
Targeting the Hallmarks of Aging with Vitamin D: Starting to Decode the Myth

 , ,
, ,  , , ,
, , ,  ,
,
Abstract
1. Introduction
2. Methods
3. The Hallmarks of Aging
3.1. Genomic Instability
3.2. Telomere Attrition
3.3. Epigenetic Alterations
3.4. Loss of Proteostasis
3.5. Disabled Macroautophagy
3.6. Deregulated Nutrient-Sensing
3.7. Mitochondrial Dysfunction
3.8. Cellular Senescence
3.9. Stem Cell Exhaustion
3.10. Altered Intercellular Communication
3.11. Chronic Inflammation
3.12. Dysbiosis
4. Vitamin D and Its Potential Impact on the Biological Hallmarks of Aging
4.1. Vitamin D and Genomic Instability
4.2. Vitamin D and Telomere Attrition
4.3. Vitamin D and Epigenetic Alterations
4.4. Vitamin D and Lack of Proteostasis
4.5. Vitamin D and Mitochondrial Dysfunction
4.6. Vitamin D and Cellular Senescence
4.7. Vitamin D and Stem Cell Exhaustion
4.8. Vitamin D and Altered Intercellular Communication
4.9. Vitamin D and Chronic Inflammation
4.10. Vitamin D and Dysbiosis

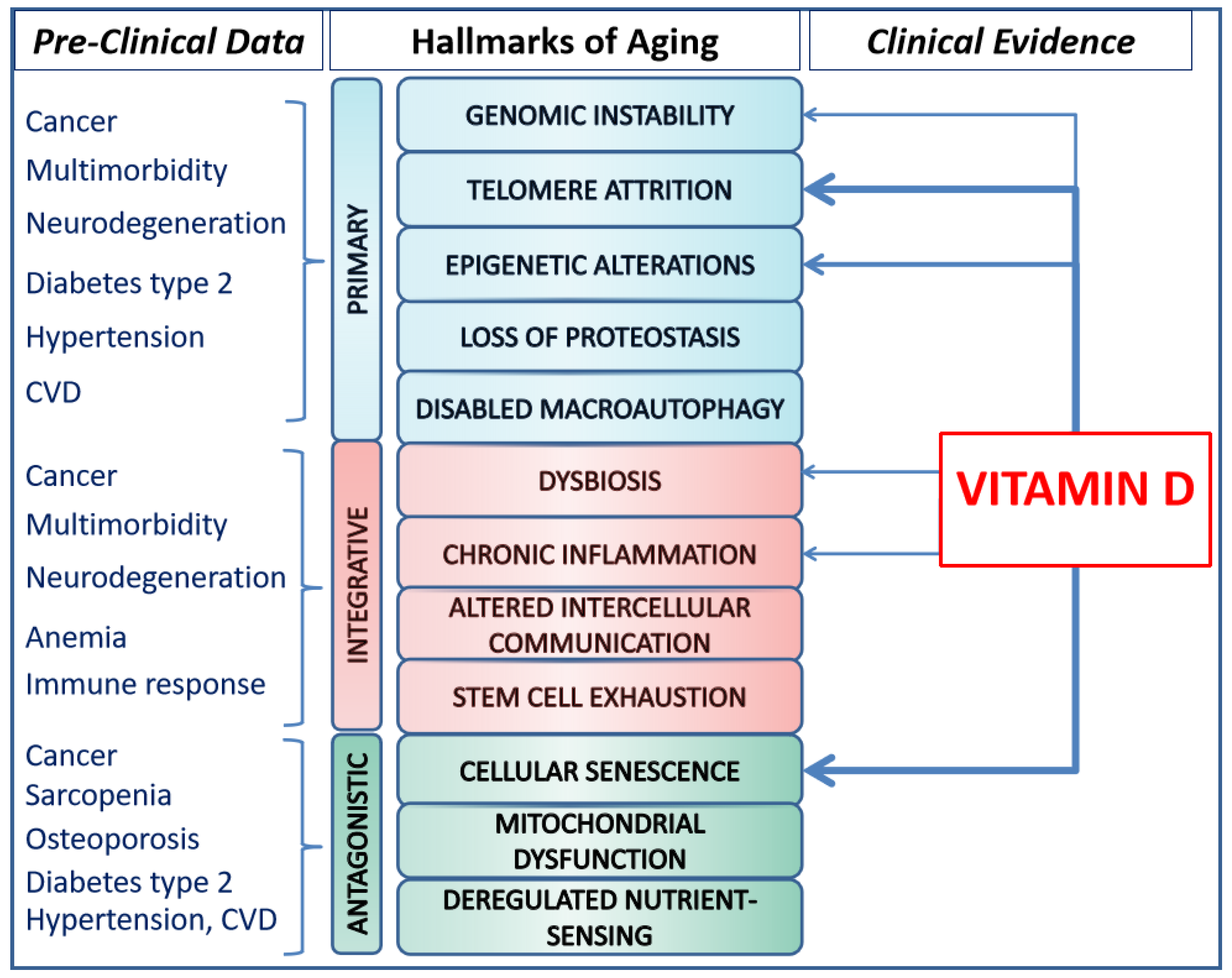{kind=link}
| Hallmark | Study | Cohort | Aim | Conclusion | Refs. |
|---|---|---|---|---|---|
| Genomic instability | Placebo-controlled | 92 patients with colorectal adenoma | Evaluate the effect of VitD supplement with 800 UI/day for 6 months | Significant increase in Bax expression (apoptosis promoter); no increase of Bcl-2 expression (apoptosis inhibitor) in VitD group | [61] |
| Randomized Clinical Trial | 92 subjects with VitD insufficiency, with and without T2DM | Evaluate the effect of VitD supplement with 2000 UI/day for 3 months | Decreased percentage of DNA damage and oxidative parameters when compared to the control group | [62] | |
| Telomere Attrition | Twins UK Cohort ObservationalRegistry | 2160 women (aged 18–79) from population-based cohort of twins | Evaluate the relationship between VitD concentrations and the rate of telomere attrition in leukocytes | Positive correlation between serum 25(OH)D levels and telomere length | [64] |
| Nurses’ Health Study Observational Registry | 1424 women | Evaluate the association between both 25(OH)D and 1,25(OH)2D and leukocyte telomere length | Positive correlation between serum 25(OH)D concentrations and leukocyte telomere length | [65] | |
| Cross sectional analysis from the Health Professional Follow-Up study | 2843 men (from studies of telomeres and cancer) | Evaluate the association between 25(OH)D, 1,25-dihydroxyvitamin D (1,25(OH)2D) and leukocyte telomere length | No association between VitD levels and leukocyte telomere length | [66] | |
| Cross sectional analysis from Northern Finland Birth Cohort 1966 | 5096 younger adults | Evaluate associations between 25(OH)D and body mass index (BMI) with leukocyte telomere length and whether associations are independent of concentrations of C-reactive protein | No association between VitD levels and leukocyte telomere length | [67] | |
| Cohort from the Newcastle 85+ Study | 775 older adults (>85 ys), community dwelling, and institutionalized older patients | Evaluate the association between serum VitD concentration and telomere length in blood cells at baseline, 18 and 36 months | Significant positive association between VitD and telomere length at baseline, but inconsistent relationships at subsequent time points | [68] | |
| Case–control study | African American Gullah women with systemic lupus erythematosus (n = 59) and healthy controls (n = 59) | Evaluate the relationships between VitD status, telomere length, and anti-telomere antibodies | Significant association between shorter telomeres and lower 25(OH)D levels in patients and healthy control; shorter telomeres at follow-up in VitD-deficient patients | [69] | |
| Retrospective case–control study | 62 stable hemodialysis patients and 60 controls | Evaluate the potential protective role of VitD supplementation on telomere length in peripheral mononuclear cells | Hemodialysis patients treated with VitD had greater telomere length in peripheral mononuclear cells | [70] | |
| Double-blind, placebo-controlled clinical trial | 37 overweight African American subjects (18 placebo and 19 intervention) | Evaluate the effect of VitD3 oral supplementation (60,000 IU/month) | VitD supplementation significantly increased telomerase activity | [74] | |
| Epigenetic alteration | Sub analysis from RCT | 51overweight/obese African Americans VitD-deficient subjects | Evaluate different effects of VitD3 supplement (600 IU/day, 2000 IU/day, and 4000 IU/day) for 16 weeks compared to placebo | DNAm age was correlated with chronological age in patients with low VitD levels, while the supplementation was able to decrease DNA metilation and slowed epigenetic aging | [75] |
| Southampton Women’s Survey: subgroup analysis from prospective cohort | 230 children from the Southampton Women’s Survey (SWS) mother–offspring study | Evaluate the correlation between VitD status and Retinoid-X receptor-alpha (RXRA) methylation in umbilical cord DNA | Methylation at RXRA locus in the umbilical cord was associated with the offspring bone mass; specific methylation at one locus was associated with maternal VitD deficiency | [78] | |
| The Maternal VitD Osteoporosis Study (MAVIDOS RCT): subgroup analysis | 479 received VitD3 supplementation and 486 were in the placebo group | Evaluate DNAm at a specific site in the RXRA locus (10 CpG) | In the cohort of mothers’ receiving VitD, the methylation levels in the RXRA-specific site were significantly lower compared to the placebo group | [79] | |
| Randomized controlled pilot study | Pregnant women receiving VitD3 400 IU (n = 6, control group) or 3800 IU (n = 7, intervention group) | Identify differentially methylated loci among infants born to mothers supplemented with VitD 400 IU/day (i.e., control) versus 3800 IU/day (i.e., intervention) | Compared to control, intervention was associated with mean 25(OH)D levels reflective of sufficient maternal VitD status by birth, as well as significant gains in maternal leukocyte DNA methylation associated with genes involved in cell migration/motility, development, and growth | [80] | |
| Cellular senescence | Case–control study | 120 individuals (30 young, 30 older, and 30 older in treatment group 1 (1 sachet of Difensil® IMMUNO/day for 12 weeks) and 20 older in treatment group 2 (2 sachet of Difensil® IMMUNO/day for 6 weeks) | Evaluate the effect of the nutraceutical supplement (comprehensive of Sambucus nigra, zinc, tyndallized Lactobacillus acidophilus (HA122), arabinogalactans, VitD, vitamin E, vitamin C, and group B vitamins) on specific signatures of immunosenescence | Older people treated for 30 days improved IL-6, CRP, and lymphocytes levels independent from the dosage of the supplements used; despite the improvement, they were not able to reach the same conditions of young patients | [95] |
| Double Blind RCT | 110 older fit or frail subjects | Evaluate the effect of alphacalcidol supplementation in inflammatory profile (Il-6, IL-10, TNF, CD4/CD8 ratio, CD8+, and CD28-) | Cohort receiving alphacalcidiol had a better immune profile across the whole cohort as well as the cohort divided according to frailty status | [96] | |
| Chronic inflammation | VITAL study sub-cohort | 25,871 women aged ≥55 and men aged ≥50 | Measure the effect of VitD supplementation (2000 UI/day) and/or n-3 FAs on systemic inflammatory biomarkers | VitD supplementation with or without n-3 FAs decreased hs-CRP by 19% at year 2, and the reduction was attenuated at 4 years Other inflammatory biomarkers (IL-6, IL-10, and TNF-α) were not significantly modified | [114] |
| Dysbiosis | Sub analysis from a RCT | 167 HIV-1 patients receiving VitD 5000 IU/day and 500 mg phenylbutyrate (PBA) for 16 weeks | VitD or PBA may have a modulatory action on microbiota composition, metabolites production, and immune activation | No significant improvement in circulating antimicrobial peptide LL-37 and an actual impact on microbiota | [115] |
| Pilot study | 24 patients with knee osteoarthritis (KOA), healthy controls, with and without VitD deficiency | Evaluate the interplay between microbiome, knee OA (KOA), and VitD levels for establishing whether VitD deficiency is associated with microbiome dysfunction | Patients with KOA and VitD-deficient showed specific bacteria species, absent in KOA, and not VitD-deficient; The different core bacteria in KOA and VitD-deficient suggested an interplay between conditions | [116] | |
| RCT | 41 patients with cystic fibrosis divided in VitD insufficient (n = 23) and VitD sufficient (n = 18) | Comparing microbiota composition in VitD insufficient and sufficient patients and evaluate the impact of VitD supplement. | Potentially pathogenic species were identified in the microbiome of VitD-deficient subjects | [117] | |
| Interventional study | 100 healthy women | Evaluate gut microbiota composition before and after VitD supplementation | Significant increment of health-promoting gut probiotics after VitD supplementation | [118] |
5. Discussion and Future Perspectives
Funding
Conflicts of Interest
References
- Lopez-Otin, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef]
- Lopez-Otin, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. Hallmarks of aging: An expanding universe. Cell 2023, 186, 243–278. [Google Scholar] [CrossRef]
- Tosato, M.; Zamboni, V.; Ferrini, A.; Cesari, M. The aging process and potential interventions to extend life expectancy. Clin. Interv. Aging 2007, 2, 401–412. [Google Scholar] [PubMed]
- Polidori, M.C. Aging hallmarks, biomarkers, and clocks for personalized medicine: (Re)positioning the limelight. Free Radic. Biol. Med. 2024, 215, 48–55. [Google Scholar] [CrossRef]
- Kennedy, B.K.; Berger, S.L.; Brunet, A.; Campisi, J.; Cuervo, A.M.; Epel, E.S.; Franceschi, C.; Lithgow, G.J.; Morimoto, R.I.; Pessin, J.E.; et al. Geroscience: Linking aging to chronic disease. Cell 2014, 159, 709–713. [Google Scholar] [CrossRef]
- Bouillon, R.; Marcocci, C.; Carmeliet, G.; Bikle, D.; White, J.H.; Dawson-Hughes, B.; Lips, P.; Munns, C.F.; Lazaretti-Castro, M.; Giustina, A.; et al. Skeletal and Extraskeletal Actions of Vitamin D: Current Evidence and Outstanding Questions. Endocr. Rev. 2019, 40, 1109–1151. [Google Scholar] [CrossRef] [PubMed]
- Luiz, M.M.; Maximo, R.; Oliveira, D.C.; Ramirez, P.C.; de Souza, A.F.; Delinocente, M.L.B.; Steptoe, A.; de Oliveira, C.; Alexandre, T. Association of Serum 25-Hydroxyvitamin D Deficiency with Risk of Incidence of Disability in Basic Activities of Daily Living in Adults >50 Years of Age. J. Nutr. 2020, 150, 2977–2984. [Google Scholar] [CrossRef] [PubMed]
- Luiz, M.M.; Maximo, R.O.; de Oliveira, D.C.; Ramirez, P.C.; de Souza, A.F.; Delinocente, M.L.B.; Steptoe, A.; de Oliveira, C.; Alexandre, T.D.S. Sex Differences in Vitamin D Status as a Risk Factor for Incidence of Disability in Instrumental Activities of Daily Living: Evidence from the ELSA Cohort Study. Nutrients 2022, 14, 2012. [Google Scholar] [CrossRef]
- Zamboni, M.; Zoico, E.; Tosoni, P.; Zivelonghi, A.; Bortolani, A.; Maggi, S.; Di Francesco, V.; Bosello, O. Relation between vitamin D, physical performance, and disability in elderly persons. J. Gerontol. A Biol. Sci. Med. Sci. 2002, 57, M7–M11. [Google Scholar] [CrossRef]
- Girgis, C.M.; Mokbel, N.; Digirolamo, D.J. Therapies for musculoskeletal disease: Can we treat two birds with one stone? Curr. Osteoporos. Rep. 2014, 12, 142–153. [Google Scholar] [CrossRef]
- National Institute of Health and Care Excellence. Vitamin D: Supplement Use in Specific Population Groups. Available online: https://www.nice.org.uk/guidance/ph56 (accessed on 30 December 2023).
- Gallagher, J.C. Vitamin D and aging. Endocrinol. Metab. Clin. N. Am. 2013, 42, 319–332. [Google Scholar] [CrossRef]
- Vetter, V.M.; Sommerer, Y.; Kalies, C.H.; Spira, D.; Bertram, L.; Demuth, I. Vitamin D supplementation is associated with slower epigenetic aging. Geroscience 2022, 44, 1847–1859. [Google Scholar] [CrossRef] [PubMed]
- Pal, E.; Ungvari, Z.; Benyo, Z.; Varbiro, S. Role of Vitamin D Deficiency in the Pathogenesis of Cardiovascular and Cerebrovascular Diseases. Nutrients 2023, 15, 334. [Google Scholar] [CrossRef]
- Tenchov, R.; Sasso, J.M.; Wang, X.; Zhou, Q.A. Aging Hallmarks and Progression and Age-Related Diseases: A Landscape View of Research Advancement. ACS Chem. Neurosci. 2024, 15, 1–30. [Google Scholar] [CrossRef]
- Zampino, M.; AlGhatrif, M.; Kuo, P.L.; Simonsick, E.M.; Ferrucci, L. Longitudinal Changes in Resting Metabolic Rates with Aging Are Accelerated by Diseases. Nutrients 2020, 12, 3061. [Google Scholar] [CrossRef]
- Blokzijl, F.; de Ligt, J.; Jager, M.; Sasselli, V.; Roerink, S.; Sasaki, N.; Huch, M.; Boymans, S.; Kuijk, E.; Prins, P.; et al. Tissue-specific mutation accumulation in human adult stem cells during life. Nature 2016, 538, 260–264. [Google Scholar] [CrossRef]
- Usman, M.; Woloshynowych, M.; Britto, J.C.; Bilkevic, I.; Glassar, B.; Chapman, S.; Ford-Adams, M.E.; Desai, A.; Bain, M.; Tewfik, I.; et al. Obesity, oxidative DNA damage and vitamin D as predictors of genomic instability in children and adolescents. Int. J. Obes. 2021, 45, 2095–2107. [Google Scholar] [CrossRef] [PubMed]
- Giardini, M.A.; Segatto, M.; da Silva, M.S.; Nunes, V.S.; Cano, M.I. Telomere and telomerase biology. Prog. Mol. Biol. Transl. Sci. 2014, 125, 1–40. [Google Scholar] [CrossRef] [PubMed]
- Raynaud, C.M.; Sabatier, L.; Philipot, O.; Olaussen, K.A.; Soria, J.C. Telomere length, telomeric proteins and genomic instability during the multistep carcinogenic process. Crit. Rev. Oncol. Hematol. 2008, 66, 99–117. [Google Scholar] [CrossRef]
- Gruber, H.J.; Semeraro, M.D.; Renner, W.; Herrmann, M. Telomeres and Age-Related Diseases. Biomedicines 2021, 9, 1335. [Google Scholar] [CrossRef] [PubMed]
- Zarei, M.; Zarezadeh, M.; Hamedi Kalajahi, F.; Javanbakht, M.H. The Relationship Between Vitamin D and Telomere/Telomerase: A Comprehensive Review. J. Frailty Aging 2021, 10, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Munoz-Najar, U.; Sedivy, J.M. Epigenetic control of aging. Antioxid. Redox Signal 2011, 14, 241–259. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Liu, H.; Hu, Q.; Wang, L.; Liu, J.; Zheng, Z.; Zhang, W.; Ren, J.; Zhu, F.; Liu, G.H. Epigenetic regulation of aging: Implications for interventions of aging and diseases. Signal Transduct. Target. Ther. 2022, 7, 374. [Google Scholar] [CrossRef] [PubMed]
- Vetter, V.M.; Spira, D.; Banszerus, V.L.; Demuth, I. Epigenetic Clock and Leukocyte Telomere Length Are Associated with Vitamin D Status but not with Functional Assessments and Frailty in the Berlin Aging Study II. J. Gerontol. A Biol. Sci. Med. Sci. 2020, 75, 2056–2063. [Google Scholar] [CrossRef] [PubMed]
- Mark, K.A.; Dumas, K.J.; Bhaumik, D.; Schilling, B.; Davis, S.; Oron, T.R.; Sorensen, D.J.; Lucanic, M.; Brem, R.B.; Melov, S.; et al. Vitamin D Promotes Protein Homeostasis and Longevity via the Stress Response Pathway Genes skn-1, ire-1, and xbp-1. Cell Rep. 2016, 17, 1227–1237. [Google Scholar] [CrossRef]
- Bhutia, S.K. Vitamin D in autophagy signaling for health and diseases: Insights on potential mechanisms and future perspectives. J. Nutr. Biochem. 2022, 99, 108841. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.B.; Aleksic, S.; Gao, T.; Weiss, E.F.; Demetriou, E.; Verghese, J.; Holtzer, R.; Barzilai, N.; Milman, S. Insulin-like Growth Factor-1 and IGF Binding Proteins Predict All-Cause Mortality and Morbidity in Older Adults. Cells 2020, 9, 1368. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Hamidu, S.; Yang, X.; Yan, Y.; Wang, Q.; Li, L.; Oduro, P.K.; Li, Y. Dietary Supplements and Natural Products: An Update on Their Clinical Effectiveness and Molecular Mechanisms of Action During Accelerated Biological Aging. Front. Genet. 2022, 13, 880421. [Google Scholar] [CrossRef] [PubMed]
- Healthy Ageing and Functional Ability. Available online: https://www.who.int/news-room/questions-and-answers/item/healthy-ageing-and-functional-ability (accessed on 30 December 2023).
- Gerencser, A.A.; Doczi, J.; Torocsik, B.; Bossy-Wetzel, E.; Adam-Vizi, V. Mitochondrial swelling measurement in situ by optimized spatial filtering: Astrocyte-neuron differences. Biophys. J. 2008, 95, 2583–2598. [Google Scholar] [CrossRef] [PubMed]
- Seo, A.Y.; Joseph, A.M.; Dutta, D.; Hwang, J.C.; Aris, J.P.; Leeuwenburgh, C. New insights into the role of mitochondria in aging: Mitochondrial dynamics and more. J. Cell Sci. 2010, 123, 2533–2542. [Google Scholar] [CrossRef]
- Srivastava, S. The Mitochondrial Basis of Aging and Age-Related Disorders. Genes 2017, 8, 398. [Google Scholar] [CrossRef] [PubMed]
- Chee, C.; Shannon, C.E.; Burns, A.; Selby, A.L.; Wilkinson, D.; Smith, K.; Greenhaff, P.L.; Stephens, F.B. Increasing skeletal muscle carnitine content in older individuals increases whole-body fat oxidation during moderate-intensity exercise. Aging Cell 2021, 20, e13303. [Google Scholar] [CrossRef] [PubMed]
- Coradduzza, D.; Congiargiu, A.; Chen, Z.; Cruciani, S.; Zinellu, A.; Carru, C.; Medici, S. Humanin and Its Pathophysiological Roles in Aging: A Systematic Review. Biology 2023, 12, 558. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Son, J.M.; Benayoun, B.A.; Lee, C. The Mitochondrial-Encoded Peptide MOTS-c Translocates to the Nucleus to Regulate Nuclear Gene Expression in Response to Metabolic Stress. Cell Metab. 2018, 28, 516–524.e517. [Google Scholar] [CrossRef] [PubMed]
- Matta Reddy, A.; Iqbal, M.; Chopra, H.; Urmi, S.; Junapudi, S.; Bibi, S.; Kumar Gupta, S.; Nirmala Pangi, V.; Singh, I.; Abdel-Daim, M.M. Pivotal role of vitamin D in mitochondrial health, cardiac function, and human reproduction. EXCLI J. 2022, 21, 967–990. [Google Scholar] [CrossRef]
- Hernandez-Segura, A.; Nehme, J.; Demaria, M. Hallmarks of Cellular Senescence. Trends Cell Biol. 2018, 28, 436–453. [Google Scholar] [CrossRef] [PubMed]
- Diwan, B.; Sharma, R. Nutritional components as mitigators of cellular senescence in organismal aging: A comprehensive review. Food Sci. Biotechnol. 2022, 31, 1089–1109. [Google Scholar] [CrossRef]
- Chaib, S.; Tchkonia, T.; Kirkland, J.L. Cellular senescence and senolytics: The path to the clinic. Nat. Med. 2022, 28, 1556–1568. [Google Scholar] [CrossRef] [PubMed]
- Teale, M.A.; Schneider, S.; Eibl, D.; van den Bos, C.; Neubauer, P.; Eibl, R. Mesenchymal and induced pluripotent stem cell-based therapeutics: A comparison. Appl. Microbiol. Biotechnol. 2023, 107, 4429–4445. [Google Scholar] [CrossRef]
- Brunet, A.; Goodell, M.A.; Rando, T.A. Ageing and rejuvenation of tissue stem cells and their niches. Nat. Rev. Mol. Cell Biol. 2023, 24, 45–62. [Google Scholar] [CrossRef]
- Garay, R.P. Recent clinical trials with stem cells to slow or reverse normal aging processes. Front. Aging 2023, 4, 1148926. [Google Scholar] [CrossRef]
- Fantini, C.; Corinaldesi, C.; Lenzi, A.; Migliaccio, S.; Crescioli, C. Vitamin D as a Shield against Aging. Int. J. Mol. Sci. 2023, 24, 4546. [Google Scholar] [CrossRef]
- Baechle, J.J.; Chen, N.; Makhijani, P.; Winer, S.; Furman, D.; Winer, D.A. Chronic inflammation and the hallmarks of aging. Mol. Metab. 2023, 74, 101755. [Google Scholar] [CrossRef]
- Korf, H.; Wenes, M.; Stijlemans, B.; Takiishi, T.; Robert, S.; Miani, M.; Eizirik, D.L.; Gysemans, C.; Mathieu, C. 1,25-Dihydroxyvitamin D3 curtails the inflammatory and T cell stimulatory capacity of macrophages through an IL-10-dependent mechanism. Immunobiology 2012, 217, 1292–1300. [Google Scholar] [CrossRef]
- Ghosh, T.S.; Shanahan, F.; O’Toole, P.W. The gut microbiome as a modulator of healthy ageing. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 565–584. [Google Scholar] [CrossRef]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M.; et al. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: A proof-of-concept exploratory study. Nat. Med. 2019, 25, 1096–1103. [Google Scholar] [CrossRef] [PubMed]
- Ratiner, K.; Ciocan, D.; Abdeen, S.K.; Elinav, E. Utilization of the microbiome in personalized medicine. Nat. Rev. Microbiol. 2023. [Google Scholar] [CrossRef] [PubMed]
- Warren, T.; McAllister, R.; Morgan, A.; Rai, T.S.; McGilligan, V.; Ennis, M.; Page, C.; Kelly, C.; Peace, A.; Corfe, B.M.; et al. The Interdependency and Co-Regulation of the Vitamin D and Cholesterol Metabolism. Cells 2021, 10, 2007. [Google Scholar] [CrossRef]
- Mizwicki, M.T.; Keidel, D.; Bula, C.M.; Bishop, J.E.; Zanello, L.P.; Wurtz, J.M.; Moras, D.; Norman, A.W. Identification of an alternative ligand-binding pocket in the nuclear vitamin D receptor and its functional importance in 1alpha,25(OH)2-vitamin D3 signaling. Proc. Natl. Acad. Sci. USA 2004, 101, 12876–12881. [Google Scholar] [CrossRef] [PubMed]
- Chishimba, L.; Thickett, D.R.; Stockley, R.A.; Wood, A.M. The vitamin D axis in the lung: A key role for vitamin D-binding protein. Thorax 2010, 65, 456–462. [Google Scholar] [CrossRef] [PubMed]
- Dancer, R.C.; Parekh, D.; Lax, S.; D’Souza, V.; Zheng, S.; Bassford, C.R.; Park, D.; Bartis, D.G.; Mahida, R.; Turner, A.M.; et al. Vitamin D deficiency contributes directly to the acute respiratory distress syndrome (ARDS). Thorax 2015, 70, 617–624. [Google Scholar] [CrossRef]
- Krasniqi, E.; Boshnjaku, A.; Wagner, K.H.; Wessner, B. Association between Polymorphisms in Vitamin D Pathway-Related Genes, Vitamin D Status, Muscle Mass and Function: A Systematic Review. Nutrients 2021, 13, 3109. [Google Scholar] [CrossRef]
- Bollen, S.E.; Bass, J.J.; Fujita, S.; Wilkinson, D.; Hewison, M.; Atherton, P.J. The Vitamin D/Vitamin D receptor (VDR) axis in muscle atrophy and sarcopenia. Cell Signal 2022, 96, 110355. [Google Scholar] [CrossRef]
- Pusceddu, I.; Farrell, C.J.; Di Pierro, A.M.; Jani, E.; Herrmann, W.; Herrmann, M. The role of telomeres and vitamin D in cellular aging and age-related diseases. Clin. Chem. Lab. Med. 2015, 53, 1661–1678. [Google Scholar] [CrossRef]
- Janubova, M.; Zitnanova, I. The effects of vitamin D on different types of cells. Steroids 2024, 202, 109350. [Google Scholar] [CrossRef]
- Lestari, M.I.; Murti, K.; Liberty, I.A.; Hafy, Z.; Linardi, V.; Khoirudin, M.; Umar, T.P. Association of vitamin D with deoxyribonucleic acid (dna) damage: A systematic review of animal and human studies. Acta Biochim. Pol. 2023, 70, 379–387. [Google Scholar] [CrossRef]
- Nair-Shalliker, V.; Armstrong, B.K.; Fenech, M. Does vitamin D protect against DNA damage? Mutat. Res. 2012, 733, 50–57. [Google Scholar] [CrossRef]
- Graziano, S.; Johnston, R.; Deng, O.; Zhang, J.; Gonzalo, S. Vitamin D/vitamin D receptor axis regulates DNA repair during oncogene-induced senescence. Oncogene 2016, 35, 5362–5376. [Google Scholar] [CrossRef] [PubMed]
- Fedirko, V.; Bostick, R.M.; Long, Q.; Flanders, W.D.; McCullough, M.L.; Sidelnikov, E.; Daniel, C.R.; Rutherford, R.E.; Shaukat, A. Effects of supplemental vitamin D and calcium on oxidative DNA damage marker in normal colorectal mucosa: A randomized clinical trial. Cancer Epidemiol. Biomarkers Prev. 2010, 19, 280–291. [Google Scholar] [CrossRef]
- Wenclewska, S.; Szymczak-Pajor, I.; Drzewoski, J.; Bunk, M.; Sliwinska, A. Vitamin D Supplementation Reduces Both Oxidative DNA Damage and Insulin Resistance in the Elderly with Metabolic Disorders. Int. J. Mol. Sci. 2019, 20, 2891. [Google Scholar] [CrossRef]
- Fagundes, G.E.; Macan, T.P.; Rohr, P.; Damiani, A.P.; Da Rocha, F.R.; Pereira, M.; Longaretti, L.M.; Vilela, T.C.; Ceretta, L.B.; Mendes, C.; et al. Vitamin D3 as adjuvant in the treatment of type 2 diabetes mellitus: Modulation of genomic and biochemical instability. Mutagenesis 2019, 34, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.B.; Valdes, A.M.; Gardner, J.P.; Paximadas, D.; Kimura, M.; Nessa, A.; Lu, X.; Surdulescu, G.L.; Swaminathan, R.; Spector, T.D.; et al. Higher serum vitamin D concentrations are associated with longer leukocyte telomere length in women. Am. J. Clin. Nutr. 2007, 86, 1420–1425. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.J.; Prescott, J.; Giovannucci, E.; Hankinson, S.E.; Rosner, B.; Han, J.; De Vivo, I. Plasma vitamin D biomarkers and leukocyte telomere length. Am. J. Epidemiol. 2013, 177, 1411–1417. [Google Scholar] [CrossRef] [PubMed]
- Julin, B.; Shui, I.M.; Prescott, J.; Giovannucci, E.L.; De Vivo, I. Plasma vitamin D biomarkers and leukocyte telomere length in men. Eur. J. Nutr. 2017, 56, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.M.; Palaniswamy, S.; Sebert, S.; Buxton, J.L.; Blakemore, A.I.; Hypponen, E.; Jarvelin, M.R. 25-Hydroxyvitamin D Concentration and Leukocyte Telomere Length in Young Adults: Findings From the Northern Finland Birth Cohort 1966. Am. J. Epidemiol. 2016, 183, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Hakeem, S.; Mendonca, N.; Aspray, T.; Kingston, A.; Martin-Ruiz, C.; Robinson, L.; Hill, T.R. The Association between 25-Hydroxyvitamin D Concentration and Telomere Length in the Very-Old: The Newcastle 85+ Study. Nutrients 2021, 13, 4341. [Google Scholar] [CrossRef]
- Hoffecker, B.M.; Raffield, L.M.; Kamen, D.L.; Nowling, T.K. Systemic lupus erythematosus and vitamin D deficiency are associated with shorter telomere length among African Americans: A case-control study. PLoS ONE 2013, 8, e63725. [Google Scholar] [CrossRef]
- Borras, M.; Panizo, S.; Sarro, F.; Valdivielso, J.M.; Fernandez, E. Assessment of the potential role of active vitamin D treatment in telomere length: A case-control study in hemodialysis patients. Clin. Ther. 2012, 34, 849–856. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.V.; Chen, Y.C.; Wu, W.T.; Shen, H.J.; Huang, K.C.; Chu, H.P.; Han, D.S. Expression of Telomeric Repeat-Containing RNA Decreases in Sarcopenia and Increases after Exercise and Nutrition Intervention. Nutrients 2020, 12, 3766. [Google Scholar] [CrossRef] [PubMed]
- Maicher, A.; Kastner, L.; Luke, B. Telomeres and disease: Enter TERRA. RNA Biol. 2012, 9, 843–849. [Google Scholar] [CrossRef]
- Mazidi, M.; Mikhailidis, D.P.; Banach, M.; Dehghan, A. Impact of serum 25-hydroxyvitamin D 25(OH) on telomere attrition: A Mendelian Randomization study. Clin. Nutr. 2020, 39, 2730–2733. [Google Scholar] [CrossRef]
- Zhu, H.; Guo, D.; Li, K.; Pedersen-White, J.; Stallmann-Jorgensen, I.S.; Huang, Y.; Parikh, S.; Liu, K.; Dong, Y. Increased telomerase activity and vitamin D supplementation in overweight African Americans. Int. J. Obes. 2012, 36, 805–809. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Dong, Y.; Bhagatwala, J.; Raed, A.; Huang, Y.; Zhu, H. Effects of Vitamin D3 Supplementation on Epigenetic Aging in Overweight and Obese African Americans With Suboptimal Vitamin D Status: A Randomized Clinical Trial. J. Gerontol. A Biol. Sci. Med. Sci. 2019, 74, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Bhagatwala, J.; Huang, Y.; Pollock, N.K.; Parikh, S.; Raed, A.; Gutin, B.; Harshfield, G.A.; Dong, Y. Race/Ethnicity-Specific Association of Vitamin D and Global DNA Methylation: Cross-Sectional and Interventional Findings. PLoS ONE 2016, 11, e0152849. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Wang, X.; Shi, H.; Su, S.; Harshfield, G.A.; Gutin, B.; Snieder, H.; Dong, Y. A genome-wide methylation study of severe vitamin D deficiency in African American adolescents. J. Pediatr. 2013, 162, 1004–1009.e1. [Google Scholar] [CrossRef] [PubMed]
- Harvey, N.C.; Sheppard, A.; Godfrey, K.M.; McLean, C.; Garratt, E.; Ntani, G.; Davies, L.; Murray, R.; Inskip, H.M.; Gluckman, P.D.; et al. Childhood bone mineral content is associated with methylation status of the RXRA promoter at birth. J. Bone Miner. Res. 2014, 29, 600–607. [Google Scholar] [CrossRef] [PubMed]
- Curtis, E.M.; Krstic, N.; Cook, E.; D’Angelo, S.; Crozier, S.R.; Moon, R.J.; Murray, R.; Garratt, E.; Costello, P.; Cleal, J.; et al. Gestational Vitamin D Supplementation Leads to Reduced Perinatal RXRA DNA Methylation: Results From the MAVIDOS Trial. J. Bone Miner. Res. 2019, 34, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.M.; Gillespie, S.L.; Thiele, D.K.; Ralph, J.L.; Ohm, J.E. Effects of Maternal Vitamin D Supplementation on the Maternal and Infant Epigenome. Breastfeed. Med. 2018, 13, 371–380. [Google Scholar] [CrossRef]
- Bollen, S.E.; Atherton, P.J. Myogenic, genomic and non-genomic influences of the vitamin D axis in skeletal muscle. Cell Biochem. Funct. 2021, 39, 48–59. [Google Scholar] [CrossRef]
- Reis, N.G.; Assis, A.P.; Lautherbach, N.; Goncalves, D.A.; Silveira, W.A.; Morgan, H.J.N.; Valentim, R.R.; Almeida, L.F.; Heck, L.C.; Zanon, N.M.; et al. Maternal vitamin D deficiency affects the morphology and function of glycolytic muscle in adult offspring rats. J. Cachexia Sarcopenia Muscle 2022, 13, 2175–2187. [Google Scholar] [CrossRef]
- Das, A.; Jawla, N.; Meena, V.; Gopinath, S.D.; Arimbasseri, G.A. Lack of vitamin D signalling shifts skeletal muscles towards oxidative metabolism. J. Cachexia Sarcopenia Muscle 2023, 15, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Ricca, C.; Aillon, A.; Bergandi, L.; Alotto, D.; Castagnoli, C.; Silvagno, F. Vitamin D Receptor Is Necessary for Mitochondrial Function and Cell Health. Int. J. Mol. Sci. 2018, 19, 1672. [Google Scholar] [CrossRef] [PubMed]
- Hua, R.; Liu, B.; He, W.; Zhang, H.; Liu, Y.; Xie, Q.; Zhou, L.; Pei, F. Calcitriol reverses age-related hypertension via downregulating renal AP1/AT(1)R pathway through regulating mitochondrial function. Clin. Exp. Hypertens. 2023, 45, 2277653. [Google Scholar] [CrossRef] [PubMed]
- Mohanad, M.; Mohamed, S.K.; Aboulhoda, B.E.; Ahmed, M.A.E. Neuroprotective effects of vitamin D in an Alzheimer’s disease rat model: Improvement of mitochondrial dysfunction via calcium/calmodulin-dependent protein kinase kinase 2 activation of Sirtuin1 phosphorylation. Biofactors 2023. [Google Scholar] [CrossRef] [PubMed]
- Russo, C.; Valle, M.S.; Casabona, A.; Spicuzza, L.; Sambataro, G.; Malaguarnera, L. Vitamin D Impacts on Skeletal Muscle Dysfunction in Patients with COPD Promoting Mitochondrial Health. Biomedicines 2022, 10, 898. [Google Scholar] [CrossRef] [PubMed]
- de Las Heras, N.; Martin Gimenez, V.M.; Ferder, L.; Manucha, W.; Lahera, V. Implications of Oxidative Stress and Potential Role of Mitochondrial Dysfunction in COVID-19: Therapeutic Effects of Vitamin D. Antioxidants 2020, 9, 897. [Google Scholar] [CrossRef]
- Lee, T.W.; Kao, Y.H.; Chen, Y.J.; Chao, T.F.; Lee, T.I. Therapeutic potential of vitamin D in AGE/RAGE-related cardiovascular diseases. Cell Mol. Life Sci. 2019, 76, 4103–4115. [Google Scholar] [CrossRef] [PubMed]
- Phillips, E.A.; Hendricks, N.; Bucher, M.; Maloyan, A. Vitamin D Supplementation Improves Mitochondrial Function and Reduces Inflammation in Placentae of Obese Women. Front. Endocrinol. 2022, 13, 893848. [Google Scholar] [CrossRef]
- Yu, M.; Wu, H.; Wang, J.; Chen, X.; Pan, J.; Liu, P.; Zhang, J.; Chen, Y.; Zhu, W.; Tang, C.; et al. Vitamin D receptor inhibits EMT via regulation of the epithelial mitochondrial function in intestinal fibrosis. J. Biol. Chem. 2021, 296, 100531. [Google Scholar] [CrossRef]
- Romani, M.; Berger, M.M.; D’Amelio, P. From the Bench to the Bedside: Branched Amino Acid and Micronutrient Strategies to Improve Mitochondrial Dysfunction Leading to Sarcopenia. Nutrients 2022, 14, 483. [Google Scholar] [CrossRef] [PubMed]
- Martinelli, R.P.; Rayego-Mateos, S.; Alique, M.; Marquez-Exposito, L.; Tejedor-Santamaria, L.; Ortiz, A.; Gonzalez-Parra, E.; Ruiz-Ortega, M. Vitamin D, Cellular Senescence and Chronic Kidney Diseases: What Is Missing in the Equation? Nutrients 2023, 15, 1349. [Google Scholar] [CrossRef] [PubMed]
- Yousefzadeh, M.J.; Flores, R.R.; Zhu, Y.; Schmiechen, Z.C.; Brooks, R.W.; Trussoni, C.E.; Cui, Y.; Angelini, L.; Lee, K.A.; McGowan, S.J.; et al. An aged immune system drives senescence and ageing of solid organs. Nature 2021, 594, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Maselli Del Giudice, A.; La Mantia, I.; Barbara, F.; Ciccarone, S.; Ragno, M.S.; de Robertis, V.; Cariti, F.; Barbara, M.; D’Ascanio, L.; Di Stadio, A. Use of Nutraceuticals in Elderly to Fight Inflammation and Immuno-Senescence: A Randomized Case-Control Study. Nutrients 2022, 14, 3476. [Google Scholar] [CrossRef] [PubMed]
- Rizka, A.; Setiati, S.; Harimurti, K.; Sadikin, M.; Mansur, I.G. Effect of Alfacalcidol on Inflammatory markers and T Cell Subsets in Elderly with Frailty Syndrome: A Double Blind Randomized Controlled Trial. Acta Med. Indones. 2018, 50, 215–221. [Google Scholar] [PubMed]
- Martin, N.; Zhu, K.; Czarnecka-Herok, J.; Vernier, M.; Bernard, D. Regulation and role of calcium in cellular senescence. Cell Calcium 2023, 110, 102701. [Google Scholar] [CrossRef]
- Mahdavi, R.; Belgheisi, G.; Haghbin-Nazarpak, M.; Omidi, M.; Khojasteh, A.; Solati-Hashjin, M. Bone tissue engineering gelatin-hydroxyapatite/graphene oxide scaffolds with the ability to release vitamin D: Fabrication, characterization, and in vitro study. J. Mater. Sci. Mater. Med. 2020, 31, 97. [Google Scholar] [CrossRef]
- Bottai, D.; Fiocco, R.; Gelain, F.; Defilippis, L.; Galli, R.; Gritti, A.; Vescovi, L.A. Neural stem cells in the adult nervous system. J. Hematotherapy Stem Cell Res. 2003, 12, 655–670. [Google Scholar] [CrossRef] [PubMed]
- Uthaiah, C.A.; Beeraka, N.M.; Rajalakshmi, R.; Ramya, C.M.; Madhunapantula, S.V. Role of Neural Stem Cells and Vitamin D Receptor (VDR)-Mediated Cellular Signaling in the Mitigation of Neurological Diseases. Mol. Neurobiol. 2022, 59, 4065–4105. [Google Scholar] [CrossRef]
- Mouchel, P.L.; Berard, E.; Tavitian, S.; Gadaud, N.; Vergez, F.; Rieu, J.B.; Luquet, I.; Sarry, A.; Huguet, F.; Largeaud, L.; et al. Vitamin C and D supplementation in acute myeloid leukemia. Blood Adv. 2023, 7, 6886–6897. [Google Scholar] [CrossRef]
- Paubelle, E.; Zylbersztejn, F.; Maciel, T.T.; Carvalho, C.; Mupo, A.; Cheok, M.; Lieben, L.; Sujobert, P.; Decroocq, J.; Yokoyama, A.; et al. Vitamin D Receptor Controls Cell Stemness in Acute Myeloid Leukemia and in Normal Bone Marrow. Cell Rep. 2020, 30, 739–754.e734. [Google Scholar] [CrossRef]
- Cao, H.; Xu, Y.; de Necochea-Campion, R.; Baylink, D.J.; Payne, K.J.; Tang, X.; Ratanatharathorn, C.; Ji, Y.; Mirshahidi, S.; Chen, C.S. Application of vitamin D and vitamin D analogs in acute myelogenous leukemia. Exp. Hematol. 2017, 50, 1–12. [Google Scholar] [CrossRef]
- Dantas, J.R.; Araujo, D.B.; Silva, K.R.; Souto, D.L.; de Fatima Carvalho Pereira, M.; Luiz, R.R.; Dos Santos Mantuano, M.; Claudio-da-Silva, C.; Gabbay, M.A.L.; Dib, S.A.; et al. Adipose tissue-derived stromal/stem cells + cholecalciferol: A pilot study in recent-onset type 1 diabetes patients. Arch. Endocrinol. Metab. 2021, 65, 342–351. [Google Scholar] [CrossRef]
- Bloemen, V.; Schoenmaker, T.; de Vries, T.J.; Everts, V. Direct cell-cell contact between periodontal ligament fibroblasts and osteoclast precursors synergistically increases the expression of genes related to osteoclastogenesis. J. Cell Physiol. 2010, 222, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Schirrmacher, K.; Bingmann, D. Effects of vitamin D3, 17beta-estradiol, vasoactive intestinal peptide, and glutamate on electric coupling between rat osteoblast-like cells in vitro. Bone 1998, 23, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Sirbe, C.; Rednic, S.; Grama, A.; Pop, T.L. An Update on the Effects of Vitamin D on the Immune System and Autoimmune Diseases. Int. J. Mol. Sci. 2022, 23, 9784. [Google Scholar] [CrossRef]
- Mansournia, M.A.; Ostadmohammadi, V.; Doosti-Irani, A.; Ghayour-Mobarhan, M.; Ferns, G.; Akbari, H.; Ghaderi, A.; Talari, H.R.; Asemi, Z. The Effects of Vitamin D Supplementation on Biomarkers of Inflammation and Oxidative Stress in Diabetic Patients: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Horm. Metab. Res. 2018, 50, 429–440. [Google Scholar] [CrossRef] [PubMed]
- Jamilian, H.; Amirani, E.; Milajerdi, A.; Kolahdooz, F.; Mirzaei, H.; Zaroudi, M.; Ghaderi, A.; Asemi, Z. The effects of vitamin D supplementation on mental health, and biomarkers of inflammation and oxidative stress in patients with psychiatric disorders: A systematic review and meta-analysis of randomized controlled trials. Prog. Neuropsychopharmacol. Biol. Psychiatry 2019, 94, 109651. [Google Scholar] [CrossRef] [PubMed]
- Akbari, M.; Ostadmohammadi, V.; Lankarani, K.B.; Tabrizi, R.; Kolahdooz, F.; Heydari, S.T.; Kavari, S.H.; Mirhosseini, N.; Mafi, A.; Dastorani, M.; et al. The Effects of Vitamin D Supplementation on Biomarkers of Inflammation and Oxidative Stress among Women with Polycystic Ovary Syndrome: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Horm. Metab. Res. 2018, 50, 271–279. [Google Scholar] [CrossRef]
- Mansouri, L.; Lundwall, K.; Moshfegh, A.; Jacobson, S.H.; Lundahl, J.; Spaak, J. Vitamin D receptor activation reduces inflammatory cytokines and plasma MicroRNAs in moderate chronic kidney disease—A randomized trial. BMC Nephrol. 2017, 18, 161. [Google Scholar] [CrossRef]
- Mousa, A.; Naderpoor, N.; Teede, H.; Scragg, R.; de Courten, B. Vitamin D supplementation for improvement of chronic low-grade inflammation in patients with type 2 diabetes: A systematic review and meta-analysis of randomized controlled trials. Nutr. Rev. 2018, 76, 380–394. [Google Scholar] [CrossRef]
- Autier, P.; Mullie, P.; Macacu, A.; Dragomir, M.; Boniol, M.; Coppens, K.; Pizot, C.; Boniol, M. Effect of vitamin D supplementation on non-skeletal disorders: A systematic review of meta-analyses and randomised trials. Lancet Diabetes Endocrinol. 2017, 5, 986–1004. [Google Scholar] [CrossRef]
- Dong, Y.; Zhu, H.; Chen, L.; Huang, Y.; Christen, W.; Cook, N.R.; Copeland, T.; Mora, S.; Buring, J.E.; Lee, I.M.; et al. Effects of Vitamin D(3) and Marine Omega-3 Fatty Acids Supplementation on Biomarkers of Systemic Inflammation: 4-Year Findings from the VITAL Randomized Trial. Nutrients 2022, 14, 5307. [Google Scholar] [CrossRef]
- Missailidis, C.; Sorensen, N.; Ashenafi, S.; Amogne, W.; Kassa, E.; Bekele, A.; Getachew, M.; Gebreselassie, N.; Aseffa, A.; Aderaye, G.; et al. Vitamin D and Phenylbutyrate Supplementation Does Not Modulate Gut Derived Immune Activation in HIV-1. Nutrients 2019, 11, 1675. [Google Scholar] [CrossRef]
- Ramasamy, B.; Magne, F.; Tripathy, S.K.; Venugopal, G.; Mukherjee, D.; Balamurugan, R. Association of Gut Microbiome and Vitamin D Deficiency in Knee Osteoarthritis Patients: A Pilot Study. Nutrients 2021, 13, 1272. [Google Scholar] [CrossRef]
- Kanhere, M.; He, J.; Chassaing, B.; Ziegler, T.R.; Alvarez, J.A.; Ivie, E.A.; Hao, L.; Hanfelt, J.; Gewirtz, A.T.; Tangpricha, V. Bolus Weekly Vitamin D3 Supplementation Impacts Gut and Airway Microbiota in Adults With Cystic Fibrosis: A Double-Blind, Randomized, Placebo-Controlled Clinical Trial. J. Clin. Endocrinol. Metab. 2018, 103, 564–574. [Google Scholar] [CrossRef]
- Singh, P.; Rawat, A.; Alwakeel, M.; Sharif, E.; Al Khodor, S. The potential role of vitamin D supplementation as a gut microbiota modifier in healthy individuals. Sci. Rep. 2020, 10, 21641. [Google Scholar] [CrossRef]
- Martin, N.; Bernard, D. Calcium signaling and cellular senescence. Cell Calcium 2018, 70, 16–23. [Google Scholar] [CrossRef]
- Terrell, K.; Choi, S.; Choi, S. Calcium’s Role and Signaling in Aging Muscle, Cellular Senescence, and Mineral Interactions. Int. J. Mol. Sci. 2023, 24, 7034. [Google Scholar] [CrossRef]
- Bertoldo, F.; Cianferotti, L.; Di Monaco, M.; Falchetti, A.; Fassio, A.; Gatti, D.; Gennari, L.; Giannini, S.; Girasole, G.; Gonnelli, S.; et al. Definition, Assessment, and Management of Vitamin D Inadequacy: Suggestions, Recommendations, and Warnings from the Italian Society for Osteoporosis, Mineral Metabolism and Bone Diseases (SIOMMMS). Nutrients 2022, 14, 4148. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruggiero, C.; Tafaro, L.; Cianferotti, L.; Tramontana, F.; Macchione, I.G.; Caffarelli, C.; Virdis, A.; Ferracci, M.; Rinonapoli, G.; Mecocci, P.; et al. Targeting the Hallmarks of Aging with Vitamin D: Starting to Decode the Myth. Nutrients 2024, 16, 906. https://doi.org/10.3390/nu16060906
Ruggiero C, Tafaro L, Cianferotti L, Tramontana F, Macchione IG, Caffarelli C, Virdis A, Ferracci M, Rinonapoli G, Mecocci P, et al. Targeting the Hallmarks of Aging with Vitamin D: Starting to Decode the Myth. Nutrients. 2024; 16(6):906. https://doi.org/10.3390/nu16060906
Chicago/Turabian StyleRuggiero, Carmelinda, Laura Tafaro, Luisella Cianferotti, Flavia Tramontana, Ilaria Giovanna Macchione, Carla Caffarelli, Agostino Virdis, Marika Ferracci, Giuseppe Rinonapoli, Patrizia Mecocci, and et al. 2024. "Targeting the Hallmarks of Aging with Vitamin D: Starting to Decode the Myth" Nutrients 16, no. 6: 906. https://doi.org/10.3390/nu16060906
APA StyleRuggiero, C., Tafaro, L., Cianferotti, L., Tramontana, F., Macchione, I. G., Caffarelli, C., Virdis, A., Ferracci, M., Rinonapoli, G., Mecocci, P., Napoli, N., & Calsolaro, V. (2024). Targeting the Hallmarks of Aging with Vitamin D: Starting to Decode the Myth. Nutrients, 16(6), 906. https://doi.org/10.3390/nu16060906