vetch | Vicia sativa L. | 1.91 ± 0.11 ng/g DW | N/A | RIA | [52] |
| Seedling | |||||
| Rice | Oryza sativa cv. Dongjin | 1.9 ng/g DW | 3 | HPLC | [96] |
| Rice (transgenic) | Oryza sativa cv. Dongjin | 2.7–5.2 ng/g DW | 3 | HPLC | [96] |
| Cucumber | Cucumis sativus L. cv. Jingyu-1 | 17.3 ng/g FW | 3 | UHPLC-ESI-MS/MS | [95] |
| Alfalfa | Medicago sativa L. | 0.13 ± 0.01 ng/g DW | 3 | ELISA | [23] |
| Lentil | Lens sculenta L. | 0.22 ± 0.0 1 ng/g DW | 3 | ELISA | [23] |
| Mung bean | Vignaradiata L. | 0.17 ± 0.01 ng/g DW | 3 | ELISA | [23] |
| Onion | Allium cepa L. | 0.30 ± 0.02 ng/g DW | 3 | ELISA | [23] |
| Broccoli | Brassica oleraceae L. | 0.44 ± 0.01 ng/g DW | 3 | ELISA | [23] |
| Red cabbage | Brassica oleraceae capitate rubra L. | 0.86 ± 0.05 ng/g DW | 3 | ELISA | [23] |
| Radish | Raphanus sativus japonicum L. | 0.54 ± 0.04 ng/g DW | 3 | ELISA | [23] |
| Raphanus sativus rambo L. | |||||
| Raphanus sativus sinicum rosae L. | |||||
| Nuts | |||||
| Pistachio | |||||
| Pistachio | Pistacia vera L. cv. Ahmad Aghaei | 233,000 ng/g DW | N/A | GC/MS | [28] |
| Pistachio | Pistacia vera L. cv. Akbari, Kalle | 226,900 ng/g DW | N/A | GC/MS | [28] |
| Pistachio | Pistacia vera L. cv. Qouchi | 231,400 ng/g DW | N/A | GC/MS | [28] |
| Pistachio | Pistacia vera L. cv. Fandoghi | 228,400 ng/g DW | N/A | GC/MS | [28] |
| Walnuts | |||||
| Walnuts | Juglans regia L. cv. Serr | 1.02 ± 0.06 ng/g FW | 4 | HPLC–MS | [97] |
| Walnuts | Juglans regia L. cv. Hartley | 1.77 ± 0.14 ng/g FW | 4 | HPLC–MS | [97] |
| Walnuts | Juglans regia L. cv. Chandler | 1.37 ± 0.37 ng/g FW | 4 | HPLC–MS | [97] |
| Walnuts | Juglans regia L. cv. Howard | 1.9 ± 0.4 ng/g FW | 4 | HPLC–MS | [97] |
| Walnuts | Not specified | 0.14 ± 0.03 ng/g DW | 5 | LC–MS/MS | [42] |
| Walnuts | Juglans regia L. | 3.5 ± 1.0 ng/g | 5 | HPLC-ECD | [78] |
| Juices and beverages | |||||
| Beer | Not specified | 0.09 ± 0.01 ng/mL | 5 | LC-MS/MS | [42] |
| Wine | |||||
| Albana must | Albana, Romagna | 1.1 ng/mL | 3 | HPLC-FD | [43] |
| Albana wine | Albana, Romagna | 0.6 ng/mL | 3 | HPLC-FD | [43] |
| Albana grappa | Albana, Romagna | 0.3 ng/mL | 3 | HPLC-FD | [43] |
| Red wine | Not specified | 0.26 ± 0.18 ng/mL | 3 | UHPLCMS/MS | [59] |
| Dessert ice wine | Not specified | 0.17 ± 0.11 ng/mL | 3 | UHPLCMS/MS | [59] |
| Groppello wines | Vitis vinifera L. cv. Groppello Gentile | 5.2 ng/mL | 3 | UHPLCMS/MS | [59] |
| Merlot wines | Merlot | 8.1 ng/mL | 3 | UHPLCMS/MS | [59] |
| Wine | Cabernet Sauvignon | 14.2 ± 0.2 ng/mL | 3 | HPLC-MS/MS | [55] |
| Wine | Cabernet Sauvignon | 0.23 ± 0.01 ng/mL | 3 | ELISA | [55] |
| Wine | Jaen Tinto | nd | 3 | HPLC-MS/MS | [55] |
| Wine | Jaen Tinto | 0.16 ± 0.01 ng/mL | 3 | ELISA | [55] |
| Wine | Vitis vinifera L. cv. Merlot | nd | 3 | HPLC-MS/MS | [55] |
| Wine | Vitis vinifera L. cv. Merlot | 0.21 ± 0.02 ng/mL | 3 | ELISA | [55] |
| Wine | Palomino Negro | nd | 3 | HPLC-MS/MS | [55] |
| Wine | Palomino Negro | 0.28 ± 0.00 ng/mL | 3 | ELISA | [55] |
| Wine | Petit Verdot | 5.1 ± 0.6 ng/mL | 3 | HPLC-MS/MS | [55] |
| Wine | Petit Verdot | 0.22 ± 0.01 ng/mL | 3 | ELISA | [55] |
| Wine | Prieto Picudo | 49.0 ± 4.7 ng/mL | 3 | HPLC-MS/MS | [55] |
| Wine | Prieto Picudo | 0.19 ± 0.01 ng/mL | 3 | ELISA | [55] |
| Wine | Syrah | 86.5 ± 2.6 ng/mL | 3 | HPLC-MS/MS | [55] |
| Wine | Syrah | 0.22 ± 0.02 ng/mL | 3 | ELISA | [55] |
| Wine | Tempranillo | 129.5 ± 3.5 ng/mL | 3 | HPLC-MS/MS | [55] |
| Wine | Tempranillo | 0.14 ± 0.01 ng/mL | 3 | ELISA | [55] |
| Wine | Vitis vinifera L. cv. Malbec | 0.24 ng/mL | 5 | CEC | [56] |
| Wine | Cabernet Sauvignon | 0.32 ng/mL | 5 | CEC | [56] |
| Wine | Chardonnay | 0.16 ng/mL | 5 | CEC | [56] |
| Wine | Sangiovese, red | 0.5 ng/mL | 3 | HPLC-FD | [43] |
| Wine | Trebbiano, white | 0.4 ng/mL | 3 | HPLC-FD | [43] |
| Coffee beans | |||||
| Green coffee | Not specified | 0.040 ± 0.01 ng/g DW | 5 | LC-MS/MS | [42] |
| Green beans | Coffea canephora L. (robusta) | 5800 ± 800 ng/g DW | 3 | LC-MS-ESI | [58] |
| Green beans | Coffea arabica L. (arabica) | 6800 ± 400 ng/g DW | 3 | LC-MS-ESI | [58] |
| Roasted beans | Coffea canephora L. (robusta) | 8000 ±.900 ng/g DW | 3 | LC-MS-ESI | [58] |
| Roasted beans | Coffea arabica L. (arabica) | 9600 ± 800 ng/g DW | 3 | LC-MS-ESI | [58] |
| Decoction (Brew) | Coffea canephora L. (robusta) | 60 ± 12 ng/mL | 3 | LC-MS-ESI | [58] |
| Decoction (Brew) | Coffea arabica L. (arabica) | 78 ± 5 ng/mL | 3 | LC-MS-ESI | [58] |
| Juices | |||||
| Orange juice | Citrus sinensis L. var. Navel late (Huelva, Spain) | 3.15–21.80 ng/mL | 3 | UHPLC-QqQ-MS/MS | [98] |
| Grape juice | Not specified | 0.5 ng/mL | 3 | HPLC-FD | [43] |
| Cacao powder | Not specified | 0.01 ng/g DW | 5 | LC-MS/MS | [42] |
| Concentrate | |||||
| Tart cherries | Prunus cerasus L. cv. Balaton | nd | 3 | HPLC-EMS | [46] |
| Tart cherries | Prunus cerasus L. cv. Montmorency | nd | 3 | HPLC-EMS | [46] |
| Sour cherries | Not specified | nd | 5 | LC-MS/MS | [42] |
| Tea | |||||
| Green tea | Not specified | nd | 5 | LC-MS/MS | [42] |
| Black tea | Not specified | nd | 5 | LC-MS/MS | [42] |
| Balsamic vinegars | 0.12 ± 0.014 ng/mL | 3 | UHPLCMS/MS | [59] | |
| Medical Herbs | |||||
| Huang-qin | Scutellaria biacalensis | 7110 ng/g DW | 2 | Not specified | [29] |
| St John’s Wort (flowers) | Hypericum perforatum | 4490 ng/g DW | 2 | Not specified | [29] |
| Chantui | Periostracum cicadae | 3771 ng/g DW | 3 | SPE-HPLC-FD | [61] |
| Gouteng | Uncaria rhynchophylla | 2460 ng/g DW | 3 | SPE-HPLC-FD | [61] |
| Diding | Viola philipica Cav. | 2368 ng/g DW | 3 | SPE-HPLC-FD | [61] |
| Shiya tea-leaf | Babreum coscluea | 2120 ng/g DW | 3 | SPE-HPLC-FD | [61] |
| Feverfew (fresh leaves) | Tanacetum parthenium | 1920–2450 ng/g DW | 2 | Not specified | [29] |
| St John’s Wort (leaves) | Hypericum perforatum | 1750 ng/g DW | 2 | Not specified | [29] |
| Sangye | Morus alba L. (Leaf) | 1510 ng/g DW | 3 | SPE-HPLC-FD | [61] |
| Huangbo | Phellodendron amurense Rupr. | 1235 ng/g DW | 3 | SPE-HPLC-FD | [61] |
| Sangbaipi | Mori Albae (Cortex) | 1110 ng/g DW | 3 | SPE-HPLC-FD | [61] |
| Yinyanghuo | Epimedium brevicornum Maxim | 1105 ng/g DW | 3 | SPE-HPLC-FD | [61] |
| Black pepper | Piper nigrum L. | 1092.7 ng/g DW | 5 | SPE HPLC ELISA | [62] |
| Huanglian | Coptis chinensis Franch | 1008 ng/g DW | 3 | SPE-HPLC-FD | [61] |
| Mulberry leaves | Morus spp. cv. Buriram 60 | 279.6 ng/g DW | 3 | HPLC-FD | [99] |
| Morus spp. cv. Sakonnakhon | 100.5 ng/g DW | 3 | HPLC-FD | [99] | |
| Morus spp. cv. Khunphai | 40.7 ng/g DW | 3 | HPLC-FD | [99] | |
| Edible oil | |||||
| Virgin Argan oil | Not specified | 0.06 ± 0.05 ng/g | 2 | HPLC-FD | [64] |
| Refined sunflower | Not specified | 0.03–0.08 ng/g | 2 | HPLC-FD | [64] |
| Primrose, | Not specified | 0.03–0.08 ng/g | 2 | HPLC-FD | [64] |
| Refined grape seed | Not specified | 0.03–0.08 ng/g | 2 | HPLC-FD | [64] |
| Refined walnut | Not specified | 0.03–0.08 ng/g | 2 | HPLC-FD | [64] |
| Virgin walnut | Not specified | 0.03–0.08 ng/g | 2 | HPLC-FD | [64] |
| Virgin linseed | Not specified | 0.03–0.08 ng/g | 2 | HPLC-FD | [64] |
| Linseed oils | Not specified | 0.03–0.08 ng/g | 2 | HPLC-FD | [64] |
| Refined linseed | Not specified | 0.29 ± 0.00 ng/g | 2 | HPLC-FD | [64] |
| Virgin sesame | Not specified | 0.03–0.08 ng/g | 2 | HPLC-FD | [64] |
| Wheat germ | Not specified | 0.03–0.08 ng/g | 2 | HPLC-FD | [64] |
| Virgin soybean | Not specified | 0.19 ± 0.00 ng/g | 2 | HPLC-FD | [64] |
| Olive oil | Extra virgin | 0.03 ± 0.00 ng/g | 2 | HPLC-FD | [64] |
| D.O. Sierra Ma′gina | Not specified | 0.11 ± 0.04 ng/mL | 3 | ELISA | [63] |
| D.O. Siurana | Not specified | 0.10 ± 0.02 ng/mL | 3 | ELISA | [63] |
| D.O. Bajo Arago′n | Not specified | 0.07 ± 0.02 ng/mL | 3 | ELISA | [63] |
| D.O. Montes de Toledo | Not specified | 0.11 ± 0.01 ng/mL | 3 | ELISA | [63] |
| D.O. Baena | Not specified | 0.12 ± 0.00 ng/mL | 3 | ELISA | [63] |
| D.O. Sierra de Segura | Not specified | 0.09 ± 0.00 ng/mL | 3 | ELISA | [63] |
| D.O. Les Garrigues | Not specified | 0.10 ± 0.00 ng/mL | 3 | ELISA | [63] |
| D.O. Toscano | Not specified | 0.11 ± 0.02 ng/mL | 3 | ELISA | [63] |
| Refined olive oil sample 1 | Not specified | 0.05 ± 0.01 ng/mL | 3 | ELISA | [63] |
| Refined olive oil sample 2 | Not specified | 0.08 ± 0.01 ng/mL | 3 | ELISA | [63] |
| Refined sunflower oil sample | Not specified | 0.05 ± 0.01 ng/mL | 3 | ELISA | [63] |
| Microorganisms | |||||
| Yeast (dried brewer) | Saccharomyces cerevisae | 2.2 ± 0.14 ng/g | 5 | HPLC | [22] |
| Study Type | Subjects | Dose (mg/kg b.w.)/(Dose Dependent) | Potential Mechanisms (Melatonin (Mel) and/or Its Metabolites) | Reference |
|---|---|---|---|---|
| Antioxidant Activities | ||||
| Directly scavenging free radical | ||||
| In vivo | Mouse | 10 mg/kg b.w. | Increasing the efficiency of electron transport chain: - lowering electron leakage and reducing free radical generation | [122] |
| In vitro | Human umbilical artery segment | 10−6, 10−5, 10−4 M (dose dependent) | Significantly scavenging the hydroxyl radical | [127] |
| Cascade effects: removing free radicals efficiently than other reductants: | ||||
| In vivo | Rat | 43 μmol/kg b.w. | - more efficient than Vitamin C | [128] |
| Rat | 2 μmol/kg b.w. | - more efficient than Vitamin E | [125] | |
| Mouse | 5 mg/kg b.w. | - more efficient than Vitamin E | [325] | |
| In vitro | Incubation medium | 1–1000 mM | - more efficient than Vitamin C & Vitamin E | [123] |
| Some metabolites more potent than its precursor in reducing oxidative stress: | ||||
| Fenton reaction-based assay | AFMK: 0.017–0.067 mM AMK: up to 0.2 mM | - the order of efficacy of scavenging ∙OH: AMK > AK > AFMK | [130] | |
| N/A | N/A | N/A | - C3-OHM is 2–3 fold more potent than Mel in reducing hypervalent hemoglobin (Tan & Reiter, unpublished observations). | [131] |
| Modulating and activating other enzymes | ||||
| Downregulating pro-oxidative enzymes | ||||
| In vitro & in vivo | Rat striatum | M & AMK: 10−11–10−3 M (dose dependent in vitro) | - Mel & AMK inhibiting nNOS activity - AMK more potent in inhibiting nNOS activity than Mel (in vivo) | [142] |
| In vitro | MCF-7 cells | 1 nM | Inhibiting the mRNA expression of COX 1 and COX-2 in MCF-7 cells | [143] |
| Stimulating the synthesis of other antioxidants | ||||
| In vitro | ECV304 cells | 1 μM | - Inducing γ-GCS expression to promote GSH synthesis | [124] |
| In vitro | 2 neuronal cell lines: PC12 cells & SK-N-SH | 1 nM | - Regulating AOEs gene expression - Increasing mRNA of SODs and GPx | [144] |
| Preventing antioxidant enzymes from oxidative stress | ||||
| In vitro | human BM-MSCs | 0 to 1000 μM (dose dependent 10–100 μM) | - significantly restoring SOD (p < 0.05) and CAT (p < 0.01) - increasing GSH (p < 0.01) with pre-treatment of Mel | [145] |
| In vivo | Sprague-Dawley rats | 10 mg/kg b.w. | GSH-Rd activity was completely or partially restored by Mel treatment | [146] |
| In vitro & in vivo | Sprague–Dawley rats | 0–0.1 mM (dose dependent in vitro) 10 mg/kg b.w. | G6PG activity dose-dependent in vitro (increased below 0.08 mM Mel concentration and reached a plateau above 0.1 mM) G6PG activity time-dependent (in vivo) | [147] |
| Synergistically working with other reductants | ||||
| Combining with other antioxidants to remove radicals synergistically | ||||
| In vitro | Rat liver homogenates | 2.5–1600 μM | - dramatically enhancing the protective effects after combining | [148] |
| Anti-inflammatory Activities | ||||
| NF-κB signaling pathway involved mechanisms | ||||
| Modulating NF-κB and its downstream pro-inflammatory target genes | ||||
| In vitro | Human colon cancer cell lines SW620 and LOVO | 1 mmol/L | - iNOS | [70] |
| In vitro | RAW 264.7 macrophages | 0.5, 1, 2 mM (dose dependent) | - COX-2, PGE2 | [158] |
| In vitro | Human neuroblastoma dopamine SH-SY5Y cell lines | 1, 10, 100 or 1000 nM | - TNF-α | [159] |
| In vitro | Rat astrocytoma C6 cells | 50–200 μM (dose dependent) | - GFAP | [160] |
| In vitro & in vivo | CHON-001 human chondrocyte cell line Rabbit with osteoarthritis (OA) | 0.1, 1, 10, 100 ng (dose- and time-dependent) 20 mg/kg | Protecting cells by blocking the activated NF-κB as well as the phosphorylation of PI3K/Akt, p38, ERK, JNK and MAPK | [161] |
| In vitro | BV2 murine microglial cell line | 1 mM | Downregulating chemokine expression | [162] |
| In vivo | Rats | 5 mg/kg | Inhibiting the inflammatory reaction | [163] |
| In vivo & in vitro | Female BALB/c mice MMECs | 5, 10, 20 mg/kg 25, 50, 100 μM (dose dependent) | Suppressing NF-κB activation and activating PPAR-γ | [164] |
| In vitro | Mast cells (RBL-2H3) | 100 nM and 1 mM (dose dependent) | Inhibiting IKK/NF-κB signal transduction | [165] |
| SIRT1 pathway involved mechanisms | ||||
| In vitro & in vivo | BV2 cell lysates PND7 rat brain | 100 µM 10 mg/kg | Activating SIRT1/Nrf2 signaling pathway to reduce oxidative stress damage | [168] |
| Other possible mechanisms | ||||
| In vivo | Pediatric patients | 10 mg (09:00 h) 60 mg ( 21:00 h ) | Regulating the expression of other pro-inflammatory genes | [150] |
| In vitro | Mouse Gsk3b knockout (Gsk3b−/−) and wild-type (Gsk3b+/+) MEF cells | 10 nM | Inhibiting the expression of inflammatory chemokines/cytokines | [169] |
| In vivo | Plasmodium | 10 µM (time dependent) | Inducing temporal up-regulation of gene expression related to UPS | [170] |
| In vivo | C57BL mice | 10 mg/kg i.p. | Downregulating mRNA of E2F2 and H2-Ab1 | [171] |
| In vivo | Rats | 5, 15, and 25 mg/kg (dose-dependent) | Activating the expression of NDRG2, which was involved in cellular differentiation, development, anti-apoptosis, anti-inflammatory cytokine, and antioxidant | [172] |
| In vivo | Carp | 10−4–10−12 M | Maintaining the pro- and anti- inflammatory balance during infection by influencing leukocyte migration and apoptosis | [151] |
| Enhancing Immune Activities | ||||
| Reciprocally regulating the nervous, endocrine, and immune systems | ||||
| In vivo & In vitro | Mice Thymus and spleen cells | 4–5 mL/day/mouse 1.5 pg/ml to 1.5 pgg/ml | Regulating thymocyte apoptosis | [174] |
| In vivo | Mice | 1.5 pg/mL to 1.5 pg/mL | The concentration of melatonin correspond with the change of seasons | [175] |
| Inhibiting the production of cAMP, cGMP and DAG, and improving the immunity | ||||
| In vitro | Human blood lymphocyte | N/A (dose-dependent) | Inhibiting adenylyl cyclase and the stimulating phospholipase C | [183] |
| In vivo | Golden hamsters | 25 μg/100 g/hamster/day | Improving immune responses | [184] |
| Protecting the immune organs, tissues and cells | ||||
| Reversing the weight loss of thymuses and spleens in pinealectomized animals | ||||
| In vivo & In vitro | Mice Thymus and spleen cells | 4–5 mL/day/mouse 1.5 pg/mL to 1.5 pg/mL | - thymus | [175] |
| In vivo | Syrian hamsters | 25 μg | - spleen | [188] |
| In vivo | Pediatric patients | N/A | Increasing tonsillar size | [189] |
| Improving proliferation, increasing activity and inhibiting apoptosis of immune cells | ||||
| In vitro | cultured monocytes | N/A | - monocyte | [190] |
| In vivo | ICR mice | 10 or 50 mg/kg | - natural killer (NK) cells | [191] |
| In vitro | Neutrophils & peripheral blood mononuclear cells | 10 mM | - neutrophils | [192] |
| In vivo | Wistar albino rats | 10 mg/kg | Increasing the sensitivity of the immune cells to some cytokines | [193] |
| In vitro & In vitro | Thymocytes of Barbari goats Thymus | 500 pg/mL 500 pg/mL | Restoring the suppressed immunity of T-cell cultured by developing some hormonal microcircuit (gonadal steroid and melatonin) in lymphatic organs | [194] |
| Modulating immune mediator production | ||||
| In vitro | Human mononuclear cells | 10−8 M | Increasing IL-2, IFN-γ and IL-6 in monocytes | [195] |
| In vitro | Neutrophils & peripheral blood mononuclear cells | 10 mM | Mel & AFMK: decreasing IL-8 and TNF-α in neutrophils | [192] |
| In vitro | RAW264.7 cells | 10, 100 or 1000 μM | Decreasing IL-1β, IL-6, IL-8, IL-10 and TNF-α in macrophages | [196] |
| Regulating the ROS production in the essential immune cells | ||||
| In vitro | Human monocytes | 10−12 M and above | Activating monocytes (above the activation threshold of 5 × 10−11 M) | [199] |
| In vitro | Lung neutrophils | 0.01, 0.1, 1 mM (dose-dependent) | Activating neutrophils | [200] |
| In vivo | Hamsters | 25 μg/100 g b.w. | Attenuating oxidative load | [201] |
| In vivo | Wild birds | 25 μg/100 g/day | Alleviating oxidative damage and suppressing the immune status induced by stress | [202]. |
| In vitro & in vivo | Heart tissue of C57BL/6 C57BL/6J mice | 3 or 4 doses of melatonin 30 mg/kg | Suppressing systemic innate immune activation by blocking the NF-κB/NLRP3 connection through a sirtuin1-dependent pathway | [154]. |
| Improving Circadian Rhythm and Sleep | ||||
| In vivo | C3H & C57BL mice | N/A | Being involved in the control of clock gene protein levels in the adrenal cortex of mice | [211] |
| In vivo | Soay sheep | N/A | Resetting circadian rhythms in the pituitary pars tuberalis | [212] |
| In vivo | Mice (C3H/HeJCrl and C57BL/6NCrl) | N/A | Influencing PER1 and CRY2 protein levels Playing a role in rhythmic regulation of pCREB levels in the mammalian retina | [213] |
| In vitro & in vivo | COS7 cells Lambs | N/A | Activating Npas4 | [215] |
| In vivo | Hamster | 20 μg/day | Coordinating the diurnal rhythm in neuronal remodeling | [217] |
| In vivo | Mice | 6 μg/day for 2 weeks | Increasing amplitude in expressional rhythms Altering the expression of genes of serotonergic neurotransmission Improve the depression-like behavior | [218] |
| In vivo | 23 patients | N/A | Being positive correlated with sleep parameters | [220] |
| Anticancer Activities | ||||
| Effects on tumor cell cycle, incl. growth, proliferation, metabolism and apoptosis | ||||
| In vitro & In vivo | Human gastric cancer cell lines (AGS and MKN) Male BALB/c nude mice | 5 mg/kg/twice/week for 33 days 1 µM to 2 mM (dose-/time- dependent, 15 min to 24 h) | Inhibiting gastric tumor growth and peritoneal metastasis Inhibiting C/EBPβ and NF-κB Inducing ER stress and inhibiting EMT | [232] |
| In vitro | T47D-BAF co-cultured | 20 nM | Suppressing breast cancer cell proliferation and inhibiting aromatase | [238] |
| In vitro & in vivo | Prostate cancer cells TRAMP male mice | 1 mM 200 µg/mL | Reducing glucose uptake and modifying the expression of GLUT1 transporter Attenuating glucose-induced tumor progression and prolonging the lifespan | [233] |
| In vitro | Hypoxic prostate cancer cell line PC-3 cells | 1 mM | Anti-angiogenic property Upregulating miRNA3195 and miRNA 374b and downregulating 16 miRNAs | [239] |
| In vitro | Colorectal cancer LoVo cells | 0.1–2.0 mM (dose-dependent) | Suppressing cell proliferation and inducing apoptosis Inducing dephosphorylation and nuclear import of histone deacetylase 4 (HDAC4) Decreasing H3 acetylation by inactivating CaMKIIα and reducing bcl-2 expression | [240] |
| In vitro | Breast cancer cell line SK-BR-3 & MDA-MB-231 | 2 mM | Changing the protein levels of Survivin, Bcl-2, and Bax Affecting cyt c release from the mitochondria to the cytosol Enhancing apoptotic cell death via sustained upregulation of Redd1 expression and inhibition of mTORC1 upstream of the activation of the p38/JNK pathways | [234] |
| Effects on invasion and metastasis of tumor cells | ||||
| In vitro | HepG2 liver cancer cells | 1 mM | Exhibiting anti-invasive and antimetastatic activities by suppressing the activity of MMP-9 Reducing IL-1β-induced HepG2 cells MMP-9 gelatinase activity and inhibiting cell invasion and motility through downregulation of MMP-9 gene expression and upregulation of the MMP-9-specific inhibitor tissue inhibitor of TIMP-1 Suppressing IL-1β-induced NF-κB translocation and transcriptional activity | [241] |
| In vitro | Renal cell carcinoma cells (Caki-1 and Achn) | 0.5–2 mM | Reducing the migration and invasion Inhibiting MMP-9 by reducing p65- and p52-DNA-binding activities Regulating MMP-9 transactivation and cell motility refer to the Akt-mediated JNK1/2 and ERK1/2 signaling pathways | [242] |
| In vivo & in vitro | Female athymic nude mice Metastatic and non-metastatic breast cancer cell lines (MDA-MB-231) | 100 mg/kg/day 1 mM | Lowering the numbers of lung metastasis Decreasing ROCK-1 protein expression in metastatic foci Reducing cell viability and invasion/migration Decreasing ROCK-1 gene expression in metastatic cells and protein expression in non-metastatic cell line | [235] |
| Therapy adjunct in tumor treatment | ||||
| In vitro | Human non-small-cell lung cancer (NSCLC) cells lines H1299 and A549 | 1 mM | Enhancing the berberine-mediated growth inhibition of lung cancer cells through simultaneous modulation of caspase/cyt C, AP-2β/hTERT, NF-κB/COX-2, and Akt/ERK signaling pathways | [157] |
| In vitro | Breast cancer cells | 1 nM | Mediating the sensitization to the ionizing radiation by decreasing around 50% the activity and expression of proteins involved in the synthesis of estrogens Reducing the amount of active estrogens at cancer cell level Inducing a 2-fold change in p53 expression compared to radiation alone | [243] |
| In vivo | Female patients | Melatonin-containing cream for twice daily use | Significantly lowering the occurrence of grade 1/2 acute radiation dermatitis in patients with breast-conserving surgery for stage 0–2 breast cancer | [244] |
| In vivo | Male Wistar rats | 10 mg/kg/week | Mitigating PVB-induced testicular dysfunction | [245] |
| In vivo & in vitro | Female athymic nude mice Human colon cancer cell lines SW620 | 25 mg/kg 1 mmol/L | Exerting synergistic anti-tumor effect by inhibiting the AKT and iNOS pathway Enhancing the 5-FU-mediated inhibition of cell proliferation, colony formation, cell migration and invasion Synergizing with 5-FU to promote the activation of the caspase/PARP-dependent apoptosis pathway and induce cell cycle arrest Synergizing anti-tumor effect of 5-FU by targeting the PI3K/AKT and NF-κB/iNOS signaling | [70] |
| In vitro | Human colorectal cancer cells | N/A | MT2 mRNA expression levels increased The profile of melatonin receptors gene expression and genes associated with their activity in colorectal cancer | [236] |
| In vitro | Estrogen receptor-positive endometrial cancer cell line, Ishikawa | 1 × 10−9 M | MT1 receptor expressing but not MT2 Attenuating ERα mRNA expression Enhancing anti-tumor effects of paclitaxel among anticancer drugs tested | [237] |
| Cardiovascular Protection | ||||
| In vivo | Patients with confirmed nocturnal hypertension | 2 mg 2 h before bedtime for 4 weeks | Reducing nocturnal systolic and diastolic BP significantly (p = 0.01) | [249] |
| In vivo | Spinal cord injury (SCI) mice model | 5, 10, 25, 50, 100 mg/kg i.p. | 50 mg/kg exhibiting significantly reduced blood spinal cord barrier permeability Restraining microvessel loss and attenuating edema Protecting the tight junction proteins, endothelial cells and pericytes Decreasing cell apoptosis and reducing MP3/AQP4/HIF-1α/VEGF/VEGFR2 expression | [251] |
| In vivo | Wistar-Kyoto (WKY) and spontaneously hypertensive rats (SHR) | 30 mg/kg/day for 4 weeks | Decreasing reflex chronotropic responses to phenylephrine and sodium nitroprusside Reducing mean arterial pressure and heart rate Improving bradycardic and tachycardic baroreflex responses without modifying catecholamine responses Increasing glutathione peroxidase activity in plasma and erythrocytes | [252] |
| In vivo | Wistar-Kyoto (WKY) and spontaneously hypertensive rats (SHR) | 30 mg/kg/day for 4 weeks | Decreasing mean arterial pressure (MAP) and heart rate Restoring the plasma noradrenaline concentrations, the chronotropic response to isoproterenol and the proportions of β1/β2-adrenoceptors in the heart in SHRs to the levels Decreasing the release of [3H] noradrenaline from isolated atria Improving the relaxation in the aorta | [253] |
| In vivo | Rats | 50 mg/kg | Preventing vasculitis Decreasing elementary pathological lesions of radiation-induced heart disease (RIHD) like fibrosis and necrosis | [254] |
| In vitro | BM-MSCs | 200, 20, and 2 µM(dose-dependent) | Reducing BM-MSC apoptotic death while increasing the levels of TGF-β, bFGF, VEGF, PDGF and Bcl-2, and decreasing Bax, p53 Upregulating modulator of apoptosis (PUMA) and caspase 3 Upregulating the phosphorylation of AMPK, which promotes ACC phosphorylation | [255] |
| In vivo & in vitro | Female C57BL/6a mice with MI Adipose-derived MSCs | 20 mg/kg/day for 28 days 5 µM | Promoting functional survival of AD-MSCs in infarcted heart and provoking a synergetic effect with AD-MSCs to restore heart function associated with alleviated inflammation, apoptosis, and oxidative stress in infarcted heart Exerting cytoprotective effects against hypoxia/serum deprivation (H/SD) injury Attenuating inflammation, apoptosis, and oxidative stress Enhancing SIRT1 signaling, with the increased expression of anti-apoptotic protein Bcl-2, and decreased the expression of Ac-FoxO1, Ac-p53, Ac-NF-KappaB, and Bax. | [256] |
| In vitro | Perfused isolated rat hearts and cultured neonatal rat cardiomyocytes | 5 µM | Improving postischemic cardiac function, decreasing infarct size, reducing apoptotic index, and diminishing lactate dehydrogenase release Upregulating the anti-apoptotic protein Bcl-2 and downregulating Bax Preserving mitochondrial redox potential and elevating SOD activity Decreasing formation of mitochondrial H2O2 and MDA | [257] |
| In vivo | Rats with sepsis | 30 mg/kg | Improving survival rates and cardiac function, attenuating myocardial injury and apoptosis Decreasing the serum LDH, decreasing inflammatory cytokines TNF-α, IL-1β, and HMGB1 Increasing anti-oxidant enzyme activity and p-Akt and Bcl-2 levels | [258] |
| In vivo | Drosophila melanogaster | 5 µM | Increasing the regularity of heartbeat, rescuing rhythmicity in flies bearing mutations, increasing cardiac regularity independent of alteration of heart rate, which is mediated via a specific G-Protein-coupled receptor encoded by the CG 4313 gene | [259] |
| In vivo | Patients with heart failure | N/A 1-year follow-up | As a predictors of left ventricular reverse remodeling (LVRR) and the adverse clinical events, increasing the area under of curve for the prediction LVRR | [260] |
| In vivo | Mice with Mst1 transgenic (Mst1 Tg) and Mst1 knockout (Mst1−/− ) | 20 mg/kg/d for 1 week | Alleviating postinfarction cardiac remodeling and dysfunction by upregulating autophagy, decreasing apoptosis, and modulating mitochondrial integrity and biogenesis via Mst1/Sirt1 signaling | [261] |
| Anti-diabetic Activities | ||||
| In vivo | Albino Wistar rats | 10 mg/kg b.w. | Increasing the inhibited activity of catalase in liver cells Restoring the dysfunctional mitochondria related to diabetes | [274] |
| In vivo | Rat | 2.8, 14, 28, and 140 nM | Inhibiting hepatic gluconeogenesis Activating hypothalamic Akt via membrane receptors MT1 and MT2 | [275] |
| In vivo | Rat | 10 mg/kg/day | Increasing Ca2+ levels in lots of organs and tissues | [11] |
| In vitro & in vivo | H9C2 cell line Rat | 0.1, 1, 10, 100, 1000 µM 20 mg/kg/day | Activating of SIRT1 signaling pathway (significant at 100 and 1000 µM) Inactivating PERK/eIF2α/ATF4 signaling pathway | [276] |
| In vitro | INS 832/13 cells | 1–100 nM | Attenuating β-cell apoptosis, improving β-cell function, prolonging β-cell survival (particularly evident at 10 nM) | [277] |
| In vivo | Rat | 10 mg/kg/day | Improving neurogenesis, synaptogenesis in hippocampi, increasing the receptors of melatonin and insulin, and restoring the downstream signaling pathway for insulin | [278] |
| In vivo | Rat | 250 µg/animal/day/i.p. | Accelerating bone healing | [279] |
| In vivo | Rat | 10 mg/kg/d, i.p. | Restoring the endothelial dysfunction and improving vascular responses | [271] |
| Anti-obese Activities | ||||
| In vivo | Rat | 10 mg/kg/day | Inducing white adipose tissue browning in rats with obesity-related type 2 diabetes | [282] |
| In vivo | Rat | 20 mg/L | Benefiting homeostasis of renal glutathione | [283] |
| In vitro | Mouse Gsk3b knockout (Gsk3b−/−) and wild-type (Gsk3b+/+) MEF cells | 10 nM | Inhibiting Akt activation Increasing GSK3B activity | [169] |
| In vivo | Mice | 100 mg/kg/day | Ameliorating obesity-induced adipokine alteration | [285] |
| In vivo | Women | N/A | Melatonin was involved in the development of obesity | [288] |
| In vitro | Mice | 20 mg/kg/day | Promoting circadian rhythm-mediated proliferation in adipose tissue | [206] |
| In vivo | Rat | 4 mg/kg/day | Decreasing myocardial infarct sizes and insulin resistant Increasing serum PKB/Akt, ERK42/44, GSK-3β and STAT3 | [289] |
| In vivo | Mice | 100 mg/kg/day | Increasing mitofusin-2 expression | [290] |
| In vivo | Mice | 10 mg/kg/day | Modulating the MAPK-JNK/p38 signaling pathway | [291] |
| Neuroprotection | ||||
| In vivo | Mice | 10 mg/kg | Increasing the activity of antioxidant enzymes Mediating the Nrf2-ARE pathway | [293] |
| In vivo | C57BL/6J mice | 10 mg/kg given twice | Reducing IR-induced mitochondrial dysfunction Activating SIRT1 signaling | [294] |
| In vivo | Rat | 150 mg/kg | Suppressing cortical expressions of proinflammatory cytokines | [295] |
| In vivo | Mice | 5 mg/kg | Reducing oxidative damage by scavenging radicals | [296] |
| In vivo | Rat | 10 mg/kg | Reversing the increased plasma TNF-α, IL-1β levels Decreasing BDNF, S100B and IL-10 values | [297] |
| In vivo | Rat | 10 mg/kg and 50 mg/kg | Preventing the decrease of the number and the diameter of sciatic nerve axons | [298] |
| In vivo | Rat | 20 mg | Preventing the decrease in VEPs and PLR Inhibiting microglial reactivity, astrocytosis, demyelination, and axon and retinal ganglion cell loss Preserving anterograde transport of cholera toxin β-subunit | [299] |
| In vivo | Mice | 10 mg/kg | Restoring mRNA and protein levels of BACE1 and PS1 | [305] |
| In vitro & in vivo | Rat hippocampal neurons Rat | 50 μM 500 mg/kg b.w. | Improving the soluble Abeta1–42-induced impairment of spatial learning and memory, synaptic plasticity and astrogliosis | [301] |
| In vivo | Rat | 10 mg/kg | Improving motor activity and muscular strength | [306] |
| In vitro | Mouse neuroblastoma cells | 1 μM | Activating transcription factor EB-dependent autophagy-lysosome | [136] |
| In vivo | Rat | 100 mg/kg | Inhibiting caspase-3 | [307] |
| In vivo | Rat | 50 mg/kg/day | Protecting the cell against neuronal damage in the hippocampus | [308] |
| Other Bioactivities | ||||
| In vivo | Rat | 10 mg/kg/day | Improving the microstructure and biomechanical properties of aged bones | [311] |
| In vivo | Patients | 10 mg/day, 60 mg/day | Reducing the hyperoxidative and inflammatory process | [150] |
| In vivo | Mice | 30 mg/kg/day | Decreasing plasma creatine kinase activity, increasing total glutathione content Lowering the oxidized/reduced glutathione ratio | [312] |
| In vitro | NCI-H292 cells | 50, 100, 200, and 400 μM (dose-dependent) | Inhibiting mucin 5AC production | [313] |
| In vivo | Rat | 4 mg/kg, i.p 10 mg/kg, i.p | Exhibits renoprotective effects against ischemia reperfusion induced AKI due to antioxidant properties and the involvement of progesterone receptors | [314] |
| In vivo | Rat | 10 mg/kg/day | Scavenging free radicals | [315] |
| In vivo | Rat | N/A | Activating SIRT1 signaling | [316] |
| In vitro | Human ASCs | 100 μM for 3 h | Enhancing human ASCs’ survival and their therapeutic effectiveness on injured tissue | [317] |
| In vivo | Rat | 10 mg/body | Interacting with other hormones | [319] |
| In vivo | Rat | 10 mg/kg/day | Decreasing the increased myeloperoxidase activities and osteoclast and neutrophil densities | [322] |
| In vivo | Rat | 10 mg/kg/day | Decreasing serum cyclophosphamide levels and increasing ALP levels | [323] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, X.; Li, Y.; Li, S.; Zhou, Y.; Gan, R.-Y.; Xu, D.-P.; Li, H.-B. Dietary Sources and Bioactivities of Melatonin. Nutrients 2017, 9, 367. https://doi.org/10.3390/nu9040367
Meng X, Li Y, Li S, Zhou Y, Gan R-Y, Xu D-P, Li H-B. Dietary Sources and Bioactivities of Melatonin. Nutrients. 2017; 9(4):367. https://doi.org/10.3390/nu9040367
Chicago/Turabian StyleMeng, Xiao, Ya Li, Sha Li, Yue Zhou, Ren-You Gan, Dong-Ping Xu, and Hua-Bin Li. 2017. "Dietary Sources and Bioactivities of Melatonin" Nutrients 9, no. 4: 367. https://doi.org/10.3390/nu9040367
APA StyleMeng, X., Li, Y., Li, S., Zhou, Y., Gan, R.-Y., Xu, D.-P., & Li, H.-B. (2017). Dietary Sources and Bioactivities of Melatonin. Nutrients, 9(4), 367. https://doi.org/10.3390/nu9040367