

Plasma One-Carbon Metabolism-Related Micronutrients and the Risk of Breast Cancer: Involvement of DNA Methylation

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Data and Sample Collection
2.3. Measurements of Plasma One-Carbon Metabolism-Related Micronutrients
2.4. DNA Methylation Assay
2.5. Data Processing for DNA Methylation Array
2.6. Statistical Analysis
3. Results
3.1. Characteristics of Study Population
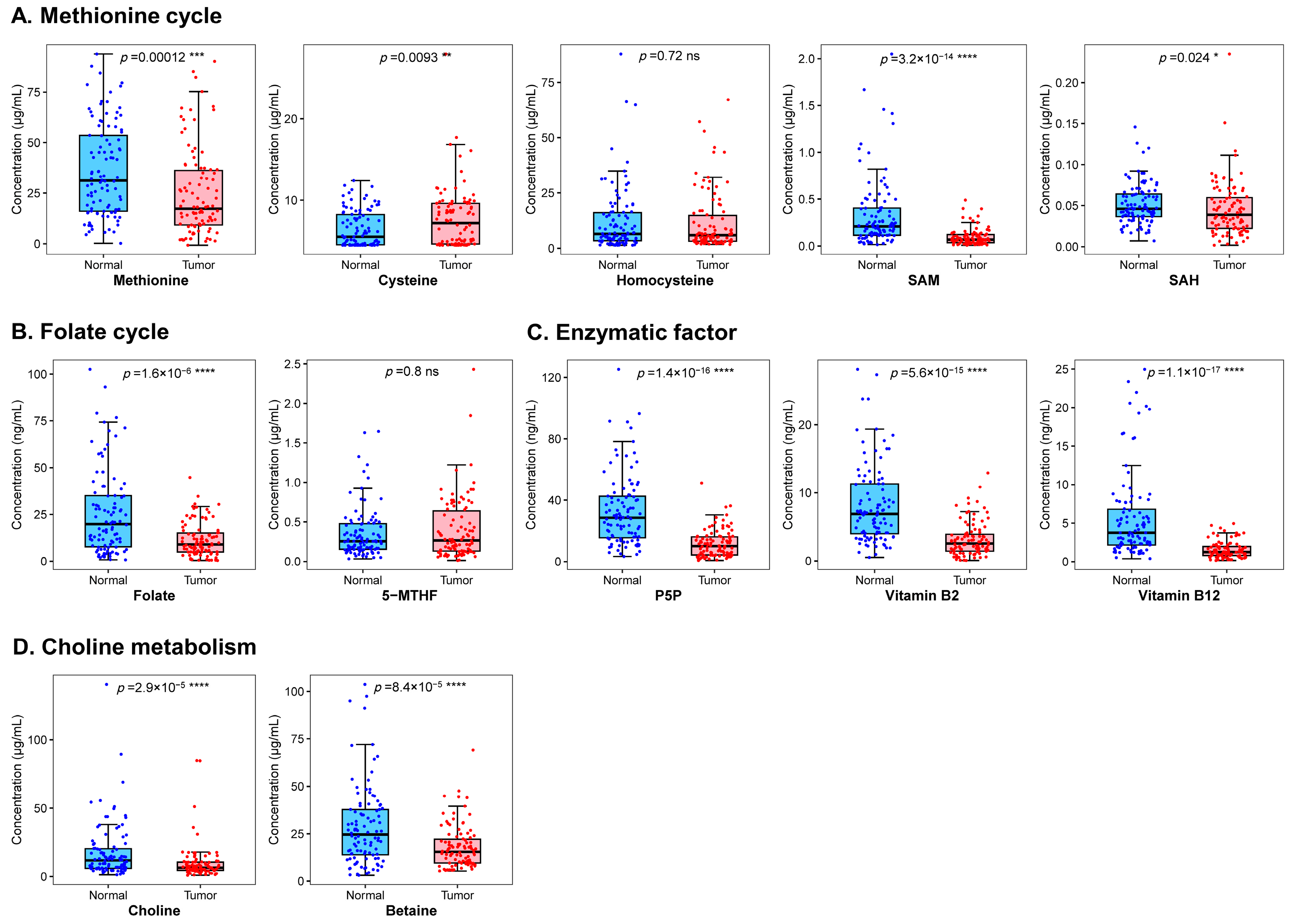
3.2. Association between One-Carbon Metabolism-Related Micronutrients and Breast Cancer Risk
3.3. Overall and Differential Analysis of DNA Methylation
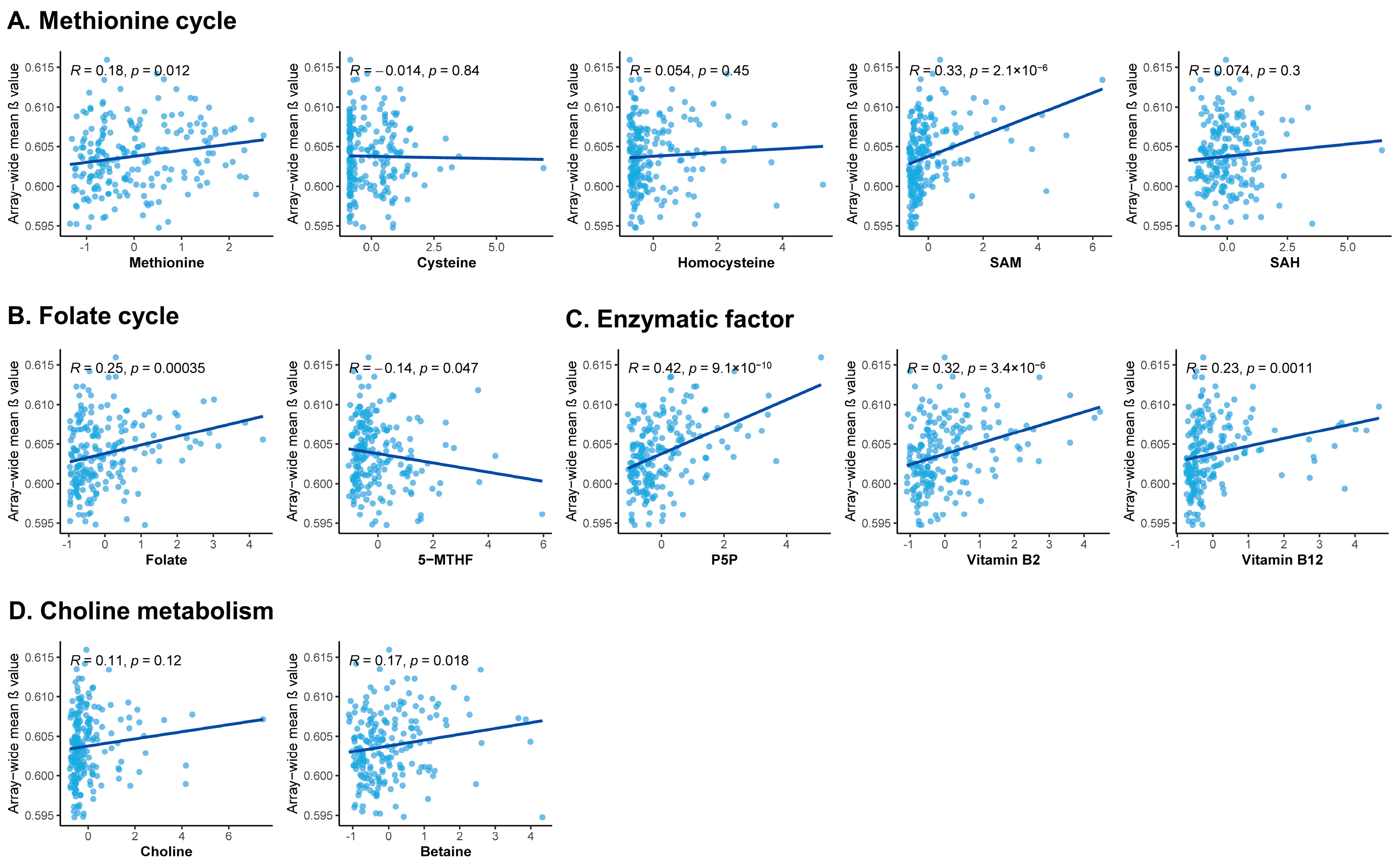
3.4. Correlation between One-Carbon Metabolism-Related Micronutrients and DNA Methylation
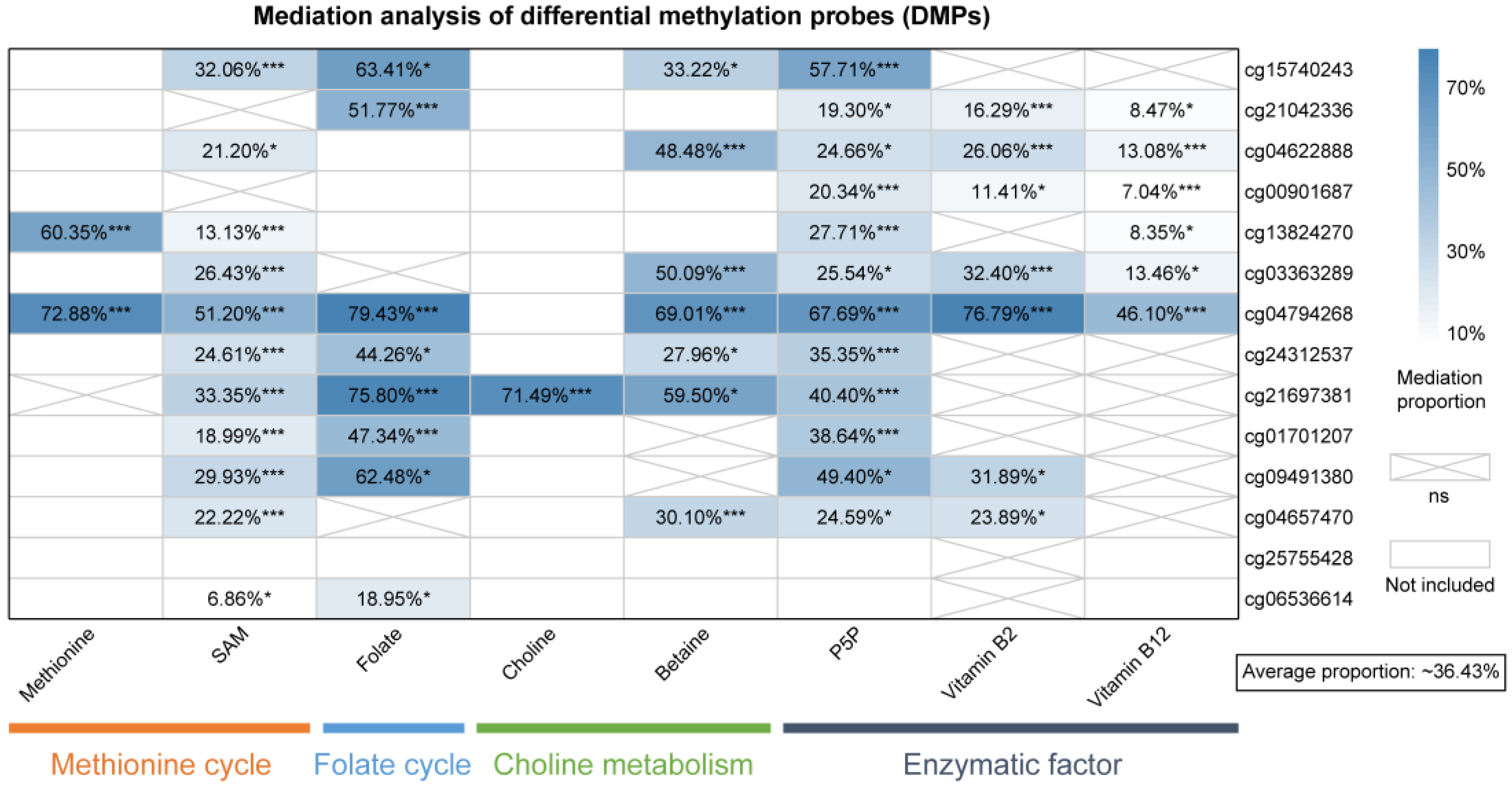
3.5. Mediation of DNA Methylation in the Micronutrients-Breast Cancer Associations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- De Cicco, P.; Catani, M.V.; Gasperi, V.; Sibilano, M.; Quaglietta, M.; Savini, I. Nutrition and Breast Cancer: A Literature Review on Prevention, Treatment and Recurrence. Nutrients 2019, 11, 1514. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zheng, R.; Baade, P.D.; Zhang, S.; Zeng, H.; Bray, F.; Jemal, A.; Yu, X.Q.; He, J. Cancer statistics in China, 2015. CA Cancer J. Clin. 2016, 66, 115–132. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Xu, P.; Zhang, D.; Liu, K.; Song, D.; Zheng, Y.; Yang, S.; Li, N.; Hao, Q.; Wu, Y.; et al. Association of folate intake and plasma folate level with the risk of breast cancer: A dose-response meta-analysis of observational studies. Aging 2020, 12, 21355–21375. [Google Scholar] [CrossRef]
- Zeng, J.; Gu, Y.; Fu, H.; Liu, C.; Zou, Y.; Chang, H. Association Between One-carbon Metabolism-related Vitamins and Risk of Breast Cancer: A Systematic Review and Meta-analysis of Prospective Studies. Clin. Breast Cancer 2020, 20, e469–e480. [Google Scholar] [CrossRef]
- Song, H.; Jeong, A.; Tran, T.X.M.; Lee, J.; Kim, M.; Park, B. Association between Micronutrient Intake and Breast Cancer Risk According to Body Mass Index in South Korean Adult Women: A Cohort Study. Nutrients 2022, 14, 2644. [Google Scholar] [CrossRef]
- Sun, Y.; Fowke, J.H.; Liang, X.; Mozhui, K.; Sen, S.; Bao, W.; Liu, B.; Snetselaar, L.G.; Wallace, R.B.; Shadyab, A.H.; et al. Changes in Dietary Intake of Methionine, Folate/Folic Acid and Vitamin B12 and Survival in Postmenopausal Women with Breast Cancer: A Prospective Cohort Study. Nutrients 2022, 14, 4747. [Google Scholar] [CrossRef]
- Hatami, M.; Vahid, F.; Akbari, M.E.; Sadeghi, M.; Ameri, F.; Eini-Zeinab, H.; Jamshidi-Naeini, Y.; Davoodi, S.H. The Vitamins Involved in One-Carbon Metabolisms are Associated with Reduced Risk of Breast Cancer in Overall and Subtypes. Int. J. Vitam. Nutr. Res. 2020, 90, 131–140. [Google Scholar] [CrossRef]
- Van Puyvelde, H.; Dimou, N.; Katsikari, A.; Indave Ruiz, B.I.; Godderis, L.; Huybrechts, I.; De Bacquer, D. The association between dietary intakes of methionine, choline and betaine and breast cancer risk: A systematic review and meta-analysis. Cancer Epidemiol. 2023, 83, 102322. [Google Scholar] [CrossRef]
- Wu, W.; Kang, S.; Zhang, D. Association of vitamin B6, vitamin B12 and methionine with risk of breast cancer: A dose-response meta-analysis. Br. J. Cancer 2013, 109, 1926–1944. [Google Scholar] [CrossRef]
- Ducker, G.S.; Rabinowitz, J.D. One-Carbon Metabolism in Health and Disease. Cell Metab. 2017, 25, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Mentch, S.J.; Locasale, J.W. One-carbon metabolism and epigenetics: Understanding the specificity. Ann. N. Y. Acad. Sci. 2016, 1363, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Koenig, K.L.; Scarmo, S.; Afanasyeva, Y.; Clendenen, T.V.; Ueland, P.M.; Zeleniuch-Jacquotte, A. Circulating unmetabolized folic acid and 5-methyltetrahydrofolate and risk of breast cancer: A nested case-control study. Eur. J. Clin. Nutr. 2020, 74, 1306–1315. [Google Scholar] [CrossRef]
- Wu, X.; Zou, T.; Cao, N.; Ni, J.; Xu, W.; Zhou, T.; Wang, X. Plasma homocysteine levels and genetic polymorphisms in folate metablism are associated with breast cancer risk in chinese women. Hered. Cancer Clin. Pract. 2014, 12, 2. [Google Scholar] [CrossRef] [PubMed]
- Houghton, S.C.; Eliassen, A.H.; Zhang, S.M.; Selhub, J.; Rosner, B.A.; Willett, W.C.; Hankinson, S.E. Plasma B-vitamins and one-carbon metabolites and the risk of breast cancer in younger women. Breast Cancer Res. Treat. 2019, 176, 191–203. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Yang, Z.; Sun, Y.; Qu, Z.; Jia, X.; Li, J.; Lin, Y.; Luo, Y. The Impact of Homocysteine on the Risk of Hormone-Related Cancers: A Mendelian Randomization Study. Front. Nutr. 2021, 8, 645371. [Google Scholar] [CrossRef]
- Houghton, S.C.; Eliassen, A.H.; Zhang, S.M.; Selhub, J.; Rosner, B.A.; Willett, W.C.; Hankinson, S.E. Plasma B-vitamin and one-carbon metabolites and risk of breast cancer before and after folic acid fortification in the United States. Int. J. Cancer 2019, 144, 1929–1940. [Google Scholar] [CrossRef]
- Lin, J.; Lee, I.M.; Song, Y.; Cook, N.R.; Selhub, J.; Manson, J.E.; Buring, J.E.; Zhang, S.M. Plasma homocysteine and cysteine and risk of breast cancer in women. Cancer Res. 2010, 70, 2397–2405. [Google Scholar] [CrossRef]
- Chou, Y.C.; Lee, M.S.; Wu, M.H.; Shih, H.L.; Yang, T.; Yu, C.P.; Yu, J.C.; Sun, C.A. Plasma homocysteine as a metabolic risk factor for breast cancer: Findings from a case-control study in Taiwan. Breast Cancer Res. Treat. 2007, 101, 199–205. [Google Scholar] [CrossRef]
- Zhang, S.M.; Willett, W.C.; Selhub, J.; Manson, J.E.; Colditz, G.A.; Hankinson, S.E. A prospective study of plasma total cysteine and risk of breast cancer. Cancer Epidemiol. Biomark. Prev. 2003, 12, 1188–1193. [Google Scholar]
- Chou, Y.C.; Wu, M.H.; Yu, J.C.; Lee, M.S.; Yang, T.; Shih, H.L.; Wu, T.Y.; Sun, C.A. Genetic polymorphisms of the methylenetetrahydrofolate reductase gene, plasma folate levels and breast cancer susceptibility: A case-control study in Taiwan. Carcinogenesis 2006, 27, 2295–2300. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.F.; Lin, F.Y.; Long, W.Q.; Luo, W.P.; Yan, B.; Xu, M.; Mo, X.F.; Zhang, C.X. Serum betaine but not choline is inversely associated with breast cancer risk: A case-control study in China. Eur. J. Nutr. 2017, 56, 1329–1337. [Google Scholar] [CrossRef] [PubMed]
- Severi, G.; Southey, M.C.; English, D.R.; Jung, C.H.; Lonie, A.; McLean, C.; Tsimiklis, H.; Hopper, J.L.; Giles, G.G.; Baglietto, L. Epigenome-wide methylation in DNA from peripheral blood as a marker of risk for breast cancer. Breast Cancer Res. Treat. 2014, 148, 665–673. [Google Scholar] [CrossRef]
- van Veldhoven, K.; Polidoro, S.; Baglietto, L.; Severi, G.; Sacerdote, C.; Panico, S.; Mattiello, A.; Palli, D.; Masala, G.; Krogh, V.; et al. Epigenome-wide association study reveals decreased average methylation levels years before breast cancer diagnosis. Clin. Epigenet. 2015, 7, 67. [Google Scholar] [CrossRef]
- Xu, X.; Gammon, M.D.; Jefferson, E.; Zhang, Y.; Cho, Y.H.; Wetmur, J.G.; Teitelbaum, S.L.; Bradshaw, P.T.; Terry, M.B.; Garbowski, G.; et al. The influence of one-carbon metabolism on gene promoter methylation in a population-based breast cancer study. Epigenetics 2011, 6, 1276–1283. [Google Scholar] [CrossRef] [PubMed]
- Song, F.; Ji, P.; Zheng, H.; Song, F.; Wang, Y.; Hao, X.; Wei, Q.; Zhang, W.; Chen, K. Definition of a functional single nucleotide polymorphism in the cell migration inhibitory gene MIIP that affects the risk of breast cancer. Cancer Res. 2010, 70, 1024–1032. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Liu, F.; Qiao, Y.; Wang, P.; Du, H.; Si, C.; Wang, X.; Chen, K.; Song, F. Genetically Modified Circulating Levels of Advanced Glycation End-Products and Their Soluble Receptor (AGEs-RAGE Axis) with Risk and Mortality of Breast Cancer. Cancers 2022, 14, 6124. [Google Scholar] [CrossRef] [PubMed]
- Butler, L.M.; Arning, E.; Wang, R.; Bottiglieri, T.; Govindarajan, S.; Gao, Y.T.; Yuan, J.M. Prediagnostic levels of serum one-carbon metabolites and risk of hepatocellular carcinoma. Cancer Epidemiol. Biomark. Prev. 2013, 22, 1884–1893. [Google Scholar] [CrossRef]
- Spratlen, M.J.; Grau-Perez, M.; Umans, J.G.; Yracheta, J.; Best, L.G.; Francesconi, K.; Goessler, W.; Bottiglieri, T.; Gamble, M.V.; Cole, S.A.; et al. Targeted metabolomics to understand the association between arsenic metabolism and diabetes-related outcomes: Preliminary evidence from the Strong Heart Family Study. Environ. Res. 2019, 168, 146–157. [Google Scholar] [CrossRef]
- Zhang, Y.; Long, H.; Wang, S.; Xiao, W.; Xiong, M.; Liu, J.; Chen, L.; Chen, R.; Wei, X.; Shu, Y.; et al. Genome-Wide DNA Methylation Pattern in Whole Blood Associated With Primary Intracerebral Hemorrhage. Front. Immunol. 2021, 12, 702244. [Google Scholar] [CrossRef]
- Morris, T.J.; Butcher, L.M.; Feber, A.; Teschendorff, A.E.; Chakravarthy, A.R.; Wojdacz, T.K.; Beck, S. ChAMP: 450k Chip Analysis Methylation Pipeline. Bioinformatics 2014, 30, 428–430. [Google Scholar] [CrossRef] [PubMed]
- Teschendorff, A.E.; Marabita, F.; Lechner, M.; Bartlett, T.; Tegner, J.; Gomez-Cabrero, D.; Beck, S. A beta-mixture quantile normalization method for correcting probe design bias in Illumina Infinium 450 k DNA methylation data. Bioinformatics 2013, 29, 189–196. [Google Scholar] [CrossRef]
- Johnson, W.E.; Li, C.; Rabinovic, A. Adjusting batch effects in microarray expression data using empirical Bayes methods. Biostatistics 2007, 8, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Xie, Z.; Pape, M.L.; Dye, T. An evaluation of statistical methods for DNA methylation microarray data analysis. BMC Bioinform. 2015, 16, 217. [Google Scholar] [CrossRef] [PubMed]
- Jaffe, A.E.; Murakami, P.; Lee, H.; Leek, J.T.; Fallin, M.D.; Feinberg, A.P.; Irizarry, R.A. Bump hunting to identify differentially methylated regions in epigenetic epidemiology studies. Int. J. Epidemiol. 2012, 41, 200–209. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. clusterProfiler: An R package for comparing biological themes among gene clusters. Omics 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Coppola, A.; Ilisso, C.P.; Stellavato, A.; Schiraldi, C.; Caraglia, M.; Mosca, L.; Cacciapuoti, G.; Porcelli, M. S-Adenosylmethionine Inhibits Cell Growth and Migration of Triple Negative Breast Cancer Cells through Upregulating MiRNA-34c and MiRNA-449a. Int. J. Mol. Sci. 2020, 22, 286. [Google Scholar] [CrossRef]
- Rezaee, M.; Akbari, H.; Momeni-Moghaddam, M.A.; Moazzen, F.; Salahi, S.; Jahankhah, R.; Tahmasebi, S. Association of C677T (rs1081133) and A1298C (rs1801131) Methylenetetrahydrofolate Reductase Variants with Breast Cancer Susceptibility Among Asians: A Systematic Review and Meta-Analysis. Biochem. Genet. 2021, 59, 367–397. [Google Scholar] [CrossRef]
- Esteller, M. Epigenetics in cancer. N. Engl. J. Med. 2008, 358, 1148–1159. [Google Scholar] [CrossRef]
- Chen, J.; Xu, X. Diet, epigenetic, and cancer prevention. Adv. Genet. 2010, 71, 237–255. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Ali, M.M. Methyl Donor Micronutrients that Modify DNA Methylation and Cancer Outcome. Nutrients 2019, 11, 608. [Google Scholar] [CrossRef] [PubMed]
- Raafat Elsayed, A.A.; Al-Marsoummi, S.; Vomhof-Dekrey, E.E.; Basson, M.D. SLFN12 Over-expression Sensitizes Triple Negative Breast Cancer Cells to Chemotherapy Drugs and Radiotherapy. Cancer Genom. Proteom. 2022, 19, 328–338. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Group | Case (N = 107) | Control (N = 107) | p |
|---|---|---|---|---|
| Age, mean ± SD, years | 53.3 ± 6.3 | 53.3 ± 5.8 | 0.991 | |
| Education, N (%) | Senior high school or above | 55 (55.0) | 66 (61.7) | 0.043 |
| Junior high school | 30 (30.0) | 36 (33.6) | ||
| Primary school and below | 15 (15.0) | 5 (4.7) | ||
| Per capita monthly household income, N (%), CNY | <500 | 4 (4.1) | 2 (1.9) | <0.001 |
| 500–999 | 15 (15.5) | 6 (5.7) | ||
| 1000–1999 | 47 (48.5) | 32 (30.2) | ||
| 2000–2999 | 20 (20.6) | 33 (31.1) | ||
| ≥3000 | 11 (11.3) | 33 (31.1) | ||
| Job, N (%) | No | 53 (54.6) | 55 (52.9) | 0.803 |
| Yes | 44 (45.4) | 49 (47.1) | ||
| BMI, N (%), kg/m2 | <23.9 | 35 (32.7) | 55 (51.9) | 0.001 |
| 24.0–27.9 | 46 (43.0) | 43 (40.6) | ||
| ≥28.0 | 26 (24.3) | 8 (7.5) | ||
| Benign breast disease, N (%) | No | 66 (65.3) | 83 (82.2) | 0.007 |
| Yes | 35 (34.7) | 18 (17.8) | ||
| Family history of cancer, N (%) | No | 66 (62.3) | 79 (75.2) | 0.042 |
| Yes | 40 (37.7) | 26 (24.8) | ||
| Family history of breast cancer, N (%) | No | 97 (94.2) | 104 (99.0) | 0.087 |
| Yes | 6 (5.8) | 1 (1.0) | ||
| Smoking, N (%) | No | 101 (97.1) | 97 (96.0) | 0.673 |
| Yes | 3 (2.9) | 4 (4.0) | ||
| Passive smoking, N (%) | No | 33 (41.3) | 31 (55.4) | 0.106 |
| Yes | 47 (58.8) | 25 (44.6) | ||
| Alcohol drinking, N (%) | No | 102 (98.1) | 100 (98.0) | 0.984 |
| Yes | 2 (1.9) | 2 (2.0) | ||
| Negative events, N (%) | No | 74 (75.5) | 89 (89.9) | 0.008 |
| Yes | 24 (24.5) | 10 (10.1) | ||
| Age of menarche, N (%), years | ≤13 | 27 (26.0) | 18 (17.1) | 0.115 |
| 14–15 | 41 (39.4) | 39 (37.1) | ||
| ≥16 | 36 (34.6) | 48 (45.7) | ||
| Number of pregnancies, N (%) | ≥3 | 53 (51.0) | 27 (25.7) | <0.001 |
| 2 | 38 (36.5) | 35 (33.3) | ||
| ≤1 | 13 (12.5) | 43 (41.0) | ||
| Age of first pregnancy, N (%), years | <30 | 87 (87.9) | 86 (85.1) | 0.572 |
| ≥30 | 12 (12.1) | 15 (14.9) | ||
| Number of live births, N (%) | ≥2 | 30 (29.1) | 13 (12.4) | 0.003 |
| ≤1 | 73 (70.9) | 92 (87.6) | ||
| Breastfeeding months, N (%), months | >12 | 63 (63.0) | 43 (43.0) | 0.005 |
| ≤12 | 37 (37.0) | 57 (57.0) | ||
| Abortion, N (%) | No | 18 (17.3) | 42 (41.2) | <0.001 |
| Yes | 86 (82.7) | 60 (58.8) | ||
| Take birth control pills, N (%) | No | 84 (84.8) | 84 (82.4) | 0.633 |
| Yes | 15 (15.2) | 18 (17.6) | ||
| Estrogen replacement therapy, N (%) | No | 87 (89.7) | 85 (94.4) | 0.232 |
| Yes | 10 (10.3) | 5 (5.6) | ||
| Sterilization surgery, N (%) | No | 93 (91.2) | 83 (91.2) | 0.994 |
| Yes | 9 (8.8) | 8 (8.8) | ||
| Menopausal status, N (%) | Premenopausal | 34 (32.1) | 33 (32.0) | 0.995 |
| Postmenopausal | 72 (67.9) | 70 (68.0) | ||
| Breast density, N (%) | Low | 72 (75.8) | 51 (63.0) | 0.064 |
| High | 23 (24.2) | 30 (37.0) |
| Group | Micronutrients | Median (P25, P75) | OR (95% CI) | ||
|---|---|---|---|---|---|
| Case (N = 107) | Control (N = 107) | Crude | Adjusted | ||
| Methionine cycle | Methionine, μmol/L | 18.06 (9.27,37.02) | 31.33 (15.50,55.15) | 0.62 (0.46–0.83) | 0.59 (0.39–0.92) |
| Cysteine, μmol/L | 6.95 (4.63,9.57) | 5.61 (4.53,8.45) | 1.44 (1.08–1.91) | 1.79 (1.08–2.97) | |
| Homocysteine, μmol/L | 4.91 (3.13,9.77) | 6.47 (3.38,16.25) | 0.75 (0.49–1.13) | 0.84 (0.51–1.40) | |
| SAM, μmol/L | 0.07 (0.04,0.13) | 0.20 (0.11,0.37) | 0.12 (0.06–0.26) | 0.07 (0.02–0.23) | |
| SAH, μmol/L | 0.04 (0.02,0.06) | 0.05 (0.04,0.07) | 0.70 (0.53–0.94) | 0.60 (0.39–0.92) | |
| Folate cycle | Folate, nmol/L | 8.75 (4.79,15.35) | 20.63 (8.37,36.57) | 0.32 (0.21–0.50) | 0.34 (0.18–0.70) |
| 5-MTHF, μmol/L | 0.24 (0.13,0.57) | 0.26 (0.16,0.47) | 1.09 (0.83–1.42) | 1.09 (0.72–1.65) | |
| Enzymatic factor | P5P, nmol/L | 9.94 (4.39,16.07) | 28.55 (15.83,42.29) | 0.10 (0.05–0.19) | 0.02 (0.004–0.17) |
| Vitamin B2, nmol/L | 2.45 (1.39,3.85) | 6.87 (3.67,11.34) | 0.13 (0.07–0.24) | 0.09 (0.03–0.28) | |
| Vitamin B12, nmol/L | 1.23 (0.69,1.97) | 3.76 (2.16,6.84) | 0.02 (0.01–0.07) | 0.01 (0.001–0.06) | |
| Choline metabolism | Choline, μmol/L | 6.58 (4.62,10.64) | 11.84 (6.15,20.17) | 0.33 (0.20–0.53) | 0.43 (0.25–0.73) |
| Betaine, μmol/L | 15.26 (9.63,22.47) | 23.83 (13.55,37.73) | 0.42 (0.28–0.62) | 0.36 (0.17–0.75) | |
| Group | Micronutrients | Median (P25, P75) | OR (95% CI) | ||
|---|---|---|---|---|---|
| Case | Control | Crude | Adjusted | ||
| Premenopausal (N = 67) | |||||
| Methionine cycle | Methionine, μmol/L | 18.62 (9.43,51.90) | 41.85 (19.70,59.58) | 0.62 (0.37–1.03) | 0.07 (0.004–0.99) |
| Cysteine, μmol/L | 7.44 (4.62–9.80) | 4.96 (4.48–6.91) | 2.30 (1.20–4.40) | 0.98 (0.39–2.46) | |
| Homocysteine, μmol/L | 5.59 (3.09–18.97) | 11.62 (4.66–18.97) | 0.71 (0.42–1.18) | 0.46 (0.08–2.55) | |
| SAM, μmol/L | 0.09 (0.03–0.16) | 0.17 (0.12–0.26) | 0.16 (0.04–0.70) | 0.19 (0.02–1.52) | |
| SAH, μmol/L | 0.04 (0.02–0.05) | 0.05 (0.04–0.07) | 0.68 (0.41–1.12) | 0.37 (0.08–1.72) | |
| Folate cycle | Folate, nmol/L | 7.86 (4.21–13.51) | 11.80 (6.96–34.68) | 0.36 (0.17–0.78) | 0.21 (0.01–4.50) |
| 5-MTHF, μmol/L | 0.20 (0.13–0.62) | 0.24 (0.15–0.34) | 1.42 (0.84–2.39) | 0.32 (0.07–1.41) | |
| Enzymatic factor | P5P, nmol/L | 11.61 (4.80–18.32) | 32.35 (16.66–50.75) | 0.17 (0.06–0.43) | 0.01 (0.00–0.82) |
| Vitamin B2, nmol/L | 2.81 (1.19–3.89) | 6.18 (3.31–8.49) | 0.18 (0.07–0.47) | 0.01 (0.00–0.83) | |
| Vitamin B12, nmol/L | 1.31 (0.73–2.01) | 3.89 (2.45–8.00) | 0.01 (0.00–0.10) | 0.00 (0.00–6.33) | |
| Choline metabolism | Choline, μmol/L | 6.09 (4.03–8.76) | 11.02 (5.98–23.63) | 0.18 (0.05–0.63) | 0.29 (0.05–1.71) |
| Betaine, μmol/L | 14.64 (9.74–23.77) | 23.64 (16.75–37.16) | 0.39 (0.19–0.81) | 0.01 (0.00–1.44) | |
| Postmenopausal (N = 143) | |||||
| Methionine cycle | Methionine, μmol/L | 16.27 (9.14–34.30) | 30.56 (15.37–51.28) | 0.60 (0.42–0.86) | 0.78 (0.38–1.59) |
| Cysteine, μmol/L | 6.95 (4.63–9.56) | 6.06 (4.59–8.68) | 1.21 (0.87–1.69) | 1.97 (0.84–4.66) | |
| Homocysteine, μmol/L | 4.91 (3.13–8.84) | 4.95 (2.67–13.62) | 0.86 (0.61–1.22) | 1.03 (0.53–1.98) | |
| SAM, μmol/L | 0.06 (0.04–0.11) | 0.23 (0.11–0.47) | 0.06 (0.02–0.19) | 0.02 (0.002–0.21) | |
| SAH, μmol/L | 0.04 (0.02–0.06) | 0.05 (0.04–0.07) | 0.73 (0.52–1.04) | 0.72 (0.34–1.51) | |
| Folate cycle | Folate, nmol/L | 10.44 (4.98–16.50) | 20.88 (10.98–40.04) | 0.30 (0.17–0.53) | 0.34 (0.13–0.92) |
| 5-MTHF, μmol/L | 0.28 (0.13–0.46) | 0.26 (0.16–0.48) | 1.04 (0.74–1.45) | 1.42 (0.69–2.91) | |
| Enzymatic factor | P5P, nmol/L | 8.98 (4.26–15.07) | 25.49 (14.23–40.81) | 0.08 (0.03–0.20) | 0.02 (0.001–0.20) |
| Vitamin B2, nmol/L | 2.48 (1.42–3.94) | 7.27 (4.15–12.65) | 0.12 (0.05–0.26) | 0.09 (0.02–0.53) | |
| Vitamin B12, nmol/L | 1.18 (0.61–1.97) | 3.29 (1.94–5.64) | 0.09 (0.04–0.22) | 0.01 (0.00–0.21) | |
| Choline metabolism | Choline, μmol/L | 7.39 (4.62–11.32) | 11.73 (6.15–18.78) | 0.54 (0.35–0.83) | 0.36 (0.14–0.90) |
| Betaine, μmol/L | 15.60 (9.54–21.02) | 23.85 (12.05–36.48) | 0.53 (0.36–0.79) | 0.58 (0.20–1.64) | |
| CpG | Δβ | Chr | CpG Position | CpG Related Traits | Gene | Gene Position | Gene Molecular Function | Gene Pathways |
|---|---|---|---|---|---|---|---|---|
| cg05397629 | −0.597 | 11 | opensea | / | RHOG | Body | Nucleotide binding; GTPase activity | PI5P, PP2A, and IER3 regulate PI3K/AKT Signaling; signaling by Rho GTPases |
| cg02589685 | −0.451 | 11 | shore | / | / | IGR | / | / |
| cg03363289 | −0.257 | 9 | island | Breast cancer prognosis; colorectal cancer | LHX6 | Body | DNA binding | / |
| cg04622888 | −0.239 | 9 | island | Gingivo-buccal oral squamous cell carcinoma; breast cancer prognosis; colorectal cancer | LHX6 | TSS200 | DNA binding | / |
| cg09491380 | −0.234 | 4 | opensea | / | MAEA | Body | Actin binding; ubiquitin protein transferase activity | Ciliary landscape |
| cg10617037 | −0.229 | 8 | shore | / | CYHR1 | 1stExon | Zinc ion binding | / |
| cg15740243 | −0.229 | 5 | shore | / | RNF145 | TSS1500 | Zinc ion binding; transferase activity | / |
| cg00901687 | −0.218 | 17 | shore | Aging; Alzheimer’s disease (AD) | MYCBPAP | TSS1500 | Protein binding; phospholipid binding; clathrin binding | / |
| cg24312537 | −0.215 | 8 | shore | Crohn’s disease (CD); gestational diabetes mellitus; mortality | HTRA4 | TSS1500 | Serine-type endopeptidase activity; protein binding | / |
| cg06536614 | −0.215 | 5 | island | Breast cancer risk | MIR886 | TSS200 | / | / |
| cg04657470 | −0.213 | 2 | island | Tetralogy of Fallot | HSPE1 | 1stExon | RNA binding; protein binding; ATP binding | Signaling by Rho GTPases; RAC2 GTPase cycle |
| cg04794268 | −0.208 | 8 | shore | Early metastasis of uveal melanoma | KIFC2 | TSS1500 | Nucleotide binding; cytoskeletal motor activity | Golgi-to-ER retrograde transport; vesicle-mediated transport |
| cg21042336 | −0.204 | 18 | shore | / | OSBPL1A | TSS1500 | Protein binding; phospholipid binding; lipid binding | Synthesis of bile acids and bile salts; metabolism |
| cg21697381 | −0.203 | 17 | opensea | B acute lymphoblastic leukemia | SLFN12 | TSS1500 | RNA nuclease activity; protein binding | 17q12 copy number variation syndrome |
| cg01701207 | −0.203 | 16 | opensea | Allergic asthma | SF3B3 | Body | Nucleic acid binding; protein binding | Processing of Capped Intron-Containing Pre-mRNA |
| cg25755428 | 0.229 | 19 | island | / | MRI1 | TSS1500 | Protein binding; isomerase activity | Methionine de novo and salvage pathway; sulfur amino acid metabolism |
| cg13824270 | 0.270 | 6 | shore | Breast cancer | PRPF4B | TSS1500 | Nucleotide binding; RNA binding; protein kinase activity | Processing of Capped Intron-Containing Pre-mRNA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, F.; Zhou, H.; Peng, Y.; Qiao, Y.; Wang, P.; Si, C.; Wang, X.; Gong, J.; Chen, K.; Song, F. Plasma One-Carbon Metabolism-Related Micronutrients and the Risk of Breast Cancer: Involvement of DNA Methylation. Nutrients 2023, 15, 3621. https://doi.org/10.3390/nu15163621
Liu F, Zhou H, Peng Y, Qiao Y, Wang P, Si C, Wang X, Gong J, Chen K, Song F. Plasma One-Carbon Metabolism-Related Micronutrients and the Risk of Breast Cancer: Involvement of DNA Methylation. Nutrients. 2023; 15(16):3621. https://doi.org/10.3390/nu15163621
Chicago/Turabian StyleLiu, Fubin, Huijun Zhou, Yu Peng, Yating Qiao, Peng Wang, Changyu Si, Xixuan Wang, Jianxiao Gong, Kexin Chen, and Fangfang Song. 2023. "Plasma One-Carbon Metabolism-Related Micronutrients and the Risk of Breast Cancer: Involvement of DNA Methylation" Nutrients 15, no. 16: 3621. https://doi.org/10.3390/nu15163621
APA StyleLiu, F., Zhou, H., Peng, Y., Qiao, Y., Wang, P., Si, C., Wang, X., Gong, J., Chen, K., & Song, F. (2023). Plasma One-Carbon Metabolism-Related Micronutrients and the Risk of Breast Cancer: Involvement of DNA Methylation. Nutrients, 15(16), 3621. https://doi.org/10.3390/nu15163621