
Menopause Hot Flashes and Molecular Mechanisms Modulated by Food-Derived Nutrients


Abstract
1. Introduction
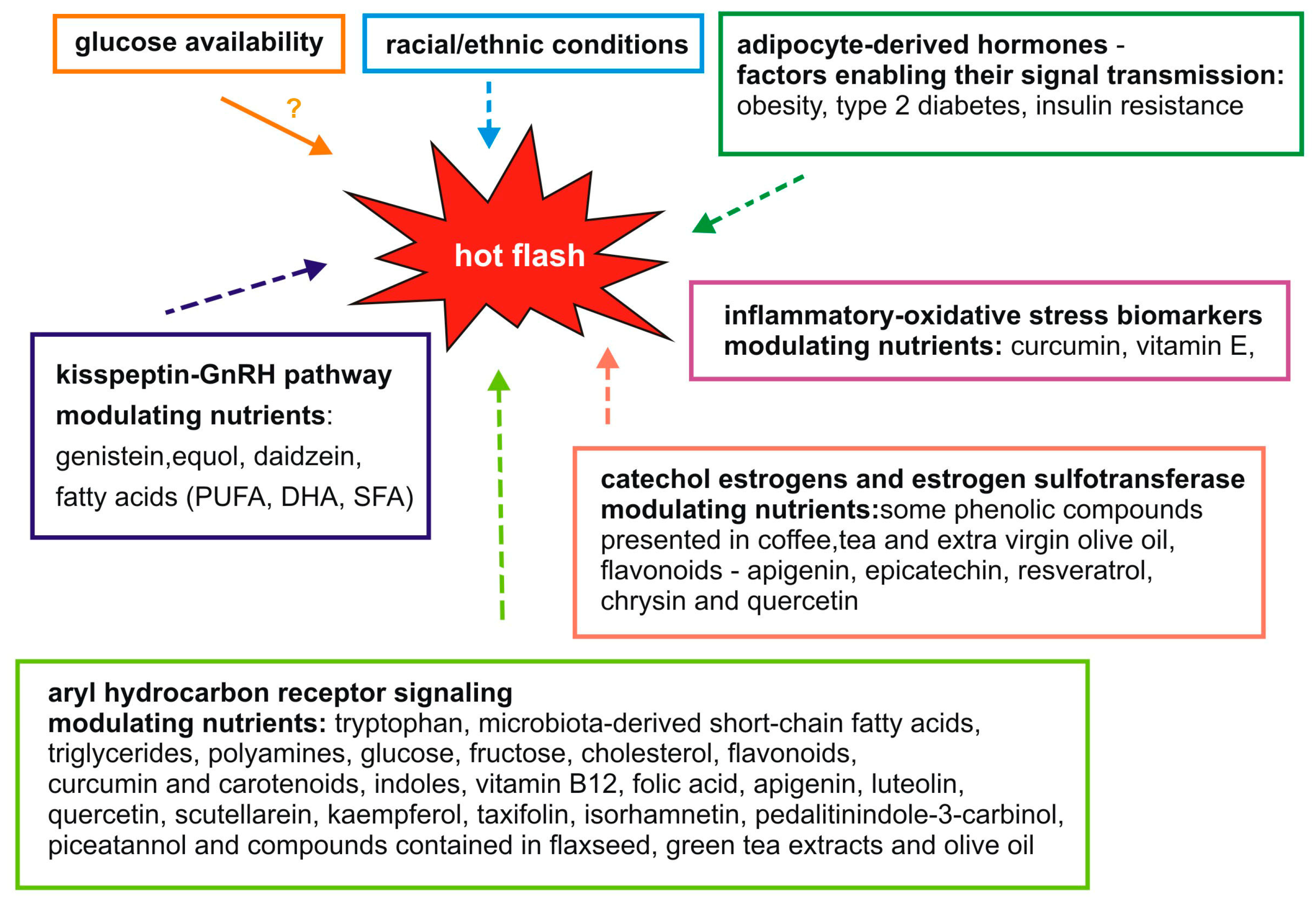
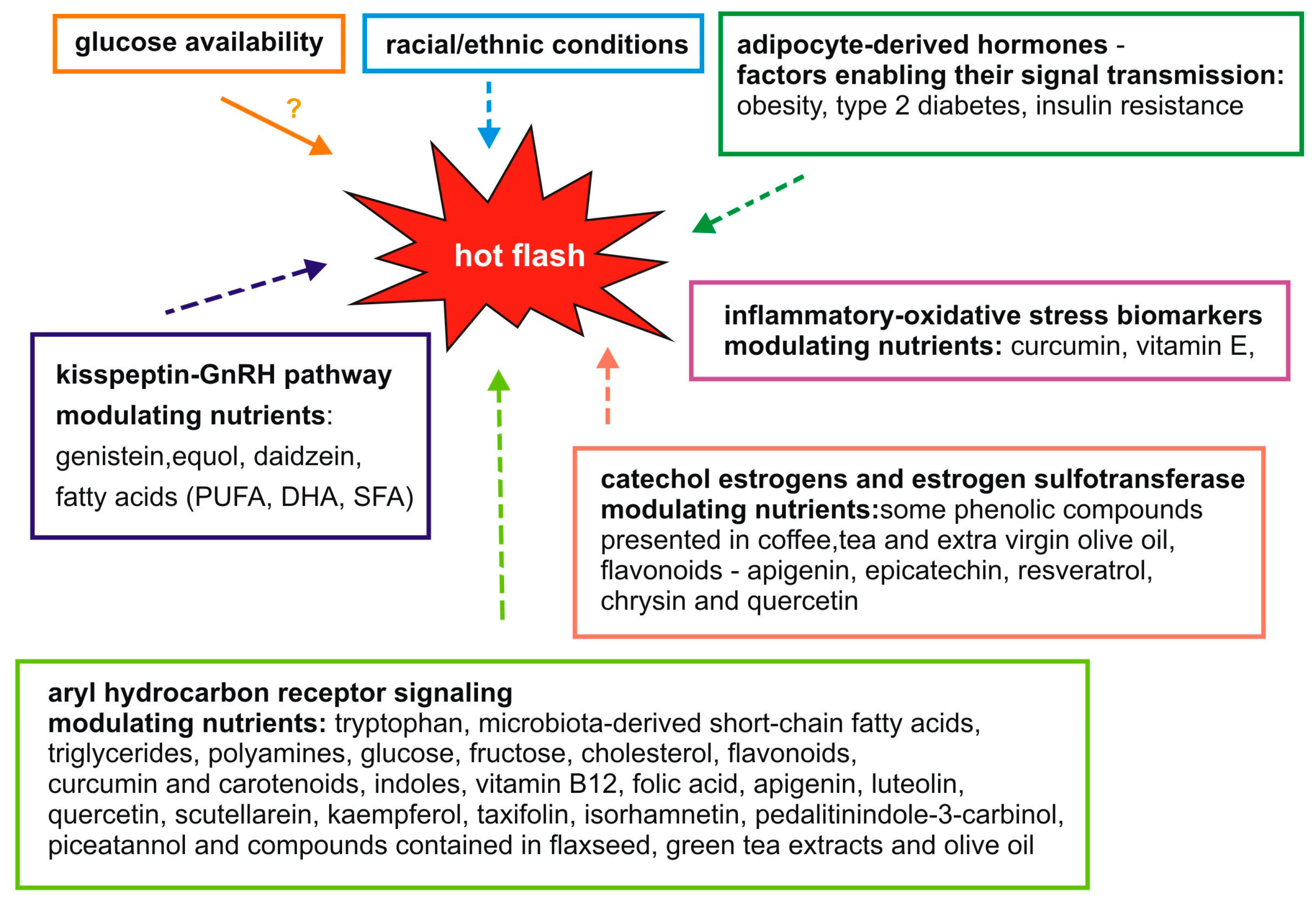
2. Molecular Factors Associated with Hot Flashes and Nutrients
3. The Kisspeptin-GnRH Pathway
4. Adipocyte-Derived Hormones
5. Aryl Hydrocarbon Receptor Signaling
6. Catechol Estrogens and Estrogen Sulfotransferase
7. Inflammatory and Oxidative Stress Biomarkers
8. Glucose Availability
9. Racial/Ethnic Conditions
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Philp, H.A. Hot flashes—A review of the literature on alternative and complementary treatment approaches. Altern. Med. Rev. 2003, 8, 284–302. [Google Scholar]
- Morrow, P.K.H.; Mattair, D.N.; Hortobagyi, G.N. Hot Flashes: A Review of Pathophysiology and Treatment Modalities. Oncologist 2011, 16, 1658–1664. [Google Scholar] [CrossRef]
- Talaulikar, V. Menopause transition: Physiology and symptoms. Best Pract. Res. Clin. Obstet. Gynaecol. 2022, 81, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Yelland, S.; Steenson, S.; Creedon, A.; Stanner, S. The role of diet in managing menopausal symptoms: A narrative review. Nutr. Bull. 2023, 48, 43–65. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Wang, B.; Hua, Q.; Jin, Q.; Xie, J.; Ma, J.; Jin, F. Investigation of the relationship between hot flashes, sweating and sleep quality in perimenopausal and postmenopausal women: The mediating effect of anxiety and depression. BMC Women’s Health 2021, 21, 293. [Google Scholar] [CrossRef] [PubMed]
- Freedman, R.R. Pathophysiology and treatment of menopausal hot flashes. Semin. Reprod. Med. 2005, 23, 117–125. [Google Scholar] [CrossRef]
- Thurston, R.C.; Maki, P.M.; Derby, C.A.; Sejdić, E.; Aizenstein, H.J. Menopausal hot flashes and the default mode network. Fertil. Steril. 2015, 103, 1572–1578.e1. [Google Scholar] [CrossRef] [PubMed]
- Gerber, L.M.; Sievert, L.L.; Schwartz, J.E. Hot flashes and midlife symptoms in relation to levels of salivary cortisol. Maturitas 2017, 96, 26–32. [Google Scholar] [CrossRef]
- Freedman, R.R. Menopausal hot flashes: Mechanisms, endocrinology, treatment. J. Steroid Biochem. Mol. Biol. 2014, 142, 115–120. [Google Scholar] [CrossRef]
- Bansal, R.; Aggarwal, N. Menopausal hot flashes: A concise review. J. Midlife Health 2019, 10, 6–13. [Google Scholar] [CrossRef]
- Sandru, F.; Dumitrascu, M.C.; Petrova, E.; Dumitrascu, A.; Tupea, C.; Carsote, M.; Ghemigian, A.; Valea, A. Hot flashes: Why? Rom. Med. J. 2021, 68, 5–9. [Google Scholar] [CrossRef]
- Saú, H.P.F.; Schmitt, A.C.B.; Cardoso, M.R.A.; Aldrighi, J.M. Prevalence of hot flashes in women of 40 to 65 years of age with metabolic syndrome. Rev. Assoc. Med. Bras. 2021, 67, 1515. [Google Scholar] [CrossRef] [PubMed]
- Patel, B.; Dhillo, W.S. Menopause review: Emerging treatments for menopausal symptoms. Best Pract. Res. Clin. Obstet. Gynaecol. 2022, 81, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, H.; Manson, J.A.E. Menopausal symptom management in women with cardiovascular disease or vascular risk factors. Maturitas 2022, 161, 1–6. [Google Scholar] [CrossRef]
- Crandall, C.J.; Diamant, A.L.; Maglione, M.; Thurston, R.C.; Sinsheimer, J. Genetic variation and hot flashes: A systematic review. J. Clin. Endocrinol. Metab. 2020, 105, e4907–e4957. [Google Scholar] [CrossRef] [PubMed]
- Zeisel, S.H. A conceptual framework for studying and investing in precision nutrition. Front. Genet. 2019, 10, 200. [Google Scholar] [CrossRef]
- Santoro, N.; Roeca, C.; Peters, B.A.; Neal-Perry, G. The Menopause Transition: Signs, Symptoms, and Management Options. J. Clin. Endocrinol. Metab. 2021, 106, 1–15. [Google Scholar] [CrossRef]
- Xiong, J.; Wang, M.; Tian, Y.; Ling, A.; Shan, S.; Zhao, L.; Cheng, G. (±)-Equol interferes with the secretion of gonadotropin-releasing hormone and the expression of neuromodulators in murine neurons. Toxicol. Environ. Chem. 2022, 104, 129–140. [Google Scholar] [CrossRef]
- Xiong, J.; Tian, Y.; Ling, A.; Liu, Z.; Zhao, L.; Cheng, G. Genistein affects gonadotrophin-releasing hormone secretion in GT1-7 cells via modulating kisspeptin receptor and key regulators. Syst. Biol. Reprod. Med. 2022, 68, 138–150. [Google Scholar] [CrossRef] [PubMed]
- Tran, A.; Loganathan, N.; McIlwraith, E.K.; Belsham, D.D. Palmitate and Nitric Oxide Regulate the Expression of Spexin and Galanin Receptors 2 and 3 in Hypothalamic Neurons: Nitric oxide and palmitate regulate Spx, Galr2 and Galr3 mRNA. Neuroscience 2020, 447, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Harlow, K.L.; Griesgraber, M.J.; Seman, A.D.; Shuping, S.L.; Sommer, J.R.; Griffith, E.H.; Hileman, S.M.; Nestor, C.C. The impact of undernutrition on KNDy (kisspeptin/neurokinin B/dynorphin) neurons in female lambs. J. Neuroendocrinol. 2022, 34, e13135. [Google Scholar] [CrossRef]
- Giordano, D.; Facchiano, A.; Moccia, S.; Meola, A.M.I.; Russo, G.L.; Spagnuolo, C. Molecular Docking of Natural Compounds for Potential Inhibition of AhR. Foods 2023, 12, 1953. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.H.; Chou, C.C.; Lee, Y.H.; Hung, C.C. Curcumin Facilitates Aryl Hydrocarbon Receptor Activation to Ameliorate Inflammatory Astrogliosis. Molecules 2022, 27, 2507. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Jeong, H.; Hur, S.; Lee, J.; Kwon, O. Efficacy and safety of kudzu flower–mandarin peel on hot flashes and bone markers in women during the menopausal transition: A randomized controlled trial. Nutrients 2020, 12, 3237. [Google Scholar] [CrossRef] [PubMed]
- Dahlem, C.; Kado, S.Y.; He, Y.; Bein, K.; Wu, D.; Haarmann-Stemmann, T.; Kado, N.Y.; Vogel, C.F.A. Ahr signaling interacting with nutritional factors regulating the expression of markers in vascular inflammation and atherogenesis. Int. J. Mol. Sci. 2020, 21, 8287. [Google Scholar] [CrossRef]
- Hubbard, T.D.; Murray, I.A.; Perdew, G.H. Special section on drug metabolism and the microbiome—Minireview indole and tryptophan metabolism: Endogenous and dietary routes to ah receptor activation. Drug Metab. Dispos. 2015, 43, 1522–1535. [Google Scholar] [CrossRef]
- Wang, L.; Tran, A.; Lee, J.; Belsham, D.D. Palmitate differentially regulates Spexin, and its receptors Galr2 and Galr3, in GnRH neurons through mechanisms involving PKC, MAPKs, and TLR4. Mol. Cell. Endocrinol. 2020, 518, 110991. [Google Scholar] [CrossRef]
- Islam, J.; Sato, S.; Watanabe, K.; Watanabe, T.; Ardiansyah; Hirahara, K.; Aoyama, Y.; Tomita, S.; Aso, H.; Komai, M.; et al. Dietary tryptophan alleviates dextran sodium sulfate-induced colitis through aryl hydrocarbon receptor in mice. J. Nutr. Biochem. 2017, 42, 43–50. [Google Scholar] [CrossRef]
- Yi, M.; Negishi, M.; Lee, S.J. Estrogen sulfotransferase (SULT1E1): Its molecular regulation, polymorphisms, and clinical perspectives. J. Pers. Med. 2021, 11, 194. [Google Scholar] [CrossRef]
- Yousefi-Nodeh, H.; Farshbaf-Khalili, A.; Sadeghzadeh Oskouei, B.; Jafarilar-Aghdam, N.; Kazemi-Zanjani, N.; Pourzeinali, S. Curcumin and vitamin E improve hot flashes, lipid profile, and fasting blood glucose without any detrimental effect on the liver and renal function in postmenopausal women: A triple-blind placebo-controlled clinical trial. Health Care Women Int. 2022, 2, 1–23. [Google Scholar] [CrossRef]
- Farshbaf-Khalili, A.; Ostadrahimi, A.; Mirghafourvand, M.; Ataei-Almanghadim, K.; Dousti, S.; Iranshahi, A.M. Clinical Efficacy of Curcumin and Vitamin e on Inflammatory-Oxidative Stress Biomarkers and Primary Symptoms of Menopause in Healthy Postmenopausal Women: A Triple-Blind Randomized Controlled Trial. J. Nutr. Metab. 2022, 2022, 6339715. [Google Scholar] [CrossRef]
- Skorupskaite, K.; George, J.T.; Anderson, R.A. The kisspeptin-GnRH pathway in human reproductive health and disease. Hum. Reprod. Update 2014, 20, 485–500. [Google Scholar] [CrossRef]
- Zhao, W.; Smith, J.A.; Yu, M.; Crandall, C.J.; Thurston, R.C.; Hood, M.M.; Ruiz-Narvaez, E.; Peyser, P.A.; Kardia, S.L.R.; Harlow, S.D. Genetic variants predictive of reproductive aging are associated with vasomotor symptoms in a multiracial/ethnic cohort. Menopause 2021, 28, 883–892. [Google Scholar] [CrossRef]
- Weems, P.W.; Coolen, L.M.; Hileman, S.M.; Hardy, S.; McCosh, R.B.; Goodman, R.L.; Lehman, M.N. Evidence that dynorphin acts upon KNDY and GnRH neurons during GnRH pulse termination in the ewe. Endocrinology 2018, 159, 3187–3199. [Google Scholar] [CrossRef]
- Marques, P.; Skorupskaite, K.; Rozario, K.S.; Anderson, R.A.; George, J.T. Physiology of GNRH and gonadotropin secretion. In Endotext [Internet]; MDText.com, Inc.: South Dartmouth, MA, USA, 2022. [Google Scholar]
- Mayo, B.; Vázquez, L.; Flórez, A.B. Equol: A bacterial metabolite from the Daidzein isoflavone and its presumed beneficial health effects. Nutrients 2019, 11, 2231. [Google Scholar] [CrossRef]
- Gong, Y.; Lv, J.; Pang, X.; Zhang, S.; Zhang, G.; Liu, L.; Wang, Y.; Li, C. Advances in the Metabolic Mechanism and Functional Characteristics of Equol. Foods 2023, 12, 2334. [Google Scholar] [CrossRef]
- Obradovic, M.; Sudar-Milovanovic, E.; Soskic, S.; Essack, M.; Arya, S.; Stewart, A.J.; Gojobori, T.; Isenovic, E.R. Leptin and Obesity: Role and Clinical Implication. Front. Endocrinol. 2021, 12, 585887. [Google Scholar] [CrossRef]
- Karim, R.; Dang, H.M.; Hodis, H.N.; Stanczyk, F.Z.; Brinton, R.D.; Mack, W.J. Association of hot flushes with ghrelin and adipokines in early versus late postmenopausal women. Menopause 2020, 27, 512–518. [Google Scholar] [CrossRef] [PubMed]
- Kazama, M.; Terauchi, M.; Odai, T.; Kato, K.; Miyasaka, N. Associations of fat mass index with hot flashes and lean mass index with insomnia in middle-aged women. Climacteric 2023, 26, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Li, X.; Wang, Y.; Cao, Y.; Yao, D.; Sun, L.; Qin, L.; Qiu, H.; Zhan, X. Adipocyte-derived extracellular vesicles modulate appetite and weight through mTOR signalling in the hypothalamus. Acta Physiol. 2020, 228, e13339. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Zhou, Y.; Jin, B.; Shu, J. Role of estrogen in the regulation of central and peripheral energy homeostasis: From a menopausal perspective. Ther. Adv. Endocrinol. Metab. 2023, 14, 20420188231199359. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.Y.; Hsin, I.L.; Chen, D.R.; Chang, C.C.; Kor, C.T.; Chen, T.Y.; Wu, H.M. Circulating interleukin-8 and tumor necrosis factor-α are associated with hot flashes in healthy postmenopausal women. PLoS ONE 2017, 12, e0184011. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Wang, H.; Zhang, X. Genetic aspects of early menopause. J. Bio-X Res. 2019, 2, 105–111. [Google Scholar] [CrossRef]
- Bungsu, I.; Kifli, N.; Ahmad, S.R.; Ghani, H.; Cunningham, A.C. Herbal Plants: The Role of AhR in Mediating Immunomodulation. Front. Immunol. 2021, 12, 697663. [Google Scholar] [CrossRef] [PubMed]
- Gong, P.; Madak-Erdogan, Z.; Flaws, J.A.; Shapiro, D.J.; Katzenellenbogen, J.A.; Katzenellenbogen, B.S. Estrogen receptor-α and aryl hydrocarbon receptor involvement in the actions of botanical estrogens in target cells. Mol. Cell. Endocrinol. 2016, 437, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Göttel, M.; Le Corre, L.; Dumont, C.; Schrenk, D.; Chagnon, M.C. Estrogen receptor α and aryl hydrocarbon receptor cross-talk in a transfected hepatoma cell line (HepG2) exposed to 2,3,7,8-tetrachlorodibenzo-p-dioxin. Toxicol. Rep. 2014, 1, 1029–1036. [Google Scholar] [CrossRef]
- Larigot, L.; Juricek, L.; Dairou, J.; Coumoul, X. AhR signaling pathways and regulatory functions. Biochim. Open 2018, 7, 1–9. [Google Scholar] [CrossRef]
- Ghiboub, M.; Verburgt, C.M.; Sovran, B.; Benninga, M.A.; de Jonge, W.J.; Van Limbergen, J.E. Nutritional therapy to modulate tryptophan metabolism and aryl hydrocarbon-receptor signaling activation in human diseases. Nutrients 2020, 12, 2846. [Google Scholar] [CrossRef]
- Sun, M.; Ma, N.; He, T.; Johnston, L.J.; Ma, X. Tryptophan (Trp) modulates gut homeostasis via aryl hydrocarbon receptor (AhR). Crit. Rev. Food Sci. Nutr. 2020, 60, 1760–1768. [Google Scholar] [CrossRef]
- De Juan, A.; Segura, E. Modulation of Immune Responses by Nutritional Ligands of Aryl Hydrocarbon Receptor. Front. Immunol. 2021, 12, 645168. [Google Scholar] [CrossRef]
- Matthews, J.; Gustafsson, J.-Å. Estrogen receptor and aryl hydrocarbon receptor signaling pathways. Nucl. Recept. Signal. 2006, 4, e016. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.J.; Venkataraman, A.; Jain, P.C.; Wiesler, E.P.; DeBlasio, M.; Klein, J.; Tu, S.S.; Lee, S.; Medzhitov, R.; Iwasaki, A. Vitamin B12 and folic acid alleviate symptoms of nutritional deficiency by antagonizing aryl hydrocarbon receptor. Proc. Natl. Acad. Sci. USA 2020, 117, 15837–15845. [Google Scholar] [CrossRef]
- Shah, B.R.; Xu, W.; Mraz, J. Cytochrome P450 1B1: Role in health and disease and effect of nutrition on its expression. RSC Adv. 2019, 9, 21050–21062. [Google Scholar] [CrossRef] [PubMed]
- Odoemelam, C.S.; Hunter, E.; Ahmad, Z.; Kamerlin, C.L.; White, S.; Wilson, P.B. Computational Investigation of Ligand Binding of Flavonoids in Cytochrome P450 Receptors. Curr. Pharm. Des. 2022, 28, 3637–3648. [Google Scholar] [CrossRef]
- Barbosa, A.C.S.; Feng, Y.; Yu, C.; Huang, M.; Xie, W. Estrogen sulfotransferase in the metabolism of estrogenic drugs and in the pathogenesis of diseases. Expert Opin. Drug Metab. Toxicol. 2019, 15, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Cox, F.F.; Misiou, A.; Vierkant, A.; Ale-Agha, N.; Grandoch, M.; Haendeler, J.; Altschmied, J. Protective Effects of Curcumin in Cardiovascular Diseases—Impact on Oxidative Stress and Mitochondria. Cells 2022, 11, 342. [Google Scholar] [CrossRef]
- Miyazawa, T.; Burdeos, G.C.; Itaya, M.; Nakagawa, K.; Miyazawa, T. Vitamin E: Regulatory Redox Interactions. IUBMB Life 2019, 71, 430–441. [Google Scholar] [CrossRef]
- Dormire, S.L. The potential role of glucose transport changes in hot flash physiology: A hypothesis. Biol. Res. Nurs. 2009, 10, 241–247. [Google Scholar] [CrossRef]
- Prakapenka, A.V.; Korol, D.L. Estradiol selectively regulates metabolic substrates across memory systems in models of menopause. Climacteric 2021, 24, 366–372. [Google Scholar] [CrossRef]
- Gregorio, K.C.R.; Laurindo, C.P.; Machado, U.F. Estrogen and Glycemic Homeostasis: The Fundamental Role of Nuclear Estrogen Receptors ESR1/ESR2 in Glucose Transporter GLUT4 Regulation. Cells 2021, 10, 99. [Google Scholar] [CrossRef]
- Mahboobifard, F.; Pourgholami, M.H.; Jorjani, M.; Dargahi, L.; Amiri, M.; Sadeghi, S.; Tehrani, F.R. Estrogen as a key regulator of energy homeostasis and metabolic health. Biomed. Pharmacother. 2022, 156, 113808. [Google Scholar] [CrossRef] [PubMed]
- Duarte, A.I.; Santos, M.S.; Oliveira, C.R.; Moreira, P.I. Brain Insulin Signalling, Glucose Metabolism and Females’ Reproductive Aging: A Dangerous Triad in Alzheimer’s Disease; Elsevier Ltd.: Amsterdam, The Netherlands, 2018; Volume 136, ISBN 3512398227. [Google Scholar]
- Aronica, L.; Ordovas, J.M.; Volkov, A.; Lamb, J.J.; Stone, P.M.; Minich, D.; Leary, M.; Class, M.; Metti, D.; Larson, I.A.; et al. Genetic Biomarkers of Metabolic Detoxification for Personalized Lifestyle Medicine. Nutrients 2022, 14, 768. [Google Scholar] [CrossRef] [PubMed]
- Visioli, F.; Marangoni, F.; Poli, A.; Ghiselli, A.; Martini, D. Nutrition and health or nutrients and health? Int. J. Food Sci. Nutr. 2022, 73, 141–148. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Compound Name, Concentration Used | Model | Property | Chemical Structure, IUPAC Name | Reference |
|---|---|---|---|---|
| kisspeptin-GnRH (gonadotropin-releasing hormone) pathway-modulating nutrients | ||||
| (±)-equol 0–20 µmol/L | GT1-7 cells (mouse hypothalamic GnRH neuronal cell line) | increase in gonadotropin-releasing hormone secretion, and disruption of neurokinin B receptor expression |  (3S)-isoflavan-4,7′-diol | [18] |
| genistein 0–80 µM for 48 h | GT1-7 cells (mouse hypothalamic GnRH neuronal cell line) | increase in GnRH secretion | 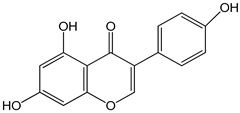 (5,7-dihydroxy-3-(4-hydroxyphenyl)chromen-4-one) | [19] |
| DHA (docosahexaenoic acid) 100 µM | mHypoA-59 cell line (primary hypothalamic culture isolated from 2-month old NPY-GFP mice) | upregulation of GnRH mRNA expression | 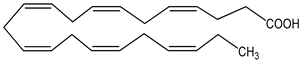 (docosahexaenoic acid; cis-4,7,10,13,16,19-docosahexaenoic acid) | [20] |
| PUFA (polyunsaturated fatty acid) 100 µM | mHypoA-59 cell line (primary hypothalamic culture isolated from 2-month old NPY-GFP mice) | upregulation of GnRH mRNA expression | linoleic acid as representative (cis, cis-9,12-octadecadienoic acid) | [20] |
| SFA (saturated fatty acid) 100 µM | mHypoA-59 cell line (primary hypothalamic culture isolated from 2-month old NPY-GFP mice) | upregulation of GnRH mRNA expression | palmitic acid as epresentative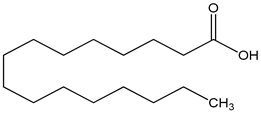 (hexadecanoic acid) | [20] |
| undernutrition feed-restricted to lose 20% of pre-study body weight over 13 weeks | Suffolk ewe lambs (fifteen lambs from single, twin, or triplet pregnancies; age 4–5 months) | decreased kisspeptin mRNA expression within kisspeptin-neurokinin B-dynorphin (KNDy) neurons | [21] | |
| aryl hydrocarbon receptor (AHR) signaling-modulating nutrients | ||||
| β-carotene 1 µM | HepG2 (hepatoma) cell line | aryl hydrocarbon receptor (AHR) activation |  | [22] |
| curcumin 1 µM | astrocyte culture derived from cerebral cortices collected from 2-day-old Sprague Dawley rats HepG2 (hepatoma) cell line | aryl hydrocarbon receptor (AHR) activation | 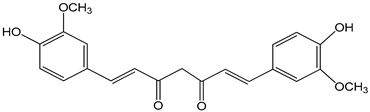 ((E,E)-1,7-bis(4-hydroxy-3-methoxyphenyl)-1,6-heptadiene-3,5-dione) | [22,23] |
| folic acid (pteroyl-L-glutamic acid) folic acid-deficient diet | HepG2 (hepatoma) cell line and C57BL/6 (B6) mice (B6 mice harboring the nonresponsive Ahrd allele (AhR null)) | suppression of aryl hydrocarbon receptor (AHR) transcriptional activity |  N-(4-{[(2-amino-4-oxo-3,4-dihydropteridin-6-yl)methyl]amino}benzoyl)-L-glutamic acid | [24] |
| high-fat diet (HFD) components: cholesterol (5-cholesten-3β-ol) (10 µg/mL), fructose (25 mM), palmitic acid (5 µM) | human aortic endothelial cells (HAEC) and human U937-derived macrophages (Umac) | aryl hydrocarbon receptor (AHR) activation | cholesterol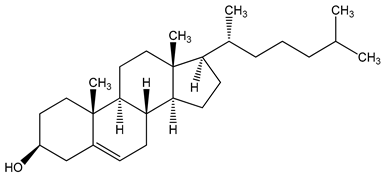 cholesterol (5-cholesten-3β-ol) cholesterol (5-cholesten-3β-ol)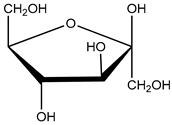 fructose | [25] |
| indole as tryptophan metabolite | aryl hydrocarbon receptor (AHR) activation | 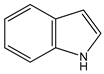 (1H-benzo[b]pyrrole) | [26] | |
| palmitate 100 µM | mHypoA-GnRH/GFP cell line (generated from the hypothalamus of a 2-month old transgenic GnRH-GFP female mouse) | modulation of spexin, and its receptors Galr2 and Galr3, in GnRH neurons | 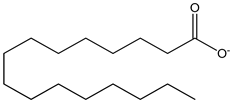 | [27] |
| tryptophan metabolites AIN93M standard diet for rodents supplemented with 0.5% tryptophan | female C57BL/6 WT and KO mice | aryl hydrocarbon receptor (AHR) activation | 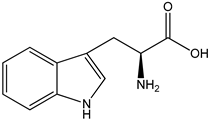 ((S)-2-amino-3-(3-indolyl)propionic acid) | [26,28] |
| vitamin B12 B12-deficient diet | HepG2 (hepatoma) cells and C57BL/6 (B6) mice (B6 mice harboring the nonresponsive Ahrd allele (AhR null)) | suppression of aryl hydrocarbon receptor (AHR) transcriptional activity |  α-(5,6-dimethylbenzimidazolyl)cobamidcyanide | [24] |
| catechol estrogens and estrogen sulfotransferase-modulating nutrients | ||||
| apigenin 0–50 µM | Sf9 cell line derived from pupa ovarian tissue of a fall armyworm; MCF-7 breast cancer cells | sulfotransferase family 1A member 1 (SULT1A1) induction | 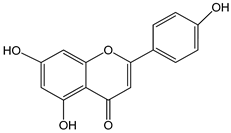 (5,7-dihydroxy-2-(4-hydroxyphenyl)-4-benzopyrone) | [29] |
| chrysin 0–10 µM | Sf9 cell line derived from pupa ovarian tissue of a fall armyworm MCF-7 breast cancer cells | sulfotransferase family 1E member 1 (SULT1E1) induction | 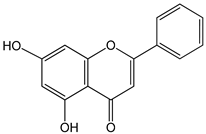 (5,7-dihydroxyflavone) | [29] |
| epicatechin 0–3000 µM | Sf9 cell line derived from pupa ovarian tissue of a fall armyworm MCF-7 breast cancer cells | sulfotransferase family 1A member 1 (SULT1A1) activation | 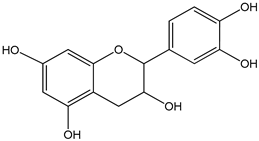 ((−)-cis-3,3′,4′,5,7-pentahydroxyflavane) | [29] |
| quercetin 0–10 µM | Sf9 cell line derived from pupa ovarian tissue of a fall armyworm MCF-7 breast cancer cells | sulfotransferase family 1E member 1 (SULT1E1) activation | 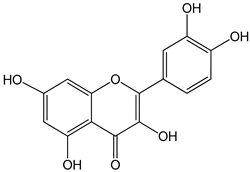 (2-(3,4-dihydroxyphenyl)-3,5,7-trihydroxychromen-4-one) | [29] |
| resveratrol 0–100 µM | Sf9 cell line derived from pupa ovarian tissue of a fall armyworm MCF-7 breast cancer cells | sulfotransferase family 1A member 1 (SULT1A1) activation | 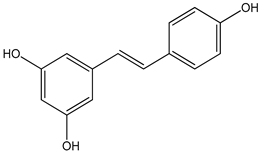 (3,4′,5-trihydroxy-trans-stilbene) | [29] |
| inflammatory–oxidative stress biomarker-modulating nutrients | ||||
| vitamin E 200 international units of alpha-tocopherol/d/8 weeks | postmenopausal women | reduction in hot flash episodes during the week improve the antioxidant status by increasing the total antioxidant capacity (TAC) levels | 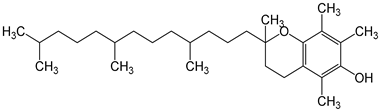 | [30,31] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Forma, E.; Urbańska, K.; Bryś, M. Menopause Hot Flashes and Molecular Mechanisms Modulated by Food-Derived Nutrients. Nutrients 2024, 16, 655. https://doi.org/10.3390/nu16050655
Forma E, Urbańska K, Bryś M. Menopause Hot Flashes and Molecular Mechanisms Modulated by Food-Derived Nutrients. Nutrients. 2024; 16(5):655. https://doi.org/10.3390/nu16050655
Chicago/Turabian StyleForma, Ewa, Karina Urbańska, and Magdalena Bryś. 2024. "Menopause Hot Flashes and Molecular Mechanisms Modulated by Food-Derived Nutrients" Nutrients 16, no. 5: 655. https://doi.org/10.3390/nu16050655
APA StyleForma, E., Urbańska, K., & Bryś, M. (2024). Menopause Hot Flashes and Molecular Mechanisms Modulated by Food-Derived Nutrients. Nutrients, 16(5), 655. https://doi.org/10.3390/nu16050655