Wine Resveratrol: From the Ground Up




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
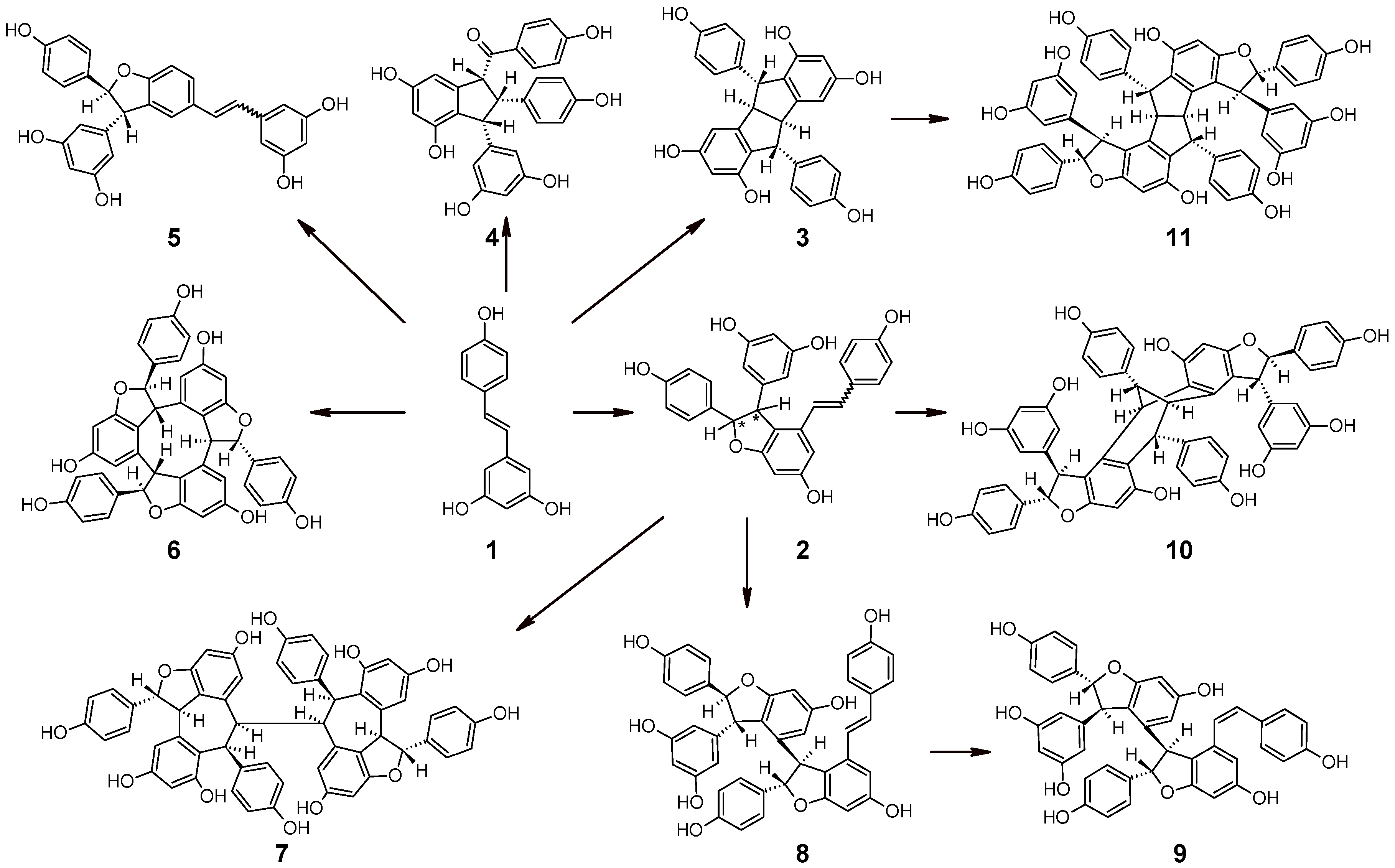
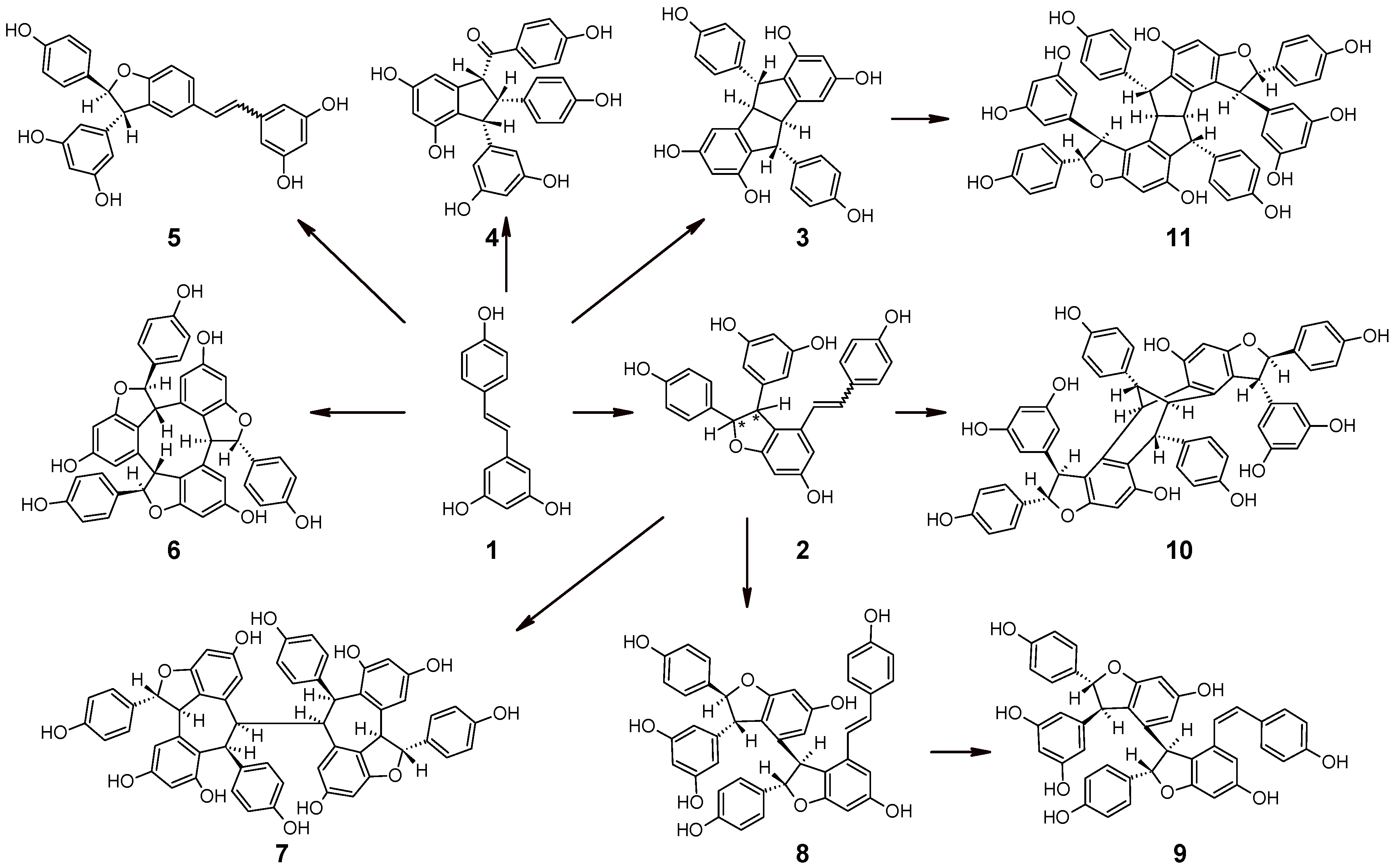
:1. Resveratrol and its Oligomers in Grape
2. Viticultural Factors and Grape Resveratrol
3. Oenological Factors and Resveratrol in Wine
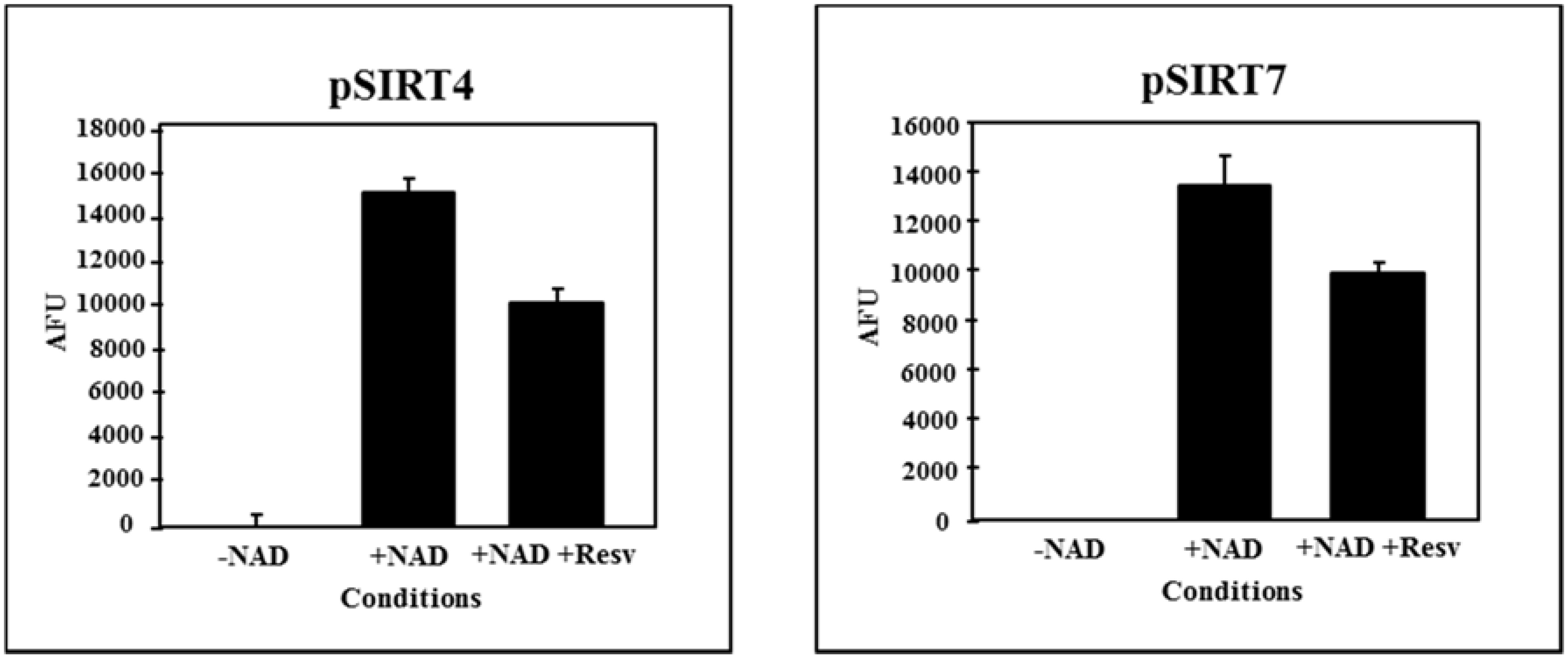
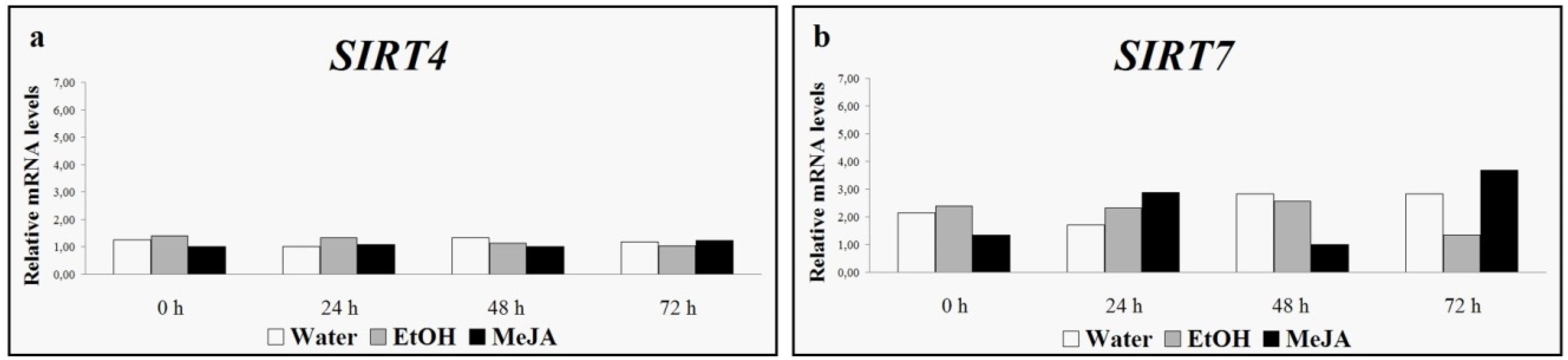
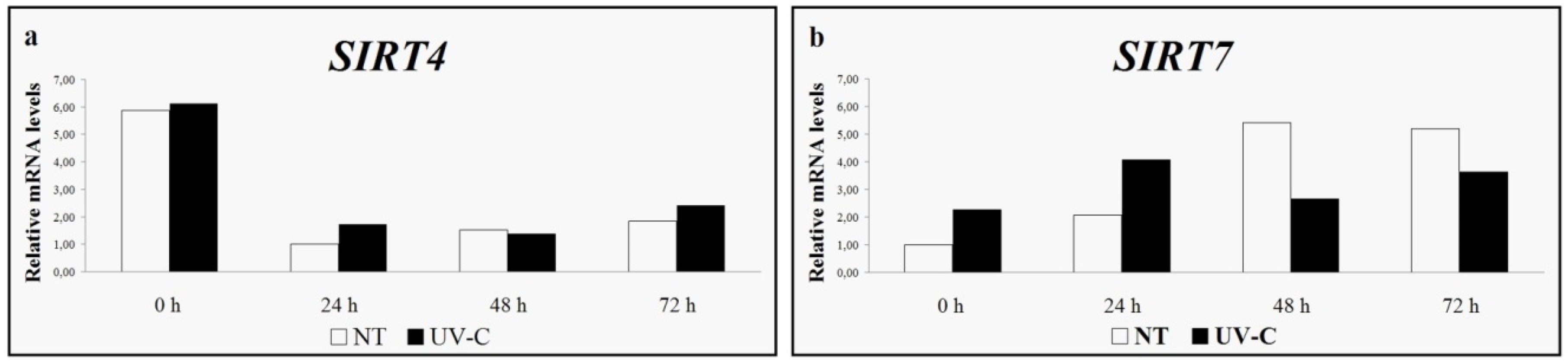
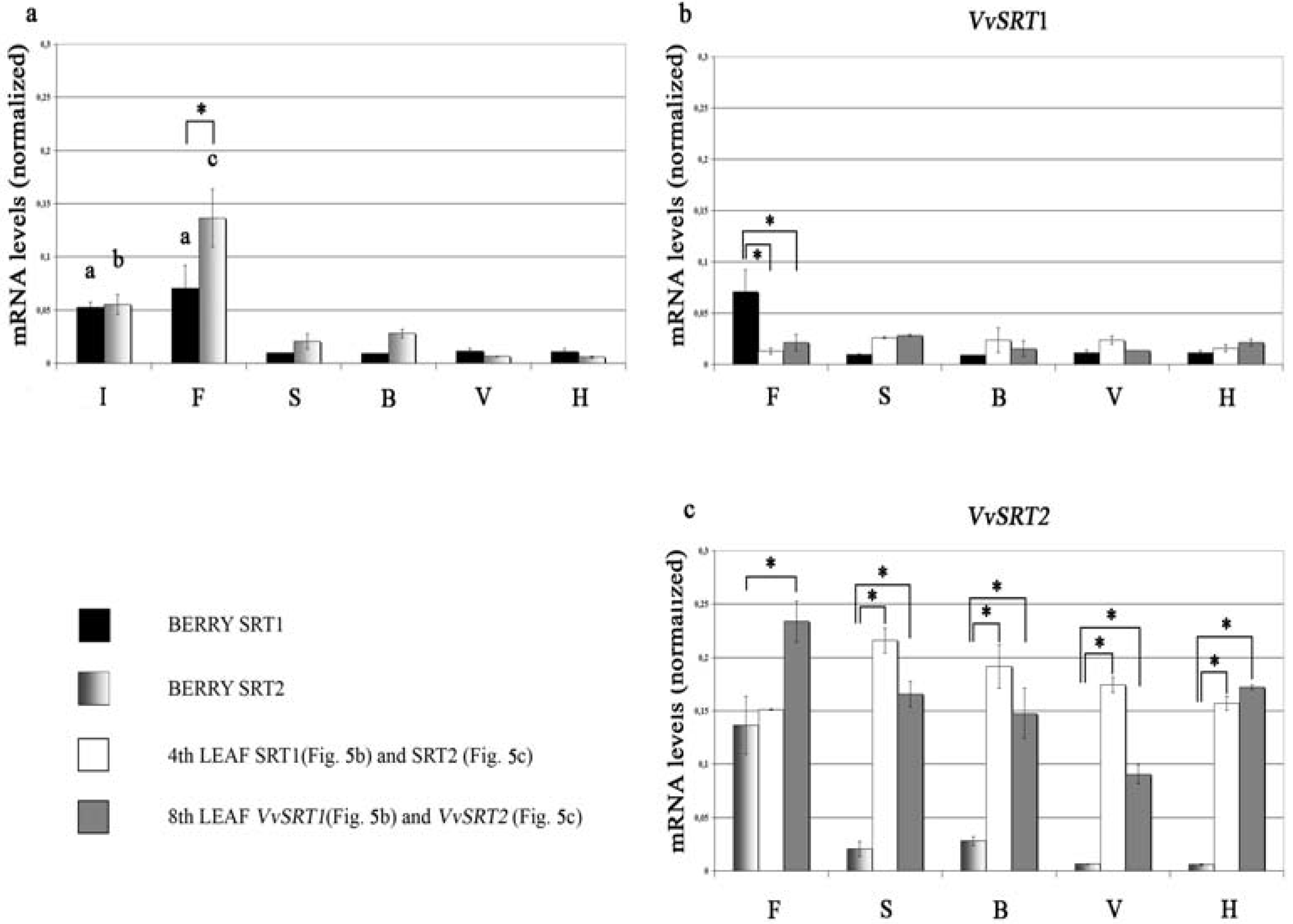
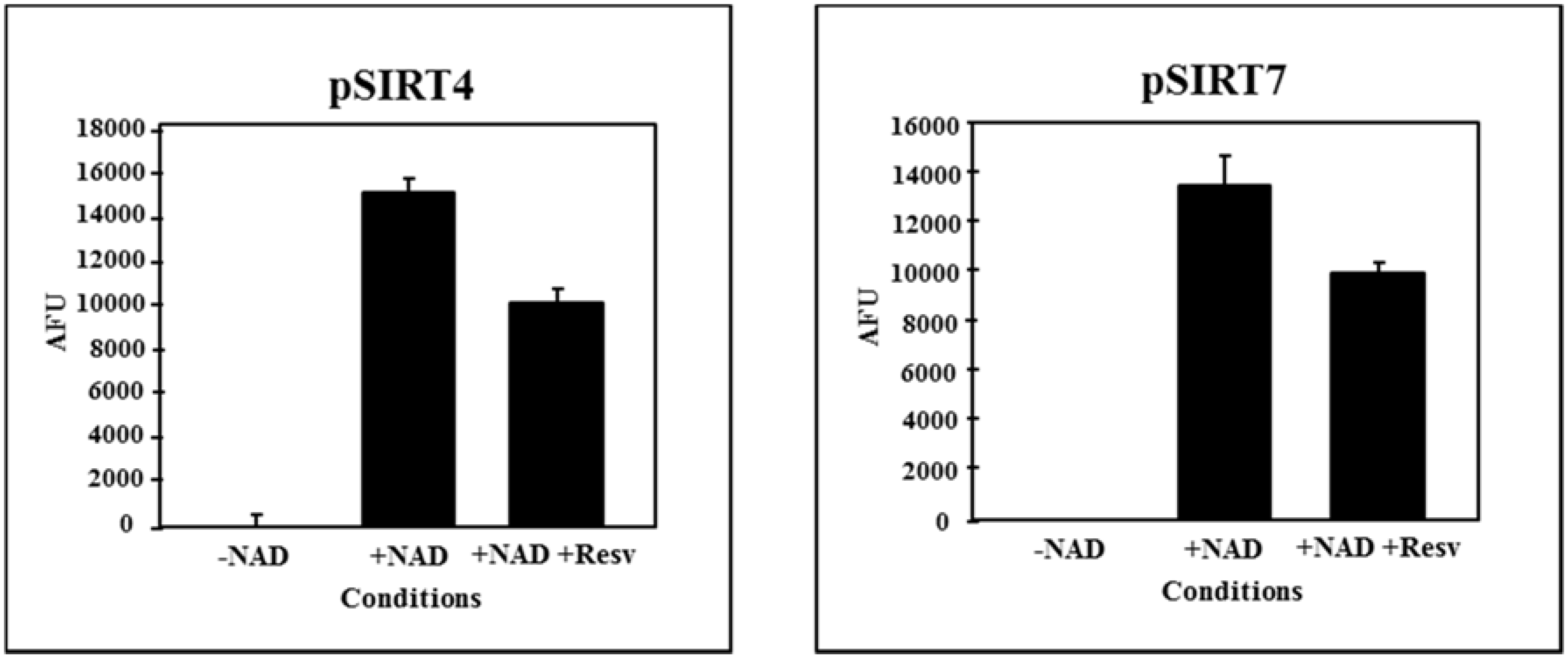
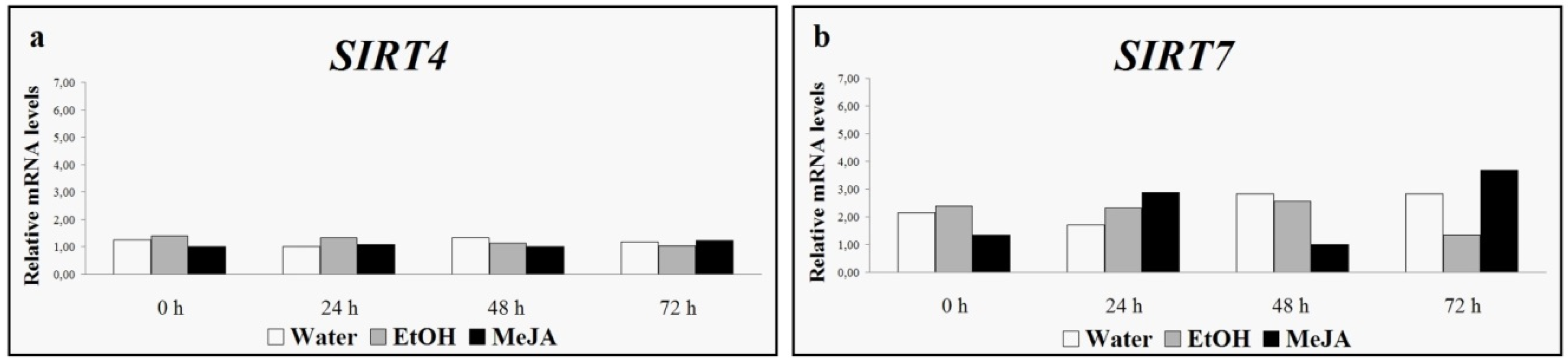
4. Grapevine Sirtuins
5. Conclusions
Conflicts of Interest
References
- Flamini, R.; Zanzotto, A.; de Rosso, M.; Lucchetta, G.; Vedova, A.D.; Bavaresco, L. Biosynthesis of stilbene oligomer phytoalexins in grape as a response to A. carbonarius infection. Physiol. Mol. Plant Pathol. 2016, 93, 112–118. [Google Scholar] [CrossRef]
- Mattivi, F.; Vrhovsek, U.; Malacarne, G.; Masuero, D.; Zulini, L.; Stefanini, M.; Moser, C.; Velasco, R.; Guella, G. Profiling of resveratrol oligomers, important stress metabolites accumulating in the leaves of hybrid V. vinifera (Merzling × Teroldego) genotypes infected with Plasmopara viticola. J. Agric. Food Chem. 2011, 59, 5364–5375. [Google Scholar] [CrossRef] [PubMed]
- Korhammer, S.; Reniero, F.; Mattivi, F. An oligostilbene from Vitis roots. Phytochemistry 1995, 38, 1501–1504. [Google Scholar] [CrossRef]
- Mattivi, F.; Reniero, F. Oligostilbenes from the roots of genus Vitis. Bull. Liaison Groupe Polyphen. 1992, 16, 116–118. [Google Scholar]
- Reniero, F.; Rudolph, M.; Angioni, A.; Bernreuther, A.; Cabras, P.; Mattivi, F. Identification of two stilbenoids from Vitis roots. Vitis 1996, 35, 125–127. [Google Scholar]
- Bavaresco, L.; Cantù, E.; Fregoni, M.; Trevisan, M. Constitutive stilbene contents of grapevine cluster stems as potential source of resveratrol in wine. Vitis 1997, 36, 115–118. [Google Scholar]
- Waffo-Teguo, P.; Lee, D.; Cuendet, M.; Merillon, J.M.; Pezzuto, J.M.; Kinghorn, A.D. Two new stilbene dimer glucosides from grape (Vitis vinifera) cell cultures. J. Nat. Prod. 2001, 64, 136–138. [Google Scholar] [CrossRef] [PubMed]
- Pezet, R.; Gindro, K.; Viret, O.; Richter, H. Effects of resveratrol, viniferins and pterostilbene on Plasmopara viticola zoospore mobility and disease development. Vitis 2004, 43, 145–148. [Google Scholar]
- Cichewicz, R.H.; Kouzi, S.A.; Hamann, M.T. Dimerization of resveratrol by the grapevine pathogen Botrytis cinerea. J. Nat. Prod. 2000, 63, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Goetz, G.; Fkyerat, A.; Métais, N.; Kunz, M.; Tabacchi, R.; Pezet, R.; Pont, V. Resistance factors to grey mould in grape berries: Identification of some phenolics inhibitors of Botrytis cinerea stilbene oxidase. Phytochemistry 1999, 52, 759–767. [Google Scholar] [CrossRef]
- Sbaghi, M.; Jeandet, P.; Bessis, R.; Leroux, P. Degradation of stilbene-type phytoalexins in relation to the pathogenicity of Botrytis cinerea to grapevines. Plant Pathol. 1996, 45, 139–144. [Google Scholar] [CrossRef]
- Hart, J.H. Role of phytostilbenes in decay and disease resistance. Annu. Rev. Phytopathol. 1981, 19, 437–458. [Google Scholar] [CrossRef]
- Bavaresco, L.; Fregoni, C.; van Zeller de Macedo Basto Goncalves, M.I.; Vezzulli, S. Physiology and molecular biology of grapevine stilbene: An update. In Grapevine Molecular Physiology and Biotechnology, 2nd ed.; Roubelakis-Angelakis, K.A., Ed.; Springer Science + Business Media B. V: London, UK, 2009; pp. 341–364. [Google Scholar]
- Jeandet, P.; Douillet-Breuil, A.C.; Bessis, R.; Debord, S.; Sbaghi, M.; Adrian, M. Phytoalexins from the Vitaceae: Biosynthesis, phytoalexin gene expression in transgenic plants, antifungal activity, and metabolism. J. Agric. Food Chem. 2002, 50, 2731–2741. [Google Scholar] [CrossRef] [PubMed]
- Bavaresco, L.; Morreale, G.; Flamini, R. Genetic and physiological bases of healthy attributes of grapes and wine. Acta Hortic. 2015, 1082, 263–274. [Google Scholar] [CrossRef]
- Bavaresco, L.; Pezzutto, S.; Gatti, M.; Mattivi, F. Role of the variety and some environmental factors on grape stilbenes. Vitis 2007, 46, 57–61. [Google Scholar]
- Gatto, P.; Vrhovsek, U.; Muth, J.; Segala, C.; Romualdi, C.; Fontana, P.; Pruefer, D.; Stefanini, M.; Moser, C.; Mattivi, F.; et al. Ripening and genotype control stilbene accumulation in healthy grapes. J. Agric. Food Chem. 2008, 56, 11773–11785. [Google Scholar] [CrossRef] [PubMed]
- Gatti, M.; Civardi, S.; Ferrari, F.; Fernandes, N.; van Zeller de Basto Gançalves, M.I.; Bavaresco, L. Viticultural performances of different Cabernet Sauvignon clones. Acta Hortic. 2014, 1046, 659–664. [Google Scholar] [CrossRef]
- Bavaresco, L.; Civardi, S.; Pezzutto, S.; Vezzulli, S.; Ferrari, F. Grape production, technological parameters, and stilbenic compounds as affected by lime induced chlorosis. Vitis 2005, 44, 63–65. [Google Scholar]
- Bavaresco, L.; Pezzutto, S.; Ragga, A.; Ferrari, F.; Trevisan, M. Effect of nitrogen supply on trans-resveratrol concentration in berries of Vitis vinifera L. cv. Cabernet Sauvignon. Vitis 2001, 40, 229–230. [Google Scholar]
- Bavaresco, L.; Gatti, M.; Pezzutto, S.; Fregoni, M.; Mattivi, F. Effect of leaf removal on grape yield, berry composition, and stilbene concentration. Am. J. Enol. Viticult. 2008, 59, 292–298. [Google Scholar]
- Gatti, M.; Civardi, S.; Zamboni, M.; Ferrari, F.; Elothmani, D.; Bavaresco, L. Preliminary results on the effect of cluster thinning on stilbene concentration and antioxidant capacity of V. vinifera L. “Barbera” wine. Vitis 2011, 50, 43–44. [Google Scholar]
- Gebbia, N.; Bavaresco, L.; Fregoni, M.; Civardi, S.; Crosta, L.; Ferrari, F.; Grippi, F.; Tolomeo, M.; Trevisan, M. The occurrence of the stilbene piceatannol in some wines from Sicily. Vignevini 2003, 30, 87–94. [Google Scholar]
- Trela, B.C.; Waterhouse, A.L. Resveratrol: Isomeric molar absorptivities and stability. J. Agric. Food Chem. 1996, 44, 1253–1257. [Google Scholar] [CrossRef]
- Vrhovsek, U.; Wendelin, S.; Eder, R. Effects of various vinification techniques on the concentration of cis- and trans-resveratrol and resveratrol glucoside isomers in wine. Am. J. Enol. Viticult. 1997, 48, 214–219. [Google Scholar]
- Threlfall, R.T.; Morris, J.R.; Mauromoustakos, A. Effect of variety, ultraviolet light exposure, and enological methods on the trans-resveratrol level of wine. Am. J. Enol. Viticult. 1999, 50, 57–64. [Google Scholar]
- Mattivi, F.; Nicolini, G. Influence of the winemaking technique on the resveratrol content of wines. L’Enotecnico 1993, 29, 81–88. [Google Scholar]
- Roldan, A.; Palacios, V.; Caro, I.; Perez, L. Evolution of resveratrol and piceid contents during the industrial winemaking process of sherry wine. J. Agric. Food Chem. 2010, 58, 4268–4273. [Google Scholar] [CrossRef] [PubMed]
- Paronetto, L.; Mattivi, F. The resveratrol in oenology and an application of UVC rays to increase its concentration in Amarone wines. L’Enotecnico 1999, 35, 73–81. [Google Scholar]
- Howitz, K.T.; Bitterman, K.J.; Cohen, H.Y.; Lamming, D.W.; Lavu, S.; Wood, J.G.; Zipkin, R.E.; Chung, P.; Kisielewski, A.; Zhang, L.L.; et al. Small molecule activators of sirtuins extend Saccharomyces cerevisiae lifespan. Nature 2003, 425, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.G.; Rogina, B.; Lavu, S.; Howitz, K.; Helfand, S.L.; Tatar, M.; Sinclair, D. Sirtuin activators mimic caloric restriction and delay aging in metazoans. Nature 2004, 430, 686–689. [Google Scholar] [CrossRef] [PubMed]
- Lamming, D.W.; Wood, J.G.; Sinclair, D.A. Small molecules that regulate lifespan: Evidence for xenohormesis. Mol. Microbiol. 2004, 53, 1003–1009. [Google Scholar] [CrossRef] [PubMed]
- North, B.J.; Verdin, E. Sirtuins: Sir2-related NAD-dependent protein deacetylases. Genome Biol. 2004, 5, 224. [Google Scholar] [CrossRef] [PubMed]
- Kaeberlein, M.; McVey, M.; Guarente, L. The SIR2/3/4 complex and SIR2 alone promote longevity in Saccharomyces cerevisiae by two different mechanisms. Genes Dev. 1999, 13, 2570–2580. [Google Scholar] [CrossRef] [PubMed]
- Tissenbaum, H.A.; Guarente, L. Increased dosage of a sir-2 gene extends lifespan in Caenorhabditis elegans. Nature 2001, 410, 227–230. [Google Scholar] [CrossRef] [PubMed]
- Rogina, B.; Helfand, S.L. Sir2 mediates longevity in the fly through a pathway related to calorie restriction. Proc. Natl. Acad. Sci. USA 2004, 101, 15998–16003. [Google Scholar] [CrossRef] [PubMed]
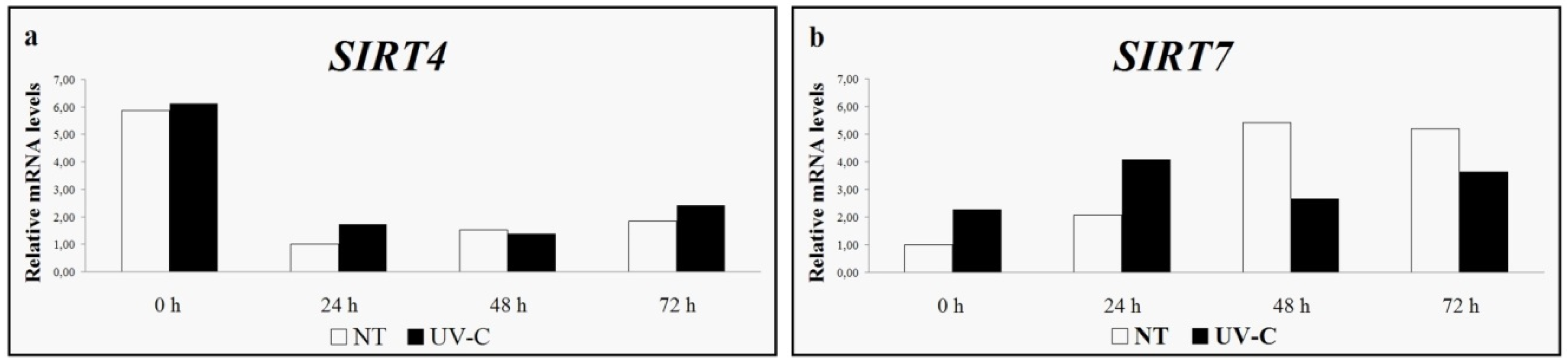
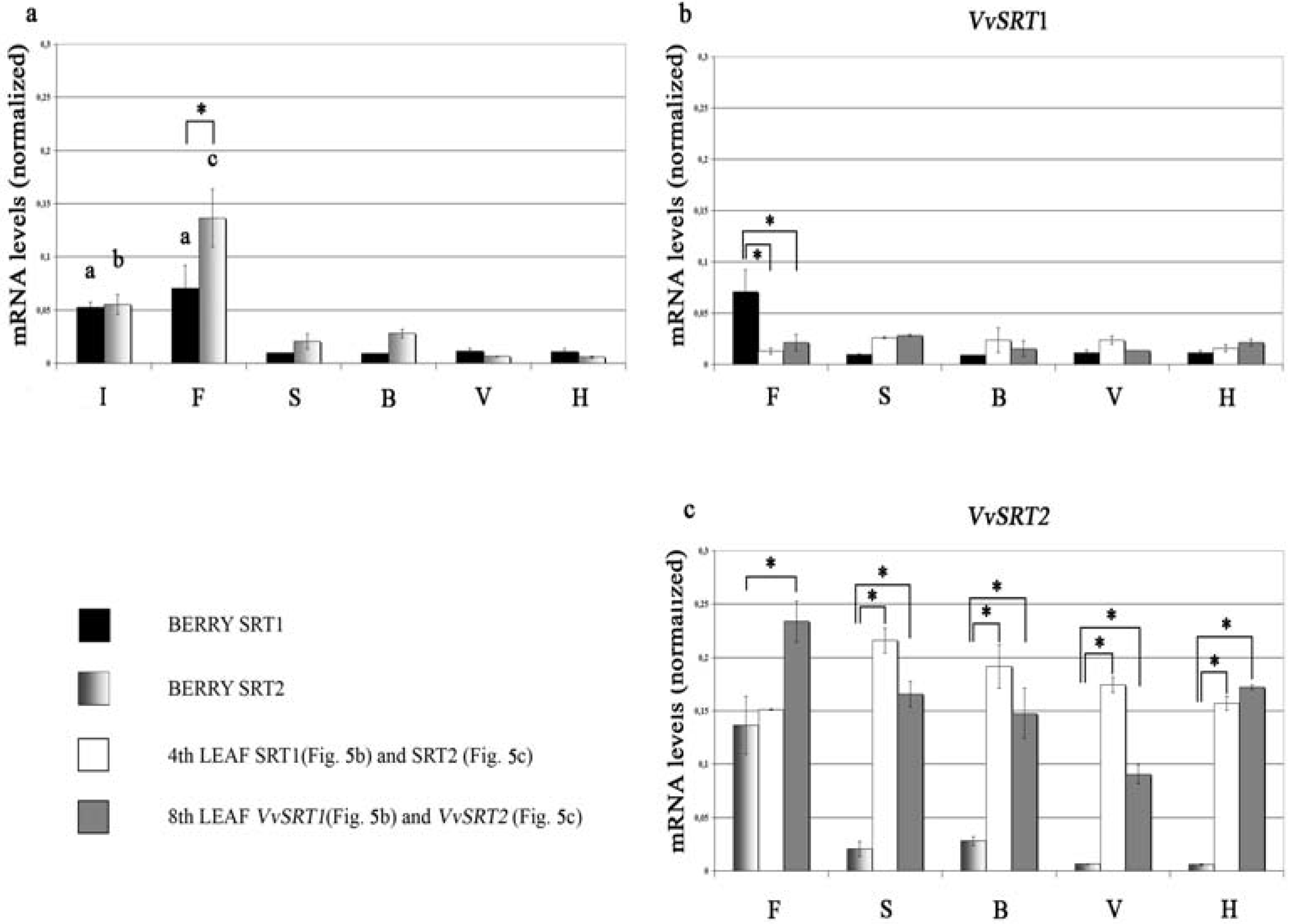
- Busconi, M.; Reggi, S.; Fogher, C.; Bavaresco, L. Evidence of a sirtuin gene family in grapevine (Vitis vinifera L.). Plant Physiol. Biochem. 2009, 47, 650–652. [Google Scholar] [CrossRef] [PubMed]
- Cucurachi, M.; Busconi, M.; Fogher, C.; Hubbard, B.; Sinclair, D.A.; Kovacs, L.G.; Olah, R.; Winterhagen, P.; Perl, A.; Bavaresco, L. Preliminary observations on the role of sirtuin genes in grapevine (Vitis vinifera L.) physiology. Acta Hort. 2014, 1046, 363–369. [Google Scholar] [CrossRef]
- Cucurachi, M.; Busconi, M.; Morreale, G.; Zanetti, A.; Bavaresco, L.; Fogher, C. Characterization and differential expression analysis of complete coding sequence of Vitis vinifera L. sirtuin genes. Plant Physiol. Biochem. 2012, 54, 123–132. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bavaresco, L.; Lucini, L.; Busconi, M.; Flamini, R.; De Rosso, M. Wine Resveratrol: From the Ground Up. Nutrients 2016, 8, 222. https://doi.org/10.3390/nu8040222
Bavaresco L, Lucini L, Busconi M, Flamini R, De Rosso M. Wine Resveratrol: From the Ground Up. Nutrients. 2016; 8(4):222. https://doi.org/10.3390/nu8040222
Chicago/Turabian StyleBavaresco, Luigi, Luigi Lucini, Matteo Busconi, Riccardo Flamini, and Mirko De Rosso. 2016. "Wine Resveratrol: From the Ground Up" Nutrients 8, no. 4: 222. https://doi.org/10.3390/nu8040222
APA StyleBavaresco, L., Lucini, L., Busconi, M., Flamini, R., & De Rosso, M. (2016). Wine Resveratrol: From the Ground Up. Nutrients, 8(4), 222. https://doi.org/10.3390/nu8040222