

Glutamine Administration Attenuates Poly(I:C)-Induced Lung Injury by Reducing Neutrophil Infiltration and Activating the TLR-3 Antiviral Pathway

 , ,
, , 
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Procedures
2.3. Quantification of GLN Concentration in Plasma
2.4. Concentrations of Inflammatory Cytokines and Chemokines in Plasma
2.5. Populations of Macrophages and Neutrophils in BALF
2.6. Isolating Leukocytes, Endothelial Cells, and Epithelial Cells from Lung Tissues
2.7. Concentrations of Inflammatory Cytokines and Chemokines in BALF
2.8. Protein Expression of Phosphorylated (p)-IRF3 in Lung Tissues
2.9. The Preparation of Messenger (m)RNA and Analysis of a Real-Time Reverse-Transcription (RT) Quantitative Polymerase Chain Reaction (qPCR)
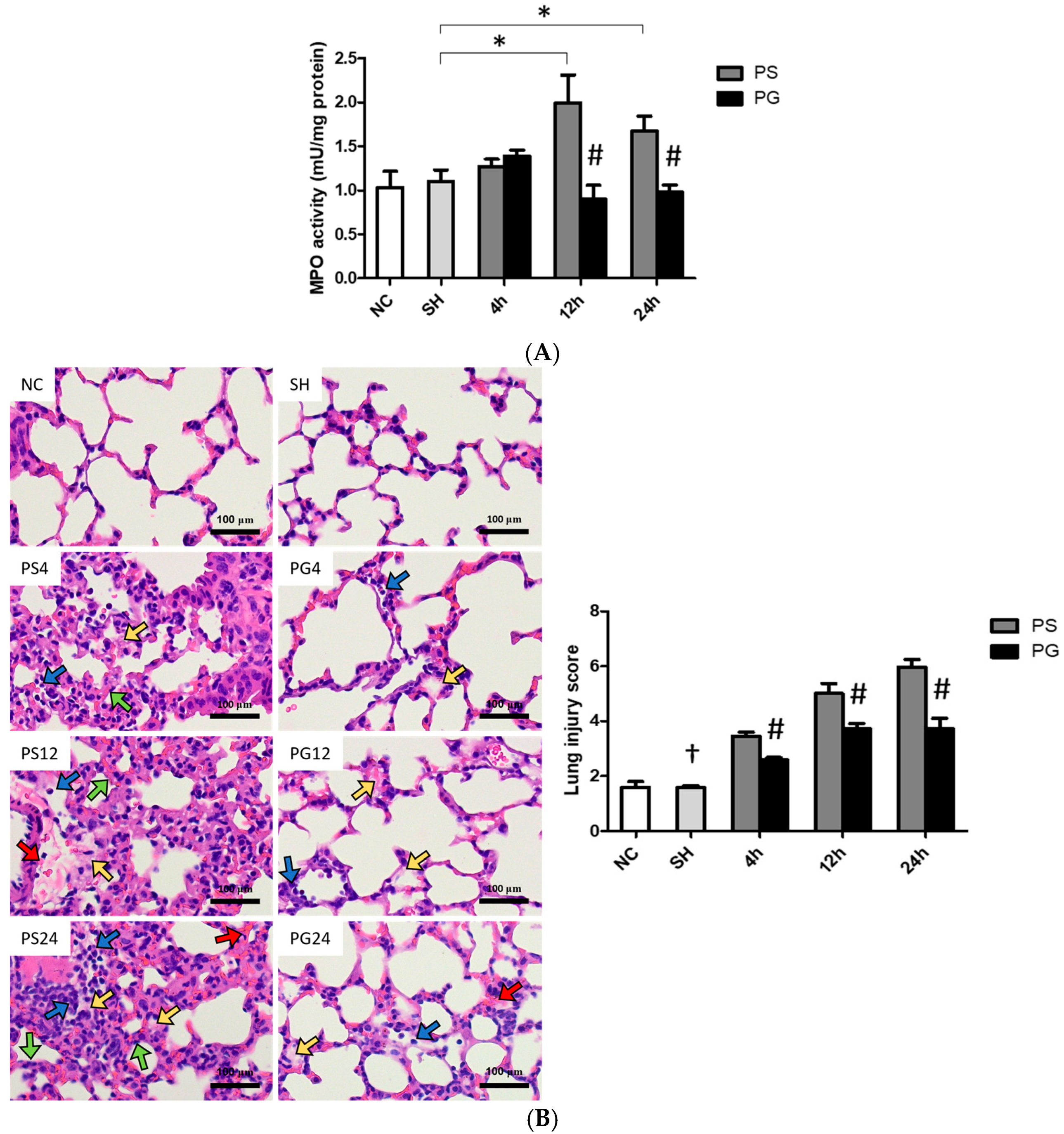
2.10. Analysis of Myeloperoxidase (MPO) Activities in Lung Tissues
2.11. Hematoxylin and Eosin (H&E) Staining and Histopathological Scoring of Lung Tissues
2.12. Statistical Analysis
3. Results
3.1. BWs and Weights of Lung Tissues/BW Ratio
3.2. Concentration of GLN in Plasma
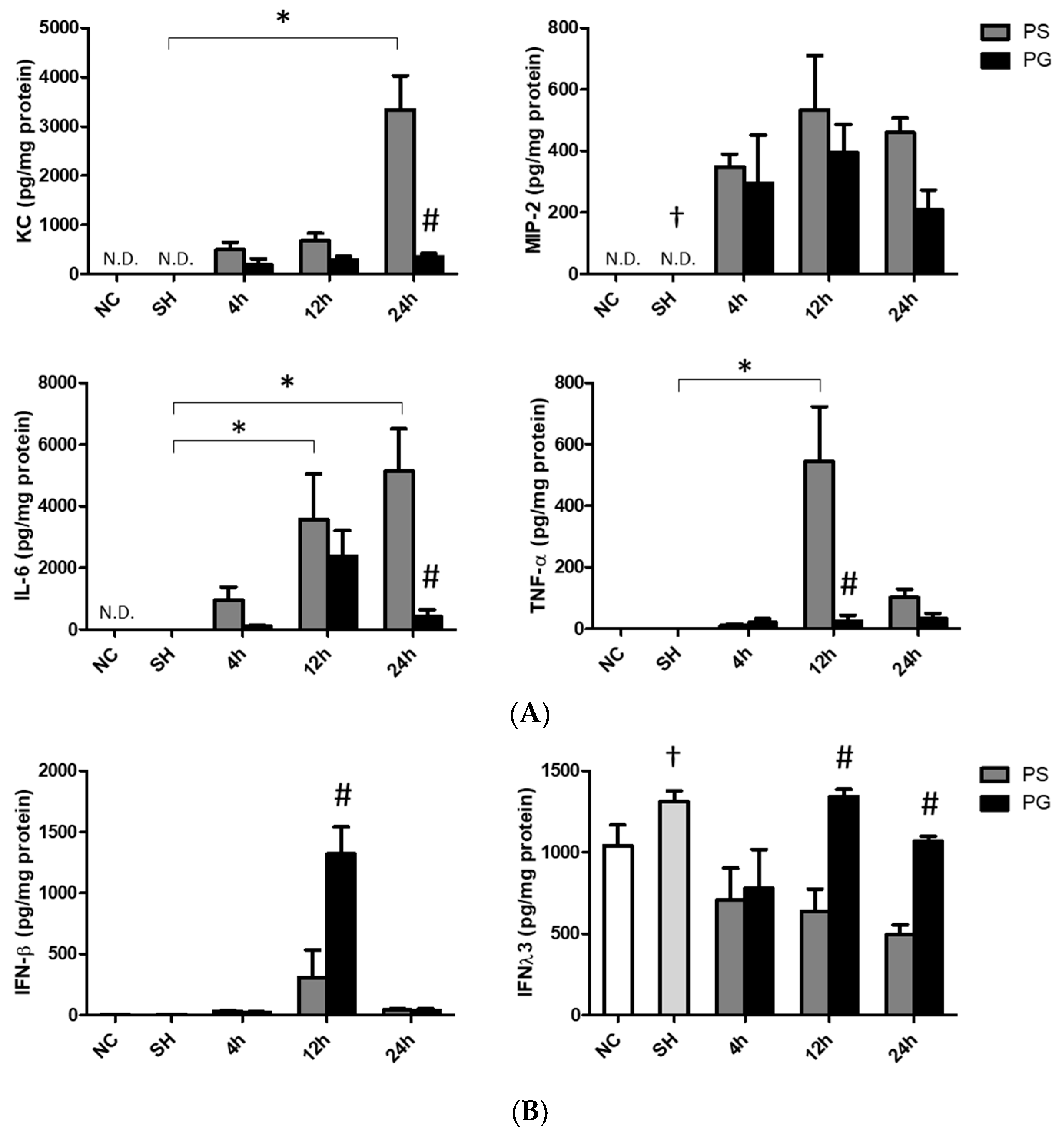
3.3. Concentrations of KC, MIP-2, IL-6, and TNF-α in Plasma
3.4. Concentrations of Chemokines and Cytokines in BALF
3.5. Distributions of Leukocytes, Macrophages, M1 Macrophages, and Neutrophils in BALF
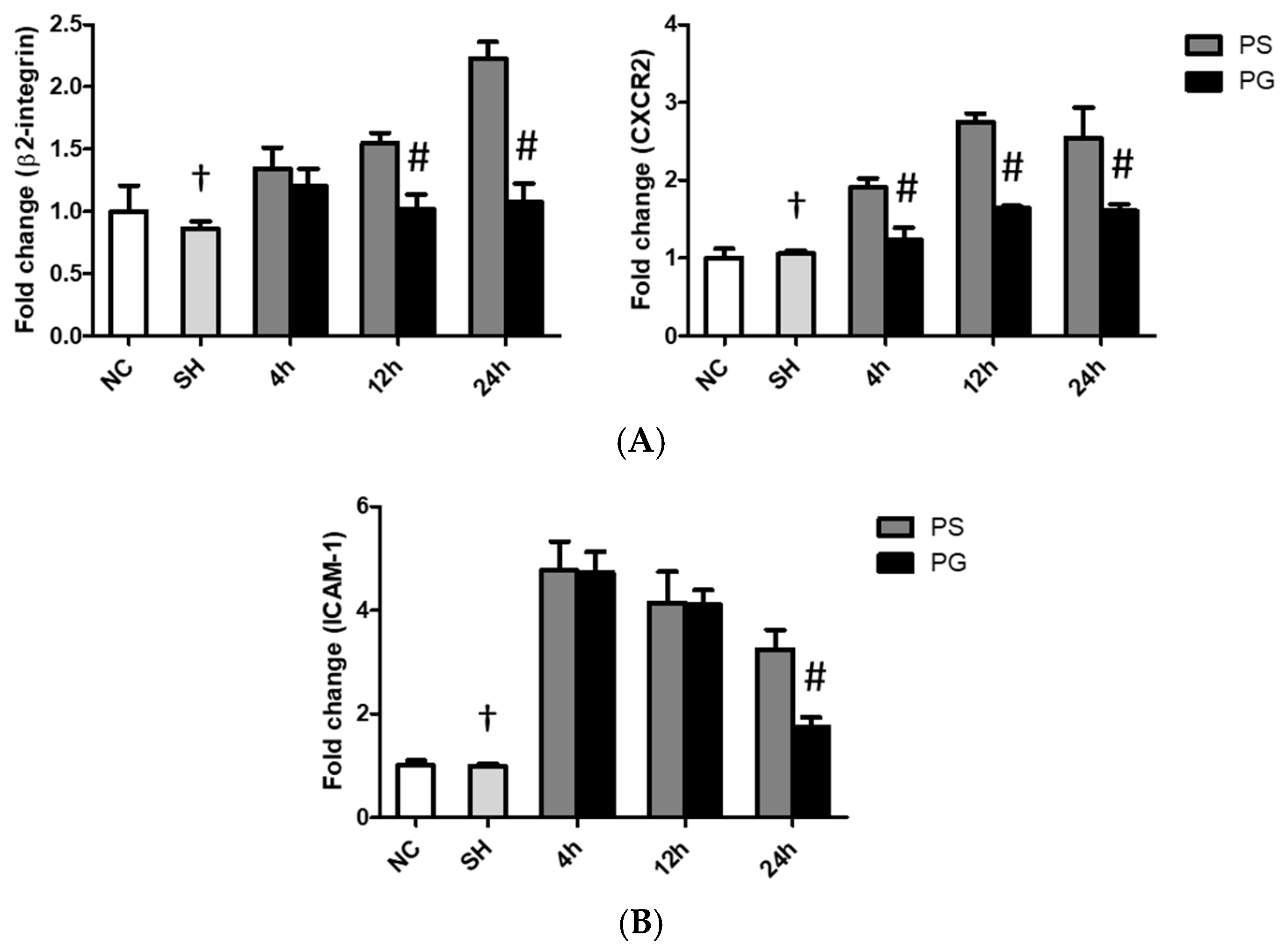
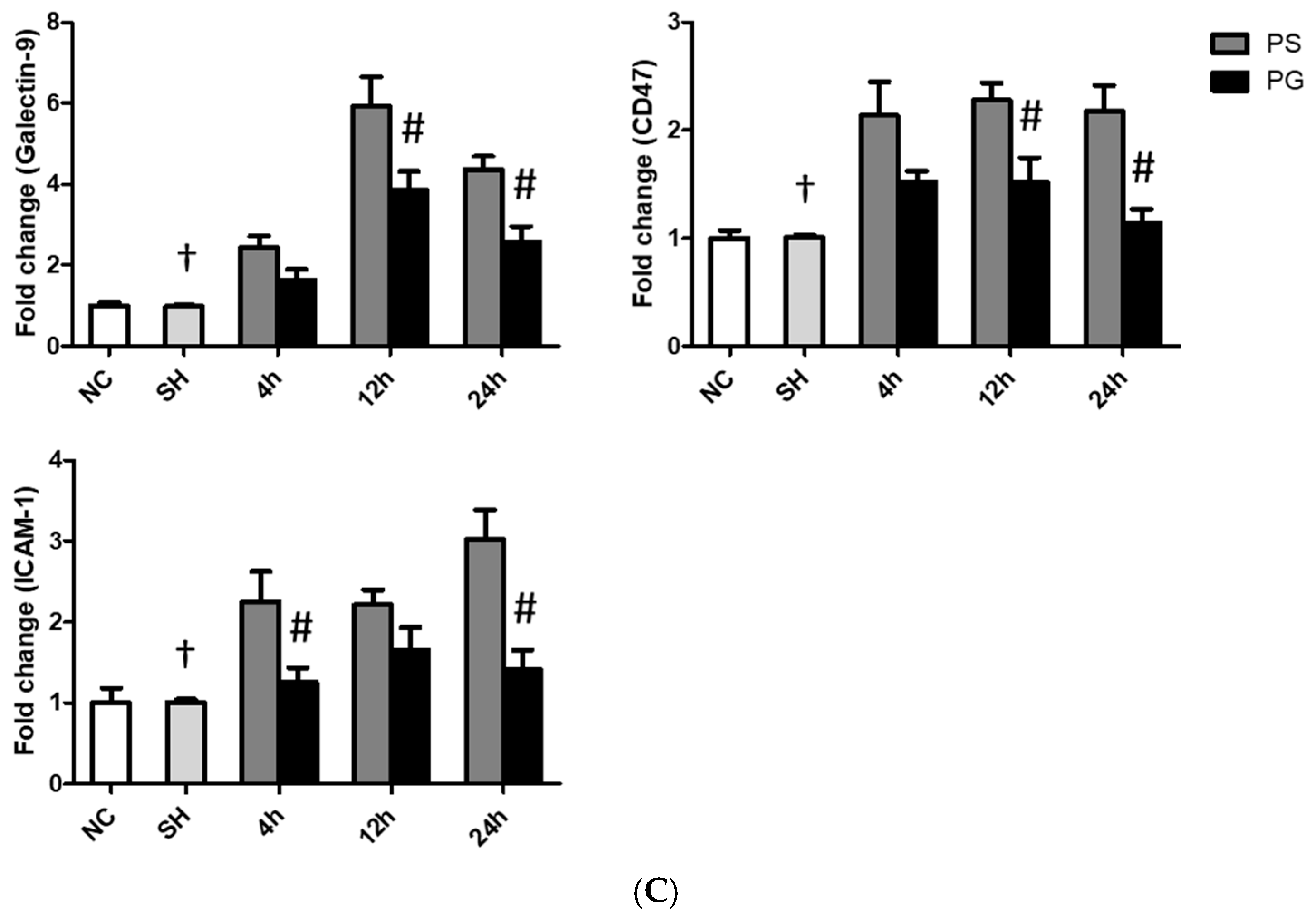
3.6. mRNA Expressions of Leukocyte Migration-Related Factors in Lung Leukocytes, Endothelial Cells, and Epithelial Cells
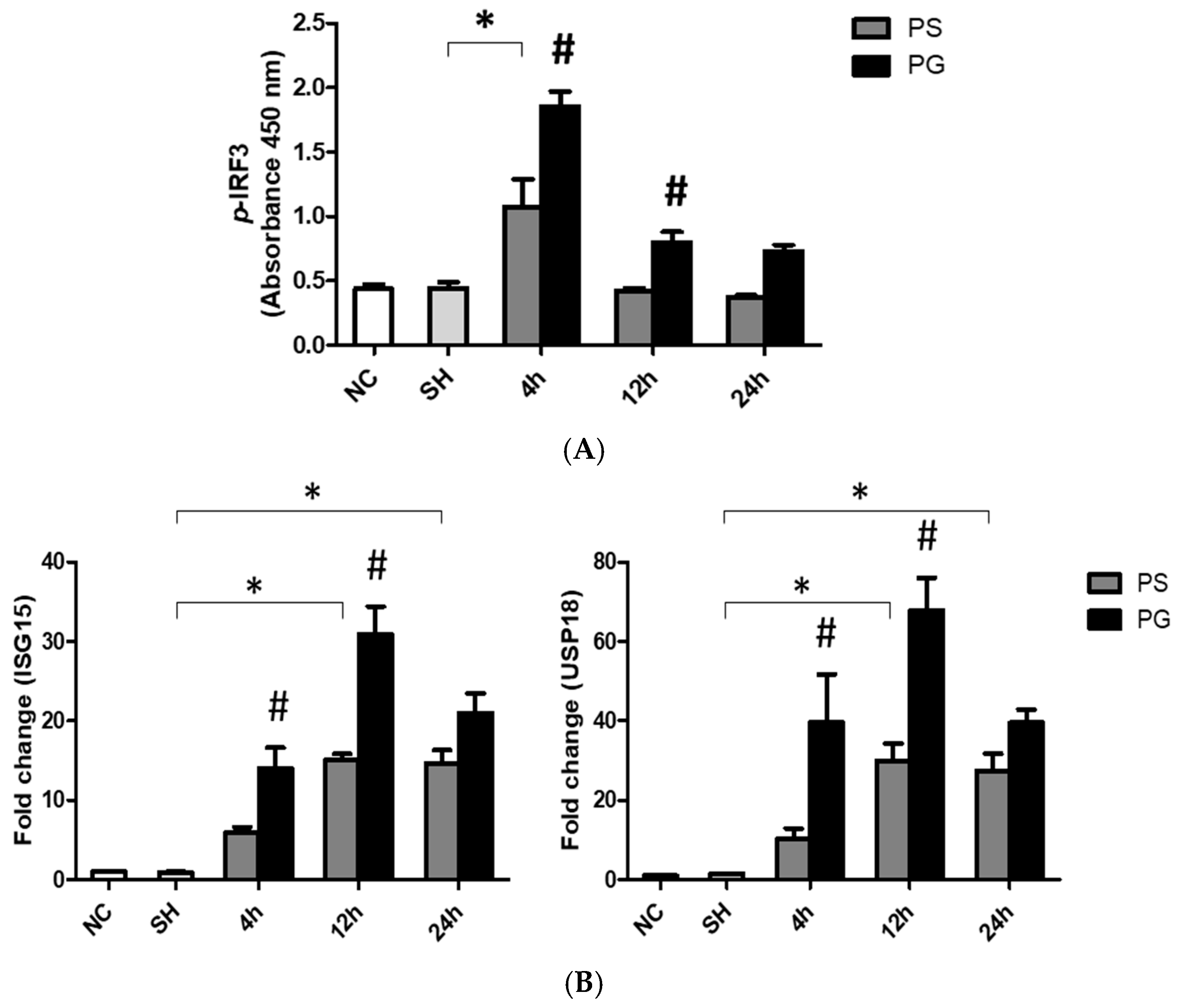
3.7. mRNA Expressions of TLR-3 Pathway-Related Factors and Protein Expressions of p-IRF3 in Lung Tissues
3.8. mRNA Expressions of Adherens and Tight Junction Proteins in Lung Endothelial and Epithelial Cells
3.9. Histopathological Findings of Lung Tissues
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ALI | Acute lung injury |
| ARDS | Acute respiratory distress syndrome |
| BALF | Bronchoalveolar lavage fluid |
| BW | Body weight |
| CXCR2 | C-X-C motif chemokine receptor 2 |
| dsRNA | Double-stranded RNA |
| ELISA | Enzyme-linked immunosorbent assay |
| GLN | Glutamine |
| H&E | Hematoxylin and eosin |
| ICAM-1 | Intercellular adhesion molecule 1 |
| IFN | Interferon |
| IL | Interleukin |
| IRF3 | IFN-regulatory factor 3 |
| ISG15 | IFN-stimulated gene 15 |
| IV | intravenous |
| KC | Keratinocyte-derived chemokine |
| LPS | Lipopolysaccharide |
| MIP-2 | Macrophage inflammatory protein-2 |
| MPO | Myeloperoxidase |
| PBS | Phosphate-buffered saline |
| ROS | Reactive oxygen species |
| TLR | Toll-like receptor |
| TNF-α | Tumor necrosis factor-α |
| TRIF | Toll-interleukin-1 receptor-domain-containing adapter-inducing interferon-β |
| UPLC | Ultraperformance liquid chromatography |
| USP18 | Ubiquitin-specific peptidase 18 |
| VE-cadherin | Vascular endothelial-cadherin |
References
- Mowery, N.T.; Terzian, W.T.H.; Nelson, A.C. Acute lung injury. Curr. Probl. Surg. 2020, 57, 100777. [Google Scholar] [CrossRef] [PubMed]
- Hodinka, R.L. Respiratory RNA Viruses. Microbiol. Spectr. 2016, 4, 233–271. [Google Scholar] [CrossRef]
- Hsieh, P.C.; Wu, Y.K.; Yang, M.C.; Su, W.L.; Kuo, C.Y.; Lan, C.C. Deciphering the role of damage-associated molecular patterns and inflammatory responses in acute lung injury. Life Sci. 2022, 305, 120782. [Google Scholar] [CrossRef]
- Cheng, P.; Li, S.; Chen, H. Macrophages in Lung Injury, Repair, and Fibrosis. Cells 2021, 10, 436. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, M.; Niisato, M.; Kawasaki, Y.; Karaman, S.; Robciuc, M.R.; Shibata, Y.; Ishida, Y.; Nishio, R.; Masuda, T.; Sugai, T.; et al. VEGF-C/VEGFR-3 signalling in macrophages ameliorates acute lung injury. Eur. Respir. J. 2022, 59, 2100880. [Google Scholar] [CrossRef]
- Rosales, C. Neutrophil: A Cell with Many Roles in Inflammation or Several Cell Types? Front. Physiol. 2018, 9, 113. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.C.; Tsai, Y.F.; Pan, Y.L.; Hwang, T.L. Understanding the role of neutrophils in acute respiratory distress syndrome. Biomed. J. 2021, 44, 439–446. [Google Scholar] [CrossRef]
- Akgul, C.; Moulding, D.A.; Edwards, S.W. Molecular control of neutrophil apoptosis. FEBS Lett. 2001, 487, 318–322. [Google Scholar] [CrossRef]
- Chen, Y.; Lin, J.; Zhao, Y.; Ma, X.; Yi, H. Toll-like receptor 3 (TLR3) regulation mechanisms and roles in antiviral innate immune responses. J. Zhejiang Univ. Sci. B 2021, 22, 609–632. [Google Scholar] [CrossRef]
- Gibbert, K.; Francois, S.; Sigmund, A.M.; Harper, M.S.; Barrett, B.S.; Kirchning, C.J.; Lu, M.; Santiago, M.L.; Dittmer, U. Friend retrovirus drives cytotoxic effectors through Toll-like receptor 3. Retrovirology 2014, 11, 126. [Google Scholar] [CrossRef]
- Mohajeri, M.; Horriatkhah, E.; Mohajery, R. The effect of glutamine supplementation on serum levels of some inflammatory factors, oxidative stress, and appetite in COVID-19 patients: A case-control study. Inflammopharmacology 2021, 29, 1769–1776. [Google Scholar] [CrossRef] [PubMed]
- Bharadwaj, S.; Singh, M.; Kirtipal, N.; Kang, S.G. SARS-CoV-2 and Glutamine: SARS-CoV-2 Triggered Pathogenesis via Metabolic Reprograming of Glutamine in Host Cells. Front. Mol. Biosci. 2020, 7, 627842. [Google Scholar] [CrossRef] [PubMed]
- Kim, H. Glutamine as an immunonutrient. Yonsei Med. J. 2011, 52, 892–897. [Google Scholar] [CrossRef]
- Santos, A.C.A.; Hebeba, C.B.; Hastreiter, A.A.; de Oliveira, D.C.; Naoto Makiyama, E.; Farsky, S.H.P.; Borelli, P.; Fock, R.A. Exogenous glutamine impairs neutrophils migration into infections sites elicited by lipopolysaccharide by a multistep mechanism. Amino Acids 2019, 51, 451–462. [Google Scholar] [CrossRef]
- Hou, Y.C.; Chiu, W.C.; Yeh, C.L.; Yeh, S.L. Glutamine modulates lipopolysaccharide-induced activation of NF-κB via the Akt/mTOR pathway in lung epithelial cells. Am. J. Physiol. Lung Cell Mol. Physiol. 2012, 302, L174–L183. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.L.; Hsu, C.S.; Yeh, S.L.; Chen, W.J. Dietary glutamine supplementation modulates Th1/Th2 cytokine and interleukin-6 expressions in septic mice. Cytokine 2005, 31, 329–334. [Google Scholar] [CrossRef]
- Yeh, C.L.; Su, L.H.; Wu, J.M.; Yang, P.J.; Lee, P.C.; Chen, P.D.; Huang, C.C.; Hsieh, D.Y.; Wang, H.J.; Yeh, S.L.; et al. Effects of the Glutamine Administration on T Helper Cell Regulation and Inflammatory Response in Obese Mice Complicated with Polymicrobial Sepsis. Mediat. Inflamm. 2020, 2020, 8869017. [Google Scholar] [CrossRef]
- Hu, Y.M.; Yeh, C.L.; Pai, M.H.; Lee, W.Y.; Yeh, S.L. Glutamine administration modulates lung γδ T lymphocyte expression in mice with polymicrobial sepsis. Shock 2014, 41, 115–122. [Google Scholar] [CrossRef]
- Amemiya, K.; Dankmeyer, J.L.; Bernhards, R.C.; Fetterer, D.P.; Waag, D.M.; Worsham, P.L.; DeShazer, D. Activation of Toll-Like Receptors by Live Gram-Negative Bacterial Pathogens Reveals Mitigation of TLR4 Responses and Activation of TLR5 by Flagella. Front. Cell Infect. Microbiol. 2021, 11, 745325. [Google Scholar] [CrossRef]
- Stowell, N.C.; Seideman, J.; Raymond, H.A.; Smalley, K.A.; Lamb, R.J.; Egenolf, D.D.; Bugelski, P.J.; Murray, L.A.; Marsters, P.A.; Bunting, R.A.; et al. Long-term activation of TLR3 by poly(I:C) induces inflammation and impairs lung function in mice. Respir. Res. 2009, 10, 43. [Google Scholar] [CrossRef]
- Ulbrich, H.; Eriksson, E.E.; Lindbom, L. Leukocyte and endothelial cell adhesion molecules as targets for therapeutic interventions in inflammatory disease. Trends Pharmacol. Sci. 2003, 24, 640–647. [Google Scholar] [CrossRef]
- Nolte, D.; Kuebler, W.M.; Muller, W.A.; Wolff, K.D.; Messmer, K. Attenuation of leukocyte sequestration by selective blockade of PECAM-1 or VCAM-1 in murine endotoxemia. Eur. Surg. Res. 2004, 36, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.L.; Wu, J.M.; Chen, K.Y.; Wu, M.H.; Yang, P.J.; Lee, P.C.; Chen, P.D.; Kuo, T.C.; Yeh, S.L.; Lin, M.T. Calcitriol attenuates poly(I:C)-induced lung injury in obese mice via modulating toll-like receptor 3- and renin-angiotensin system-associated signal pathways. Int. Immunopharmacol. 2024, 128, 111522. [Google Scholar] [CrossRef]
- Oliveira, G.P.; Oliveira, M.B.; Santos, R.S.; Lima, L.D.; Dias, C.M.; Ab’Saber, A.M.; Teodoro, W.R.; Capelozzi, V.L.; Gomes, R.N.; Bozza, P.T.; et al. Intravenous glutamine decreases lung and distal organ injury in an experimental model of abdominal sepsis. Crit. Care 2009, 13, R74. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Dawood, F.S.; Iuliano, A.D.; Reed, C.; Meltzer, M.I.; Shay, D.K.; Cheng, P.Y.; Bandaranayake, D.; Breiman, R.F.; Brooks, W.A.; Buchy, P.; et al. Estimated global mortality associated with the first 12 months of 2009 pandemic influenza A H1N1 virus circulation: A modelling study. Lancet Infect. Dis. 2012, 12, 687–695. [Google Scholar] [CrossRef]
- Curi, R.; Newsholme, P.; Pithon-Curi, T.C.; Pires-de-Melo, M.; Garcia, C.; Homem-de-Bittencourt Júnior, P.I.; Guimarães, A.R. Metabolic fate of glutamine in lymphocytes, macrophages and neutrophils. Braz. J. Med. Biol. Res. 1999, 32, 15–21. [Google Scholar] [CrossRef]
- Robinson, N.; Ganesan, R.; Hegedűs, C.; Kovács, K.; Kufer, T.A.; Virág, L. Programmed necrotic cell death of macrophages: Focus on pyroptosis, necroptosis, and parthanatos. Redox Biol. 2019, 26, 101239. [Google Scholar] [CrossRef]
- Li, K.; McCaw, J.M.; Cao, P. Modelling within-host macrophage dynamics in influenza virus infection. J. Theor. Biol. 2021, 508, 110492. [Google Scholar] [CrossRef]
- Yu, S.; Ge, H.; Li, S.; Qiu, H.J. Modulation of Macrophage Polarization by Viruses: Turning Off/On Host Antiviral Responses. Front. Microbiol. 2022, 13, 839585. [Google Scholar] [CrossRef]
- Cassetta, L.; Kajaste-Rudnitski, A.; Coradin, T.; Saba, E.; Della Chiara, G.; Barbagallo, M.; Graziano, F.; Alfano, M.; Cassol, E.; Vicenzi, E.; et al. M1 polarization of human monocyte-derived macrophages restricts pre and postintegration steps of HIV-1 replication. Aids 2013, 27, 1847–1856. [Google Scholar] [CrossRef] [PubMed]
- Puneet, P.; Moochhala, S.; Bhatia, M. Chemokines in acute respiratory distress syndrome. Am. J. Physiol. Lung Cell Mol. Physiol. 2005, 288, L3–L15. [Google Scholar] [CrossRef]
- Griffith, J.W.; Sokol, C.L.; Luster, A.D. Chemokines and chemokine receptors: Positioning cells for host defense and immunity. Annu. Rev. Immunol. 2014, 32, 659–702. [Google Scholar] [CrossRef] [PubMed]
- Girbl, T.; Lenn, T.; Perez, L.; Rolas, L.; Barkaway, A.; Thiriot, A.; Del Fresno, C.; Lynam, E.; Hub, E.; Thelen, M.; et al. Distinct Compartmentalization of the Chemokines CXCL1 and CXCL2 and the Atypical Receptor ACKR1 Determine Discrete Stages of Neutrophil Diapedesis. Immunity 2018, 49, 1062–1076.e1066. [Google Scholar] [CrossRef]
- Bednarczyk, M.; Stege, H.; Grabbe, S.; Bros, M. β2 Integrins-Multi-Functional Leukocyte Receptors in Health and Disease. Int. J. Mol. Sci. 2020, 21, 1402. [Google Scholar] [CrossRef]
- Reutershan, J.; Morris, M.A.; Burcin, T.L.; Smith, D.F.; Chang, D.; Saprito, M.S.; Ley, K. Critical role of endothelial CXCR2 in LPS-induced neutrophil migration into the lung. J. Clin. Investig. 2006, 116, 695–702. [Google Scholar] [CrossRef]
- Lin, W.C.; Fessler, M.B. Regulatory mechanisms of neutrophil migration from the circulation to the airspace. Cell Mol. Life Sci. 2021, 78, 4095–4124. [Google Scholar] [CrossRef] [PubMed]
- Steichen, A.L.; Simonson, T.J.; Salmon, S.L.; Metzger, D.W.; Mishra, B.B.; Sharma, J. Alarmin function of galectin-9 in murine respiratory tularemia. PLoS ONE 2015, 10, e0123573. [Google Scholar] [CrossRef]
- Effah, C.Y.; Drokow, E.K.; Agboyibor, C.; Ding, L.; He, S.; Liu, S.; Akorli, S.Y.; Nuamah, E.; Sun, T.; Zhou, X.; et al. Neutrophil-Dependent Immunity During Pulmonary Infections and Inflammations. Front. Immunol. 2021, 12, 689866. [Google Scholar] [CrossRef]
- Mullane, K.M.; Kraemer, R.; Smith, B. Myeloperoxidase activity as a quantitative assessment of neutrophil infiltration into ischemic myocardium. J. Pharmacol. Methods 1985, 14, 157–167. [Google Scholar] [CrossRef]
- Park, A.; Iwasaki, A. Type I and Type III Interferons—Induction, Signaling, Evasion, and Application to Combat COVID-19. Cell Host Microbe 2020, 27, 870–878. [Google Scholar] [CrossRef]
- Han, L.; Zhuang, M.W.; Deng, J.; Zheng, Y.; Zhang, J.; Nan, M.L.; Zhang, X.J.; Gao, C.; Wang, P.H. SARS-CoV-2 ORF9b antagonizes type I and III interferons by targeting multiple components of the RIG-I/MDA-5-MAVS, TLR3-TRIF, and cGAS-STING signaling pathways. J. Med. Virol. 2021, 93, 5376–5389. [Google Scholar] [CrossRef]
- Kessler, D.S.; Levy, D.E.; Darnell, J.E., Jr. Two interferon-induced nuclear factors bind a single promoter element in interferon-stimulated genes. Proc. Natl. Acad. Sci. USA 1988, 85, 8521–8525. [Google Scholar] [CrossRef]
- Jiménez Fernández, D.; Hess, S.; Knobeloch, K.P. Strategies to Target ISG15 and USP18 Toward Therapeutic Applications. Front. Chem. 2019, 7, 923. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Bogunovic, D.; Payelle-Brogard, B.; Francois-Newton, V.; Speer, S.D.; Yuan, C.; Volpi, S.; Li, Z.; Sanal, O.; Mansouri, D.; et al. Human intracellular ISG15 prevents interferon-α/β over-amplification and auto-inflammation. Nature 2015, 517, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Jewell, N.A.; Cline, T.; Mertz, S.E.; Smirnov, S.V.; Flaño, E.; Schindler, C.; Grieves, J.L.; Durbin, R.K.; Kotenko, S.V.; Durbin, J.E. Lambda interferon is the predominant interferon induced by influenza A virus infection in vivo. J. Virol. 2010, 84, 11515–11522. [Google Scholar] [CrossRef] [PubMed]
- Dellgren, C.; Gad, H.H.; Hamming, O.J.; Melchjorsen, J.; Hartmann, R. Human interferon-lambda3 is a potent member of the type III interferon family. Genes. Immun. 2009, 10, 125–131. [Google Scholar] [CrossRef]
- Bolen, C.R.; Ding, S.; Robek, M.D.; Kleinstein, S.H. Dynamic expression profiling of type I and type III interferon-stimulated hepatocytes reveals a stable hierarchy of gene expression. Hepatology 2014, 59, 1262–1272. [Google Scholar] [CrossRef]
- Vestweber, D. VE-cadherin: The major endothelial adhesion molecule controlling cellular junctions and blood vessel formation. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 223–232. [Google Scholar] [CrossRef]
- Dejana, E.; Tournier-Lasserve, E.; Weinstein, B.M. The control of vascular integrity by endothelial cell junctions: Molecular basis and pathological implications. Dev. Cell 2009, 16, 209–221. [Google Scholar] [CrossRef]
- Grommes, J.; Soehnlein, O. Contribution of neutrophils to acute lung injury. Mol. Med. 2011, 17, 293–307. [Google Scholar] [CrossRef]
- Kotton, D.N. Claudin-18: Unexpected regulator of lung alveolar epithelial cell proliferation. J. Clin. Investig. 2018, 128, 903–905. [Google Scholar] [CrossRef] [PubMed]
- Yaqoob, P.; Calder, P.C. Cytokine production by human peripheral blood mononuclear cells: Differential senstivity to glutamine availability. Cytokine 1998, 10, 790–794. [Google Scholar] [CrossRef] [PubMed]
- Field, C.J.; Johnson, I.R.; Schley, P.D. Nutrients and their role in host resistance to infection. J. Leukoc. Biol. 2002, 71, 16–32. [Google Scholar] [CrossRef] [PubMed]
- Murphy, C.; Newsholme, P. Importance of glutamine metabolism in murine macrophages and human monocytes to L-arginine biosynthesis and rates of nitrite or urea production. Clin. Sci. 1998, 95, 397–407. [Google Scholar] [CrossRef]
- Pithon-Curi, T.C.; Schumacher, R.I.; Freitas, J.J.; Lagranha, C.; Newsholme, P.; Palanch, A.C.; Doi, S.Q.; Curi, R. Glutamine delays spontaneous apoptosis in neutrophils. Am. J. Physiol. Cell Physiol. 2003, 284, C1355–C1361. [Google Scholar] [CrossRef]
- Singleton, K.D.; Beckey, V.E.; Wischmeyer, P.E. Glutamine prevents activation of nf-kappab and stress kinase pathways, attenuates inflammatory cytokine release, and prevents acute respiratory distress syndrome (ARDS) following sepsis. Shock 2005, 24, 583–589. [Google Scholar] [CrossRef]
- Sato, N.; Moore, F.A.; Kone, B.C.; Zou, L.; Smith, M.A.; Childs, M.A.; Moore-Olufemi, S.; Schultz, S.G.; Kozar, R.A. Differential induction of PPAR-gamma by luminal glutamine and iNOS by luminal arginine in the rodent postischemic small bowel. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 290, G616–G623. [Google Scholar] [CrossRef]
- Liu, N.; Ma, X.; Luo, X.; Zhang, Y.; He, Y.; Dai, Z.; Yang, Y.; Wu, G.; Wu, Z. l-Glutamine Attenuates Apoptosis in Porcine Enterocytes by Regulating Glutathione-Related Redox Homeostasis. J. Nutr. 2018, 148, 526–534. [Google Scholar] [CrossRef]
- Zhang, Y.L.; Li, Q.Q.; Guo, W.; Huang, Y.; Yang, J. Effects of chronic ethanol ingestion on tight junction proteins and barrier function of alveolar epithelium in the rat. Shock 2007, 28, 245–252. [Google Scholar] [CrossRef]
- Roth, E.; Oehler, R.; Manhart, N.; Exner, R.; Wessner, B.; Strasser, E.; Spittler, A. Regulative potential of glutamine--relation to glutathione metabolism. Nutrition 2002, 18, 217–221. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, L.-H.; Tsai, W.-C.; Shirakawa, H.; Tsai, Y.-L.; Yeh, S.-L.; Yeh, C.-L. Glutamine Administration Attenuates Poly(I:C)-Induced Lung Injury by Reducing Neutrophil Infiltration and Activating the TLR-3 Antiviral Pathway. Nutrients 2025, 17, 1700. https://doi.org/10.3390/nu17101700
Su L-H, Tsai W-C, Shirakawa H, Tsai Y-L, Yeh S-L, Yeh C-L. Glutamine Administration Attenuates Poly(I:C)-Induced Lung Injury by Reducing Neutrophil Infiltration and Activating the TLR-3 Antiviral Pathway. Nutrients. 2025; 17(10):1700. https://doi.org/10.3390/nu17101700
Chicago/Turabian StyleSu, Li-Han, Wen-Chiuan Tsai, Hitoshi Shirakawa, Yu-Ling Tsai, Sung-Ling Yeh, and Chiu-Li Yeh. 2025. "Glutamine Administration Attenuates Poly(I:C)-Induced Lung Injury by Reducing Neutrophil Infiltration and Activating the TLR-3 Antiviral Pathway" Nutrients 17, no. 10: 1700. https://doi.org/10.3390/nu17101700
APA StyleSu, L.-H., Tsai, W.-C., Shirakawa, H., Tsai, Y.-L., Yeh, S.-L., & Yeh, C.-L. (2025). Glutamine Administration Attenuates Poly(I:C)-Induced Lung Injury by Reducing Neutrophil Infiltration and Activating the TLR-3 Antiviral Pathway. Nutrients, 17(10), 1700. https://doi.org/10.3390/nu17101700