Using NMR-Based Metabolomics to Evaluate Postprandial Urinary Responses Following Consumption of Minimally Processed Wheat Bran or Wheat Aleurone by Men and Women

 , ,
, , 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants and Study Design
2.2. Preparation and Analysis of the Treatments
2.3. NMR Spectroscopy
2.4. NMR Spectra Pre-Processing
2.5. Data Analysis
3. Results
3.1. Composition of the Treatments
3.2. Participant Characteristics and Compliance
3.3. Metabolomic Analysis of Urine Samples
3.4. Metabolite Identification
3.5. Comparisons between Sexes for Effects of Treatments
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Seal, C.J.; Brownlee, I.A. Whole-grain foods and chronic disease: Evidence from epidemiological and intervention studies. Proc. Nutr. Soc. 2015, 74, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Fardet, A. New hypotheses for the health-protective mechanisms of whole-grain cereals: What is beyond fibre? Nutr. Res. Rev. 2010, 23, 65–134. [Google Scholar] [CrossRef] [PubMed]
- Buri, R.C.; von Reding, W.; Gavin, M.H. Description and characterization of wheat aleurone. Cereal Foods World 2004, 49, 274–282. [Google Scholar]
- Price, R.K.; Keaveney, E.M.; Hamill, L.L.; Wallace, J.M.W.; Ward, M.; Ueland, P.M.; McNulty, H.; Strain, J.J.; Parker, M.J.; Welch, R.W. Consumption of wheat aleurone-rich foods increases fasting plasma betaine and modestly decreases fasting homocysteine and LDL-cholesterol in adults. J. Nutr. 2010, 140, 2153–2157. [Google Scholar] [CrossRef] [PubMed]
- Price, R.K.; Wallace, J.M.W.; Hamill, L.L.; Keaveney, E.M.; Strain, J.J.; Parker, M.J.; Welch, R.W. Evaluation of the effect of wheat aleurone-rich foods on markers of antioxidant status, inflammation and endothelial function in apparently healthy men and women. Br. J. Nutr. 2012, 108, 1644–1651. [Google Scholar] [CrossRef] [PubMed]
- Brennan, L. NMR-based metabolomics: From sample preparation to applications in nutrition research. Prog. Nucl. Mag. Reson. Spectrosc. 2014, 83, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Bertram, H.C.; Knudsen, K.E.B.; Serena, A.; Malmendal, A.; Nielsen, N.C.; Frette, X.C.; Andersen, H.J. NMR-based metabonomic studies reveal changes in the bio-chemical profile of plasma and urine from pigs fed high-fibre rye bread. Br. J. Nutr. 2006, 95, 955–962. [Google Scholar] [CrossRef] [PubMed]
- Fardet, A.; Canlet, C.; Gottardi, G.; Lyan, B.; Llorach, R.; Rémésy, C.; Mazur, A.; Paris, A.; Scalbert, A. Whole-grain and refined wheat flours show distinct metabolic profiles in rats as assessed by a 1H NMR-based metabonomic approach. J. Nutr. 2007, 137, 923–929. [Google Scholar] [PubMed]
- Liu, G.; Xiao, L.; Fang, T.; Cai, Y.; Jia, G.; Zhao, H.; Wang, J.; Chen, X. Pea fiber and wheat bran fiber show distinct metabolic profiles in rats as investigated by a 1H NMR-based metabolomic approach. PLoS ONE 2014, 9, 1–23. [Google Scholar]
- Moazzami, A.A.; Zhang, J.X.; Kamal-Eldin, A.; Aman, P.; Hallmans, G.; Johansson, J.E.; Andersson, S.O. Nuclear Magnetic Resonance-Based Metabolomics Enables Detection of the Effects of a Whole Grain Rye and Rye Bran Diet on the Metabolic Profile of Plasma in Prostate Cancer Patients. J. Nutr. 2011, 141, 2126–2132. [Google Scholar] [CrossRef] [PubMed]
- Moazzami, A.A.; Shrestha, A.; Morrison, D.A.; Poutanen, K.; Mykkänen, H. Metabolomics reveals differences in postprandial responses to breads and fasting metabolic characteristics associated with postprandial insulin demand in postmenopausal women. J. Nutr. 2014, 144, 807–814. [Google Scholar] [CrossRef] [PubMed]
- Pekkinen, J.; Rosa, N.N.; Savolainen, O.I.; Keski-Rahkonen, P.; Mykkänen, H.; Poutanen, K.; Micard, V.; Hanhineva, K. Disintegration of wheat aleurone structure has an impact on the bioavailability of phenolic compounds and other phytochemicals as evidenced by altered urinary metabolite profile of diet-induced obese mice. Nutr. Metab. 2014, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Ross, A.B.; Pere-Trépat, E.; Montoliu, I.; Martin, F.P.J.; Collino, S.; Moco, S.; Godin, J.P.; Cléroux, M.; Guy, P.A.; Breton, I.; et al. Whole-grain-rich diet reduces urinary excretion of markers of protein catabolism and gut microbiota metabolism in healthy men after one week. J. Nutr. 2013, 143, 766–733. [Google Scholar] [CrossRef] [PubMed]
- Ross, A.B. Whole grains beyond fibre: What can metabolomics tell us about mechanisms? Proc. Nutr. Soc. 2014, 74, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Keaveney, E.M.; Price, R.K.; Hamill, L.L.; Wallace, J.M.; McNulty, H.; Ward, M.; Strain, J.J.; Ueland, P.M.; Molloy, A.M.; Piironen, V.; et al. Postprandial plasma betaine and other methyl donor-related responses after consumption of minimally processed wheat bran or wheat aleurone, or wheat aleurone incorporated into bread. Br. J. Nutr. 2015, 113, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Hamill, L.L.; Keaveney, E.M.; Price, R.K.; Wallace, J.M.W.; Strain, J.J.; Welch, R.W. Absorption of ferulic acid in human subjects after consumption of wheat-bran and wheat-aleurone fractions. Proc. Nutr. Soc. 2009, 67, E255. [Google Scholar] [CrossRef]
- Corol, D.I.; Ravel, C.; Raksegi, M.; Bedo, Z.; Charmet, G.; Beale, M.H.; Shewry, P.R.; Ward, J.L. Effects of genotype and environment on the contents of betaine, choline, and trigonelline in cereal grains. J. Agric. Food Chem. 2012, 60, 5471–5481. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Shewry, P.R.; Ward, J.L. Phenolic acids in wheat varieties in the Healthgrain diversity screen. J. Agric. Food Chem. 2008, 56, 9732–9739. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.W.; Cagney, G.; Cartwright, H.M. SpecAlign—Processing and alignment of mass spectra datasets. Bioinformatics 2005, 21, 2088–2090. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Aluko, R.E.; Corke, H. Partial least-squares regression study of the effects of wheat flour composition, protein and starch quality characteristics on oil content of steamed-and-fried instant noodles. J. Cereal Sci. 2006, 44, 117–126. [Google Scholar] [CrossRef]
- Wold, S.; Johansson, E.; Cocchi, M. PLS—Partial least squares projections to latent structures. In 3D QSAR in Drug Design, Theory, Methods, and Applications; Kubinyi, H., Ed.; ESCOM Science Publishers: Leiden, Holland, 1993; pp. 523–550. [Google Scholar]
- Vander Heiden, M.G.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg effect: The metabolic requirements of cell proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [PubMed]
- DeBerardinis, R.J.; Lum, J.L.; Hatzivassiliou, G.; Thompson, C.B. The Biology of Cancer: Metabolic Reprogramming Fuels Cell Growth and Proliferation. Cell Metab. 2008, 7, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Castellano, G.; Dias, C.S.B.; Foerster, B.; Li, L.M.; Covolan, R.J.M. NAA and NAAG variation in neuronal activation during visual stimulation. Braz. J. Med. Biol. Res. 2012, 45, 1031–1036. [Google Scholar] [CrossRef] [PubMed]
- Moffett, J.R.; Ross, B.; Arun, P.; Madhavarao, C.N.; Namboodiri, A.M. N-Acetylaspartate in the CNS: From neurodiagnostics to neurobiology. Prog. Neurobiol. 2007, 81, 89–131. [Google Scholar] [CrossRef] [PubMed]
- Benarroch, E.E. N-Acetylaspartate and N-acetylaspartylglutamate. Neurology 2008, 70, 1353–1357. [Google Scholar] [CrossRef] [PubMed]
- Clark, J.B. N-acetyl aspartate: A marker for neuronal loss or mitochondrial dysfunction. Dev. Neurosci. 1998, 20, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Zeisel, S.H. Metabolic crosstalk between choline/1-carbon metabolism and energy homeostasis. Clin. Chem. Lab. Med. 2013, 51, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Lee, I. Betaine is a positive regulator of mitochondrial respiration. Biochem. Biophys. Res. Commun. 2015, 456, 621–625. [Google Scholar] [CrossRef] [PubMed]
- Lever, M.; Slow, S. The clinical significance of betaine, an osmolyte with a key role in methyl group metabolism. Clin. Biochem. 2010, 43, 732–744. [Google Scholar] [CrossRef] [PubMed]
- Finkelstein, J.D.; Martin, J.J.; Harris, B.J.; Kyle, W.E. Regulation of hepatic betaine-homocysteine methyltransferase by dietary betaine. J. Nutr. 1983, 113, 519–521. [Google Scholar] [PubMed]
- Lees, H.J.; Swann, J.R.; Wilson, I.D.; Nicholson, J.K.; Holmes, E. Hippurate: The natural history of a mammalian-microbial cometabolite. J. Proteome Res. 2013, 12, 1527–1546. [Google Scholar] [CrossRef] [PubMed]
- Gavaghan McKee, C.L.; Wilson, I.D.; Nicholson, J.K. Metabolic phenotyping of nude and normal (Alpk:ApfCD, C57BL10J) mice. J. Proteome Res. 2006, 5, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.E.; Lenz, E.M.; Evans, J.A.; Wilson, I.D.; Granger, J.H.; Plumb, R.S.; Stumpf, C.L. A combined 1H NMR and HPLC-MS-based metabonomic study of urine from obese (fa/fa) Zucker and normal Wistar-derived rats. J. Pharm. Biomed. Anal. 2005, 38, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Siqueira, M.E.; Paiva, M.J. Hippuric acid in urine: Reference values. Rev. Saúde Pública 2002, 36, 723–727. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Yang, S.; Cai, S.; Dong, J.; Li, X.; Chen, Z. Identification of biochemical changes in lactovegetarian urine using 1H NMR spectroscopy and pattern recognition. Anal. Bioanal. Chem. 2010, 396, 1451–1463. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, H.; Nicholson, J.K.; Hylands, P.J.; Sampson, J.; Holmes, E. A metabonomic strategy for the detection of the metabolic effects of chamomile (Matricaria recutita L.) ingestion. J. Agric. Food Chem. 2005, 53, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Lenz, E.M.; Bright, J.; Wilson, I.D.; Morgan, S.R.; Nash, A.F. A 1H NMR-based metabonomic study of urine and plasma samples obtained from healthy human subjects. J. Pharm. Biomed. Anal. 2003, 33, 1103–1115. [Google Scholar] [CrossRef]
- Rasmussen, L.G.; Savorani, F.; Larsen, T.M.; Dragsted, L.O.; Astrup, A.; Engelsen, S.B. Standardization of factors that influence human urine metabolomics. Metabolomics 2011, 7, 71–83. [Google Scholar] [CrossRef]
- Walsh, M.C.; Brennan, L.; Malthouse, J.P.G.; Roche, H.M.; Gibney, M.J. Effect of acute dietary standardization on the urinary, plasma, and salivary metabolomic profiles of healthy humans. Am. J. Clin. Nutr. 2006, 84, 531–539. [Google Scholar] [PubMed]
- Walsh, M.C.; Brennan, L.; Pujos-Guillot, E.; Sébédio, J.L.; Scalbert, A.; Fagan, A.; Higgins, D.G.; Gibney, M.J. Influence of acute phytochemical intake on human urinary metabolomic profiles. Am. J. Clin. Nutr. 2007, 86, 1687–1693. [Google Scholar] [PubMed]
- Garg, R.; Brennan, L.; Price, R.K.; Wallace, J.M.W.; Strain, J.J.; Gibney, M.J.; Welch, R.W. Evaluating the potential of the wheat aleurone fraction in humans using 1H NMR-based metabolomic analysis. Proc. Nutr. Soc. 2010, 69, E348. [Google Scholar] [CrossRef]
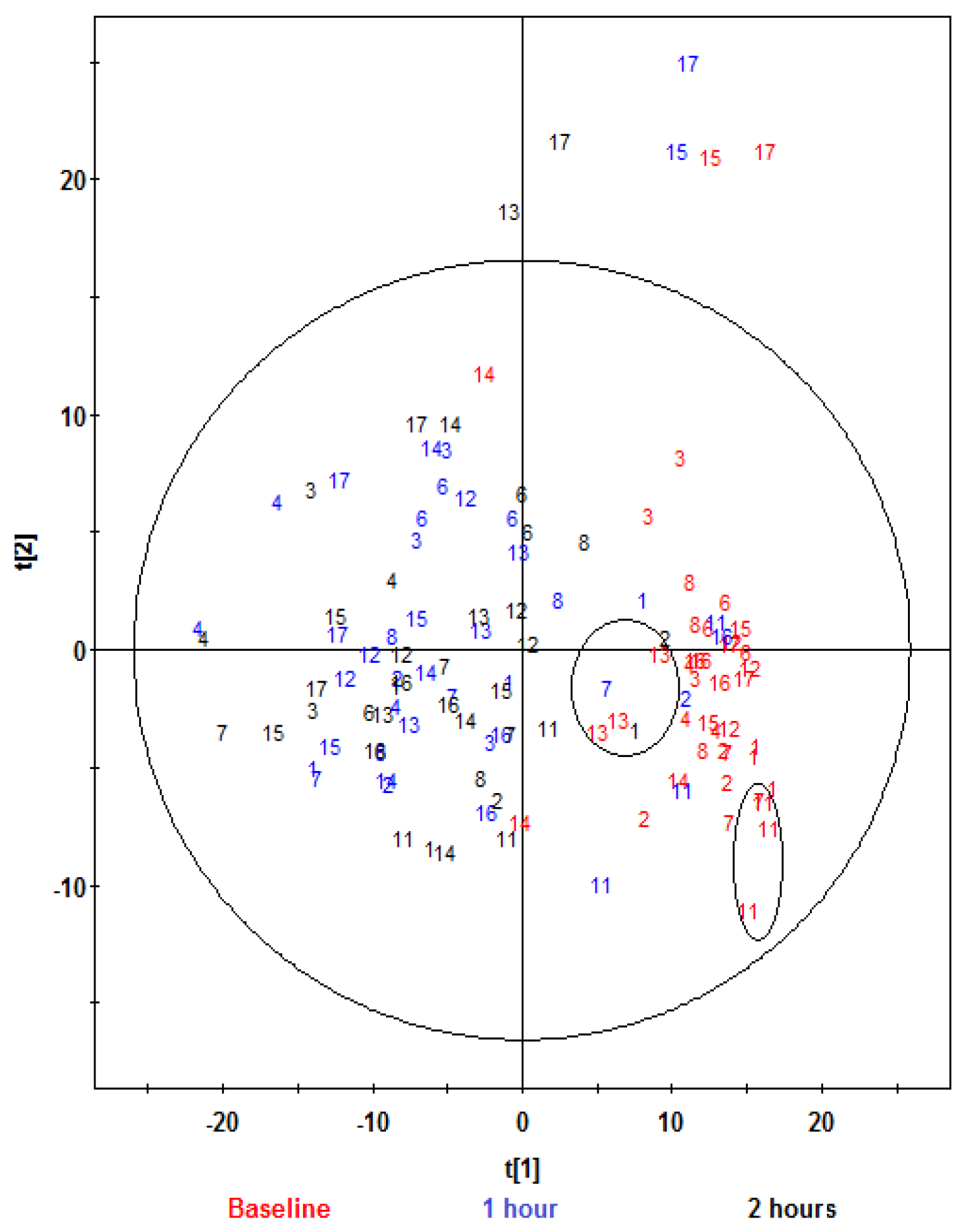
 , aleurone;
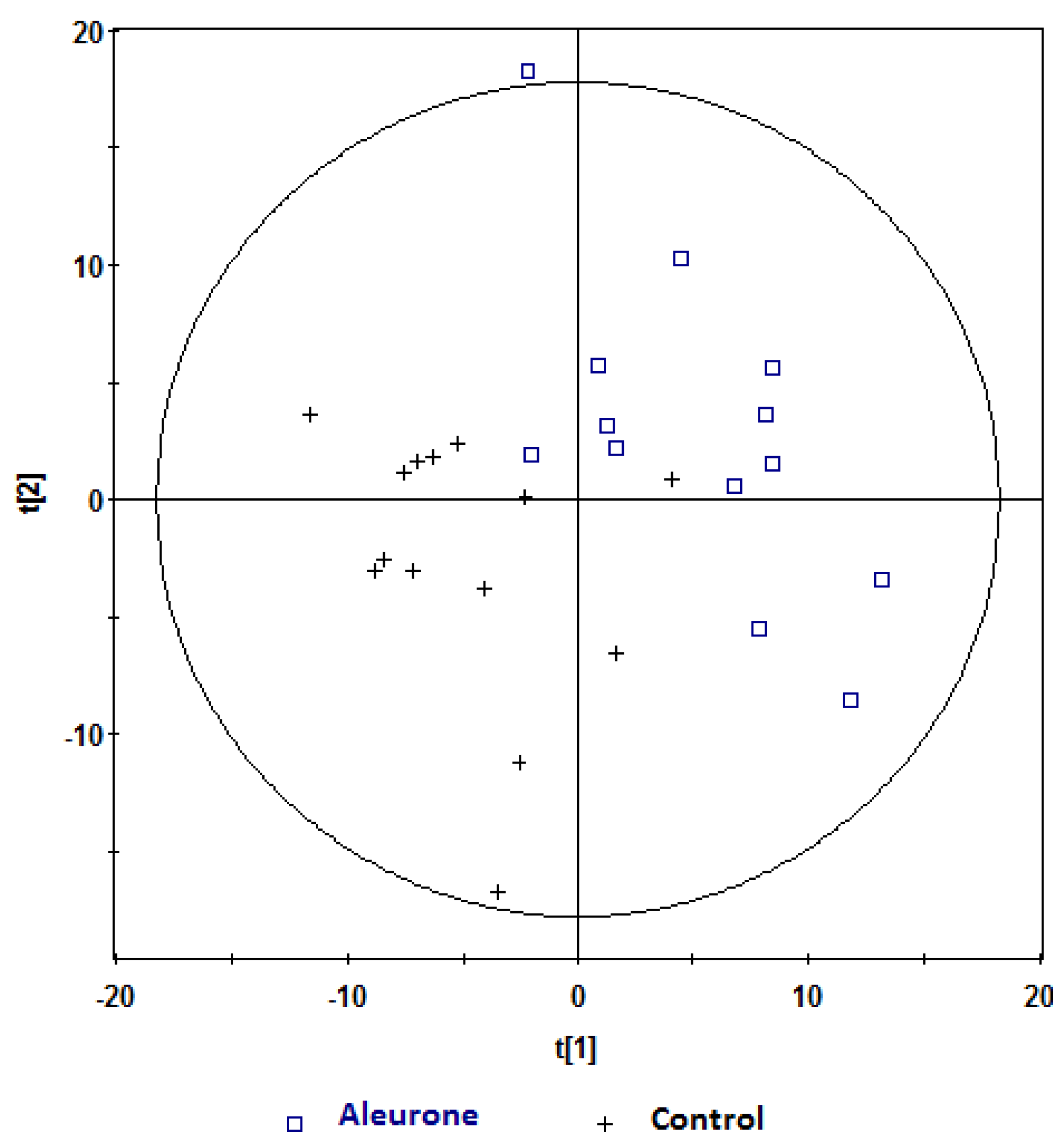
, aleurone;  , control. Variables (K) = 181. PLS component 1: R2 = 0.196, Q2 = 0.315; PLS component 2: R2 = 0.18, Q2 = 0.117. The ellipse represents 95% confidence region of the model based on Hotelling T2.
, aleurone; , control. Variables (K) = 181. PLS component 1: R2 = 0.196, Q2 = 0.315; PLS component 2: R2 = 0.18, Q2 = 0.117. The ellipse represents 95% confidence region of the model based on Hotelling T2.
, control. Variables (K) = 181. PLS component 1: R2 = 0.196, Q2 = 0.315; PLS component 2: R2 = 0.18, Q2 = 0.117. The ellipse represents 95% confidence region of the model based on Hotelling T2.
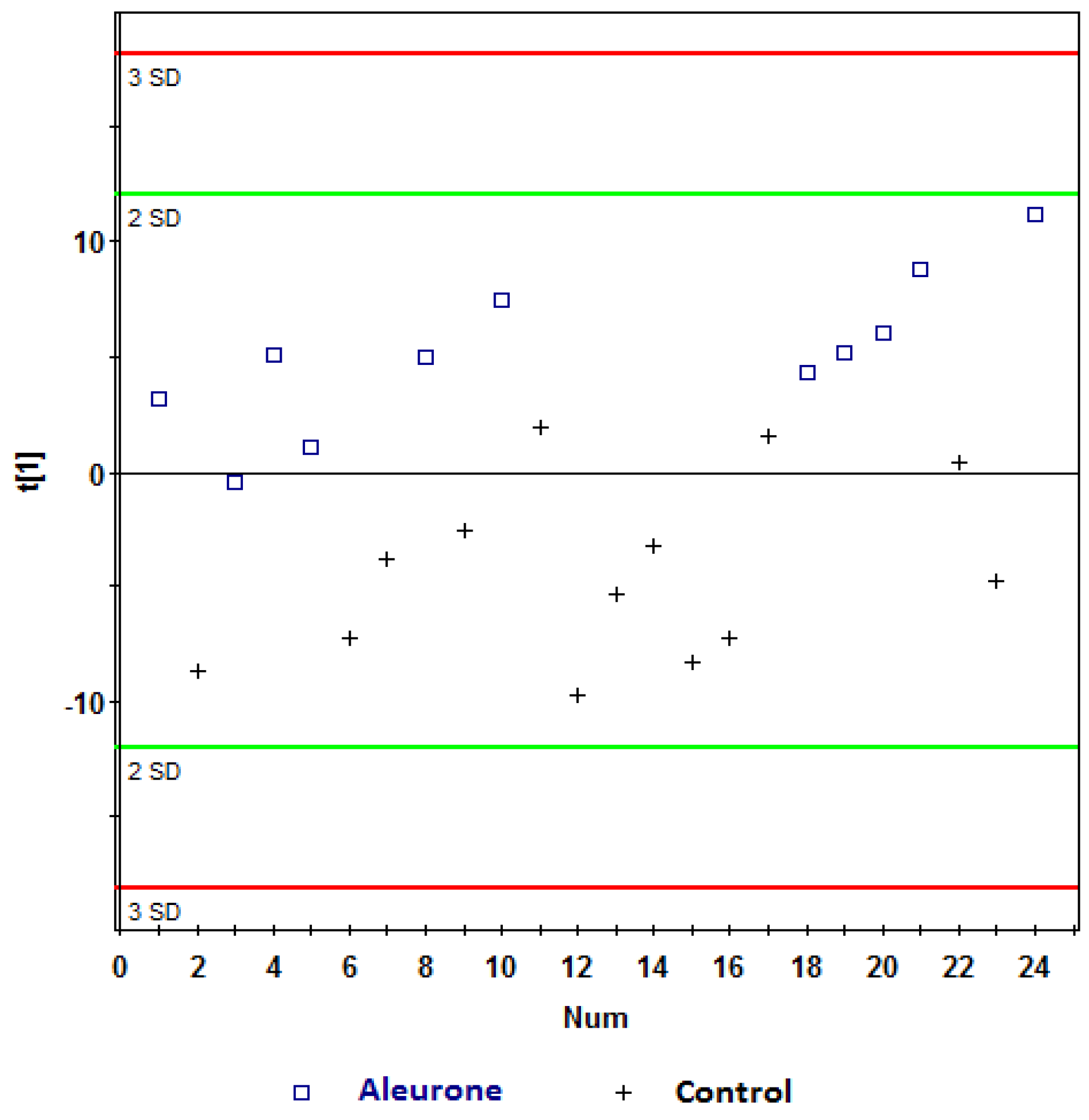
, aleurone; , control. Variables (K) = 181. PLS component 1: R2 = 0.196, Q2 = 0.315; PLS component 2: R2 = 0.18, Q2 = 0.117. The ellipse represents 95% confidence region of the model based on Hotelling T2.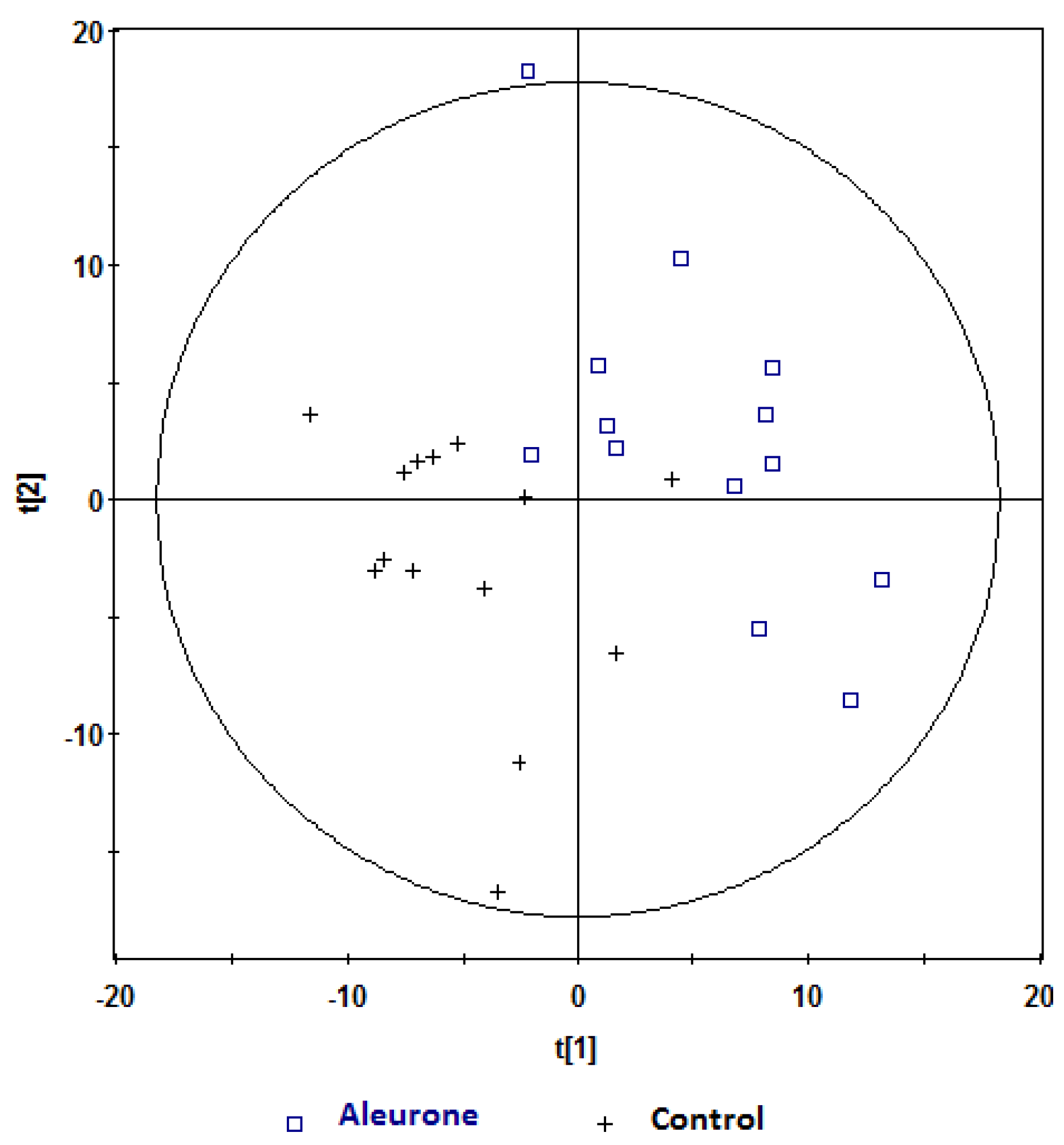 , aleurone; , control. Variables (K) = 181. PLS component 1: R2 = 0.148, Q2 = 0.339.
, aleurone; , control. Variables (K) = 181. PLS component 1: R2 = 0.148, Q2 = 0.339.
, aleurone; , control. Variables (K) = 181. PLS component 1: R2 = 0.148, Q2 = 0.339.
, aleurone; , control. Variables (K) = 181. PLS component 1: R2 = 0.148, Q2 = 0.339.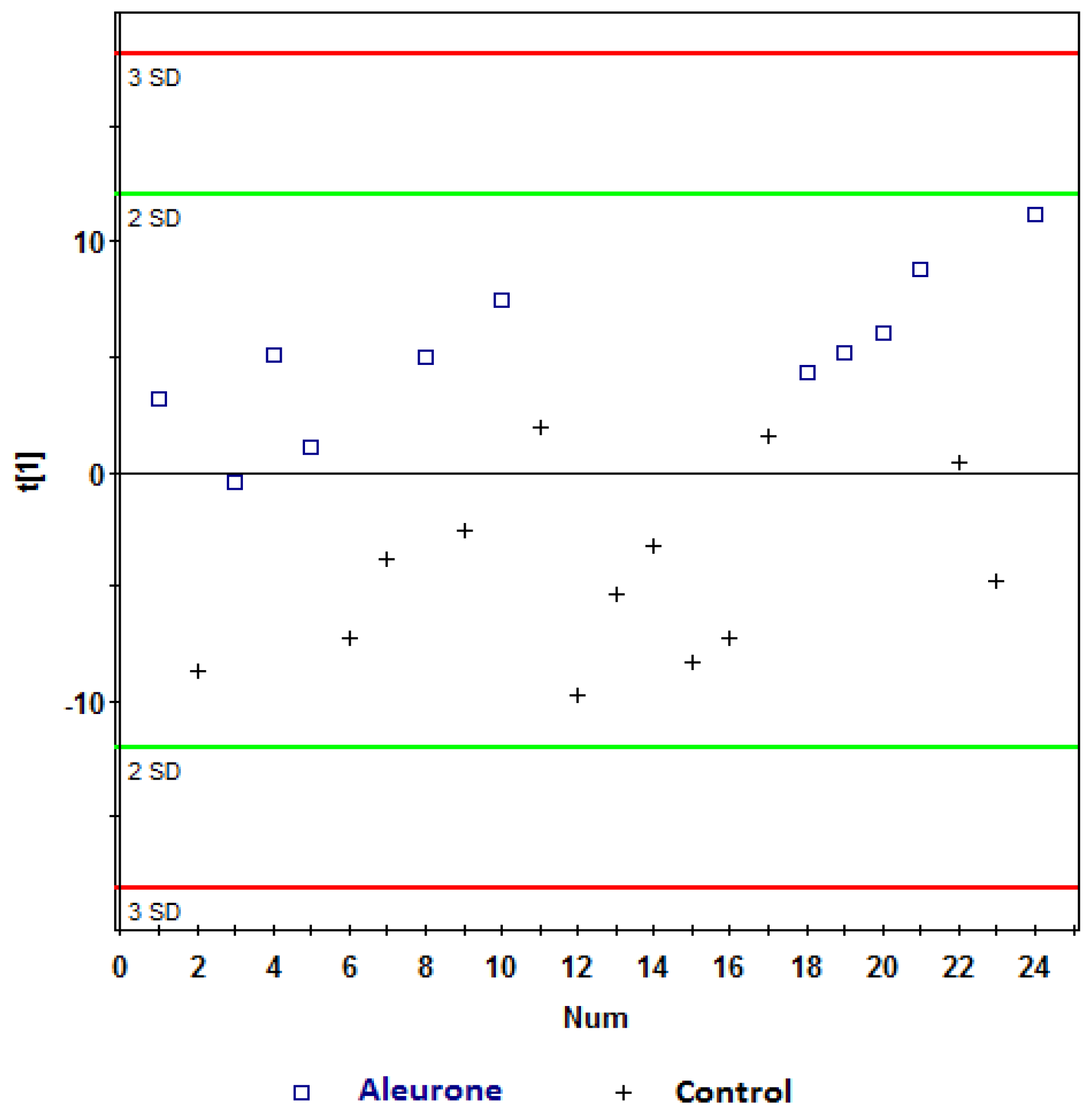
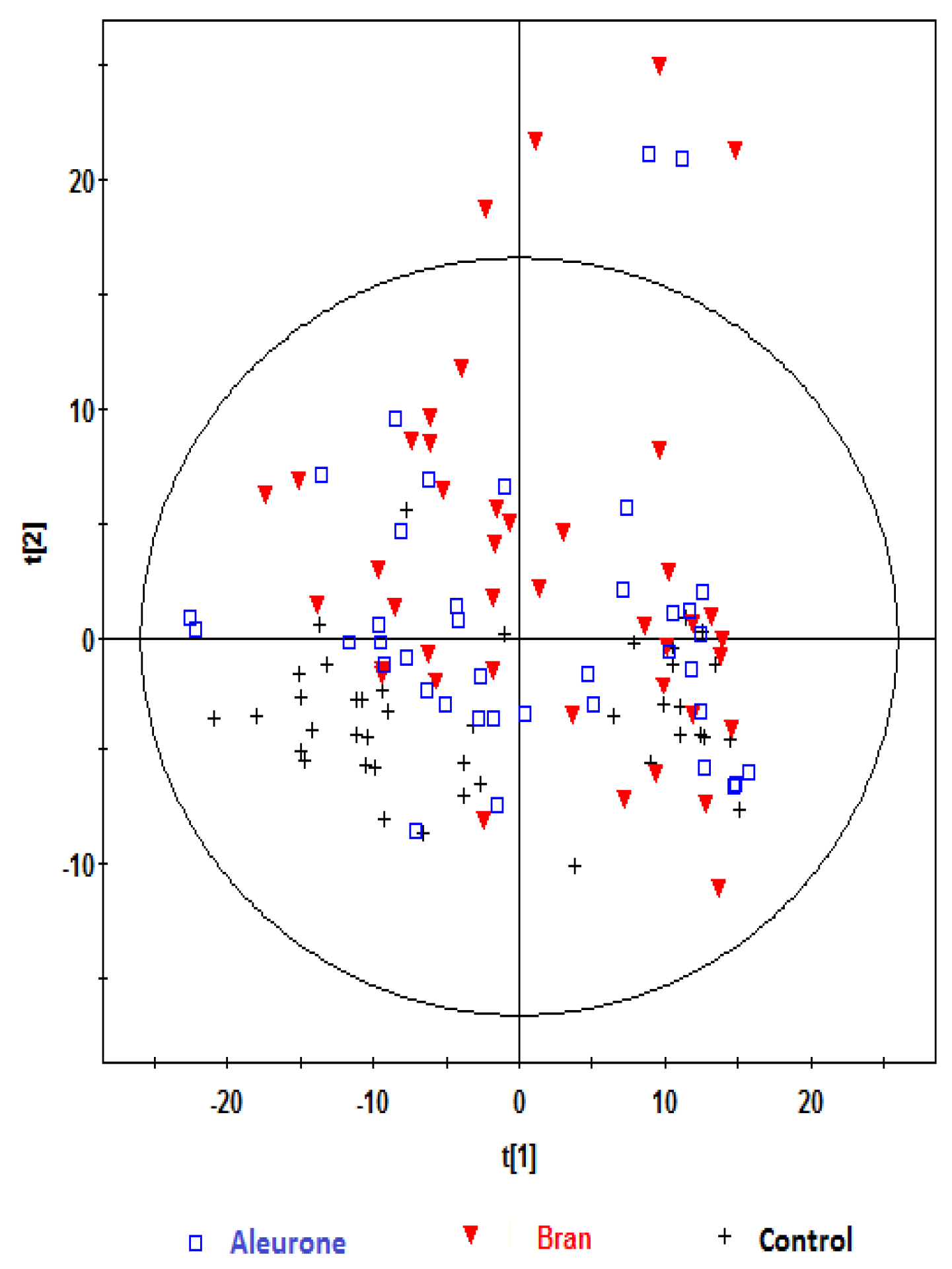
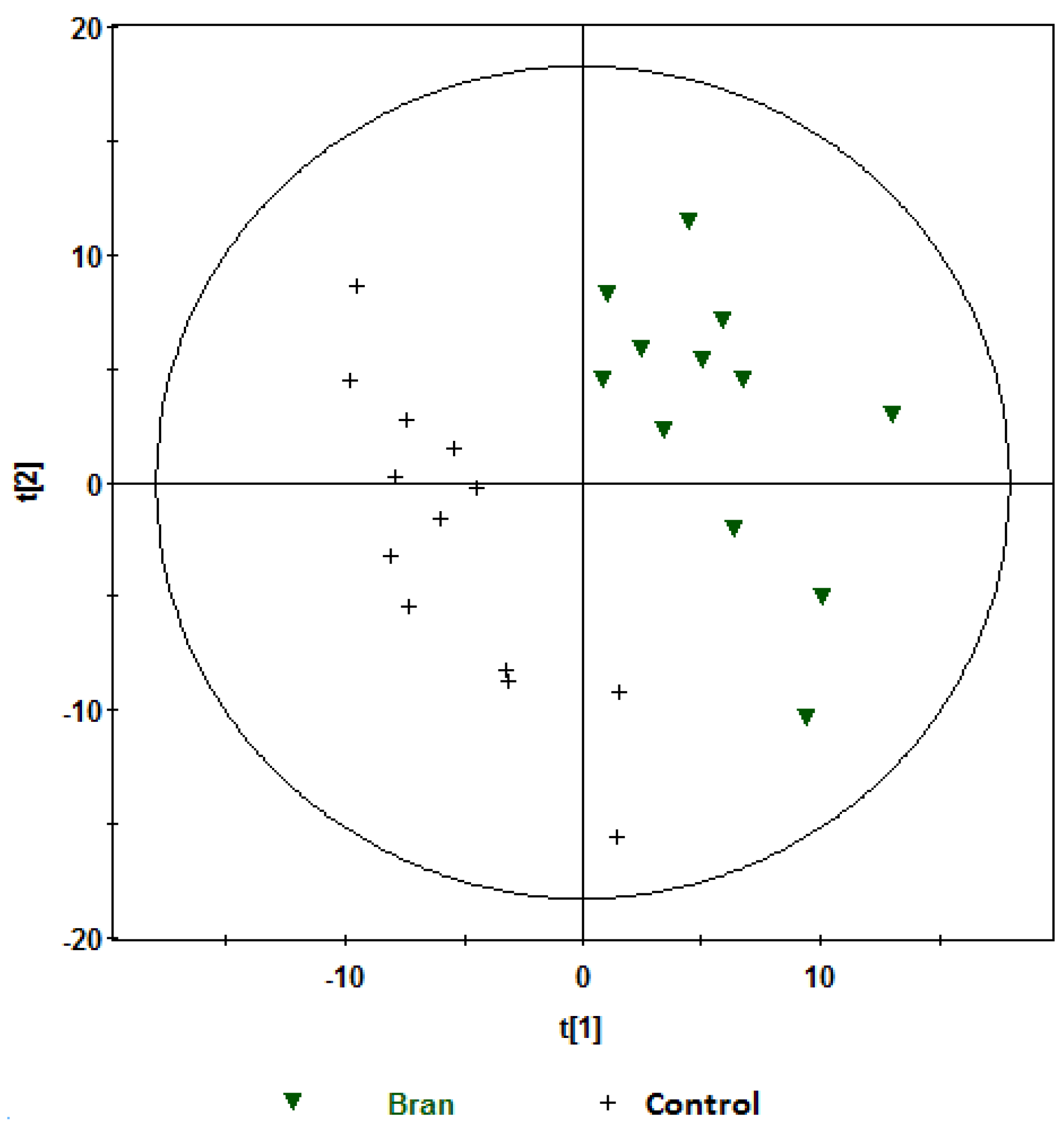
 , bran; , control. Variables (K) = 181. PLS component 1: R2 = 0.204, Q2 = 0.51; PLS component 2: R2 = 0.176, Q2 = 0.111. The ellipse represents 95% confidence region of the model based on Hotelling T2.
, bran; , control. Variables (K) = 181. PLS component 1: R2 = 0.204, Q2 = 0.51; PLS component 2: R2 = 0.176, Q2 = 0.111. The ellipse represents 95% confidence region of the model based on Hotelling T2.
, bran; , control. Variables (K) = 181. PLS component 1: R2 = 0.204, Q2 = 0.51; PLS component 2: R2 = 0.176, Q2 = 0.111. The ellipse represents 95% confidence region of the model based on Hotelling T2.
, bran; , control. Variables (K) = 181. PLS component 1: R2 = 0.204, Q2 = 0.51; PLS component 2: R2 = 0.176, Q2 = 0.111. The ellipse represents 95% confidence region of the model based on Hotelling T2.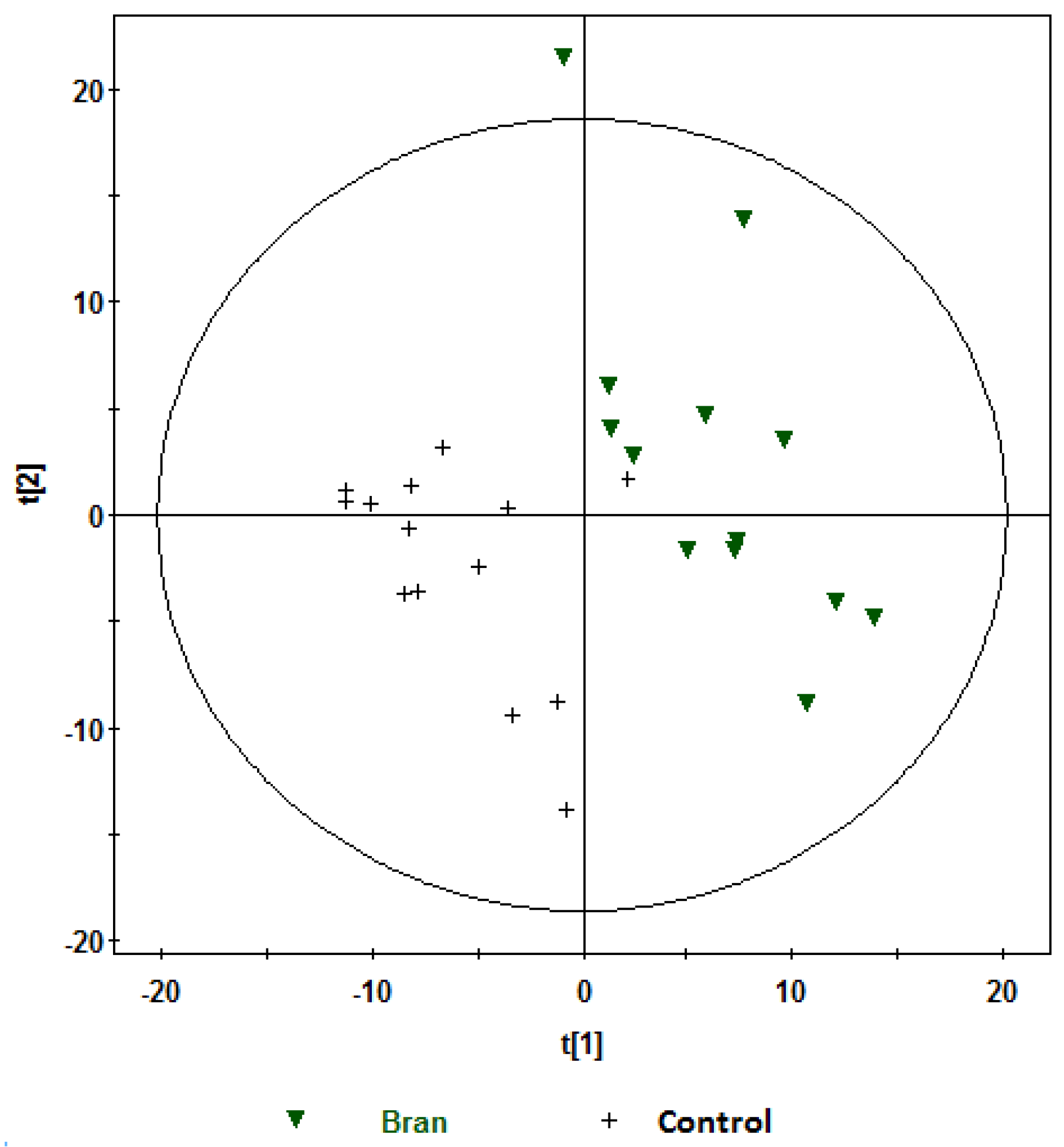 , bran; , control. Variables (K) = 181. PLS component 1: R2 = 0.179, Q2 = 0.379; PLS component 2: R2 = 0.168, Q2 = 0.327. The ellipse represents 95% confidence region of the model based on Hotelling T2.
, bran; , control. Variables (K) = 181. PLS component 1: R2 = 0.179, Q2 = 0.379; PLS component 2: R2 = 0.168, Q2 = 0.327. The ellipse represents 95% confidence region of the model based on Hotelling T2.
, bran; , control. Variables (K) = 181. PLS component 1: R2 = 0.179, Q2 = 0.379; PLS component 2: R2 = 0.168, Q2 = 0.327. The ellipse represents 95% confidence region of the model based on Hotelling T2.
, bran; , control. Variables (K) = 181. PLS component 1: R2 = 0.179, Q2 = 0.379; PLS component 2: R2 = 0.168, Q2 = 0.327. The ellipse represents 95% confidence region of the model based on Hotelling T2.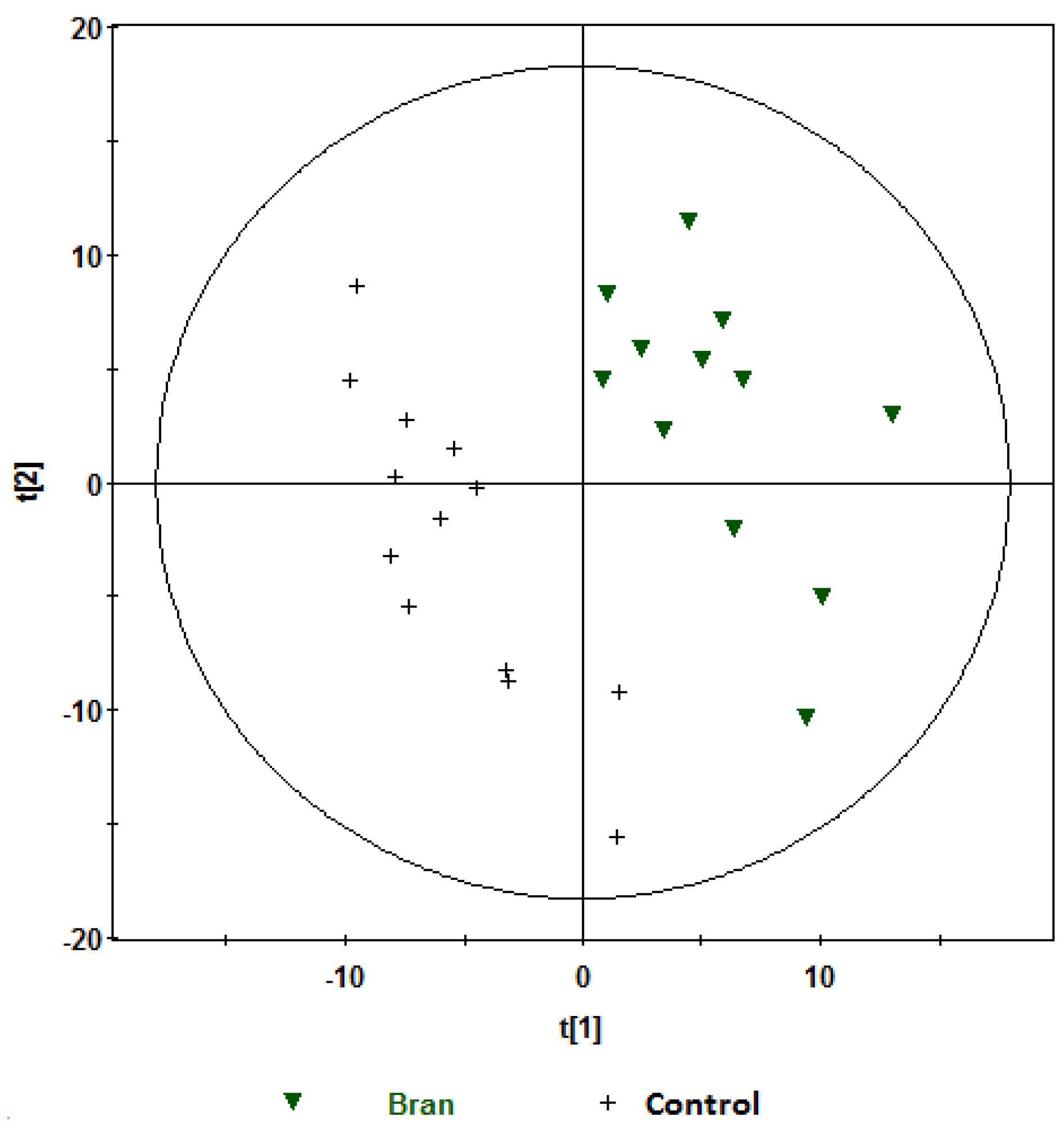

{kind=link}
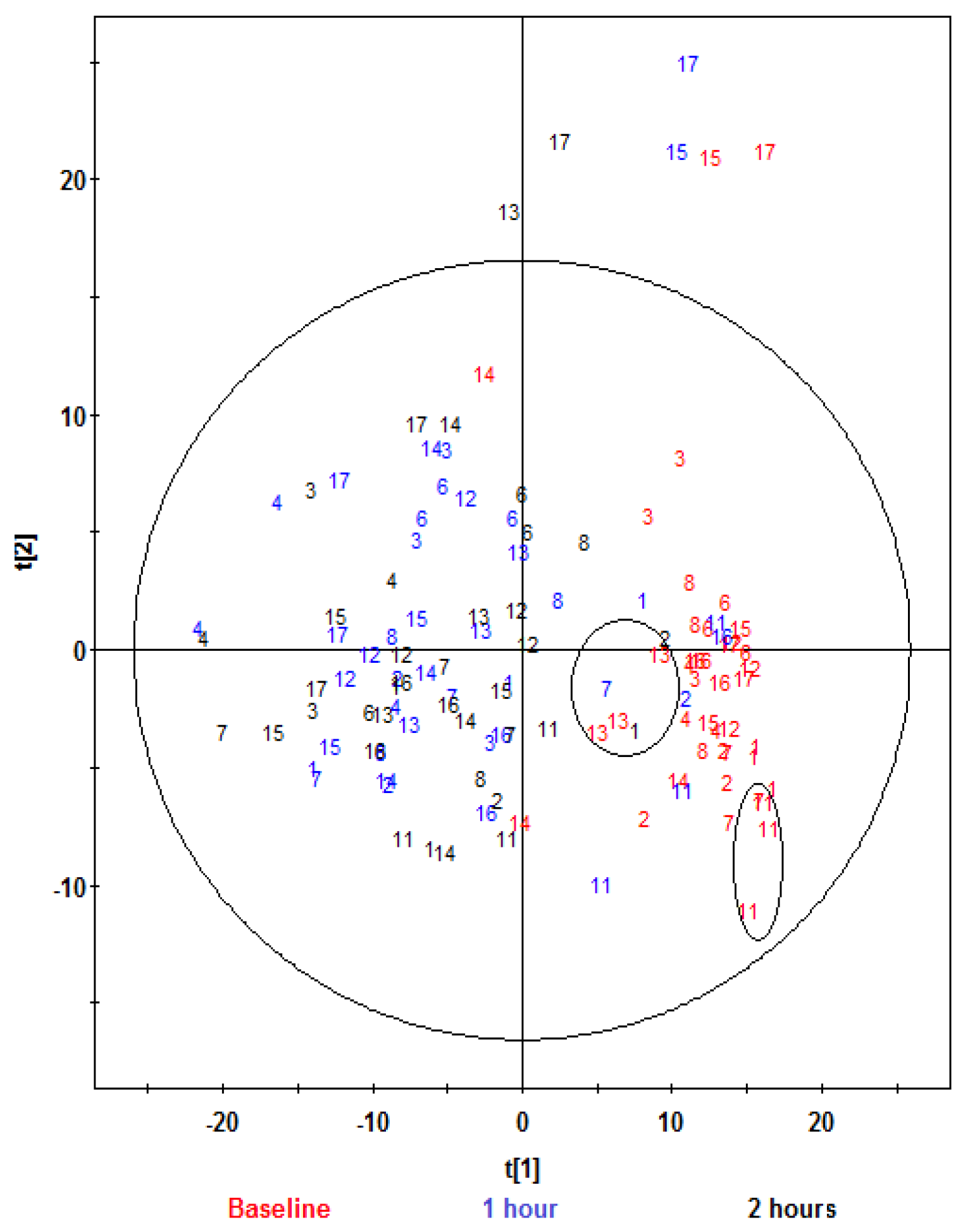{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aleurone | Bran | Control | |
|---|---|---|---|
| Ingredients (g per portion) a | |||
| Wheat bran b | - | 50.0 | - |
| Wheat aleurone b | 50.0 | - | - |
| Wheat fiber c | 3.6 | - | 27.0 |
| Wheat protein d | - | 2.3 | 8.9 |
| Wheat starch e | - | 0.3 | 5.6 |
| Vegetable fat f | - | 0.6 | 2.0 |
| Sugar g | 2.5 | 2.5 | 2.5 |
| Composition per portion | |||
| Energy (kJ) | 352 | 348 | 336 |
| Carbohydrate (g) | 8.1 | 8.0 | 7.4 |
| Protein (g) | 8.9 | 8.7 | 8.4 |
| Fat (g) | 2.0 | 2.0 | 2.1 |
| Fiber (g) | 27 | 27 | 25 |
| Betaine (mg) | 515 | 301 | 31 |
| Total phenolic acids (mg) h | 213 | 162 | 1.5 |
| Total ferulic acid (mg) i | 177 | 124 | 0.9 |
| Free ferulic acid (mg) | 2.2 | 1.2 | 0.0 |
| Treatments | Time-Point (h) | No. of Components | a R2 | b Q2 | Cross Validation (%) c |
|---|---|---|---|---|---|
| Aleurone vs. control | 1 | 2 | 0.377 | 0.395 | 74 |
| Aleurone vs. control | 2 | 1 | 0.148 | 0.339 | 71 |
| Bran vs. control | 1 | 2 | 0.380 | 0.564 | 93 |
| Bran vs. control | 2 | 2 | 0.347 | 0.582 | 93 |
| Chemical Shift (ppm) a | Discriminating Metabolites | Percentage (%) Change in Metabolite Concentration with Respect to Control | |||
|---|---|---|---|---|---|
| Aleurone (1 h) | Aleurone (2 h) | Bran (1 h) | Bran (2 h) | ||
| 1.32 | Lactate | 97 | 109 | 94 | 187 |
| 3.24 | Betaine | −47 | −43 | −27 | −27 |
| 1.52 | Alanine | 43 | 40 | 52 | 61 |
| 2.16, 2.20 | NAA/NAAG b | 65 | 59 | 78 | 110 |
| 2.52, 2.56, 2.64, 2.68 | Citrate | 24 | - | - | - |
| 7.56, 7.84 | Hippurate | - | 35 | - | - |
| 3.48, 3.52, 3.60 | Unknown | - | - | 66 | - |
| 1.28 | 3-hydroxyisovalerate | - | - | 69 | 70 |
| 1.36 | 2-hydroxyisobutyrate | - | - | - | 27 |
| 3.68 | Unknown | - | - | 18 | 34 |
| 3.88 | Unknown | - | - | 25 | 41 |
| Treatment | Timepoint (h) | Females (F) vs Males (M) | No. of Components | a R2 | b Q2 | Cross Validation (%) c | Discriminating Metabolites d |
|---|---|---|---|---|---|---|---|
| None | 0 | F vs. M | 2 | 0.236 | 0.721 | 95 | In females: citrate↑, hippurate↑, betaine↓ |
| Aleurone | 1 | F vs. M | 3 | 0.567 | 0.903 | 92 | In females: citrate↑, hippurate↑, betaine↓ |
| Aleurone | 2 | F vs. M | 2 | 0.454 | 0.717 | 72 | In females: citrate↑, hippurate↑ |
| Bran | 1 | F vs. M | 0 | - | - | - | - |
| Bran | 2 | F vs. M | 1 | 0.194 | 0.287 | Not validated | In females: citrate↑, lactate↑ |
| Control | 1 | F vs. M | 2 | 0.372 | 0.304 | 61 | In females: hippurate↑, betaine↓ |
| Control | 2 | F vs. M | 2 | 0.484 | 0.587 | 83 | In females: hippurate↑, citrate↑, betaine↓ |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garg, R.; Brennan, L.; Price, R.K.; Wallace, J.M.W.; Strain, J.J.; Gibney, M.J.; Shewry, P.R.; Ward, J.L.; Garg, L.; Welch, R.W. Using NMR-Based Metabolomics to Evaluate Postprandial Urinary Responses Following Consumption of Minimally Processed Wheat Bran or Wheat Aleurone by Men and Women. Nutrients 2016, 8, 96. https://doi.org/10.3390/nu8020096
Garg R, Brennan L, Price RK, Wallace JMW, Strain JJ, Gibney MJ, Shewry PR, Ward JL, Garg L, Welch RW. Using NMR-Based Metabolomics to Evaluate Postprandial Urinary Responses Following Consumption of Minimally Processed Wheat Bran or Wheat Aleurone by Men and Women. Nutrients. 2016; 8(2):96. https://doi.org/10.3390/nu8020096
Chicago/Turabian StyleGarg, Ramandeep, Lorraine Brennan, Ruth K. Price, Julie M. W. Wallace, J. J. Strain, Mike J. Gibney, Peter R. Shewry, Jane L. Ward, Lalit Garg, and Robert W. Welch. 2016. "Using NMR-Based Metabolomics to Evaluate Postprandial Urinary Responses Following Consumption of Minimally Processed Wheat Bran or Wheat Aleurone by Men and Women" Nutrients 8, no. 2: 96. https://doi.org/10.3390/nu8020096
APA StyleGarg, R., Brennan, L., Price, R. K., Wallace, J. M. W., Strain, J. J., Gibney, M. J., Shewry, P. R., Ward, J. L., Garg, L., & Welch, R. W. (2016). Using NMR-Based Metabolomics to Evaluate Postprandial Urinary Responses Following Consumption of Minimally Processed Wheat Bran or Wheat Aleurone by Men and Women. Nutrients, 8(2), 96. https://doi.org/10.3390/nu8020096