
American Basil, Ocimum americanum, Has Neuroprotective Properties in the Aging Process
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Preparation of the Ethanol Extract
2.2. Animals
2.3. Ex Vivo Experiments
2.3.1. Animals, Preparation, and Incubation of Hippocampal Slices
2.3.2. Cellular Viability
2.3.3. Cellular Damage
2.4. In Vivo Acute Experiments
2.4.1. Animals and Treatment
2.4.2. Inhibitory Avoidance Task
2.4.3. Tissue Preparation
2.4.4. Reactive Species Levels
2.4.5. Thiobarbituric Acid Reactive Substances (TBARS)
2.4.6. Acetylcholinesterase Activity
2.5. In Vivo Chronic Experiments
2.5.1. Animals and EEOA Supplementation
2.5.2. Tissue Preparation
2.5.3. Measurement of Cytokines, TNF-α, and IL-1β Content
2.6. Protein Determination
2.7. Statistical Analysis
3. Results
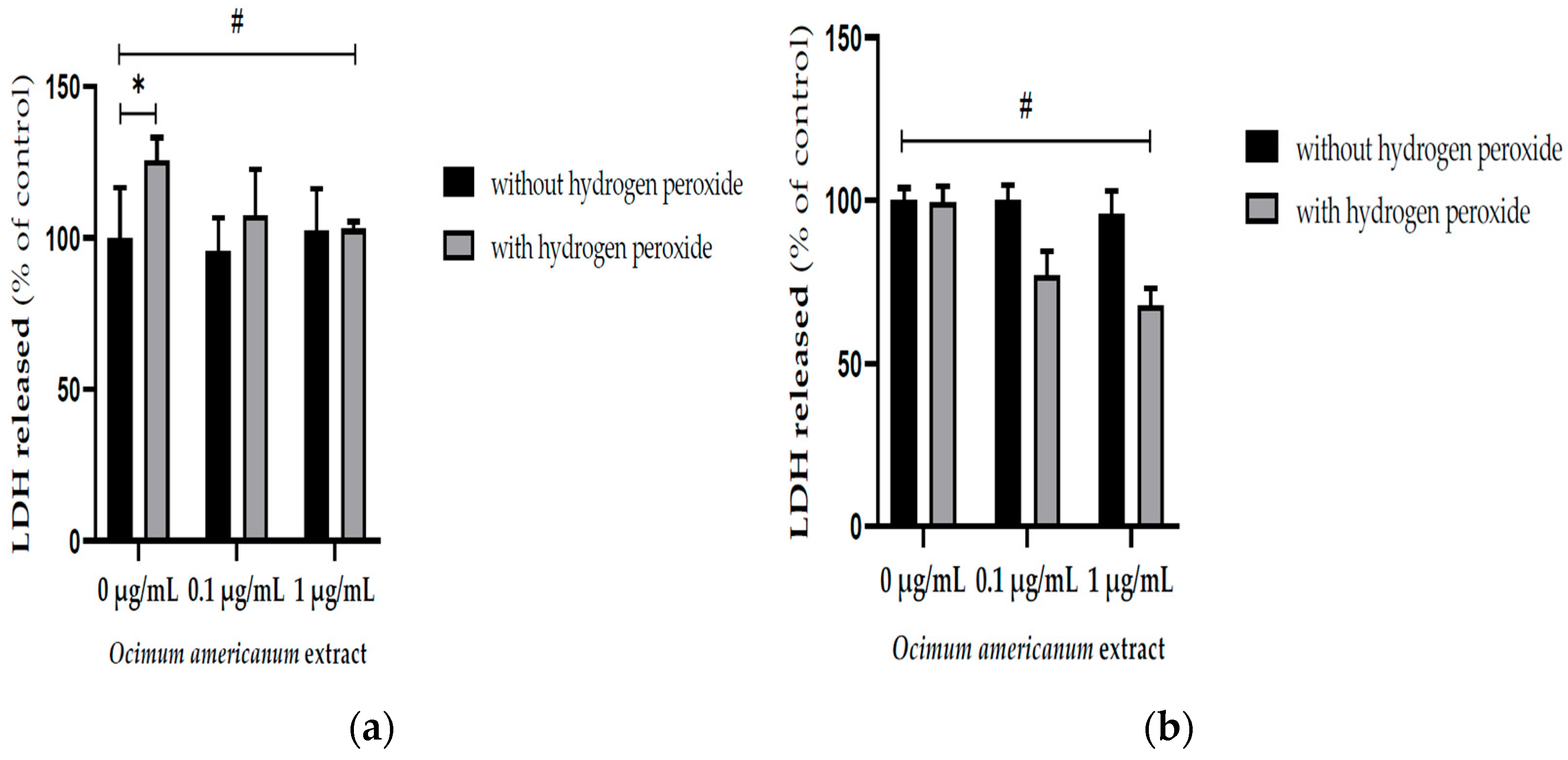
3.1. Ex Vivo Neuroprotective Effect of Ocimum Americanum Extract in Young Adult and Late Middle-Aged Rats
3.2. Effect of Acute Treatment with Ocimum Americanum Extract in Young Adult Rats
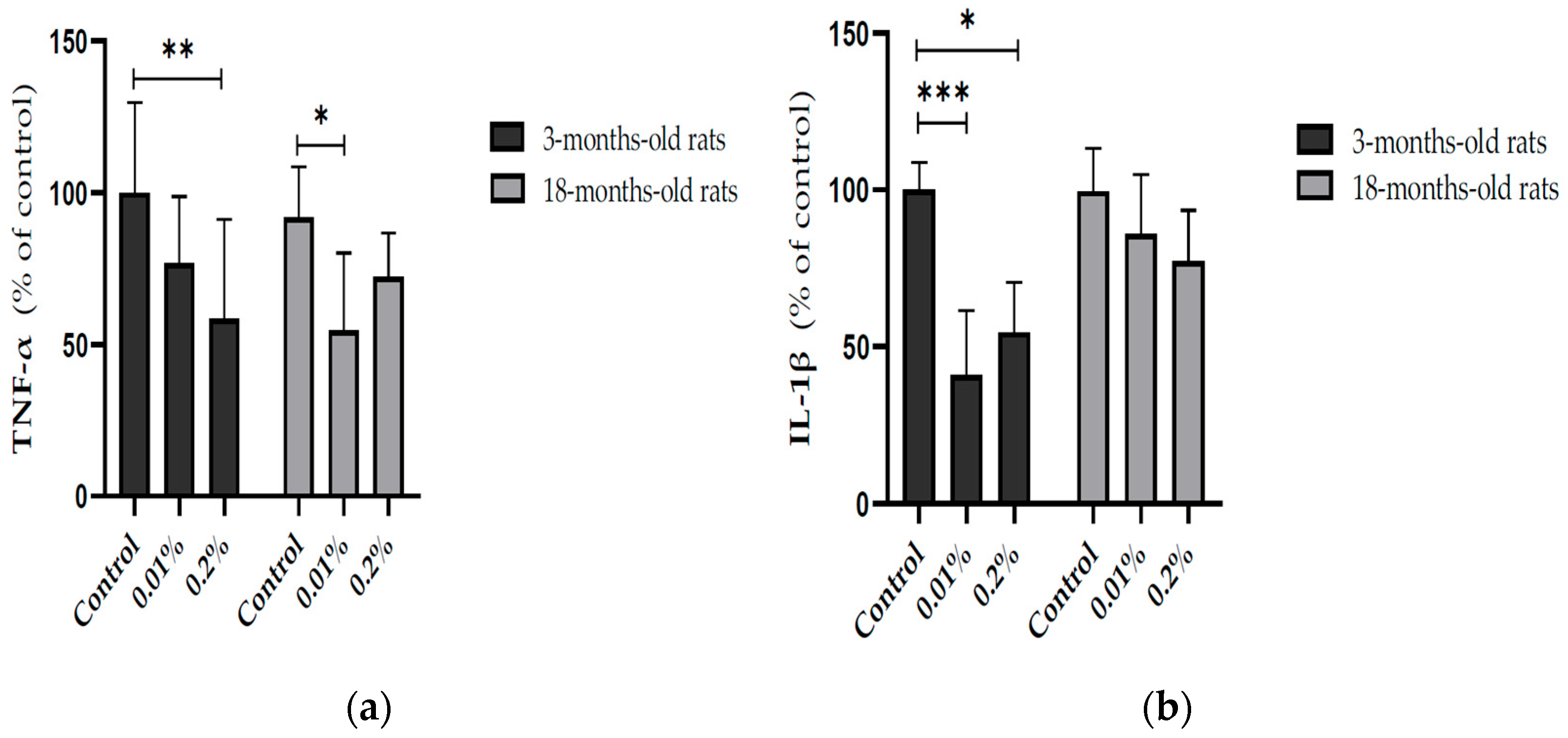
3.3. Effect of Ocimum Americanum Extract Supplementation in Young Adult and Aged Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| OAEE | Ocimum americanum ethanol extract |
| MTT | 3(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide |
| LDH | lactate dehydrogenase |
| EDTA | ethylenediaminetetraacetic acid |
| PMSF | phenylmethylsulfonyl fluoride |
| DCFH-DA | 2′-7′-dichlorofluorescein diacetate |
| LPO | lipid peroxidation |
| TBARS | thiobarbituric acid reactive substances |
| MDA | malondialdehyde |
| AChE | acetylcholinesterase |
| ASCh | acetylthiocholine iodide |
| PBS | phosphate-buffered saline |
| EGTA | ethylene glycol tetraacetic acid |
| IL-1β | interleukin-1β |
| TNF-α | Tumor Necrosis Factor-α |
References
- Leyane, T.S.; Jere, S.W.; Houreld, N.N. Oxidative Stress in Ageing and Chronic Degenerative Pathologies: Molecular Mechanisms Involved in Counteracting Oxidative Stress and Chronic Inflammation. Int. J. Mol. Sci. 2022, 23, 7273. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Yang, L.; Gao, L.; Du, G.; Qin, X.; Zhou, Y. The leaves of Scutellaria baicalensis Georgi attenuate brain aging in D-galactose-induced rats via regulating glutamate metabolism and Nrf2 signaling pathway. Exp. Gerontol. 2022, 170, 111978. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, D.O.; Scholey, A.B. The Psychopharmacology of European Herbs with Cognition-Enhancing Properties. Curr. Pharm. Des. 2006, 12, 4613–4623. [Google Scholar] [CrossRef] [PubMed]
- Eidi, M.; Eidi, A.; Bahar, M. Effects of Salvia officinalis L. (sage) leaves on memory retention and its interaction with the cholinergic system in rats. Nutrition 2006, 22, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Miroddi, M.; Navarra, M.; Quattropani, M.C.; Calapai, F.; Gangemi, S.; Calapai, G. Systematic review of clinical trials assessing pharmacological properties of Salvia species on memory, cognitive impairment and Alzheimer’s disease. CNS Neurosci. Ther. 2014, 20, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Yanpallewar, S.U.; Rai, S.; Kumar, M.; Acharya, S.B. Evaluation of antioxidant and neuroprotective effect of Ocimum sanctum on transient cerebral ischemia and long-term cerebral hypoperfusion. Pharmacol. Biochem. Behav. 2004, 74, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Mataram, M.B.A.; Hening, P.; Harjanti, F.N.; Karnati, S.; Wasityastuti, W.; Nugrahaningsih, D.A.A.; Kusindarta, D.L.; Wihadmadyatami, H. The neuroprotective effect of ethanolic extract Ocimum sanctum Linn. in the regulation of neuronal density in hippocampus areas as a central autobiography memory on the rat model of Alzheimer’s disease. J. Chem. Neuroanat. 2021, 111, 101885. [Google Scholar] [CrossRef] [PubMed]
- Bora, K.S.; Arora, S.; Shri, R. Role of Ocimum basilicum L. in prevention of ischemia and reperfusion-induced cerebral damage, and motor dysfunctions in mice brain. J. Ethnopharmacol. 2011, 137, 1360–1365. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Krishan, P.; Shri, R. Improvement of memory and neurological deficit with Ocimum basilicum L. extract after ischemia reperfusion induced cerebral injury in mice. Metab. Brain Dis. 2018, 33, 1111–1120. [Google Scholar] [CrossRef] [PubMed]
- Pires, D.C.M.; Moraes, A.S. Long-term food supplementation with sweet basil (Ocimum basilicum L.) prevents age-associated cognitive decline in female mice. Nutr. Health 2024, 28, 2601060241281765. [Google Scholar] [CrossRef] [PubMed]
- Beheshti, F.; Vakilian, A.; Navari, M.; Zare Moghaddam, M.; Dinpanah, H.; Ahmadi-Soleimani, S.M. Effects of Ocimum basilicum L. Extract on Hippocampal Oxidative Stress, Inflammation, and BDNF Expression in Amnesic Aged Rats. Exp. Aging Res. 2024, 50, 443–458. [Google Scholar] [CrossRef] [PubMed]
- Venuprasad, M.P.; Hemanth Kumar, K.; Khanum, F. Neuroprotective effects of hydroalcoholic extract of Ocimum sanctum against H2O2 induced neuronal cell damage in SH-SY5Y cells via its antioxidative defence mechanism. Neurochem. Res. 2013, 38, 2190–2200. [Google Scholar] [CrossRef] [PubMed]
- Ionescu-Tucker, A.; Cotman, C.W. Emerging roles of oxidative stress in brain aging and Alzheimer’s disease. Neurobiol Aging. 2021, 107, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Kaurinovic, B.; Popovic, M.; Vlaisavljevic, S.; Trivic, S. Antioxidant capacity of Ocimum basilicum L. and Origanum vulgare L. extracts. Molecules 2011, 16, 7401–7414. [Google Scholar] [CrossRef] [PubMed]
- Zengin, G.; Ferrante, C.; Gnapi, D.E.; Sinan, K.I.; Orlando, G.; Recinella, L.; Diuzheva, A.; Jekő, J.; Cziáky, Z.; Chiavaroli, A.; et al. Comprehensive approaches on the chemical constituents and pharmacological properties of flowers and leaves of American basil (Ocimum americanum L). Food Res. Int. 2019, 125, 108610. [Google Scholar] [CrossRef] [PubMed]
- Lovatel, G.A.; Bertoldi, K.; Elsner, V.R.; Piazza, F.V.; Basso, C.G.; Moysés, F.d.S.; Worm, P.V.; Netto, C.A.; Marcuzzo, S.; Siqueira, I.R. Long-term effects of pre and post-ischemic exercise following global cerebral ischemia on astrocyte and microglia functions in hippocampus from Wistar rats. Brain Res. 2014, 1587, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Porcher, L.; Bruckmeier, S.; Burbano, S.D.; Finnell, J.E.; Gorny, N.; Klett, J.; Wood, S.K.; Kelly, M.P. Aging triggers an upregulation of a multitude of cytokines in the male and especially the female rodent hippocampus but more discrete changes in other brain regions. J. Neuroinflamm. 2021, 18, 219. [Google Scholar] [CrossRef] [PubMed]
- Amini-Khoei, H.; Taei, N.; Dehkordi, H.T.; Lorigooini, Z.; Bijad, E.; Farahzad, A.; Madiseh, M.R. Therapeutic Potential of Ocimum basilicum L. Extract in Alleviating Autistic-Like Behaviors Induced by Maternal Separation Stress in Mice: Role of Neuroinflammation and Oxidative Stress. Phytother. Res. 2025, 39, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Mitra, S.; Turconi, G.; Darreh-Shori, T.; Mätlik, K.; Aquilino, M.; Eriksdotter, M.; Andressoo, J.O. Increased Endogenous GDNF in Mice Protects Against Age-Related Decline in Neuronal Cholinergic Markers. Front. Aging Neurosci. 2021, 13, 714186. [Google Scholar] [CrossRef] [PubMed]
- Castilhos, L.G.; Oliveira, J.S.; Adefegha, S.A.; Manzoni, A.G.; Passos, D.F.; Assmann, C.E.; Silveira, L.L.; Trelles, K.B.; Kronbauer, M.; Doleski, P.H.; et al. Uncaria tomentosa improves cognition, memory and learning in middle-aged rats. Exp. Gerontol. 2020, 138, 111016. [Google Scholar] [CrossRef] [PubMed]
- Perry, N.S.; Houghton, P.J.; Theobald, A.; Jenner, P.; Perry, E.K. In-vitro Inhibition of Human Erythrocyte Acetylcholinesterase by Salvia lavandulaefolia Essential Oil and Constituent Terpenes. J. Pharm. Pharmacol. 2000, 52, 895–902. [Google Scholar] [CrossRef] [PubMed]
- Dabaghian, F.; Delnavazi, M.R.; Hariri, R.; Akbarzadeh, T.; Tayarani-Najaran, Z.; Shams Ardekani, M.R.; Sharifzadeh, M.; Khanavi, M. Neuroprotective, and memory enhancement effects of Salvia aristata and its phenolic constituents: An in vitro, and in vivo study. BMC Complement. Med. Ther. 2025, 25, 181. [Google Scholar] [CrossRef] [PubMed]
- Loizzo, M.R.; Tundis, R.; Conforti, F.; Menichini, F.; Bonesi, M.; Nadjafi, F.; Frega, N.G.; Menichini, F. Salvia leriifolia Benth (Lamiaceae) extract demonstrates in vitro antioxidant properties and cholinesterase inhibitory activity. Nutr. Res. 2010, 30, 823–830. [Google Scholar] [CrossRef] [PubMed]
- Vieira, R.E.; Simon, J.E. Chemical characterization of basil (Ocimum sp.) found in the markets and used in traditional medicine in Brazil. Econ. Bot. 2000, 54, 207–216. [Google Scholar] [CrossRef]
- Ivanova, D.; Gerova, D.; Chervenkov, T.; Yankova, T. Polyphenols and antioxidant capacity of Bulgarian medicinal plants. J. Ethnopharmacol. 2005, 96, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Siqueira, I.R.; Elsner, V.R.; Leite, M.C.; Vanzella, C.F.; Moysés, D.S.; Spindler, C.; Godinho, G.; Battú, C.; Wofchuk, S.; Souza, D.O.; et al. Ascorbate uptake is decreased in the hippocampus of ageing rats. Neurochem. Int. 2011, 58, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Zanella, C.A.; Treichel, H.; Cansian, R.L.; Roman, S.S. The effects of acute administration of the hydroalcoholic extract of rosemary (Rosmarinus officinalis L.) (Lamiaceae) in animal models of memory. Braz. J. Pharm. Sci. 2012, 48, 389–397. [Google Scholar] [CrossRef]
- Siqueira, I.R.; Fochesatto, C.; de Andrade, A.; Santos, M.; Hagen, M.; Bello-Klein, A.; Netto, C.A. Total antioxidant capacity is impaired in different structures from aged rat brain. Int. J. Dev. Neurosci. 2005, 23, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Kolosova, N.G.; Shcheglova, T.V.; Sergeeva, S.V.; Loskutova, L.V. Long-term antioxidant supplementation attenuates oxidative stress markers and cognitive deficits in senescent-accelerated OXYS rats. Neurobiol. Aging 2006, 27, 1289–1297. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Mahendran, G.; Vimolmangkang, S. Chemical compositions, antioxidant, antimicrobial, and mosquito larvicidal activity of Ocimum americanum L. and Ocimum basilicum L. leaf essential oils. BMC Complement. Med. Ther. 2023, 23, 390. [Google Scholar] [CrossRef] [PubMed]
- Sanmartín-Suárez, C.; Soto-Otero, R.; Sánchez-Sellero, I.; Méndez-Álvarez, E. Antioxidant properties of dimethyl sulfoxide and its viability as a solvent in the evaluation of neuroprotective antioxidants. J. Pharmacol. Toxicol. Methods 2011, 63, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Bora, K.S.; Shri, R.; Monga, J. Cerebroprotective effect of Ocimum gratissimum against focal ischemia and reperfusion-induced cerebral injury. Pharm. Biol. 2011, 49, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Asadi, S.; Ahmadiani, A.; Esmaeili, M.A.; Sonboli, A.; Ansari, N.; Khodagholi, F. In vitro antioxidant activities and an investigation of neuroprotection by six Salvia species from Iran: A comparative study. Food Chem. Toxicol. 2010, 48, 1341–1349. [Google Scholar] [CrossRef] [PubMed]
- Park, S.E.; Kim, S.; Sapkota, K.; Kim, S.J. Neuroprotective effect of Rosmarinus officinalis extract on human dopaminergic cell line, SH-SY5Y. Cell. Mol. Neurobiol. 2010, 30, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Yadav, I.; Sharma, N.; Velayudhan, R.; Fatima, Z.; Maras, J.S. Ocimum sanctum Alters the Lipid Landscape of the Brain Cortex and Plasma to Ameliorate the Effect of Photothrombotic Stroke in a Mouse Model. Life 2023, 13, 1877. [Google Scholar] [CrossRef] [PubMed]
- Farag, M.A.; Ezzat, S.M.; Salama, M.M.; Tadros, M.G. Anti-acetylcholinesterase potential and metabolome classification of 4 Ocimum species as determined via UPLC/qTOF/MS and chemometric tools. J. Pharm. Biomed. Anal. 2016, 125, 292–302. [Google Scholar] [CrossRef] [PubMed]
- Giridharan, V.V.; Thandavarayan, R.A.; Mani, V.; Ashok Dundapa, T.; Watanabe, K.; Konishi, T. Ocimum sanctum Linn. leaf extracts inhibit acetylcholinesterase and improve cognition in rats with experimentally induced dementia. J. Med. Food 2011, 14, 912–919. [Google Scholar] [CrossRef] [PubMed]
- Tadros, M.G.; Ezzat, S.M.; Salama, M.M.; Farag, M.A. In vitro and in vivo Anticholinesterase Activity of the Volatile Oil of the Aerial Parts of Ocimum basilicum L. and O. africanum Lour. Growing in Egypt. World Acad. Sci. Eng. Technol. Int. J. Pharmacol. Pharm. Sci. 2014, 1, 157–161. [Google Scholar]
- Schliebs, R.; Arendt, T. The cholinergic system in aging and neuronal degeneration. Behav. Brain Res. 2011, 221, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Chiang, N.; Ray, S.; Lomax, J.; Goertzen, S.; Komarnytsky, S.; Ho, C.T.; Munafo, J.P., Jr. Modulation of Brain-Derived Neurotrophic Factor (BDNF) Signaling Pathway by Culinary Sage (Salvia officinalis L.). Int. J. Mol. Sci. 2021, 22, 7382. [Google Scholar] [CrossRef] [PubMed]
- Chwang, W.B.; O’Riordan, K.J.; Levenson, J.M.; Sweatt, J.D. ERK/MAPK regulates hippocampal histone phosphorylation following contextual fear conditioning. Learn. Mem. 2006, 13, 322–328. [Google Scholar] [CrossRef] [PubMed]
- de Meireles, L.C.; Bertoldi, K.; Elsner, V.R.; Moysés, F.d.S.; Siqueira, I.R. Treadmill exercise alters histone acetylation differently in rats exposed or not exposed to aversive learning context. Neurobiol. Learn. Mem. 2014, 116, 193–196. [Google Scholar] [CrossRef] [PubMed]
- Siqueira, I.R.; Cimarosti, H.; Fochesatto, C.; Salbego, C.; Netto, C.A. Age-related susceptibility to oxygen and glucose deprivation damage in rat hippocampal slices. Brain Res. 2004, 1025, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Burke, S.N.; Foster, T.C. Animal models of cognitive aging and circuit-specific vulnerability. Handb. Clin. Neurol. 2019, 167, 19–36. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siqueira, I.R.; Vanzella, C.; Lovatel, G.A.; Bertoldi, K.; Spindler, C.; Moysés, F.d.S.; Vizuete, A.; Poser, G.L.v.; Netto, C.A. American Basil, Ocimum americanum, Has Neuroprotective Properties in the Aging Process. Nutrients 2025, 17, 2368. https://doi.org/10.3390/nu17142368
Siqueira IR, Vanzella C, Lovatel GA, Bertoldi K, Spindler C, Moysés FdS, Vizuete A, Poser GLv, Netto CA. American Basil, Ocimum americanum, Has Neuroprotective Properties in the Aging Process. Nutrients. 2025; 17(14):2368. https://doi.org/10.3390/nu17142368
Chicago/Turabian StyleSiqueira, Ionara Rodrigues, Cláudia Vanzella, Gisele Agustini Lovatel, Karine Bertoldi, Christiano Spindler, Felipe dos Santos Moysés, Adriana Vizuete, Gilsane Lino von Poser, and Carlos Alexandre Netto. 2025. "American Basil, Ocimum americanum, Has Neuroprotective Properties in the Aging Process" Nutrients 17, no. 14: 2368. https://doi.org/10.3390/nu17142368
APA StyleSiqueira, I. R., Vanzella, C., Lovatel, G. A., Bertoldi, K., Spindler, C., Moysés, F. d. S., Vizuete, A., Poser, G. L. v., & Netto, C. A. (2025). American Basil, Ocimum americanum, Has Neuroprotective Properties in the Aging Process. Nutrients, 17(14), 2368. https://doi.org/10.3390/nu17142368