Intra Amniotic Administration of Raffinose and Stachyose Affects the Intestinal Brush Border Functionality and Alters Gut Microflora Populations

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Design
2.2. Intra-Amniotic Administration
2.3. Tissue Collection
2.4. Isolation of Total RNA
2.5. Gene Expression Analysis
2.6. Bacterial Analysis
2.7. Assessment of Liver Ferritin
2.8. Blood Analysis and Hb Measurements
2.9. Morphological Examination of the Intestinal Villi
2.10. Goblet Cell Diameter
2.11. Statistical Analysis
3. Results
3.1. Intestinal Content Bacterial Genera- and Species-Level Analysis
3.2. BBM Functional Genes
3.3. Fe Metabolism Genes
3.4. Cecum-to-Body-Weight Ratio
3.5. Morphometric Data for Villi
3.6. Goblet Cell Diameters
3.7. Liver Ferritin and Hb
4. Discussion
5. Conclusions
Author Contributions
Conflicts of Interest
Abbreviations
| Fe | iron |
| Hb | hemoglobin |
| DMT1 | divalent metal transporter 1 |
| DcytB | duodenal cytochrome B |
| AP | amino peptidase |
| SI | sucrose isomaltase |
| SGLT-1 | sodium glucose transporter 1 |
| BBM | brush border membrane |
References
- World Health Organization. Iron Deficiency Anemia: Assessment, Prevention and Control: A Guide for Programme Managers; World Health Organization: Geneva, Switzerland, 2001; pp. 15–21. [Google Scholar]
- World Health Organization. The Global Prevalence of Anaemia in 2011; World Health Organization: Geneva, Switzerland, 2015; pp. 3–6. [Google Scholar]
- Pollitt, E.; Soemantri, A.G.; Yunis, F.; Scrimshaw, N.S. Cognitive effects of iron-deficiency Anaemia. Lancet 1985, 325, 158. [Google Scholar] [CrossRef]
- Johnson, C.R.; Thavarajah, D.; Combs, G.F.; Thavarajah, P. Lentil (Lens culinaris L.): A prebiotic-rich whole food legume. Food Res. Int. 2013, 51, 107–113. [Google Scholar] [CrossRef]
- Dwivedi, S.; Sahrawat, K.; Puppala, N.; Ortiz, R. Plant prebiotics and human health: Biotechnology to breed prebiotic-rich nutritious food crops. Electron. J. Biotechnol. 2014, 17, 238–245. [Google Scholar] [CrossRef]
- Nestel, P.; Bouis, H.E.; Meenakshi, J.V.; Pfeiffer, W. Biofortification of staple food crops. J. Nutr. 2006, 136, 1064–1067. [Google Scholar] [PubMed]
- Welch, R.M.; Graham, R.D. Breeding for micronutrients in staple food crops from a human nutrition perspective. J. Exp. Bot. 2004, 55, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Wiesinger, J.A.; Cichy, K.A.; Glahn, R.P.; Grusak, M.A.; Brick, M.A.; Thompson, H.J.; Tako, E. Demonstrating a Nutritional Advantage to the Fast-Cooking Dry Bean ( Phaseolus vulgaris L.). J. Agric. Food Chem. 2016, 64, 8592–8603. [Google Scholar] [CrossRef] [PubMed]
- Tako, E.; Beebe, S.E.; Reed, S.; Hart, J.J.; Glahn, R.P. Polyphenolic compounds appear to limit the nutritional benefit of biofortified higher iron black bean (Phaseolus vulgaris L.). Nutr. J. 2014, 13, 1. [Google Scholar] [CrossRef] [PubMed]
- Petry, N.; Egli, I.; Zeder, C.; Walczyk, T.; Hurrell, R. Polyphenols and phytic acid contribute to the low iron bioavailability from common beans in young women. J. Nutr. 2010, 140, 1977–1982. [Google Scholar] [CrossRef] [PubMed]
- Petry, N.; Egli, I.; Gahutu, J.B.; Tugirimana, P.L.; Boy, E.; Hurrell, R. Phytic acid concentration influences iron bioavailability from biofortified beans in Rwandese women with low iron status. J. Nutr. 2014, 144, 1681–1687. [Google Scholar] [CrossRef] [PubMed]
- Tako, E.; Glahn, R.P.; Knez, M.; Stangoulis, J.C. The effect of wheat prebiotics on the gut bacterial population and iron status of iron deficient broiler chickens. Nutr. J. 2014, 13, 1. [Google Scholar] [CrossRef] [PubMed]
- Tako, E.; Rutzke, M.A.; Glahn, R.P. Using the domestic chicken (Gallus gallus) as an in vivo model for iron bioavailability1. Poult. Sci. 2010, 89, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Tako, E.; Reed, S.M.; Budiman, J.; Hart, J.J.; Glahn, R.P. Higher iron pearl millet (Pennisetum glaucum L.) provides more absorbable iron that is limited by increased polyphenolic content. Nutr. J. 2015, 14, 1. [Google Scholar] [CrossRef] [PubMed]
- Hillier, L.W.; Miller, W.; Birney, E.; Warren, W.; Hardison, R.C.; Ponting, C.P.; Bork, P.; Burt, D.W.; Groenen, M.A.M.; Delany, M.E.; et al. Sequence and comparative analysis of the chicken genome provide unique perspectives on vertebrate evolution. Nature 2004, 432, 695–716. [Google Scholar] [CrossRef] [PubMed]
- Tako, E.; Glahn, R.P. Intra-amniotic administration and dietary inulin affect the iron status and intestinal functionality of iron-deficient broiler chickens. Poult. Sci. 2012, 91, 1361–1370. [Google Scholar] [CrossRef] [PubMed]
- Tako, E.; Ferket, P.; Uni, Z. Changes in chicken intestinal zinc exporter mRNA expression and small intestinal functionality following intra-amniotic zinc-methionine administration. J. Nutr. Biochem. 2005, 16, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Hartono, K.; Reed, S.; Ankrah, N.A.; Glahn, R.P.; Tako, E. Alterations in gut microflora populations and brush border functionality following intra-amniotic daidzein administration. RSC Adv. 2015, 5, 6407–6412. [Google Scholar] [CrossRef]
- Zhu, X.Y.; Zhong, T.; Pandya, Y.; Joerger, R.D. 16S rRNA-Based Analysis of Microbiota from the Cecum of Broiler Chickens. Appl. Environ. Microbiol. 2002, 68, 124–137. [Google Scholar] [CrossRef] [PubMed]
- Tako, E.; Glahn, R.P.; Welch, R.M.; Lei, X.; Yasuda, K.; Miller, D.D. Dietary inulin affects the expression of intestinal enterocyte iron transporters, receptors and storage protein and alters the microbiota in the pig intestine. Br. J. Nutr. 2008, 99, 472–480. [Google Scholar] [CrossRef] [PubMed]
- Passaniti, A.; Roth, T.F. Purification of chicken liver ferritin by two novel methods and structural comparison with horse spleen ferritin. Biochem. J. 1989, 258, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Mete, A.; van Zeeland, Y.R.A.; Vaandrager, A.B.; van Dijk, J.E.; Marx, J.J.M.; Dorrestein, G.M. Partial purification and characterization of ferritin from the liver and intestinal mucosa of chickens, turtledoves and mynahs. Avian Pathol. 2005, 34, 430–434. [Google Scholar] [CrossRef] [PubMed]
- Reed, S.; Qin, X.; Ran-Ressler, R.; Brenna, J.; Glahn, R.; Tako, E. Dietary zinc deficiency affects blood linoleic acid: Dihomo-γ-linolenic acid (LA:DGLA) ratio; a sensitive physiological marker of zinc status in vivo (Gallus gallus). Nutrients 2014, 6, 1164–1180. [Google Scholar] [CrossRef] [PubMed]
- Smirnov, A.; Tako, E.; Ferket, P.R.; Uni, Z. Mucin gene expression and mucin content in the chicken intestinal goblet cells are affected by in ovo feeding of carbohydrates. Poult. Sci. 2006, 85, 669–673. [Google Scholar] [CrossRef] [PubMed]
- Ludwiczek, S.; Theurl, I.; Artner-Dworzak, E.; Chorney, M.; Weiss, G. Duodenal HFE expression and hepcidin levels determine body iron homeostasis: Modulation by genetic diversity and dietary iron availability. J. Mol. Med. 2004, 82, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.; Beatty, E.; Wang, X.; Cummings, J.H. Selective stimulation of Bifidobacteria in the Human Colon by Oligofructose and Inulin. Gastroenterology 1995, 108, 975–982. [Google Scholar] [CrossRef]
- Roberfroid, M.; Van Loot, J.; Gibson, G. The bifidogenic nature of chicory inulin and its hydrolysis products. J. Nutr. 1998, 128, 11–19. [Google Scholar] [PubMed]
- Patterson, J.K.; Lei, X.G.; Miller, D.D. The pig as an experimental model for elucidating the mechanisms governing dietary influence on mineral absorption. Exp. Biol. Med. 2008, 233, 651–664. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Rastall, R.A. Prebiotics: Development & Application; John Wiley & Sons: Chichester, UK; Hoboken, NJ, USA, 2006. [Google Scholar]
- Tzortzis, G.; Goulas, A.K.; Gibson, G.R. Synthesis of prebiotic galactooligosaccharides using whole cells of a novel strain, Bifidobacterium bifidum NCIMB 41171. Appl. Microbiol. Biotechnol. 2005, 68, 412–416. [Google Scholar] [CrossRef] [PubMed]
- Tako, E.; Glahn, R.P. Iron status of the late term broiler (Gallus gallus) embryo and hatchling. Int. J. Poul. Sci. 2011, 10, 42–48. [Google Scholar] [CrossRef]
- Tako, E.; Ferket, P.R.; Uni, Z. Effects of in ovo feeding of carbohydrates and beta-hydroxy-beta-methylbutyrate on the development of chicken intestine. Poult. Sci. 2004, 83, 2023–2028. [Google Scholar] [CrossRef] [PubMed]
- Scholz-Ahrens, K.; Ade, P.; Marten, B.; Weber, P.; Timm, W.; Acil, Y.; Gluer, C.; Schrezenmeir, J. Prebiotics, probiotics, and synbiotics affect mineral absorption, bone mineral content, and bone structure. J. Nutr. 2007, 137, 838S–846S. [Google Scholar] [PubMed]
- Preidis, G.A.; Saulnier, D.M.; Blutt, S.E.; Mistretta, T.-A.; Riehle, K.P.; Major, A.M.; Venable, S.F.; Finegold, M.J.; Petrosino, J.F.; Conner, M.E.; et al. Probiotics stimulate enterocyte migration and microbial diversity in the neonatal mouse intestine. FASEB J. 2012, 26, 1960–1969. [Google Scholar] [CrossRef] [PubMed]
- Ouwehand, A.C.; Derrien, M.; de Vos, W.; Tiihonen, K.; Rautonen, N. Prebiotics and other microbial substrates for gut functionality. Curr. Opin. Biotechnol. 2005, 16, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Deplancke, B.; Gaskins, H.R. Microbial modulation of innate defense: Goblet cells and the intestinal mucus layer. Am. J. Clin. Nutr. 2001, 73, 1131S–1141S. [Google Scholar] [PubMed]
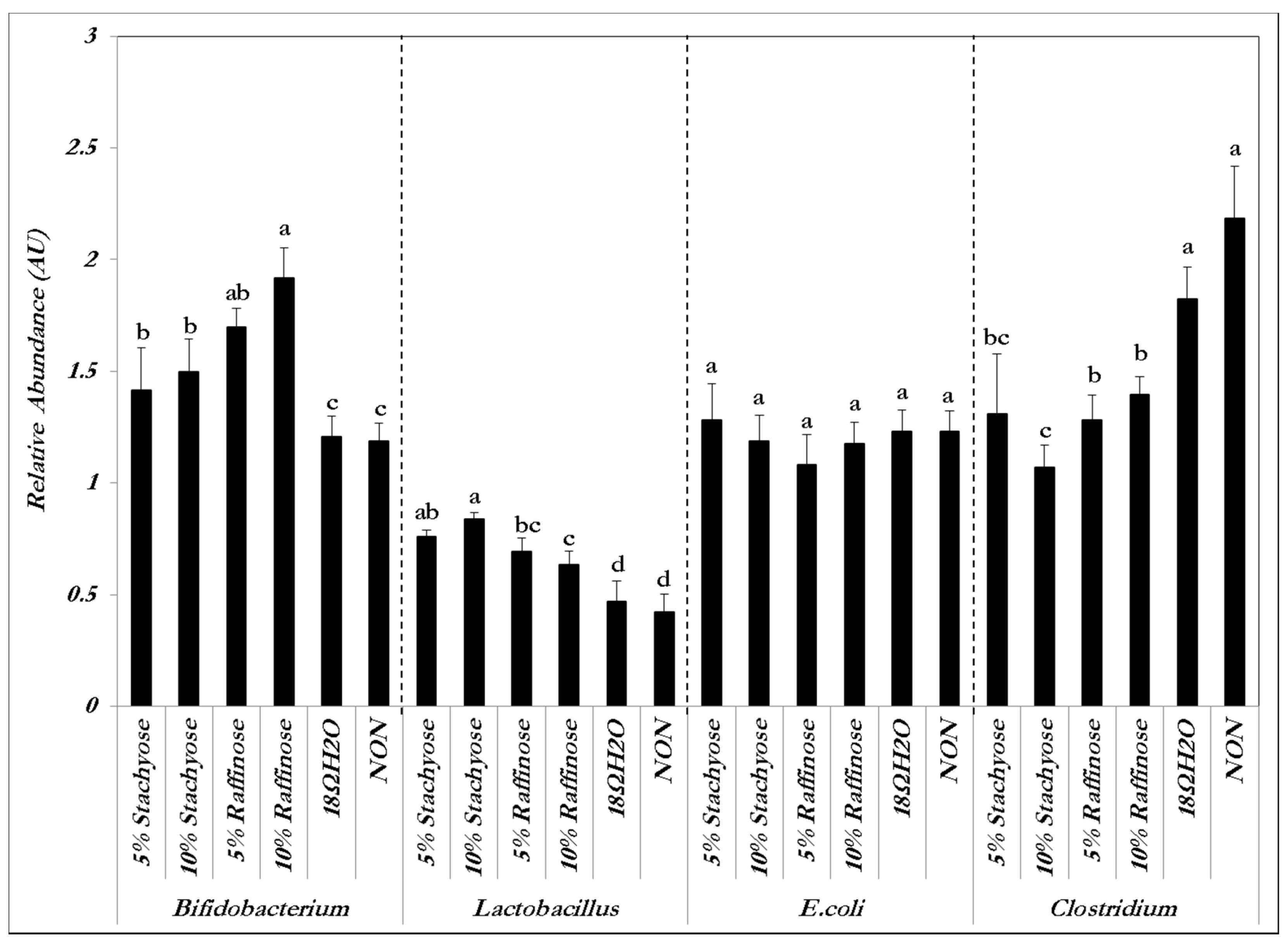
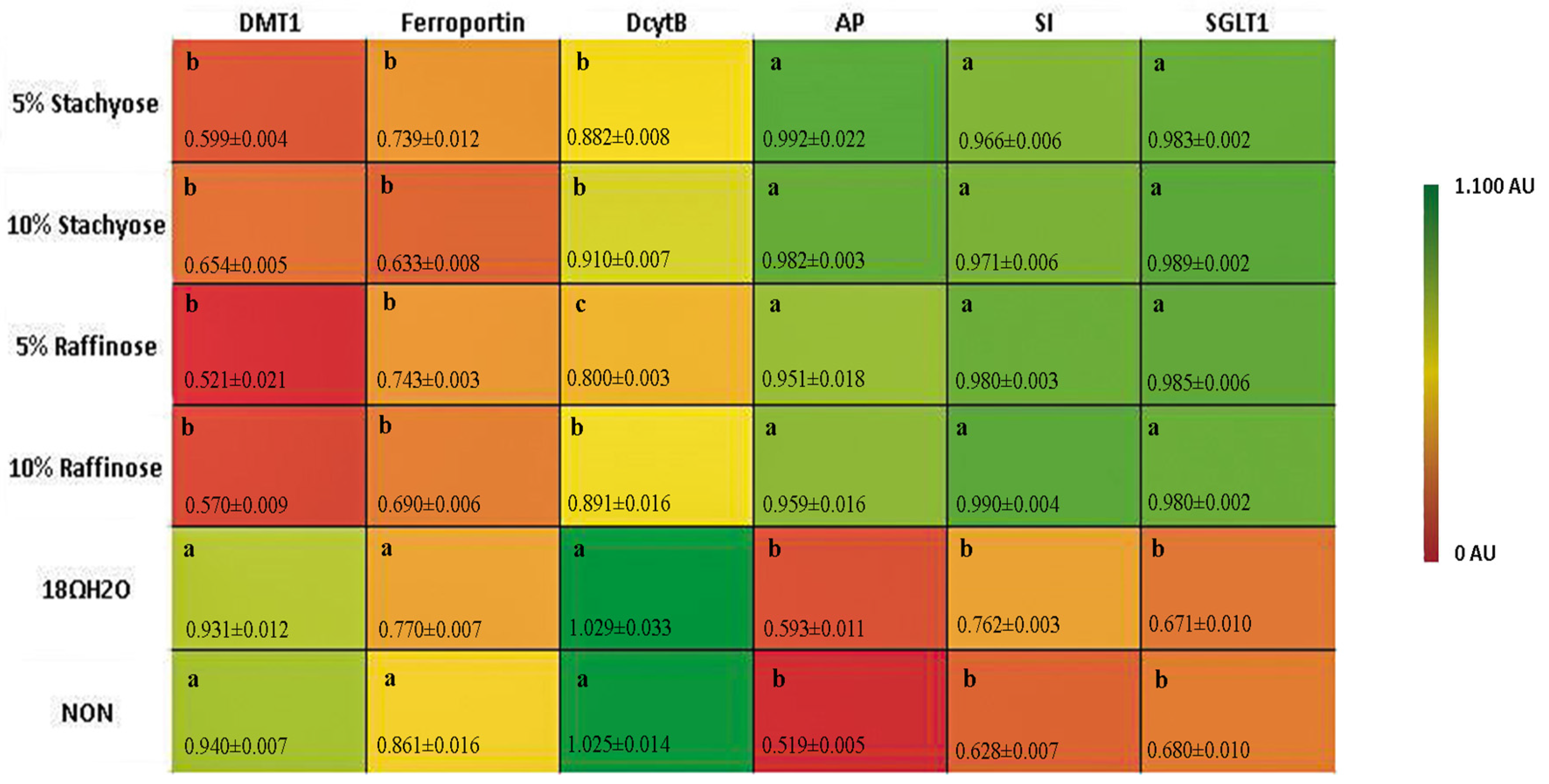

{kind=link}
{kind=link}
| Treatment Group | Cecum/Body Weight Ratio (%) |
|---|---|
| 5% Stachyose | 1.67 ± 0.21 a |
| 10% Stachyose | 1.63 ± 0.18 a |
| 5% Raffinose | 1.83 ± 0.20 a |
| 10% Raffinose | 1.55 ± 0.13 a |
| 18ΩH2O | 1.35 ± 0.08 b |
| Non-injected | 1.22 ± 0.07 b |
| Treatment Group | Villus Surface Area (mm2) | Goblet Cell Diameter (µm) |
|---|---|---|
| 5% Stachyose | 459.2 ± 32.09 a,b | 18.2 ± 0.143 c |
| 10% Stachyose | 493.8 ± 10.31 a | 19.1 ± 0.152 b |
| 5% Raffinose | 467.5 ± 35.55 a,b | 22.5 ± 0.180 a |
| 10% Raffinose | 425.2 ± 24.04 b,c | 19.5 ± 0.156 b |
| 18ΩH2O | 384.4 ± 14.16 c | 15.4 ± 0.123 d |
| Non-injected | 353.1 ± 13.24 c | 13.2 ± 0.105 e |
| Treatment Group | Ferritin (AU) | Hb (g/dL) |
|---|---|---|
| 5% Stachyose | 0.48 ± 0.09 a | 10.7 ± 0.54 a |
| 10% Stachyose | 0.48 ± 0.08 a | 11.1 ± 0.55 a |
| 5% Raffinose | 0.47 ± 0.10 a | 10.5 ± 0.68 a |
| 10% Raffinose | 0.48 ± 0.09 a | 11.0 ± 0.56 a |
| 18ΩH2O | 0.47 ± 0.09 a | 10.4 ± 0.42 a |
| Non-injected | 0.47 ± 0.07 a | 10.3 ± 0.65 a |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pacifici, S.; Song, J.; Zhang, C.; Wang, Q.; Glahn, R.P.; Kolba, N.; Tako, E. Intra Amniotic Administration of Raffinose and Stachyose Affects the Intestinal Brush Border Functionality and Alters Gut Microflora Populations. Nutrients 2017, 9, 304. https://doi.org/10.3390/nu9030304
Pacifici S, Song J, Zhang C, Wang Q, Glahn RP, Kolba N, Tako E. Intra Amniotic Administration of Raffinose and Stachyose Affects the Intestinal Brush Border Functionality and Alters Gut Microflora Populations. Nutrients. 2017; 9(3):304. https://doi.org/10.3390/nu9030304
Chicago/Turabian StylePacifici, Sarina, Jaehong Song, Cathy Zhang, Qiaoye Wang, Raymond P. Glahn, Nikolai Kolba, and Elad Tako. 2017. "Intra Amniotic Administration of Raffinose and Stachyose Affects the Intestinal Brush Border Functionality and Alters Gut Microflora Populations" Nutrients 9, no. 3: 304. https://doi.org/10.3390/nu9030304
APA StylePacifici, S., Song, J., Zhang, C., Wang, Q., Glahn, R. P., Kolba, N., & Tako, E. (2017). Intra Amniotic Administration of Raffinose and Stachyose Affects the Intestinal Brush Border Functionality and Alters Gut Microflora Populations. Nutrients, 9(3), 304. https://doi.org/10.3390/nu9030304