
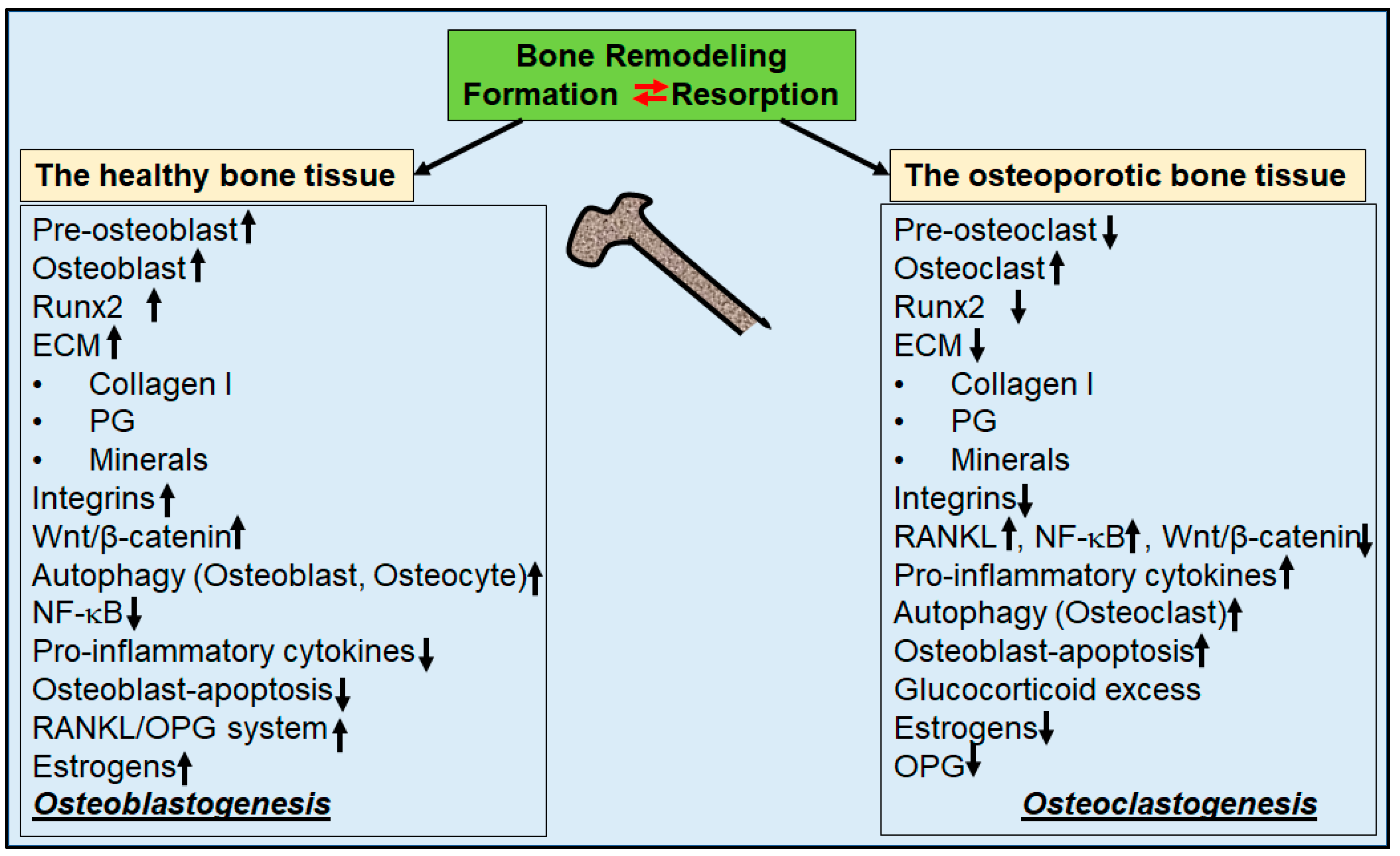
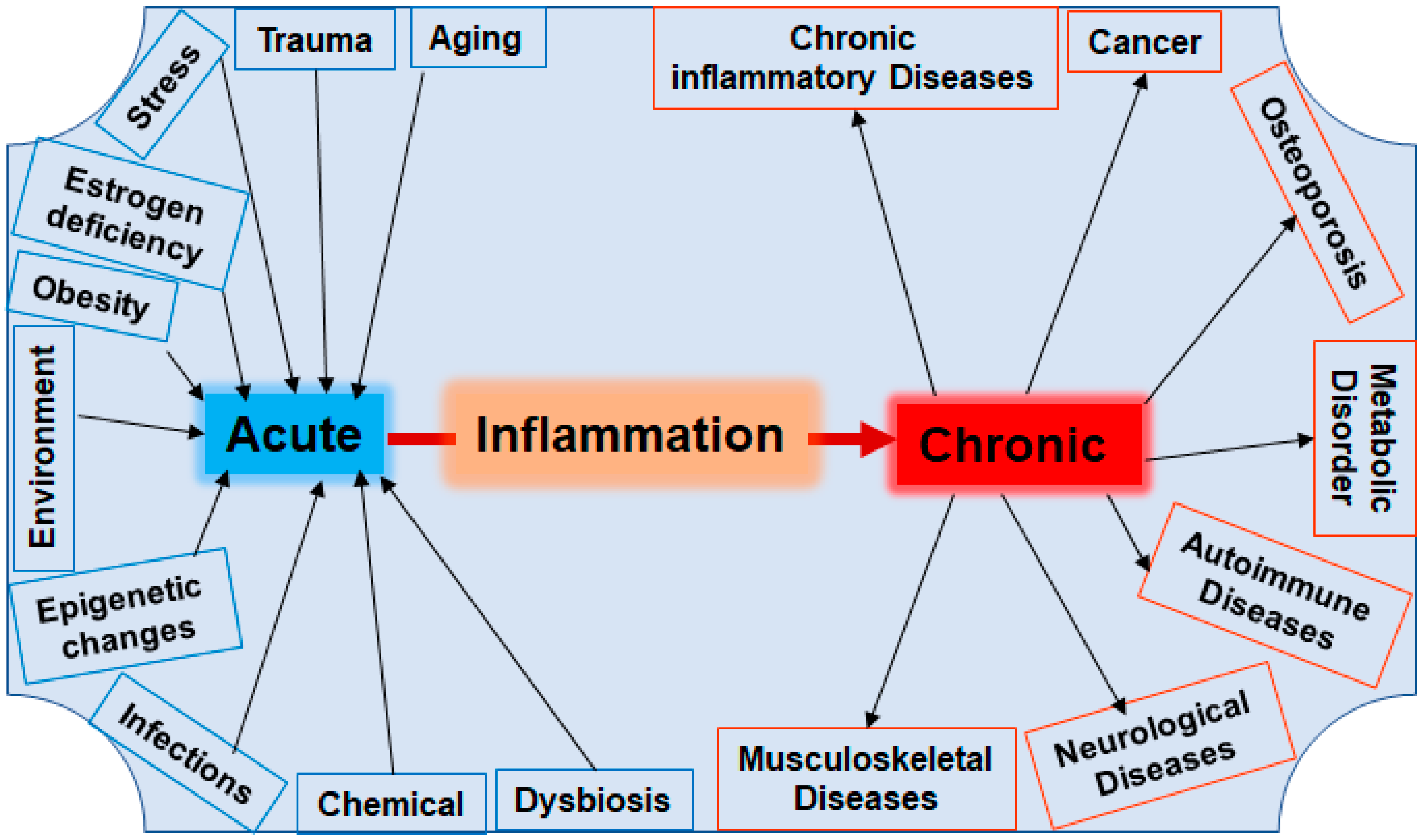
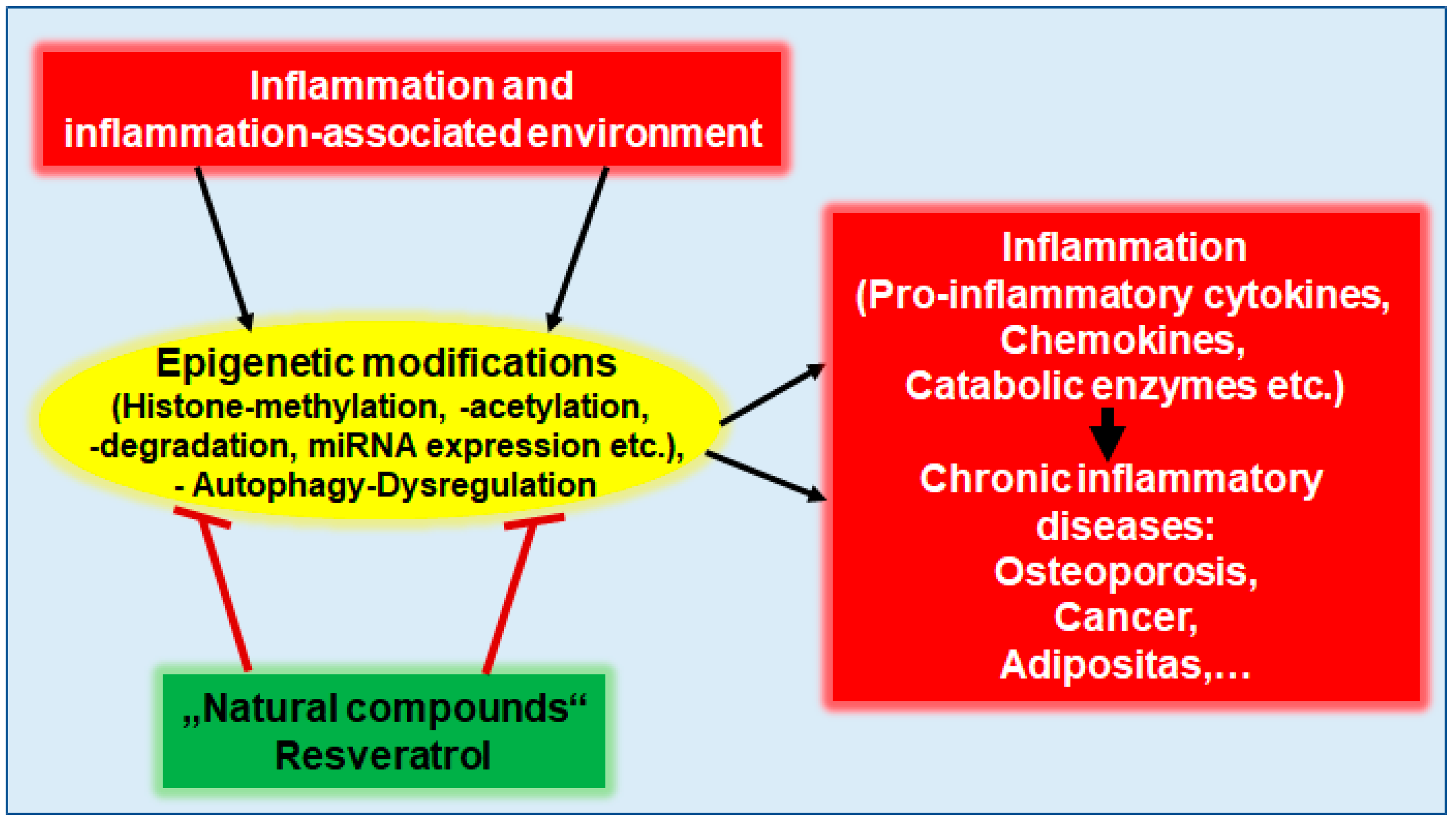
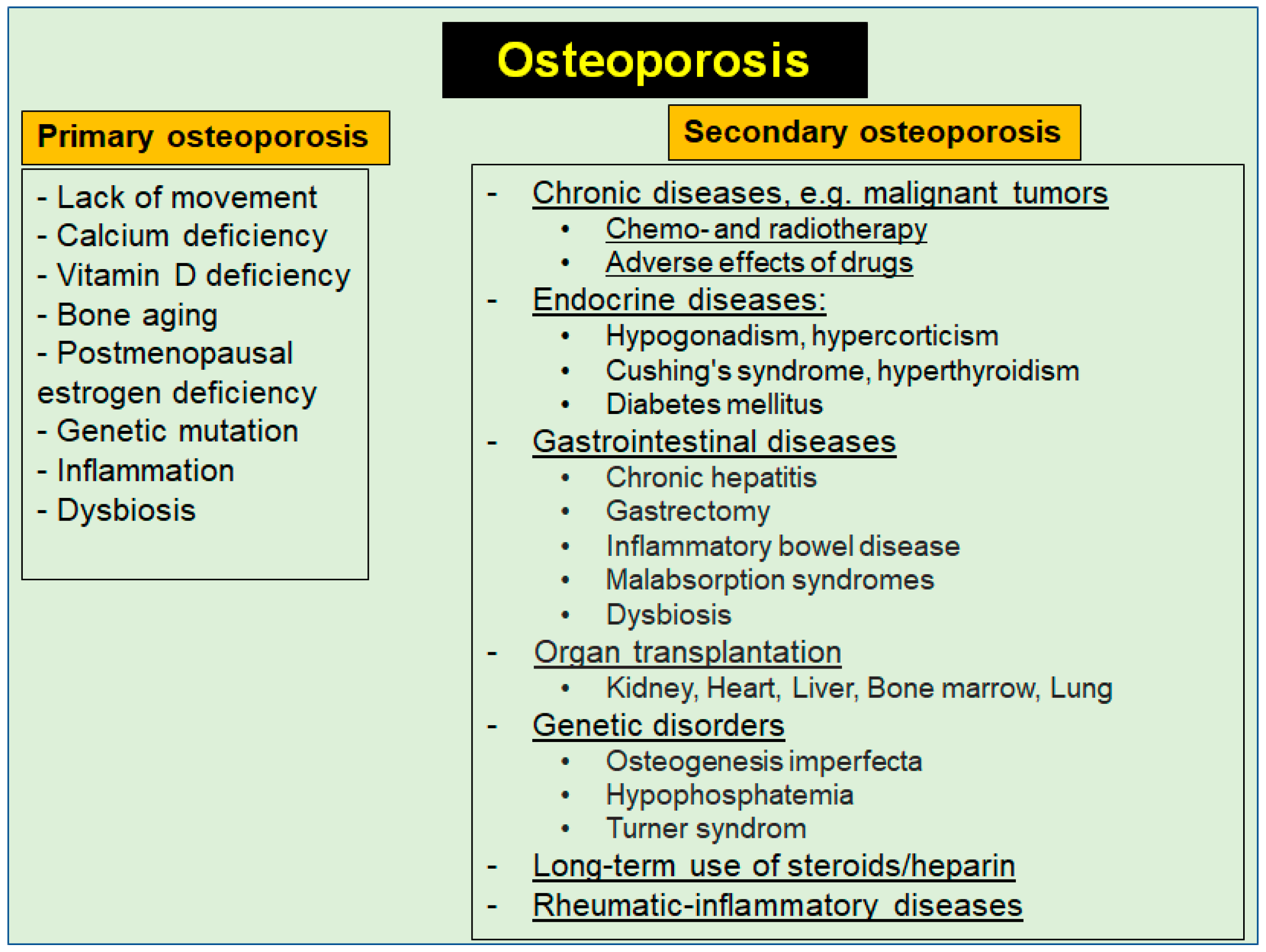
Prevention and Co-Management of Breast Cancer-Related Osteoporosis Using Resveratrol



{kind=link}
{kind=link}
{kind=link}
{kind=link}
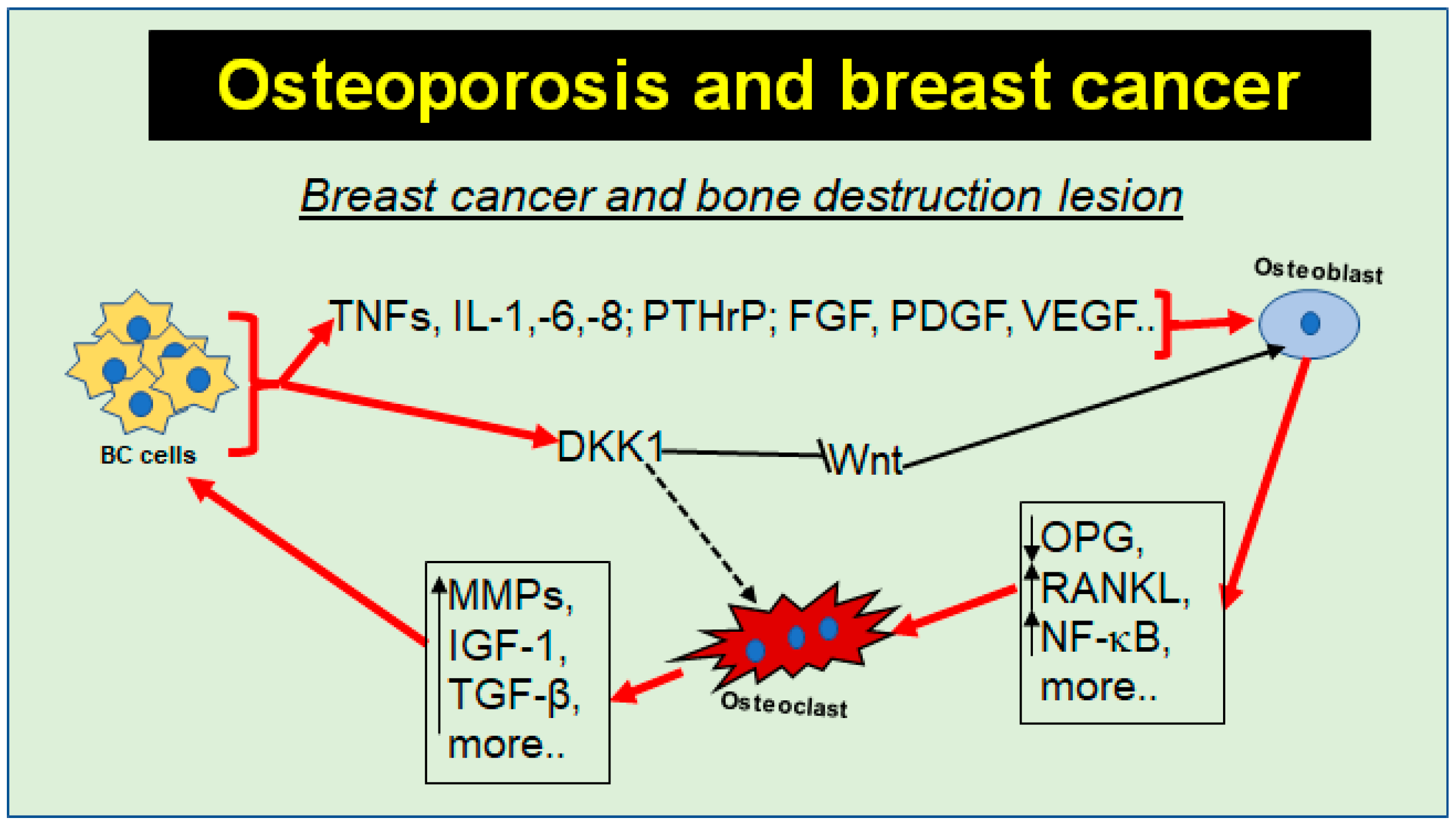{kind=link}
{kind=link}
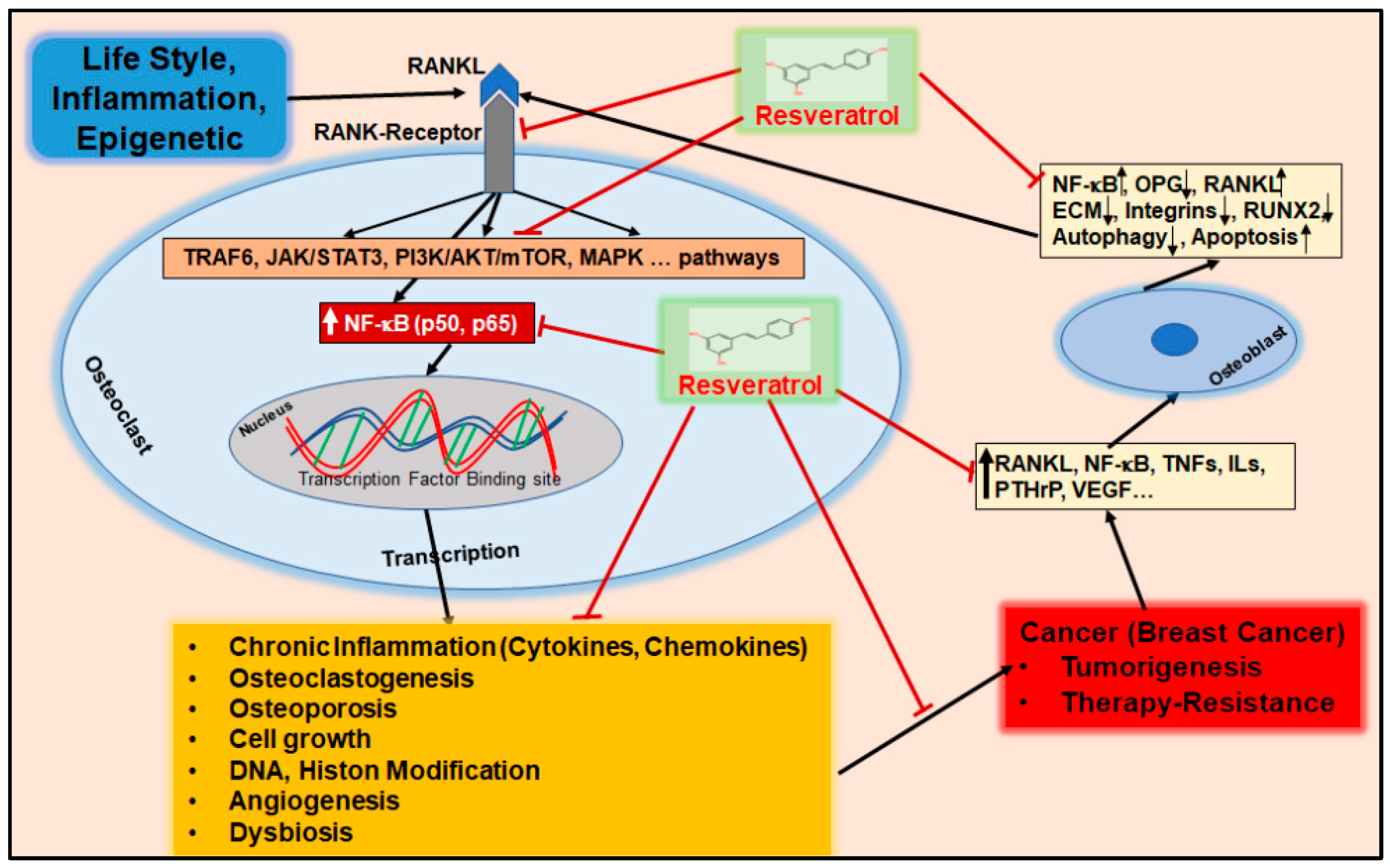{kind=link}
Abstract
Share and Cite
Meyer, C.; Brockmueller, A.; Buhrmann, C.; Shakibaei, M. Prevention and Co-Management of Breast Cancer-Related Osteoporosis Using Resveratrol. Nutrients 2024, 16, 708. https://doi.org/10.3390/nu16050708
Meyer C, Brockmueller A, Buhrmann C, Shakibaei M. Prevention and Co-Management of Breast Cancer-Related Osteoporosis Using Resveratrol. Nutrients. 2024; 16(5):708. https://doi.org/10.3390/nu16050708
Chicago/Turabian StyleMeyer, Christine, Aranka Brockmueller, Constanze Buhrmann, and Mehdi Shakibaei. 2024. "Prevention and Co-Management of Breast Cancer-Related Osteoporosis Using Resveratrol" Nutrients 16, no. 5: 708. https://doi.org/10.3390/nu16050708
APA StyleMeyer, C., Brockmueller, A., Buhrmann, C., & Shakibaei, M. (2024). Prevention and Co-Management of Breast Cancer-Related Osteoporosis Using Resveratrol. Nutrients, 16(5), 708. https://doi.org/10.3390/nu16050708