
The Role of G Protein-Coupled Receptors in the Regulation of Orthopaedic Diseases by Gut Microbiota

Abstract
1. Introduction
2. The Relationship Between Gut Microbiota and Orthopaedic Disease
2.1. Gut Microbiota and Rheumatoid Arthritis
2.2. Gut Microbiota and Osteoarthritis
2.3. Gut Microbiota and Osteoporosis
2.4. Gut Microbiota and Bone Metastasis
3. Interaction Between Metabolites of Gut Microbiota with GPCRs
3.1. GPCRs as Key Receptors for Sensing Gut Microbiota Metabolites
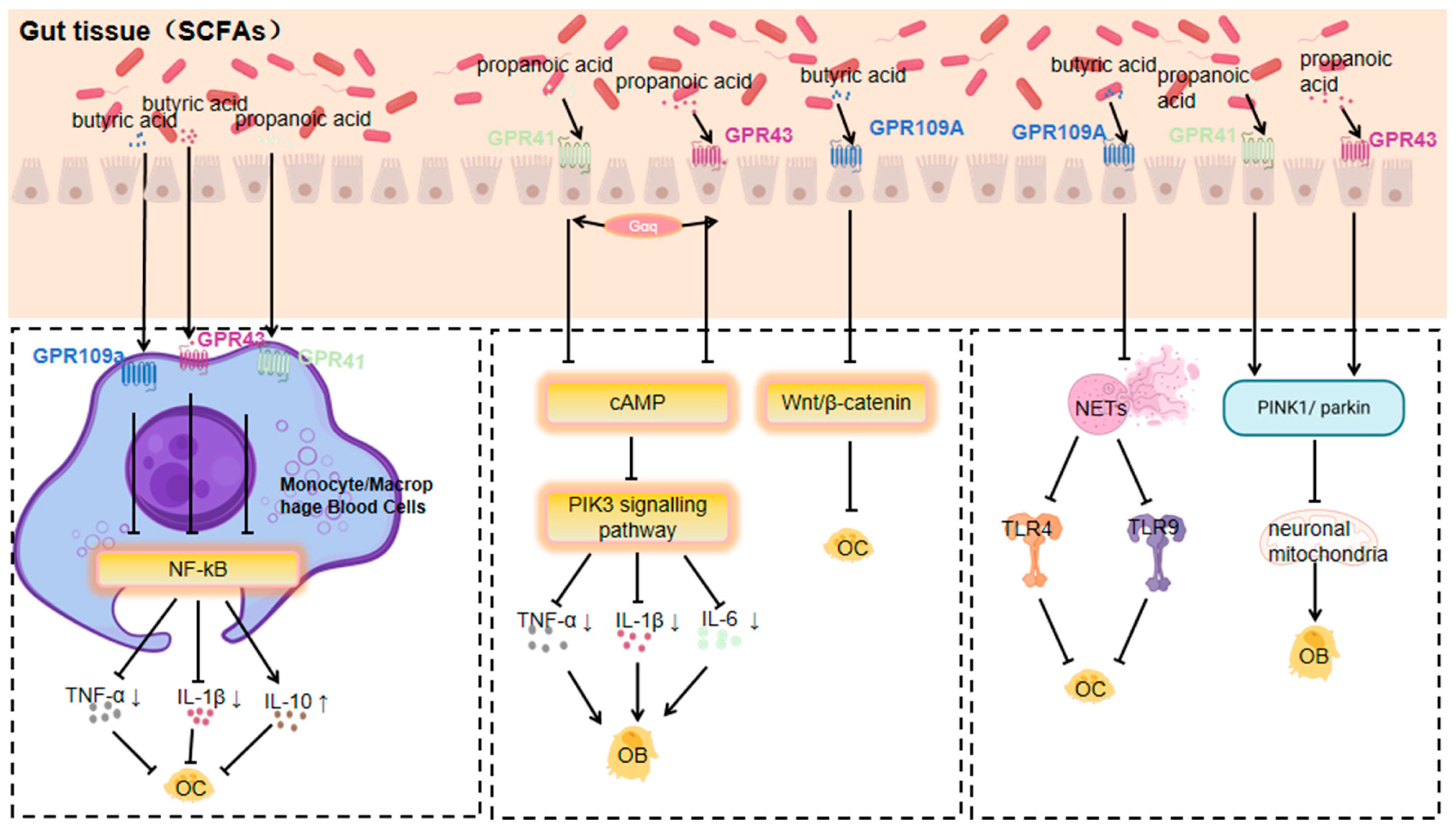
3.1.1. Response of GPR41, GPR43 and GPR109a to SCFAs
3.1.2. TGR5 Response to Bile Acids
3.1.3. GPR81 Response to Lactate
3.1.4. GPR84 Response to MCFAs
3.1.5. Response of GPR91 to Succinate
3.1.6. GPR4 Response to Acids
3.1.7. Response of GPR120 to Medium- and Long-Chain Unsaturated Fatty Acids
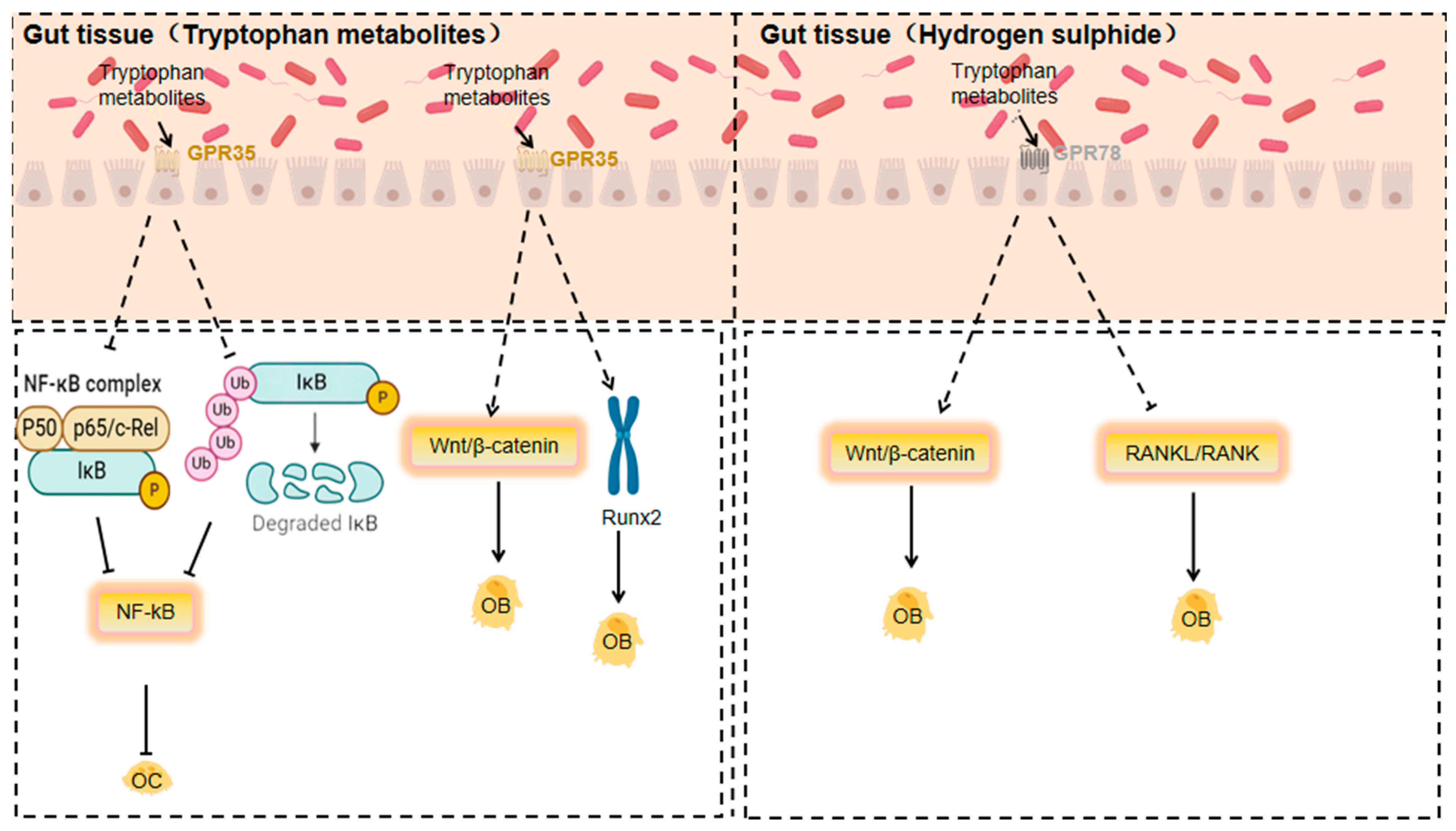
3.1.8. Response of GPR35 to Tryptophan Metabolites
3.1.9. Response of GPR78 to Hydrogen Sulphide
4. Role of Metabolites and GPCRs in Orthopaedic Diseases and Molecular Mechanisms
Signal Transduction Pathways and Their Effects on Orthopaedic Diseases
5. Mechanisms of GPCR Regulation of Gut Microbiota
5.1. Immune Regulation
5.2. Regulation of Bacterial Homeostasis by GPCRs
6. Research Progress of GPCR Activation in Orthopaedic Diseases
6.1. Drug Development
6.2. Exercise Regulates Gut Microbiota and Thus Activates GPCR
6.3. Dietary Modulation of Gut Microbiota and Thus Activation of GPCR
7. Future Research Directions
7.1. New GPCR Discovery
7.2. Strategies for Modulating the Gut Microbiota
7.3. Personalised Therapeutic Strategies
8. Conclusions
- (1)
- In terms of the discovery and function of new GPCRs, the discovery of new GPCRs is expected to provide new avenues for personalised treatment of orthopaedic diseases.
- (2)
- Moderate exercise and dietary interventions affecting the activity of GPCRs through the regulation of gut microbiota are expected to provide new protocols and strategies for personalised treatment of orthopaedic diseases.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AGE | advanced glycosylation end |
| CIA | collagen-induced arthritis |
| DHA | docosahexaenoic acid |
| ERK | extracellular signal-regulated kinase |
| FMT | faecal microbiota transplantation |
| GPCRs | G protein-coupled receptors |
| HA | hippuric acid |
| IGF-1 | insulin-like growth factor 1 |
| IL-6 | interleukin 6 |
| IL-10 | interleukin-10 |
| LPC | lysophosphatidyl choline |
| MAPK | mitogen-activated protein kinase |
| MCFAs | medium-chain fatty acids |
| MMPs | matrix metalloproteinases |
| MM | multiple myeloma |
| micro-CT | micro-computed tomography |
| MSCs | mesenchymal stem cells |
| OA | osteoarthritis |
| OM | osteogenic medium |
| OP | osteoporosis |
| OPN | Osteopontin |
| OPG | Osteoprotegerin |
| PI3K | phosphatidylinositol 3 kinase |
| PLC | phospholipase C |
| PGE2 | Prostaglandin E2 |
| RA | rheumatoid arthritis |
| RNS | reactive nitrogen species |
| SCFAs | short-chain fatty acids |
| SPC | phingosylphosphatidyl choline |
| SPF | specific pathogen-free |
| Tregs | T cells |
| TMAO | Trimethylamine N-oxide |
References
- Safiri, S.; Kolahi, A.A.; Cross, M.; Hill, C.; Smith, E.; Carson-Chahhoud, K.; Mansournia, M.A.; Almasi-Hashiani, A.; Ashrafi-Asgarabad, A.; Kaufman, J.; et al. Prevalence, Deaths, and Disability-Adjusted Life Years Due to Musculoskeletal Disorders for 195 Countries and Territories 1990–2017. Arthritis Rheumatol. 2021, 73, 702–714. [Google Scholar] [CrossRef] [PubMed]
- Salari, N.; Darvishi, N.; Bartina, Y.; Larti, M.; Kiaei, A.; Hemmati, M.; Shohaimi, S.; Mohammadi, M. Global prevalence of osteoporosis among the world older adults: A comprehensive systematic review and meta-analysis. J. Orthop. Surg. Res. 2021, 16, 669. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.-M.; Bisignano, C.; James, S.L.; Abady, G.G.; Abedi, A.; Abu-Gharbieh, E.; Alhassan, R.K.; Alipour, V.; Arabloo, J.; Asaad, M.; et al. Global, regional, and national burden of bone fractures in 204 countries and territories, 1990–2019: A systematic analysis from the Global Burden of Disease Study 2019. Lancet Healthy Longev. 2021, 2, e580–e592. [Google Scholar] [CrossRef] [PubMed]
- Guan, S.Y.; Zheng, J.-X.; Sam, N.B.; Xu, S.; Shuai, Z.; Pan, F. Global burden and risk factors of musculoskeletal disorders among adolescents and young adults in 204 countries and territories, 1990–2019. Autoimmun. Rev. 2023, 22, 103361. [Google Scholar] [CrossRef]
- Cao, F.; Xu, Z.; Li, X.-X.; Fu, Z.-Y.; Han, R.-Y.; Zhang, J.-L.; Wang, P.; Hou, S.; Pan, H.-F. Trends and cross-country inequalities in the global burden of osteoarthritis, 1990–2019: A population-based study. Ageing Res. Rev. 2024, 99, 102382. [Google Scholar] [CrossRef]
- Zhang, X.; Shen, D.; Fang, Z.; Jie, Z.; Qiu, X.; Zhang, C.; Chen, Y.; Ji, L. Human gut microbiota changes reveal the progression of glucose intolerance. PLoS ONE 2013, 8, e71108. [Google Scholar] [CrossRef]
- Rajani, C.; Jia, W. Disruptions in gut microbial-host co-metabolism and the development of metabolic disorders. Clin. Sci. 2018, 132, 791–811. [Google Scholar] [CrossRef]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The Role of Short-Chain Fatty Acids from Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef]
- Fusco, W.; Lorenzo, M.B.; Cintoni, M.; Porcari, S.; Rinninella, E.; Kaitsas, F.; Lener, E.; Mele, M.C.; Gasbarrini, A.; Collado, M.C.; et al. Short-Chain Fatty-Acid-Producing Bacteria: Key Components of the Human Gut Microbiota. Nutrients 2023, 15, 2211. [Google Scholar] [CrossRef]
- den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef]
- Wallimann, A.; Magrath, W.; Thompson, K.; Moriarty, T.; Richards, R.; Akdis, C.; O’Mahony, L.; Hernandez, C.J. Gut microbial-derived short-chain fatty acids and bone: A potential role in fracture healing. Eur. Cells Mater. 2021, 41, 454–470. [Google Scholar] [CrossRef] [PubMed]
- Said, B.; Charlie, L.; Getachew, E.; Wanjiru, C.L.; Abebe, M.; Manyazewal, T. Molecular bacterial load assay versus culture for monitoring treatment response in adults with tuberculosis. SAGE Open Med. 2021, 9, 20503121211033470. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Rosen, C.E.; González-Hernández, J.A.; Song, D.; Potempa, J.; Ring, A.M.; Palm, N.W. Highly multiplexed bioactivity screening reveals human and microbiota metabolome-GPCRome interactions. Cell 2023, 186, 3095–3110.e19. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.J.; Goldsworthy, S.M.; Barnes, A.A.; Eilert, M.M.; Tcheang, L.; Daniels, D.; Muir, A.I.; Wigglesworth, M.J.; Kinghorn, I.; Fraser, N.J.; et al. The Orphan G protein-coupled receptors GPR41 and GPR43 are activated by propionate and other short chain carboxylic acids. J. Biol. Chem. 2003, 278, 11312–11319. [Google Scholar] [CrossRef]
- Hauser, A.S.; Chavali, S.; Masuho, I.; Jahn, L.J.; Martemyanov, K.A.; Gloriam, D.E.; Babu, M.M. Pharmacogenomics of GPCR Drug Targets. Cell 2018, 172, 41–54.e19. [Google Scholar] [CrossRef]
- Wang, X.; Yuan, W.; Yang, C.; Wang, Z.; Zhang, J.; Xu, D.; Sun, X.; Sun, W. Emerging role of gut microbiota in autoimmune diseases. Front. Immunol. 2024, 15, 1365554. [Google Scholar] [CrossRef]
- Ley, R.E.; Peterson, D.A.; Gordon, J.I. Ecological and evolutionary forces shaping microbial diversity in the human intestine. Cell 2006, 124, 837–848. [Google Scholar] [CrossRef]
- Belkaid, Y.; Hand, T.W. Role of the microbiota in immunity and inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef]
- Sender, R.; Fuchs, S.; Milo, R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef]
- Ma, F.; Li, Z.; Liu, H.; Chen, S.; Zheng, S.; Zhu, J.; Shi, H.; Ye, H.; Qiu, Z.; Gao, L.; et al. Dietary-timing-induced gut microbiota diurnal oscillations modulate inflammatory rhythms in rheumatoid arthritis. Cell Metab. 2024, 36, 2367–2382.e5. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, S.-X.; Chang, M.-J.; Qiao, J.; Wang, C.-H.; Li, X.-F.; Yu, Q.; He, P.-F. Characteristics of the Gut Microbiome and Its Relationship with Peripheral CD4(+) T Cell Subpopulations and Cytokines in Rheumatoid Arthritis. Front. Microbiol. 2022, 13, 799602. [Google Scholar] [CrossRef] [PubMed]
- Round, J.L.; Lee, S.M.; Li, J.; Tran, G.; Jabri, B.; Chatila, T.A.; Mazmanian, S.K. The Toll-like receptor 2 pathway establishes colonization by a commensal of the human microbiota. Science 2011, 332, 974–977. [Google Scholar] [CrossRef]
- Telesford, K.M.; Yan, W.; Ochoa-Reparaz, J.; Pant, A.; Kircher, C.; Christy, M.A.; Begum-Haque, S.; Kasper, D.L.; Kasper, L.H. A commensal symbiotic factor derived from Bacteroides fragilis promotes human CD39(+)Foxp3(+) T cells and Treg function. Gut Microbes 2015, 6, 234–242. [Google Scholar] [CrossRef]
- Mei, L.; Yang, Z.; Zhang, X.; Liu, Z.; Wang, M.; Wu, X.; Chen, X.; Huang, Q.; Huang, R. Sustained Drug Treatment Alters the Gut Microbiota in Rheumatoid Arthritis. Front. Immunol. 2021, 12, 704089. [Google Scholar] [CrossRef]
- Scher, J.U.; Sczesnak, A.; Longman, R.S.; Segata, N.; Ubeda, C.; Bielski, C.; Rostron, T.; Cerundolo, V.; Pamer, E.G.; Abramson, S.B.; et al. Expansion of intestinal Prevotella copri correlates with enhanced susceptibility to arthritis. eLife 2013, 2, e01202. [Google Scholar] [CrossRef]
- Maeda, Y.; Takeda, K. Role of Gut Microbiota in Rheumatoid Arthritis. J. Clin. Med. 2017, 6, 60. [Google Scholar] [CrossRef]
- Granfors, K.; Isomäki, H.; Von Essen, R.; Maatela, J.; Kalliomäki, J.L.; Toivanen, A. Yersinia antibodies in inflammatory joint diseases. Clin. Exp. Rheumatol. 1983, 1, 215–218. [Google Scholar]
- Chen, J.; Wright, K.; Davis, J.M.; Jeraldo, P.; Marietta, E.V.; Murray, J.; Nelson, H.; Matteson, E.L.; Taneja, V. An expansion of rare lineage intestinal microbes characterizes rheumatoid arthritis. Genome Med. 2016, 8, 43. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Peng, F.; Wei, X.; Hao, H.; Li, W.; Zhao, Y. Cedrol from ginger alleviates rheumatoid arthritis through dynamic regulation of intestinal microenvironment. Food Funct. 2022, 13, 11825–11839. [Google Scholar] [CrossRef]
- Blenkinsopp, H.C.; Seidler, K.; Barrow, M. Microbial Imbalance and Intestinal Permeability in the Pathogenesis of Rheumatoid Arthritis: A Mechanism Review with a Focus on Bacterial Translocation, Citrullination, and Probiotic Intervention. J. Am. Nutr. Assoc. 2024, 43, 59–76. [Google Scholar] [CrossRef]
- Chen, C.; Zhang, Y.; Yao, X.; Li, S.; Wang, G.; Huang, Y.; Yang, Y.; Zhang, A.; Liu, C.; Zhu, D.; et al. Characterizations of the Gut Bacteriome, Mycobiome, and Virome in Patients with Osteoarthritis. Microbiol. Spectr. 2023, 11, e0171122. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Chen, J.; Li, B.; Zeng, B.; Chou, C.-H.; Zheng, X.; Xie, J.; Li, H.; Hao, Y.; Chen, G.; et al. Faecal microbiota transplantation from metabolically compromised human donors accelerates osteoarthritis in mice. Ann. Rheum. Dis. 2020, 79, 646–656. [Google Scholar] [CrossRef]
- Wei, Z.; Li, F.; Pi, G. Association Between Gut Microbiota and Osteoarthritis: A Review of Evidence for Potential Mechanisms and Therapeutics. Front. Cell. Infect. Microbiol. 2022, 12, 812596. [Google Scholar] [CrossRef]
- Mendez, M.E.; Murugesh, D.K.; Christiansen, B.A.; Loots, G.G. Antibiotic Treatment Prior to Injury Abrogates the Detrimental Effects of LPS in STR/ort Mice Susceptible to Osteoarthritis Development. JBMR Plus 2023, 7, e10759. [Google Scholar] [CrossRef]
- Jiang, S.; Shen, B. Research progress on the relationship between gut microbiota dysbiosis and osteoarthritis. Zhongguo Xiu Fu Chong Jian Wai Ke Za Zhi 2023, 37, 371–376. [Google Scholar]
- Marchese, L.; Contartese, D.; Giavaresi, G.; Di Sarno, L.; Salamanna, F. The Complex Interplay between the Gut Microbiome and Osteoarthritis: A Systematic Review on Potential Correlations and Therapeutic Approaches. Int. J. Mol. Sci. 2024, 25, 143. [Google Scholar] [CrossRef]
- Longo, U.G.; Lalli, A.; Bandini, B.; de Sire, R.; Angeletti, S.; Lustig, S.; Ammendolia, A.; Budhiparama, N.C.; de Sire, A. Role of the Gut Microbiota in Osteoarthritis, Rheumatoid Arthritis, and Spondylarthritis: An Update on the Gut-Joint Axis. Int. J. Mol. Sci. 2024, 25, 3242. [Google Scholar] [CrossRef]
- Manafi, S.; Mirjalili, F.; Reshadi, R. Synthesis and evaluation of the bioactivity of fluorapatite-45S5 bioactive glass nanocomposite. Prog. Biomater. 2019, 8, 77–89. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Y.; Gao, W.; Wang, B.; Zhao, H.; Zeng, Y.; Ji, Y.; Hao, D. Diversity analysis of gut microbiota in osteoporosis and osteopenia patients. PeerJ 2017, 5, e3450. [Google Scholar] [CrossRef]
- Ling, C.W.; Miao, Z.; Xiao, M.-L.; Zhou, H.; Jiang, Z.; Fu, Y.; Xiong, F.; Zuo, L.-S.; Liu, Y.-P.; Wu, Y.-Y.; et al. The Association of Gut Microbiota with Osteoporosis Is Mediated by Amino Acid Metabolism: Multiomics in a Large Cohort. J. Clin. Endocrinol. Metab. 2021, 106, e3852–e3864. [Google Scholar] [CrossRef]
- Yan, J.; Herzog, J.W.; Tsang, K.; Brennan, C.A.; Bower, M.A.; Garrett, W.S.; Sartor, B.R.; Aliprantis, A.O.; Charles, J.F. Gut microbiota induce IGF-1 and promote bone formation and growth. Proc. Natl. Acad. Sci. USA 2016, 113, E7554–E7563. [Google Scholar] [CrossRef]
- Mikkelsen, K.H.; Vilsbøll, T.; Holst, J.J.; Hartmann, B.; Knop, F.K.; Frost, M. No changes in levels of bone formation and resorption markers following a broad-spectrum antibiotic course. BMC Endocr. Disord. 2018, 18, 60. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, J.; Xue, Q.; Liu, B.; Qin, R.; Li, Y.; Qiu, Y.; Wang, R.; Goltzman, D.; Miao, D.; et al. Pyrroloquinoline quinone alleviates natural aging-related osteoporosis via a novel MCM3-Keap1-Nrf2 axis-mediated stress response and Fbn1 upregulation. Aging Cell 2023, 22, e13912. [Google Scholar] [CrossRef] [PubMed]
- Qin, Q.; Yan, S.; Yang, Y.; Chen, J.; Yan, H.; Li, T.; Gao, X.; Wang, Y.; Li, A.; Wang, S.; et al. The Relationship Between Osteoporosis and Intestinal Microbes in the Henan Province of China. Front. Cell Dev. Biol. 2021, 9, 752990. [Google Scholar] [CrossRef]
- Xu, Z.; Xie, Z.; Sun, J.; Huang, S.; Chen, Y.; Li, C.; Sun, X.; Xia, B.; Tian, L.; Guo, C.; et al. Gut Microbiome Reveals Specific Dysbiosis in Primary Osteoporosis. Front. Cell. Infect. Microbiol. 2020, 10, 160. [Google Scholar] [CrossRef]
- Huang, R.; Liu, P.; Bai, Y.; Huang, J.; Pan, R.; Li, H.; Su, Y.; Zhou, Q.; Ma, R.; Zong, S.; et al. Changes in the gut microbiota of osteoporosis patients based on 16S rRNA gene sequencing: A systematic review and meta-analysis. J. Zhejiang Univ. Sci. B 2022, 23, 1002–1013. [Google Scholar] [CrossRef]
- Zhang, Y.W.; Cao, M.-M.; Li, Y.-J.; Chen, X.-X.; Yu, Q.; Rui, Y.-F. A narrative review of the moderating effects and repercussion of exercise intervention on osteoporosis: Ingenious involvement of gut microbiota and its metabolites. J. Transl. Med. 2022, 20, 490. [Google Scholar] [CrossRef]
- Sevcikova, A.; Martiniakova, M.; Omelka, R.; Stevurkova, V.; Ciernikova, S. The Link Between the Gut Microbiome and Bone Metastasis. Int. J. Mol. Sci. 2024, 25, 12086. [Google Scholar] [CrossRef]
- Jian, X.; Zhu, Y.; Ouyang, J.; Wang, Y.; Lei, Q.; Xia, J.; Guan, Y.; Zhang, J.; Guo, J.; He, Y.; et al. Alterations of gut microbiome accelerate multiple myeloma progression by increasing the relative abundances of nitrogen-recycling bacteria. Microbiome 2020, 8, 74. [Google Scholar] [CrossRef]
- Nasr, R.; Shamseddine, A.; Mukherji, D.; Nassar, F.; Temraz, S. The Crosstalk between Microbiome and Immune Response in Gastric Cancer. Int. J. Mol. Sci. 2020, 21, 6586. [Google Scholar] [CrossRef]
- Qasem, H.H.; El-Sayed, W.M. The bacterial microbiome and cancer: Development, diagnosis, treatment, and future directions. Clin. Exp. Med. 2024, 25, 12. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, T.W.; Mimpen, I.L.; Traets, J.J.; van Hoeck, A.; Zeverijn, L.J.; Geurts, B.S.; de Wit, G.F.; Noë, M.; Hofland, I.; Vos, J.L.; et al. A pan-cancer analysis of the microbiome in metastatic cancer. Cell 2024, 187, 2324–2335.e19. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, T.; Zheng, B.; Lu, Y.; Liang, Y.; Xu, G.; Zhao, L.; Tao, Y.; Song, Q.; You, H.; et al. Lymphotoxin-beta promotes breast cancer bone metastasis colonization and osteolytic outgrowth. Nat. Cell Biol. 2024, 26, 1597–1612. [Google Scholar] [CrossRef]
- Yuan, L.; Pan, L.; Wang, Y.; Zhao, J.; Fang, L.; Zhou, Y.; Xia, R.; Ma, Y.; Jiang, Z.; Xu, Z.; et al. Characterization of the landscape of the intratumoral microbiota reveals that Streptococcus anginosus increases the risk of gastric cancer initiation and progression. Cell Discov. 2024, 10, 117. [Google Scholar] [CrossRef] [PubMed]
- Zeng, R.; Gou, H.; Lau, H.C.H.; Yu, J. Stomach microbiota in gastric cancer development and clinical implications. Gut 2024, 73, 2062–2073. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, D.; Chen, S. Unveiling the gastric microbiota: Implications for gastric carcinogenesis, immune responses, and clinical prospects. J. Exp. Clin. Cancer Res. 2024, 43, 118. [Google Scholar] [CrossRef]
- Doocey, C.M.; Finn, K.; Murphy, C.; Guinane, C.M. The impact of the human microbiome in tumorigenesis, cancer progression, and biotherapeutic development. BMC Microbiol. 2022, 22, 53. [Google Scholar] [CrossRef]
- Han, E.J.; Ahn, J.-S.; Choi, Y.-J.; Kim, D.-H.; Choi, J.-S.; Chung, H.-J. Exploring the gut microbiome: A potential biomarker for cancer diagnosis, prognosis, and therapy. Biochim. Biophys. Acta Rev. Cancer 2024, 1880, 189251. [Google Scholar] [CrossRef]
- Yu, D.; Du, J.; Pu, X.; Zheng, L.; Chen, S.; Wang, N.; Li, J.; Chen, S.; Pan, S.; Shen, B. The Gut Microbiome and Metabolites Are Altered and Interrelated in Patients with Rheumatoid Arthritis. Front. Cell. Infect. Microbiol. 2021, 11, 763507. [Google Scholar] [CrossRef]
- Peng, J.; Lu, X.; Xie, K.; Xu, Y.; He, R.; Guo, L.; Han, Y.; Wu, S.; Dong, X.; Lu, Y.; et al. Dynamic Alterations in the Gut Microbiota of Collagen-Induced Arthritis Rats Following the Prolonged Administration of Total Glucosides of Paeony. Front. Cell. Infect. Microbiol. 2019, 9, 204. [Google Scholar] [CrossRef]
- Majidi, M.; Heidarnejad, F.; Naseri, M.; Bonakdar, S.; Salimi, M.; Yaraee, R. Marham-Mafasel decrease joint inflammation and IL-1beta gene expression in rheumatoid arthritis animal model. Vet. Med. Sci. 2021, 7, 1417–1425. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yu, Y.; Liao, J.; Hu, W.; Bian, X.; Wu, J.; Zhu, Y.Z. S-Propargyl-Cysteine Remodels the Gut Microbiota to Alleviate Rheumatoid Arthritis by Regulating Bile Acid Metabolism. Front. Cell. Infect. Microbiol. 2021, 11, 670593. [Google Scholar] [CrossRef] [PubMed]
- Lai, W.; Wang, C.; Lai, R.; Peng, X.; Luo, J. Lycium barbarum polysaccharide modulates gut microbiota to alleviate rheumatoid arthritis in a rat model. NPJ Sci. Food 2022, 6, 34. [Google Scholar] [CrossRef]
- Niu, S.; Zhu, X.; Zhang, J.; Ma, Y.; Lang, X.; Luo, L.; Li, W.; Zhao, Y.; Zhang, Z. Arsenic trioxide modulates the composition and metabolic function of the gut microbiota in a mouse model of rheumatoid arthritis. Int. Immunopharmacol. 2022, 111, 109159. [Google Scholar] [CrossRef]
- Wang, X.; Wu, Y.; Liu, Y.; Chen, F.; Chen, S.; Zhang, F.; Li, S.; Wang, C.; Gong, Y.; Huang, R.; et al. Altered gut microbiome profile in patients with knee osteoarthritis. Front. Microbiol. 2023, 14, 1153424. [Google Scholar] [CrossRef]
- Schott, E.M.; Farnsworth, C.W.; Grier, A.; Lillis, J.A.; Soniwala, S.; Dadourian, G.H.; Bell, R.D.; Doolittle, M.L.; Villani, D.A.; Awad, H.; et al. Targeting the gut microbiome to treat the osteoarthritis of obesity. JCI Insight 2018, 3, e95997. [Google Scholar] [CrossRef]
- Stevens, C.; Norris, S.; Arbeeva, L.; Carter, S.; Enomoto, M.; Nelson, A.E.; Lascelles, B.D.X. Gut Microbiome and Osteoarthritis: Insights From the Naturally Occurring Canine Model of Osteoarthritis. Arthritis Rheumatol. 2024, 76, 1758–1763. [Google Scholar] [CrossRef]
- Liu, J.H.; Chen, C.; Liu, Z.; Luo, Z.; Rao, S.; Jin, L.; Wan, T.; Yue, T.; Tan, Y.; Yin, H.; et al. Extracellular Vesicles from Child Gut Microbiota Enter into Bone to Preserve Bone Mass and Strength. Adv. Sci. 2021, 8, 2004831. [Google Scholar] [CrossRef]
- Guo, M.; Liu, H.; Yu, Y.; Zhu, X.; Xie, H.; Wei, C.; Mei, C.; Shi, Y.; Zhou, N.; Qin, K.; et al. Lactobacillus rhamnosus GG ameliorates osteoporosis in ovariectomized rats by regulating the Th17/Treg balance and gut microbiota structure. Gut Microbes 2023, 15, 2190304. [Google Scholar] [CrossRef]
- Wenhui, Y.; Zhongyu, X.; Kai, C.; Zhaopeng, C.; Jinteng, L.; Mengjun, M.; Zepeng, S.; Yunshu, C.; Peng, W.; Yanfeng, W.; et al. Variations in the Gut Microbiota in Breast Cancer Occurrence and Bone Metastasis. Front. Microbiol. 2022, 13, 894283. [Google Scholar] [CrossRef]
- Dutta, R.K.; Abu, Y.F.; Tao, J.; Chupikova, I.; Oleas, J.; Singh, P.K.; Vitari, N.A.; Qureshi, R.; Ramakrishnan, S.; Roy, S. Altered gut microbiome drives heightened pain sensitivity in a murine model of metastatic triple-negative breast cancer. Am. J. Cancer Res. 2024, 14, 274–299. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Kim, M.T.; Han, J.H. GPR41 and GPR43: From development to metabolic regulation. Biomed. Pharmacother. 2024, 175, 116735. [Google Scholar] [CrossRef]
- Katono, T.; Kawato, T.; Tanabe, N.; Suzuki, N.; Iida, T.; Morozumi, A.; Ochiai, K.; Maeno, M. Sodium butyrate stimulates mineralized nodule formation and osteoprotegerin expression by human osteoblasts. Arch. Oral Biol. 2008, 53, 903–909. [Google Scholar] [CrossRef]
- Tian, H.; Chen, F.; Wang, Y.; Liu, Y.; Ma, G.; Zhao, Y.; Ma, Y.; Tian, T.; Ma, R.; Yu, Y.; et al. Nur77 Prevents Osteoporosis by Inhibiting the NF-kappaB Signalling Pathway and Osteoclast Differentiation. J. Cell. Mol. Med. 2022, 26, 2163–2176. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, C.; Yuan, B.; Liu, L.; Zhang, H.; Zhu, M.; Chai, H.; Peng, J.; Huang, Y.; Zhou, S.; et al. Akkermansia muciniphila and its metabolite propionic acid maintains neuronal mitochondrial division and autophagy homeostasis during Alzheimer’s disease pathologic process via GPR41 and GPR43. Microbiome 2025, 13, 16. [Google Scholar] [CrossRef]
- Zhao, N.; Zhang, X.; Li, B.; Wang, J.; Zhang, C.; Xu, B. Treadmill Exercise Improves PINK1/Parkin-Mediated Mitophagy Activity Against Alzheimer’s Disease Pathologies by Upregulated SIRT1-FOXO1/3 Axis in APP/PS1 Mice. Mol. Neurobiol. 2023, 60, 277–291. [Google Scholar] [CrossRef]
- Geisler, C.E.; Miller, K.E.; Ghimire, S.; Renquist, B.J. The Role of GPR109a Signaling in Niacin Induced Effects on Fed and Fasted Hepatic Metabolism. Int. J. Mol. Sci. 2021, 22, 4001. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, J.; Zhang, M.; Li, B.; Pan, X.; Dong, X.; Pan, L.; Sun, J. GPR109a Regulates Phenotypic and Functional Alterations in Macrophages and the Progression of Type 1 Diabetes. Mol. Nutr. Food Res. 2022, 66, e2200300. [Google Scholar] [CrossRef]
- Gong, Y.; Jin, X.; Yuan, B.; Lv, Y.; Yan, G.; Liu, M.; Xie, C.; Liu, J.; Tang, Y.; Gao, H.; et al. G Protein-Coupled Receptor 109A Maintains the Intestinal Integrity and Protects Against ETEC Mucosal Infection by Promoting IgA Secretion. Front. Immunol. 2020, 11, 583652. [Google Scholar] [CrossRef]
- Chen, J.R.; Zhao, H.; Wankhade, U.D.; Chintapalli, S.V.; Li, C.; Gai, D.; Shankar, K.; Zhan, F.; Lazarenko, O.P. GPR109A mediates the effects of hippuric acid on regulating osteoclastogenesis and bone resorption in mice. Commun. Biol. 2021, 4, 53. [Google Scholar] [CrossRef]
- Chen, J.R.; Lazarenko, O.P.; Blackburn, M.L. GPR109A gene deletion ameliorates gonadectomy-induced bone loss in mice. Bone 2022, 161, 116422. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Gong, Q.; Zong, X.; Wu, D.; Li, Y.; Xiao, H.; Song, J.; Zhang, S.; Fu, S.; Feng, Z.; et al. GPR109A controls neutrophil extracellular traps formation and improve early sepsis by regulating ROS/PAD4/Cit-H3 signal axis. Exp. Hematol. Oncol. 2023, 12, 15. [Google Scholar] [CrossRef]
- Smith, P.M.; Howitt, M.R.; Panikov, N.; Michaud, M.; Gallini, C.A.; Bohlooly, Y.M.; Glickman, J.N.; Garrett, W.S. The microbial metabolites, short-chain fatty acids, regulate colonic Treg cell homeostasis. Science 2013, 341, 569–573. [Google Scholar] [CrossRef]
- Deutschmann, K.; Reich, M.; Klindt, C.; Dröge, C.; Spomer, L.; Häussinger, D.; Keitel, V. Bile acid receptors in the biliary tree: TGR5 in physiology disease. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864 Pt 4, 1319–1325. [Google Scholar] [CrossRef]
- He, S.; Li, L.; Yao, Y.; Su, J.; Lei, S.; Zhang, Y.; Zeng, H. Bile acid and its bidirectional interactions with gut microbiota: A review. Crit. Rev. Microbiol. 2024, 50, 684–701. [Google Scholar] [CrossRef]
- Jia, W.; Li, Y.; Cheung, K.C.P.; Zheng, X. Bile acid signaling in the regulation of whole body metabolic and immunological homeostasis. Sci. China Life Sci. 2024, 67, 865–878. [Google Scholar] [CrossRef]
- Li, Z.; Huang, J.; Wang, F.; Li, W.; Wu, X.; Zhao, C.; Zhao, J.; Wei, H.; Wu, Z.; Qian, M.; et al. Dual Targeting of Bile Acid Receptor-1 (TGR5) and Farnesoid X Receptor (FXR) Prevents Estrogen-Dependent Bone Loss in Mice. J. Bone Miner. Res. 2019, 34, 765–776. [Google Scholar] [CrossRef]
- Guo, C.; Chen, W.D.; Wang, Y.D. TGR5, Not Only a Metabolic Regulator. Front. Physiol. 2016, 7, 646. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, G.; Wang, B.; Yang, H. Activation of TGR5 promotes osteoblastic cell differentiation and mineralization. Biomed. Pharmacother. 2018, 108, 1797–1803. [Google Scholar] [CrossRef]
- Pokhrel, S.; Dilts, M.; Stahl, Z.; Boehme, S.; Frame, G.; Chiang, J.Y.L.; Ferrell, J.M. Tgr5-/- mice are protected from ethanol-induced metabolic alterations through enhanced leptin and Fgf21 signaling. Hepatol. Commun. 2023, 7, e0138. [Google Scholar] [CrossRef]
- Williams, G.A.; Callon, K.E.; Watson, M.; Costa, J.L.; Ding, Y.; Dickinson, M.; Wang, Y.; Naot, D.; Reid, I.R.; Cornish, J. Skeletal phenotype of the leptin receptor-deficient db/db mouse. J. Bone Miner. Res. 2011, 26, 1698–1709. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Kim, T.-Y.; Kim, Y.; Lee, S.-H.; Kim, S.; Kang, S.W.; Yang, J.-Y.; Baek, I.-J.; Sung, Y.H.; Park, Y.-Y.; et al. Microbiota-Derived Lactate Accelerates Intestinal Stem-Cell-Mediated Epithelial Development. Cell Host Microbe 2018, 24, 833–846.e6. [Google Scholar] [CrossRef] [PubMed]
- Dinu, L.D.; Gatea, F.; Israel, F.R.; Lakicevic, M.; Dedović, N.; Vamanu, E. The Modulation Effect of a Fermented Bee Pollen Postbiotic on Cardiovascular Microbiota and Therapeutic Perspectives. Biomedicines 2023, 11, 2712. [Google Scholar] [CrossRef]
- Wu, H.; Mu, C.; Li, X.; Fan, W.; Shen, L.; Zhu, W. Breed-Driven Microbiome Heterogeneity Regulates Intestinal Stem Cell Proliferation via Lactobacillus-Lactate-GPR81 Signaling. Adv. Sci. 2024, 11, e2400058. [Google Scholar] [CrossRef]
- Whitfield, J.F. Leptin: Brains and bones. Expert. Opin. Investig. Drugs 2001, 10, 1617–1622. [Google Scholar] [CrossRef]
- Ranganathan, P.; Shanmugam, A.; Swafford, D.; Suryawanshi, A.; Bhattacharjee, P.; Hussein, M.S.; Koni, P.A.; Prasad, P.D.; Kurago, Z.B.; Thangaraju, M.; et al. GPR81, a Cell-Surface Receptor for Lactate, Regulates Intestinal Homeostasis and Protects Mice from Experimental Colitis. J. Immunol. 2018, 200, 1781–1789. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, M.; Zhang, K.; Li, Y.; Xu, M.; Tang, S.; Qu, X.; Li, C. Lactate enhanced the effect of parathyroid hormone on osteoblast differentiation via GPR81-PKC-Akt signaling. Biochem. Biophys. Res. Commun. 2018, 503, 737–743. [Google Scholar] [CrossRef]
- van den Bosch, M.H.; Blom, A.B.; van de Loo, F.A.; Koenders, M.I.; Lafeber, F.P.; van den Berg, W.B.; van der Kraan, P.M.; van Lent, P.L. Brief Report: Induction of Matrix Metalloproteinase Expression by Synovial Wnt Signaling and Association with Disease Progression in Early Symptomatic Osteoarthritis. Arthritis Rheumatol. 2017, 69, 1978–1983. [Google Scholar] [CrossRef]
- Mohammad, N.M.; Chemtob, S. 3-OBA Is Not an Antagonist of GPR81. Front. Pharmacol. 2021, 12, 803907. [Google Scholar] [CrossRef]
- Jia, M.; Zhang, Y.; Gao, Y.; Ma, X. Effects of Medium Chain Fatty Acids on Intestinal Health of Monogastric Animals. Curr. Protein Pept. Sci. 2020, 21, 777–784. [Google Scholar] [CrossRef]
- Hill, J.O.; Peters, J.C.; Yang, D.; Sharp, T.; Kaler, M.; Abumrad, N.N.; Greene, H.L. Thermogenesis in humans during overfeeding with medium-chain triglycerides. Metabolism 1989, 38, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Bach, A.C.; Ingenbleek, Y.; Frey, A. The usefulness of dietary medium-chain triglycerides in body weight control: Fact or fancy? J. Lipid Res. 1996, 37, 708–726. [Google Scholar] [CrossRef] [PubMed]
- Van Wymelbeke, V.; Himaya, A.; Louis-Sylvestre, J.; Fantino, M. Influence of medium-chain and long-chain triacylglycerols on the control of food intake in men. Am. J. Clin. Nutr. 1998, 68, 226–234. [Google Scholar] [CrossRef]
- St-Onge, M.P.; Ross, R.; Parsons, W.D.; Jones, P.J. Medium-chain triglycerides increase energy expenditure and decrease adiposity in overweight men. Obes. Res. 2003, 11, 395–402. [Google Scholar] [CrossRef]
- Puengel, T.; De Vos, S.; Hundertmark, J.; Kohlhepp, M.; Guldiken, N.; Pujuguet, P.; Auberval, M.; Marsais, F.; Shoji, K.F.; Saniere, L.; et al. The Medium-Chain Fatty Acid Receptor GPR84 Mediates Myeloid Cell Infiltration Promoting Steatohepatitis and Fibrosis. J. Clin. Med. 2020, 9, 1140. [Google Scholar] [CrossRef]
- Wang, S.W.; Zhang, Q.; Lu, D.; Fang, Y.-C.; Yan, X.-C.; Chen, J.; Xia, Z.-K.; Yuan, Q.-T.; Chen, L.-H.; Zhang, Y.-M.; et al. GPR84 regulates pulmonary inflammation by modulating neutrophil functions. Acta Pharmacol. Sin. 2023, 44, 1665–1675. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, Y.; Supekar, S.; Cao, X.; Zhou, J.; Dang, J.; Chen, S.; Jenkins, L.; Marsango, S.; Li, X.; et al. Pro-phagocytic function and structural basis of GPR84 signaling. Nat. Commun. 2023, 14, 5706. [Google Scholar] [CrossRef]
- Jian, L.; Shi-Wei, L.; Dan, J.; Juan, W.; Wei, Z. GPR84 potently inhibits osteoclastogenesis and alleviates osteolysis in bone metastasis of colorectal cancer. J. Orthop. Surg. Res. 2023, 18, 3. [Google Scholar] [CrossRef]
- Guarnieri, M.T.; Chou, Y.-C.; Salvachúa, D.; Mohagheghi, A.; John, P.C.S.; Peterson, D.J.; Bomble, Y.J.; Beckham, G.T. Metabolic Engineering of Actinobacillus succinogenes Provides Insights into Succinic Acid Biosynthesis. Appl. Environ. Microbiol. 2017, 83, e00996-17. [Google Scholar] [CrossRef]
- Zhu, F.; Wang, C.; San, K.Y.; Bennett, G.N. Metabolic engineering of Escherichia coli to produce succinate from woody hydrolysate under anaerobic conditions. J. Ind. Microbiol. Biotechnol. 2020, 47, 223–232. [Google Scholar] [CrossRef]
- De Vadder, F.; Kovatcheva-Datchary, P.; Zitoun, C.; Duchampt, A.; Bäckhed, F.; Mithieux, G. Microbiota-Produced Succinate Improves Glucose Homeostasis via Intestinal Gluconeogenesis. Cell Metab. 2016, 24, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Mao, M.; Zhang, Y.; Yu, K.; Zhu, W. Succinate Modulates Intestinal Barrier Function and Inflammation Response in Pigs. Biomolecules 2019, 9, 486. [Google Scholar] [CrossRef] [PubMed]
- Van der Meulen, R.; Adriany, T.; Verbrugghe, K.; De Vuyst, L. Kinetic analysis of bifidobacterial metabolism reveals a minor role for succinic acid in the regeneration of NAD+ through its growth-associated production. Appl. Environ. Microbiol. 2006, 72, 5204–5210. [Google Scholar] [CrossRef]
- Xu, J.; Yang, Y.; Li, X.; Ding, S.; Zheng, L.; Xiong, C.; Yang, Y. Pleiotropic activities of succinate: The interplay between gut microbiota and cardiovascular diseases. iMETA 2023, 2, e124. [Google Scholar] [CrossRef]
- Ariza, A.C.; Deen, P.M.; Robben, J.H. The succinate receptor as a novel therapeutic target for oxidative and metabolic stress-related conditions. Front. Endocrinol. 2012, 3, 22. [Google Scholar] [CrossRef]
- Littlewood-Evans, A.; Sarret, S.; Apfel, V.; Loesle, P.; Dawson, J.; Zhang, J.; Muller, A.; Tigani, B.; Kneuer, R.; Patel, S.; et al. GPR91 senses extracellular succinate released from inflammatory macrophages and exacerbates rheumatoid arthritis. J. Exp. Med. 2016, 213, 1655–1662. [Google Scholar] [CrossRef]
- Li, L.; Dong, Y.-L.; Liu, T.; Luo, D.; Wei, C.; Shi, W.-Y. Increased succinate receptor GPR91 involved in the pathogenesis of Mooren’s ulcer. Int. J. Ophthalmol. 2018, 11, 1733–1740. [Google Scholar]
- Rios-Covian, D.; Arboleya, S.; Hernandez-Barranco, A.M.; Alvarez-Buylla, J.R.; Ruas-Madiedo, P.; Gueimonde, M.; de los Reyes-Gavilan, C.G. Interactions between Bifidobacterium and Bacteroides species in cofermentations are affected by carbon sources, including exopolysaccharides produced by bifidobacteria. Appl. Environ. Microbiol. 2013, 79, 7518–7524. [Google Scholar] [CrossRef]
- Stalewski, J.; Shih, A.Y.; Papazyan, R.; Ramirez, J.; Ibanez, G.; Hsiao, P.; Yue, Y.; Yin, J.; Badger, C.; Wu, S.; et al. pH Dependence of a GPR4 Selective Antagonist Hampers Its Therapeutic Potential. J. Pharmacol. Exp. Ther. 2023, 386, 35–44. [Google Scholar] [CrossRef]
- Zha, X.; Liu, X.; Wei, M.; Huang, H.; Cao, J.; Liu, S.; Bian, X.; Zhang, Y.; Xiao, F.; Xie, Y.; et al. Microbiota-derived lysophosphatidylcholine alleviates Alzheimer’s disease pathology via suppressing ferroptosis. Cell Metab. 2025, 37, 169–186.e9. [Google Scholar] [CrossRef]
- Bäckhed, F.; Roswall, J.; Peng, Y.; Feng, Q.; Jia, H.; Kovatcheva-Datchary, P.; Li, Y.; Xia, Y.; Xie, H.; Zhong, H.; et al. Dynamics and Stabilization of the Human Gut Microbiome during the First Year of Life. Cell Host Microbe 2015, 17, 690–703. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Guan, Z.; Bi, S.; Wang, F.; He, L.; Niu, X.; You, Y.; Liu, Y.; Ding, Y.; Siwko, S.; et al. The proton-activated G protein-coupled receptor GPR4 regulates the development of osteoarthritis via modulating CXCL12/CXCR7 signaling. Cell Death Dis. 2022, 13, 152. [Google Scholar] [CrossRef]
- Liu, H.; Liu, Y.; Chen, B. Antagonism of GPR4 with NE 52-QQ57 and the Suppression of AGE-Induced Degradation of Type II Collagen in Human Chondrocytes. Chem. Res. Toxicol. 2020, 33, 1915–1921. [Google Scholar] [CrossRef]
- Cani, P.D.; Lecourt, E.; Dewulf, E.M.; Sohet, F.M.; Pachikian, B.D.; Naslain, D.; De Backer, F.; Neyrinck, A.M.; Delzenne, N.M. Gut microbiota fermentation of prebiotics increases satietogenic and incretin gut peptide production with consequences for appetite sensation and glucose response after a meal. Am. J. Clin. Nutr. 2009, 90, 1236–1243. [Google Scholar] [CrossRef]
- Di Paola, M.; De Filippo, C.; Cavalieri, D.; Ramazzotti, M.; Poullet, J.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef]
- Song, X.; Zhang, H.; Zhang, Y.; Goh, B.; Bao, B.; Mello, S.S.; Sun, X.; Zheng, W.; Gazzaniga, F.S.; Wu, M.; et al. Gut microbial fatty acid isomerization modulates intraepithelial T cells. Nature 2023, 619, 837–843. [Google Scholar] [CrossRef]
- Peltomaa, E.; Johnson, M.D.; Taipale, S.J. Marine Cryptophytes Are Great Sources of EPA and DHA. Mar. Drugs 2018, 16, 3. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Kurt, E.; Lbassi, T.; Sa, L.; Xie, D. Biotechnological production of omega-3 fatty acids: Current status and future perspectives. Front. Microbiol. 2023, 14, 1280296. [Google Scholar] [CrossRef]
- Anbazhagan, A.N.; Priyamvada, S.; Gujral, T.; Bhattacharyya, S.; Alrefai, W.A.; Dudeja, P.K.; Borthakur, A. A novel anti-inflammatory role of GPR120 in intestinal epithelial cells. Am. J. Physiol. Cell Physiol. 2016, 310, C612–C621. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, D.; Ho, K.W.; Lin, S.; Suen, W.C.-W.; Zhang, H.; Zha, Z.; Li, G.; Leung, P.S. GPR120 is an important inflammatory regulator in the development of osteoarthritis. Arthritis Res. Ther. 2018, 20, 163. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, H.; Zhang, Z. Recent Advance in Regulatory Effect of GRP120 on Bone Metabolism. Aging Dis. 2023, 14, 1714–1727. [Google Scholar] [CrossRef] [PubMed]
- Kishikawa, A.; Kitaura, H.; Kimura, K.; Ogawa, S.; Qi, J.; Shen, W.-R.; Ohori, F.; Noguchi, T.; Marahleh, A.; Nara, Y.; et al. Docosahexaenoic Acid Inhibits Inflammation-Induced Osteoclast Formation and Bone Resorption in vivo Through GPR120 by Inhibiting TNF-alpha Production in Macrophages and Directly Inhibiting Osteoclast Formation. Front. Endocrinol. 2019, 10, 157. [Google Scholar] [CrossRef] [PubMed]
- Shi, T.; Shi, Y.; Gao, H.; Ma, Y.; Wang, Q.; Shen, S.; Shao, X.; Gong, W.; Chen, X.; Qin, J.; et al. Exercised accelerated the production of muscle-derived kynurenic acid in skeletal muscle and alleviated the postmenopausal osteoporosis through the Gpr35/NFkappaB p65 pathway. J. Orthop. Translat 2022, 35, 1–12. [Google Scholar] [CrossRef]
- Mackenzie, A.E.; Lappin, J.E.; Taylor, D.L.; Nicklin, S.A.; Milligan, G. GPR35 as a Novel Therapeutic Target. Front. Endocrinol. 2011, 2, 68. [Google Scholar] [CrossRef]
- Metges, C.C.; El-Khoury, A.E.; Henneman, L.; Petzke, K.J.; Grant, I.; Bedri, S.; Pereira, P.P.; Ajami, A.M.; Fuller, M.F.; Young, V.R. Availability of intestinal microbial lysine for whole body lysine homeostasis in human subjects. Am. J. Physiol. 1999, 277, E597–E607. [Google Scholar] [CrossRef]
- Wang, J.; Simonavicius, N.; Wu, X.; Swaminath, G.; Reagan, J.; Tian, H.; Ling, L. Kynurenic acid as a ligand for orphan G protein-coupled receptor GPR35. J. Biol. Chem. 2006, 281, 22021–22028. [Google Scholar] [CrossRef]
- Kennedy, P.J.; Cryan, J.F.; Dinan, T.G.; Clarke, G. Kynurenine pathway metabolism the microbiota-gut-brain axis. Neuropharmacology 2017, 112 Pt B, 399–412. [Google Scholar] [CrossRef]
- Cosi, C.; Mannaioni, G.; Cozzi, A.; Carlà, V.; Sili, M.; Cavone, L.; Maratea, D.; Moroni, F. G-protein coupled receptor 35 (GPR35) activation inflammatory pain: Studies on the antinociceptive effects of kynurenic acid zaprinast. Neuropharmacology 2011, 60, 1227–1231. [Google Scholar] [CrossRef]
- Xu, J.; Wu, H.F.; Ang, E.S.; Yip, K.; Woloszyn, M.; Zheng, M.H.; Tan, R.X. NF-kappaB modulators in osteolytic bone diseases. Cytokine Growth Factor Rev. 2009, 20, 7–17. [Google Scholar] [CrossRef]
- Du, J.; Liu, Y.; Wu, X.; Sun, J.; Shi, J.; Zhang, H.; Zheng, A.; Zhou, M.; Jiang, X. BRD9-mediated chromatin remodeling suppresses osteoclastogenesis through negative feedback mechanism. Nat. Commun. 2023, 14, 1413. [Google Scholar] [CrossRef]
- Vimalraj, S.; Arumugam, B.; Miranda, P.J.; Selvamurugan, N. Runx2: Structure, function, and phosphorylation in osteoblast differentiation. Int. J. Biol. Macromol. 2015, 78, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Munteanu, C.; Onose, G.; Poștaru, M.; Turnea, M.; Rotariu, M.; Galaction, A.I. Hydrogen Sulfide and Gut Microbiota: Their Synergistic Role in Modulating Sirtuin Activity and Potential Therapeutic Implications for Neurodegenerative Diseases. Pharmaceuticals 2024, 17, 1480. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Yang, F.; Zhang, X.; Chen, G.; Zou, J.; Yin, L.; Yang, D. Hydrogen sulfide inhibits endoplasmic reticulum stress through the GRP78/mTOR pathway in rat chondrocytes subjected to oxidative stress. Int. J. Mol. Med. 2021, 47, 34. [Google Scholar] [CrossRef]
- Jiang, L.; Wu, Y.; Xu, Z.; Hou, M.; Chen, S.; Cheng, C.; Hu, D.; Lu, D.; Zhu, X.; Li, C. Harnessing hydrogen sulfide in injectable hydrogels that guide the immune response and osteoclastogenesis balance for osteoporosis treatment. Mater. Today Bio 2024, 29, 101338. [Google Scholar] [CrossRef]
- Wang, R. Physiological implications of hydrogen sulfide: A whiff exploration that blossomed. Physiol. Rev. 2012, 92, 791–896. [Google Scholar] [CrossRef]
- Peck, S.C.; Denger, K.; Burrichter, A.; Irwin, S.M.; Balskus, E.P.; Schleheck, D. A glycyl radical enzyme enables hydrogen sulfide production by the human intestinal bacterium Bilophila wadsworthia. Proc. Natl. Acad. Sci. USA 2019, 116, 3171–3176. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, R.; Liu, X.; Zhou, Y.; Qu, C.; Kikuiri, T.; Wang, S.; Zandi, E.; Du, J.; Ambudkar, I.S.; et al. Hydrogen sulfide maintains mesenchymal stem cell function and bone homeostasis via regulation of Ca(2+) channel sulfhydration. Cell Stem Cell 2014, 15, 66–78. [Google Scholar] [CrossRef]
- Kao, R.S.; Abbott, M.J.; Louie, A.; O’Carroll, D.; Lu, W.; Nissenson, R. Constitutive protein kinase A activity in osteocytes and late osteoblasts produces an anabolic effect on bone. Bone 2013, 55, 277–287. [Google Scholar] [CrossRef]
- Luo, J.; Sun, P.; Siwko, S.; Liu, M.; Xiao, J. The role of GPCRs in bone diseases and dysfunctions. Bone Res. 2019, 7, 19. [Google Scholar] [CrossRef]
- Wang, L.; Han, L.; Xue, P.; Hu, X.; Wong, S.-W.; Deng, M.; Tseng, H.C.; Huang, B.-W.; Ko, C.-C. Dopamine suppresses osteoclast differentiation via cAMP/PKA/CREB pathway. Cell. Signal. 2021, 78, 109847. [Google Scholar] [CrossRef]
- Law, N.C.; White, M.F.; Hunzicker-Dunn, M.E. G protein-coupled receptors (GPCRs) That Signal via Protein Kinase A (PKA) Cross-talk at Insulin Receptor Substrate 1 (IRS1) to Activate the phosphatidylinositol 3-kinase (PI3K)/AKT Pathway. J. Biol. Chem. 2016, 291, 27160–27169. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Xu, X.; Zhang, Q.; Yuan, Z.; Tan, B. The PI3K/AKT pathway promotes fracture healing through its crosstalk with Wnt/beta-catenin. Exp. Cell Res. 2020, 394, 112137. [Google Scholar] [CrossRef] [PubMed]
- Naor, Z.; Benard, O.; Seger, R. Activation of MAPK cascades by G-protein-coupled receptors: The case of gonadotropin-releasing hormone receptor. Trends Endocrinol. Metab. 2000, 11, 91–99. [Google Scholar] [CrossRef]
- Greenblatt, M.B.; Shim, J.-H.; Zou, W.; Sitara, D.; Schweitzer, M.; Hu, D.; Lotinun, S.; Sano, Y.; Baron, R.; Park, J.M.; et al. The p38 MAPK pathway is essential for skeletogenesis and bone homeostasis in mice. J. Clin. Investig. 2010, 120, 2457–2473. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, X.; Xue, P.; Ma, X.; Li, J.; Zhang, J. FN1 promotes chondrocyte differentiation and collagen production via TGF-beta/PI3K/Akt pathway in mice with femoral fracture. Gene 2021, 769, 145253. [Google Scholar] [CrossRef]
- Wan, L.; Liu, F.; Wang, A.; He, Y.; Pan, J.; Liu, Y.; Xu, J.; Xu, C.; Wu, F.; Ye, Q. PI3K/Akt pathway-mediated enhancement of bone and vascular regeneration by gelatin/hyaluronic acid/exosome composite scaffold in bone tissue engineering. Biomater. Adv. 2025, 166, 214064. [Google Scholar] [CrossRef]
- Lee, J.; Song, X.; Hyun, B.; Jeon, C.O.; Hyun, S. Drosophila Gut Immune Pathway Suppresses Host Development-Promoting Effects of Acetic Acid Bacteria. Mol. Cells 2023, 46, 637–653. [Google Scholar] [CrossRef]
- Maslowski, K.M.; Vieira, A.T.; Ng, A.; Kranich, J.; Sierro, F.; Yu, D.; Schilter, H.C.; Rolph, M.S.; Mackay, F.; Artis, D.; et al. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature 2009, 461, 1282–1286. [Google Scholar] [CrossRef]
- Cheng, L.; Wu, H.; Cai, X.; Zhang, Y.; Yu, S.; Hou, Y.; Yin, Z.; Yan, Q.; Wang, Q.; Sun, T.; et al. A Gpr35-tuned gut microbe-brain metabolic axis regulates depressive-like behavior. Cell Host Microbe 2024, 32, 227–243.e6. [Google Scholar] [CrossRef]
- Ando, M.; Nagata, K.; Takeshita, R.; Ito, N.; Noguchi, S.; Minamikawa, N.; Kodama, N.; Yamamoto, A.; Yashiro, T.; Hachisu, M.; et al. The gut lactic acid bacteria metabolite, 10-oxo-cis-6,trans-11-octadecadienoic acid, suppresses inflammatory bowel disease in mice by modulating the NRF2 pathway and GPCR-signaling. Front. Immunol. 2024, 15, 1374425. [Google Scholar] [CrossRef]
- Qi, Q.; Zhang, H.; Jin, Z.; Wang, C.; Xia, M.; Chen, B.; Lv, B.; Diaz, L.P.; Li, X.; Feng, R.; et al. Hydrogen sulfide produced by the gut microbiota impairs host metabolism via reducing GLP-1 levels in male mice. Nat. Metab. 2024, 6, 1601–1615. [Google Scholar] [CrossRef] [PubMed]
- Tilyard, M.W.; Spears, G.F.; Thomson, J.; Dovey, S. Treatment of Postmenopausal Osteoporosis with Calcitriol or Calcium. N. Engl. J. Med. 1992, 326, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Black, D.M.; Delmas, P.D.; Eastell, R.; Reid, I.R.; Boonen, S.; Cauley, J.A.; Cosman, F.; Lakatos, P.; Leung, P.C.; Man, Z.; et al. Once-yearly zoledronic acid for treatment of postmenopausal osteoporosis. N. Engl. J. Med. 2007, 356, 1809–1822. [Google Scholar] [CrossRef]
- Marx, R.E.; Sawatari, Y.; Fortin, M.; Broumand, V. Bisphosphonate-induced exposed bone (osteonecrosis/osteopetrosis) of the jaws: Risk factors, recognition, prevention, and treatment. J. Oral. Maxillofac. Surg. 2005, 63, 1567–1575. [Google Scholar] [CrossRef]
- Shane, E.; Burr, D.; Abrahamsen, B.; Adler, R.A.; Brown, T.D.; Cheung, A.M.; Cosman, F.; Curtis, J.R.; Dell, R.; Dempster, D.W.; et al. Atypical subtrochanteric and diaphyseal femoral fractures: Second report of a task force of the American Society for Bone and Mineral Research. J. Bone Miner. Res. 2014, 29, 1–23. [Google Scholar] [CrossRef]
- Cummings, S.R.; Martin, J.S.; McClung, M.R.; Siris, E.S.; Eastell, R.; Reid, I.R.; Delmas, P.; Zoog, H.B.; Austin, M.; Wang, A.; et al. Denosumab for prevention of fractures in postmenopausal women with osteoporosis. N. Engl. J. Med. 2009, 361, 756–765. [Google Scholar] [CrossRef]
- Miller, P.D.; Pannacciulli, N.; Brown, J.P.; Czerwinski, E.; Nedergaard, B.S.; Bolognese, M.A.; Malouf, J.; Bone, H.G.; Reginster, J.-Y.; Singer, A.; et al. Denosumab or Zoledronic Acid in Postmenopausal Women with Osteoporosis Previously Treated with Oral Bisphosphonates. J. Clin. Endocrinol. Metab. 2016, 101, 3163–3170. [Google Scholar] [CrossRef]
- Jiang, W.; Jin, Y.; Zhang, S.; Ding, Y.; Huo, K.; Yang, J.; Zhao, L.; Nian, B.; Zhong, T.P.; Lu, W.; et al. PGE2 activates EP4 in subchondral bone osteoclasts to regulate osteoarthritis. Bone Res. 2022, 10, 27. [Google Scholar] [CrossRef]
- Silverstein, F.E.; Faich, G.; Goldstein, J.L.; Simon, L.S.; Pincus, T.; Whelton, A.; Makuch, R.; Eisen, G.; Agrawal, N.M.; Stenson, W.F.; et al. Gastrointestinal toxicity with celecoxib vs nonsteroidal anti-inflammatory drugs for osteoarthritis and rheumatoid arthritis: The CLASS study: A randomized controlled trial. Celecoxib Long-term Arthritis Safety Study. JAMA 2000, 284, 1247–1255. [Google Scholar] [CrossRef]
- Vrieze, A.; Out, C.; Fuentes, S.; Jonker, L.; Reuling, I.; Kootte, R.S.; van Nood, E.; Holleman, F.; Knaapen, M.; Romijn, J.A.; et al. Impact of oral vancomycin on gut microbiota, bile acid metabolism, and insulin sensitivity. J. Hepatol. 2014, 60, 824–831. [Google Scholar] [CrossRef]
- Gao, X.; Zhang, P. Exercise perspective: Benefits and mechanisms of gut microbiota on the body. Zhong Nan Da Xue Xue Bao Yi Xue Ban 2024, 49, 508–515. [Google Scholar]
- Mohr, A.E.; Jäger, R.; Carpenter, K.C.; Kerksick, C.M.; Purpura, M.; Townsend, J.R.; West, N.P.; Black, K.; Gleeson, M.; Pyne, D.B.; et al. The athletic gut microbiota. J. Int. Soc. Sports Nutr. 2020, 17, 24. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhang, Z.; Hu, B.; Huang, W.; Yuan, C.; Zou, L. Response of Gut Microbiota to Metabolite Changes Induced by Endurance Exercise. Front. Microbiol. 2018, 9, 765. [Google Scholar] [CrossRef]
- Durk, R.P.; Castillo, E.; Márquez-Magaña, L.; Grosicki, G.J.; Bolter, N.D.; Lee, C.M.; Bagley, J.R. Gut Microbiota Composition Is Related to Cardiorespiratory Fitness in Healthy Young Adults. Int. J. Sport. Nutr. Exerc. Metab. 2019, 29, 249–253. [Google Scholar] [CrossRef]
- Morita, E.; Yokoyama, H.; Imai, D.; Takeda, R.; Ota, A.; Kawai, E.; Hisada, T.; Emoto, M.; Suzuki, Y.; Okazaki, K. Aerobic Exercise Training with Brisk Walking Increases Intestinal Bacteroides in Healthy Elderly Women. Nutrients 2019, 11, 868. [Google Scholar] [CrossRef]
- Yang, Y.; Shi, Y.; Wiklund, P.; Tan, X.; Wu, N.; Zhang, X.; Tikkanen, O.; Zhang, C.; Munukka, E.; Cheng, S. The Association between Cardiorespiratory Fitness and Gut Microbiota Composition in Premenopausal Women. Nutrients 2017, 9, 792. [Google Scholar] [CrossRef]
- Slavin, J. Fiber and prebiotics: Mechanisms and health benefits. Nutrients 2013, 5, 1417–1435. [Google Scholar] [CrossRef]
- Tu, Q.; He, Z.; Li, Y.; Chen, Y.; Deng, Y.; Lin, L.; Hemme, C.L.; Yuan, T.; Van Nostrand, J.D.; Wu, L.; et al. Development of HuMiChip for functional profiling of human microbiomes. PLoS ONE 2014, 9, e90546. [Google Scholar] [CrossRef]
- Statovci, D.; Aguilera, M.; MacSharry, J.; Melgar, S. The Impact of Western Diet and Nutrients on the Microbiota and Immune Response at Mucosal Interfaces. Front. Immunol. 2017, 8, 838. [Google Scholar] [CrossRef]
- Olson, C.A.; Vuong, H.E.; Yano, J.M.; Liang, Q.Y.; Nusbaum, D.J.; Hsiao, E.Y. The Gut Microbiota Mediates the Anti-Seizure Effects of the Ketogenic Diet. Cell 2018, 174, 497. [Google Scholar] [CrossRef]
- Rondanelli, M.; Gasparri, C.; Peroni, G.; Faliva, M.A.; Naso, M.; Perna, S.; Bazire, P.; Sajuox, I.; Maugeri, R.; Rigon, C. The Potential Roles of Very Low Calorie, Very Low Calorie Ketogenic Diets and Very Low Carbohydrate Diets on the Gut Microbiota Composition. Front. Endocrinol. 2021, 12, 662591. [Google Scholar]
- Singh, R.K.; Chang, H.W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H.; et al. Influence of diet on the gut microbiome and implications for human health. J. Transl. Med. 2017, 15, 73. [Google Scholar] [CrossRef] [PubMed]
- Tomova, A.; Bukovsky, I.; Rembert, E.; Yonas, W.; Alwarith, J.; Barnard, N.D.; Kahleova, H. The Effects of Vegetarian and Vegan Diets on Gut Microbiota. Front. Nutr. 2019, 6, 47. [Google Scholar] [CrossRef]
- De Palma, G.; Nadal, I.; Collado, M.C.; Sanz, Y. Effects of a gluten-free diet on gut microbiota and immune function in healthy adult human subjects. Br. J. Nutr. 2009, 102, 1154–1160. [Google Scholar] [CrossRef]
- Bonder, M.J.; Tigchelaar, E.F.; Cai, X.; Trynka, G.; Cenit, M.C.; Hrdlickova, B.; Zhong, H.; Vatanen, T.; Gevers, D.; Wijmenga, C.; et al. The influence of a short-term gluten-free diet on the human gut microbiome. Genome Med. 2016, 8, 45. [Google Scholar] [CrossRef]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial ecology: Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef]
- Haro, C.; Montes-Borrego, M.; Rangel-Zuñiga, O.A.; Alcala-Diaz, J.F.; Gómez-Delgado, F.; Pérez-Martínez, P.; Delgado-Lista, J.; Quintana-Navarro, G.M.; Tinahones, F.J.; Landa, B.B.; et al. Two Healthy Diets Modulate Gut Microbial Community Improving Insulin Sensitivity in a Human Obese Population. J. Clin. Endocrinol. Metab. 2016, 101, 233–242. [Google Scholar] [CrossRef]
- Wen, Z.Q.; Liu, D.; Zhang, Y.; Cai, Z.-J.; Xiao, W.-F.; Li, Y.-S. G Protein-Coupled Receptors in Osteoarthritis: A Novel Perspective on Pathogenesis and Treatment. Front. Cell Dev. Biol. 2021, 9, 758220. [Google Scholar] [CrossRef]
- Uniyal, A.; Tiwari, V.; Tsukamoto, T.; Dong, X.; Guan, Y.; Raja, S.N. Targeting sensory neuron GPCRs for peripheral neuropathic pain. Trends Pharmacol. Sci. 2023, 44, 1009–1027. [Google Scholar] [CrossRef]
- Zhang, M.; Chen, T.; Lu, X.; Lan, X.; Chen, Z.; Lu, S. G protein-coupled receptors (GPCRs): Advances in structures, mechanisms, and drug discovery. Signal Transduct. Target. Ther. 2024, 9, 88. [Google Scholar] [CrossRef]
- Vrieze, A.; Van Nood, E.; Holleman, F.; Salojärvi, J.; Kootte, R.S.; Bartelsman, J.F.; Dallinga-Thie, G.M.; Ackermans, M.T.; Serlie, M.J.; Oozeer, R.; et al. Transfer of intestinal microbiota from lean donors increases insulin sensitivity in individuals with metabolic syndrome. Gastroenterology 2012, 143, 913–916.e7. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Chen, Y.; Huang, W.; Zhou, H.; Zhang, W. Drug-microbiota interactions: An emerging priority for precision medicine. Signal Transduct. Target. Ther. 2023, 8, 386. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Subject | Disease Model | Methods | Sample Size (n) | Changes in Gut Microbiota (Beneficial Bacteria) | Changes in Gut Microbiota (Pathogenic Bacteria) | Reference |
|---|---|---|---|---|---|---|---|
| RA | human | Rheumatoid arthritis patients | 16S rRNA sequencing | 52 | Fusicatenibacter ↓ Enterococcium ↓ | Escherichia ↑ Eisenbergiella ↑ Klebsiella ↑ | [59] |
| RA | animal | Collagen-induced arthritis (CIA) rats | 16S rRNA sequencing | around 10–20 per group | Bifidobacterium ↓ Lactobacillus ↓ Pseudomonas fragilis ↓ | Fusobacterium nucleatum ↑ Streptococcus ↑ Escherichia coli ↑ Prevotella ↑ Bacteroidetes ↑ | [60,61,62,63,64] |
| OA | human | Osteoarthritis patients | Metagenomic sequencing | 89 | Agathobacter ↓ Ruminococcus ↓ Roseburia ↓ Subdoligranulum ↓ Lactobacillus ↓ | Prevotella_7 ↑ Clostridium ↑ Flavonifractor ↑ Klebsiella ↑ | [63,65] |
| OA | animal | Mono-iodine-acetic acid (MIA) induced OA animal models | Metagenomic sequencing or 16S rRNA sequencing | around 10–20 per group or 93 | Dictyostelium parvum ↓ Ackermannia ↓ Lactobacillus ↓ Bifidobacterium bifidum ↓ | Enterobacteriaceae ↑ Staphylococcus Proteus ↑ Clostridium ↑ | [66,67] |
| OP | human | Osteoporosis patients | 16S rRNA sequencing | 2033 | Firmicutes ↓ Blautia ↓ Alistipes ↓ Megamonas ↓ Anaerostipes ↓ | Lactobacillus ↑ Ruminococcus ↑ Bacteroides of Bacteroidetes ↑ | [46] |
| OP | animal | Ovariectomized (OVX) rats | 16S rRNA sequencing | around 10–20 per group | Bifidobacterium ↓ Lactobacillus ↓ Akkermansia muciniphila ↓ | Streptococcus sanguinis ↑ Streptococcus gordonii ↑ Actinomyces odontolyticus ↑ Actinomyces graevenitzii ↑ Escherichia coli ↑ Enterococcus ↑ Bacteroides ↑ | [68,69] |
| BM | human | Tumour bone metastasis patients | Metagenomic DNA sequencing | 79 | Megamonas ↓ Clostridia ↓ Akkermansia ↓ Gemmiger ↓ Paraprevotella ↓ | Lactobacillales ↑ Bacilli ↑ Veillonella ↑ Streptococcus ↑ Campylobacter ↑ Epsilonproteobacteria ↑ Acinetobacter ↑ Pseudomonadales ↑ Moraxellaceae ↑ | [70] |
| BM | animal | Inoculation of tumour cells into animal bones | Metagenomic DNA sequencing or 16S rRNA sequencing | around 10–20 per group | Bifidobacterium ↓ Lactobacillus ↓ Akkermansia muciniphila ↓ | Fusobacterium nucleatum ↑ Escherichia coli ↑ Enterococcus faecalis ↑ Fusobacterium ↑ | [49,71] |
| Drug Development | Name of Drug | Mechanism of Action | Side Effects | Reference |
|---|---|---|---|---|
| Existing Drugs | Bisphosphonates | Used in the treatment of osteoporosis to increase bone density by activating the TGR5 | Studies have found that phosphonates are associated with an increased risk of atypical femur fractures, bisphosphonates have been associated with an increased risk of osteonecrosis of the jaw (ONJ), particularly in patients undergoing dental surgery, and oral bisphosphonates may cause oesophagitis and other gastro symptoms. | [162,163,164,165] |
| Denosumab | A monoclonal antibody against RANKL used for the treatment of osteoporosis and bone metastases that reduces bone resorption by inhibiting the RANKL-RANK signalling pathway | Studies have found that Denosumab may affect the immune system and increase the risk of infection. Denosumab treatment may cause hypocalcaemia, especially in patients with vitamin D deficiency. | [166,167] | |
| Prostaglandin | Used in the treatment of osteoarthritis to reduce inflammation and pain by activating EP2 and EP4 receptors | PGE2 analogues can cause gastro disturbances, increase the risk of cardiovascular events (especially in heart disease patients), and negatively impact renal function (especially in the elderly or those with kidney disease history). | [168,169] | |
| New Drug Candidate Molecules | GPR41 and GPR43 Agonists | Activators of short-chain fatty acid receptors being investigated to promote osteoblast differentiation and inhibit osteoclast activity for the treatment of osteoporosis | [14] | |
| TGR5 Agonists | Activators of the bile acid receptor in development to improve energy metabolism and bone density | [87] | ||
| AhR Agonists | Activators of the aromatic hydrocarbon receptor, potentially used in the treatment of osteoarthritis by modulating immune response and inflammation | [170] |
| Type | Changes in Gut Microbiota | Reference |
|---|---|---|
| High-intensity interval training | In bodybuilders, the genera Clostridium, Eisenbergiella, Faecalibacterium, Haemophilus, and Sutterella were more abundant, while Bifidobacterium and Parasutterella were less abundant. Probiotic species including Bifidobacterium adolescentis, Bifidobacterium longum, Latilactobacillus sakei, and SCFA-producing microorganisms (Blautia wexlerae, Eubacterium hallii) were less abundant in bodybuilders compared to controls. | [172] |
| Endurance training | At the phylum level, Lentisphaerae and Acidobacteria were detected after running, with their functions in the human gut remaining unknown. At the family level, there was an increase in Coriobacteriaceae and Succinivibrionaceae, where Coriobacteriaceae are involved in bile salts and steroid hormones metabolism and activation of dietary polyphenols. At the genus level, half marathon running led to a decrease in the levels of Ezakiella, Romboutsia, and Actinobacillus, and an increase in Coprococcus and Ruminococcus bicirculans. | [173] |
| Aerobic exercise | In a study by Durk et al. [174] on young healthy individuals, a higher Firmicutes/Bacteroidetes ratio was significantly correlated with higher VO2max. In a study on premenopausal women, participants with low VO2max had lower Bacteroides levels but higher Eubacterium rectale and Clostridium coccoides levels compared to those with high VO2max. A 12-week aerobic exercise training led to an increase in the relative abundance of intestinal Bacteroides and an improvement in cardiorespiratory fitness. | [174,175,176] |
| Type | Changes in Gut Microbiota | Reference |
|---|---|---|
| Western Diet | This type of diet leads to a decrease in the abundance of several beneficial bacterial species, including Akkermansia muciniphila, Faecalibacterium prausnitzii, Roseburia spp., Eubacterium spp., and the bacteria in Clostridium cluster XIVa and IV. | [178,179] |
| Ketogenic Diet | In obese patients, the Bacteroidetes/Firmicutes ratio was significantly altered. After a very-low-calorie ketogenic diet (VLCKD), there was a reduction in Eubacterium rectale and Roseburia, while the abundance of Akkermansia and Christensenellaceae increased. | [180,181] |
| Vegan Diet | High intakes of indigestible carbohydrates (wheat bran and whole grain) typically lead to an increase in Lactobacillus spp. and Bifidobacterium spp. Whole grain barley and resistant starch are associated with an increase in Ruminococcus spp., Eubacterium rectale, and Roseburia spp., and a decrease in some Firmicutes phylum taxa such as Clostridium and Enterococcus species. | [182,183] |
| Gluten-Free Diet | In healthy volunteers, one month of a gluten-free diet (GFD) led to the following microbiota changes: a decrease in the populations of Lactobacillus, Bifidobacterium, and Roseburia; an increase in E. coli, Enterobacteriaceae, Victivallaceae, and Clostridiaceae. | [184,185] |
| Mediterranean Diet | In a study of twenty obese men, a one-year Mediterranean diet (MD) led to a decrease in Prevotella and an increase in Oscillospira and Roseburia. A long-term MD resulted in an increase in the relative abundance of Parabacteroides distasonis. A two-year MD had a positive effect, increasing the levels of Bacteroides, Prevotella, and some saccharolytic genera such as Roseburia, Faecalibacterium, and Ruminococcus. | [186,187] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, P.; Liu, J.; Chen, G.; Guo, Y. The Role of G Protein-Coupled Receptors in the Regulation of Orthopaedic Diseases by Gut Microbiota. Nutrients 2025, 17, 1702. https://doi.org/10.3390/nu17101702
Sun P, Liu J, Chen G, Guo Y. The Role of G Protein-Coupled Receptors in the Regulation of Orthopaedic Diseases by Gut Microbiota. Nutrients. 2025; 17(10):1702. https://doi.org/10.3390/nu17101702
Chicago/Turabian StyleSun, Peng, Jinchao Liu, Guannan Chen, and Yilan Guo. 2025. "The Role of G Protein-Coupled Receptors in the Regulation of Orthopaedic Diseases by Gut Microbiota" Nutrients 17, no. 10: 1702. https://doi.org/10.3390/nu17101702
APA StyleSun, P., Liu, J., Chen, G., & Guo, Y. (2025). The Role of G Protein-Coupled Receptors in the Regulation of Orthopaedic Diseases by Gut Microbiota. Nutrients, 17(10), 1702. https://doi.org/10.3390/nu17101702