
The Anti-Inflammatory and Curative Exponent of Probiotics: A Comprehensive and Authentic Ingredient for the Sustained Functioning of Major Human Organs

 , , ,
, , ,
Abstract
1. Introduction
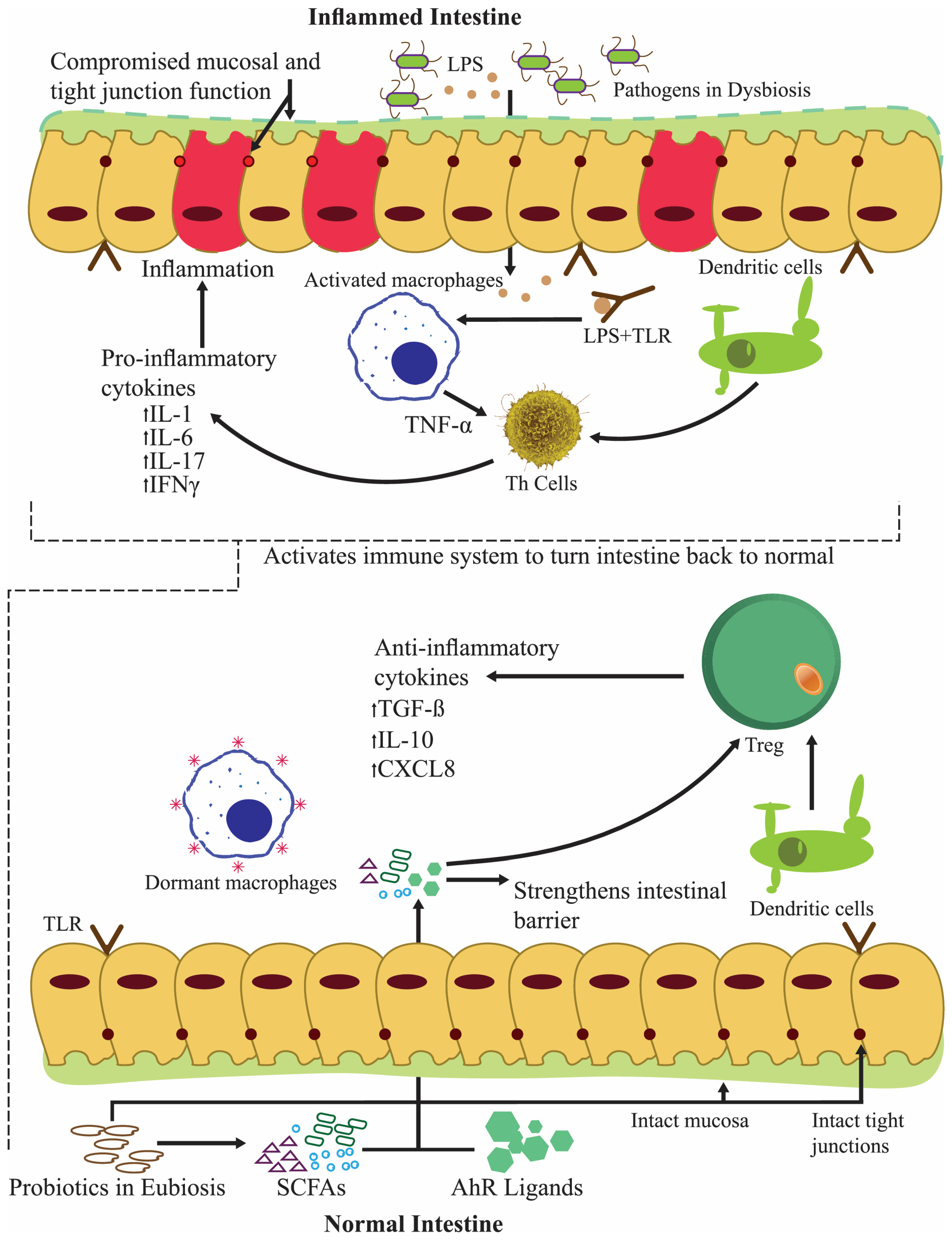
2. Probiotics Palliate Ailments in the Oral Cavity
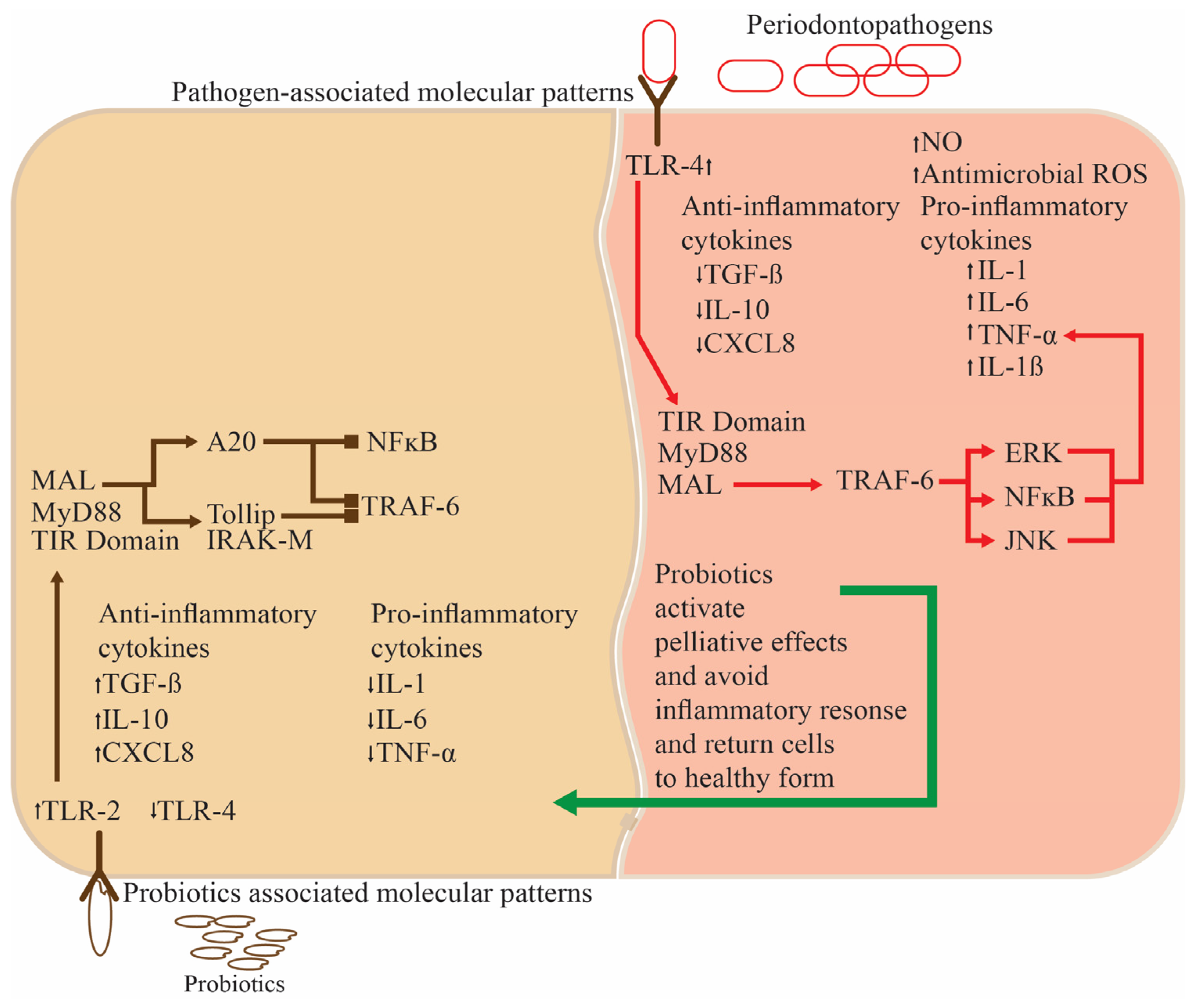
2.1. Periodontal Disorders and The Use of Probiotics to Alleviate Them
2.2. Streptococcus and Lactobacillus spp. as Therapeutic Agents against Dental Caries
2.3. Bifidobacterium and Lactobacillus spp. Curing Gingivitis
2.4. Halitosis and Its Establishment and Rectification by Probiotics
2.4.1. Streptococcus salivarius: An Efficacious Element against Halitosis
2.4.2. Streptococcus salivarius’s Enmity against Streptococcus pyogenes—An Oral Pathogen
2.5. Lactobacillus spp. as pH and Saliva Regulators in the Oral Cavity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of the Probiotic Strains | Age of the Participants | Dose of the Probiotics | Duration of Study | Outcomes of the Study | References |
|---|---|---|---|---|---|
| Probiotics Palliate Ailments in the Oral Cavity | |||||
| Lactobacillus. reuteri ATCC55730 and ATCC PTA5289 | ~38 years | 1 × 108 CFU/g each | 21 days | Gingival crevicular fluid (GCF) volume increased slightly, with a significantly increase in IL1-β and IL-18 and a significant decrease in IL-8 and MIP1-β also being found. | [58] |
| Lactobacillus reuteri PTA5289 | 31–46 years | 1 × 108 CFU/g | 14 days | A significant decrease in IL-17, TNF-α, and IL-1β, along with improved clinical indices, including clinical attachment level (CAL), periodontal probing depth (PPD), and sulcus bleeding index (SBI). | [59] |
| Lactobacillus curvatus EB10 DSM32307, Lactobacillus rhamnosus PB01 DSM14869 | 18–50 years | 1 × 108 CFU/g each | 28 days | Reduced bleeding on probing (BOP), amount of GCF, and decreased plaque levels. | [60] |
| Bifidobacterium animalis subsp. lactis DN 173010 | 16–26 years | 1 × 108 CFU/g | 28 days | Lowered gingivitis and plaque scores, lessened GCF volume and BOP, and lowered IL-1β concentration. | [24] |
| Bifidobacterium lactis BB-12, Lactobacillus rhamnosus GG | 13–15 years | 4.4 × 108 and 4.8 × 108 CFU/g | 28 days | Reduction in the gingival index (GI), plaque and Porphyromonas gingivalis in plaque, as well as a reduction in Aggregatibacter actinomycetemcomitans and Fusobacterium nucleatum in saliva. | [61] |
| Lactobacillus acidophilus, Enterococcus faecium Bifidobacterium infantis | 35–55 years | 1 × 107, 1 × 106, and 1 × 107 CFU/capsule | 30 days | A significant decrease in BOP after seven days and a reduction in the plaque index (PI), BOP, and periodontal pocket depth (PPD) after 30 days. | [62] |
| Bifidobacterium bifidum, Lactobacillus Acidophilus-HS101, Lactobacillus rhamnosus GG-HS111 | ≥60 years | 3.3 × 107 CFU/g | 60 days | Increased saliva in completely edentulous patients, which can be helpful in hyposalivation/xerostomia patients. | [51] |
| Lactobacillus rhamnosus GG, Bifidobacterium longum | 3–5 years | 7.5 × 105 and 4.5 × 105 CFU/mL of milk | 180 days | Significantly decreased Streptococcus mutans and pH, as well as the remineralization of 39.4% of caries. | [63] |
| Bifidobacterium lactis Bb-12, Lactobacillus acidophilus La-5 | 6–12 years | 1 × 106 CFU/g each | 30 days | Reduced Streptococcus mutans count after a week and also after 30 days. | [64] |
| Lactococcus reuteri ATCC PTA 5289 and DSM 17938 | 3–6 years | 1 × 108 CFU/g each | 28 days | Reduction in Mutans streptococci and lactobacilli and caries-associated bacterial counts. | [65] |
| Lactococcus rhamnosus GG, Lactobacillus helveticus, Lactococcus lactis, Lactococcus rhamnosus LC705, Propionibacterium freudenreichii ssp shermanii JS | 70–100 years | 1 × 107 CFU/g each | 120 days | Effectively controlled hyposalivation and oral Candida in the elderly. | [55] |
| Streptococcus salivarius M18 | 6–17 years | 1 × 109 CFU/ml | 90 days | Increased chances of avoiding the development of new dental caries in kids and reduced risk of tooth decay receptivity. | [66] |
| Lactobacillus salivarius WB21 | 22–67 years | 2.0 × 109 CFU/g | 14 days | Significantly decreased organoleptic test scores, the average probing pocket depth, the concentration of volatile sulfur compounds (VSCs), levels of Fusobacterium nucleatum and ubiquitous bacteria, which exhibited oral malodor and malodor-related factor control. | [67] |
| Lactobacillus salivarius Lactobacillus reuteri | 25–59 years | 2 × 109 CFU/g each | 90 days | Significantly reduced clinical and microbiological parameters and significantly improved bleeding index (BI), modified gingival index (MGI), and PI, leading to a significant decline in N-benzoyl-DL-arginine-naphthylamide and halitosis. | [68] |
| Lactobacillus reuteri ATCC PTA 5289 and DSM 17938 | 19–25 years | 1 × 108 CFU/g each | 28 days | Beneficial for oral malodor and malodourous compounds (other than VSCs) producing bacteria. | [69] |
| Streptococcus salivarius K12 | 23–44 years | 1 × 109 CFU/g | 30 days | Significantly decreased immediate organoleptic test (OLT) scores, tongue coating scores, and VSC levels in the absence of tongue coating. | [70] |
| Weissella cibaria CMU | 20–39 years | 1 × 108 CFU/g | 56 days | Significant decrease in OLT and VSC scores, along with bad breath improvement scores being reduced after eight weeks. | [71] |
| Probiotics Associated with the Small and Large Intestine | |||||
| Bifidobacterium longum BB536, Lactobacillus rhamnosus HN001 | 37–59 years | 4 × 108 and 1 × 108 CFU/g with 1.4 mg vitamin B6 | 60 days | Reduced abdominal pain, bloating, and disease severity; improved sucralose recovery (colonic permeability); increased relative abundance of hydrocarbons, butanoic, propanoic, and pentanoic acids; and decreased phenol. | [72] |
| Lactobacillus acidophilus subsp. helveticus LAFTI L10, Lactobacillus acidophilus NCFM | 30–60 years | 2.5 × 109 CFU/g each | 56 days | Significantly decreased flatus and composite scores. | [73] |
| Bifidobacterium longum, Lactobacillus paracasei | ≥18 years | 1 × 1010 CFU each | 84 days | Reduced symptoms in IBS and addition to the armamentarium of IBS management tools dependent on the IBS subtype. | [73] |
| Bifidobacterium bifidum HI-MIMBb75 | ≥18 years | 1 × 109 CFU/capsule (Non-viable) | 56 days | Substantially alleviated IBS and its symptoms and mediated specific beneficial effects independent of cell viability. | [74] |
| Bifidobacterium longum, Bifidobacterium breve, Lactobacillus paracasei HII01 | ~60 years | 2.0 × 1010, 2.0 × 1010, and 1.0 × 1010 CFU/g | 84 days | Improved intestinal barrier function (up to 48%), enhanced short-chain fatty acid levels, improved obesity-related anthropometric biomarkers, and significantly increased high-density lipoprotein–cholesterol. | [75] |
| Lactobacillus acidophilus W37, Lactococcus lactis W19 and W58, Lactobacillus brevis W63, Lactobacillus salivarius W24, Lactobacillus casei W56, Bifidobacterium lactis W52, Bifidobacterium bifidum W23, | 18–80 years | 2.5 × 109 CFU/g each | 180 days | Significantly increased production of reactive oxygen species by neutrophils and serum neopterin levels, maintaining or even improving liver functioning in sturdy cirrhosis with a slight impact on bacterial translocation and gut barrier function. | [76] |
| Clostridium butyricum | ≥18 years | 1 × 106 CFU/g | 15 days | Shortened duration of fever and constipation and significantly decreased bactericides, Escherichia coli, and Enterococcus. | [77] |
| Lactobacillus paracasei W20, Lactobacillus plantarum W1 and W62, Bifidobacterium bifidum W23, Lactobacillus acidophilus W37 and W55, Lactobacillus rhamnosus W71 Lactobacillus salivarius W24, Enterococcus faecium W54, Bifidobacterium lactis W51, | 45–65 years | 1.1 × 109 CFU/g each | 28 days | Increased probiotic strains in stool and improved microbiome composition and functional diversity, successfully modulating the microbiome and ultimately intervening in sepsis. | [78] |
| Bifidobacterium spp. | 27–55 years | - | 60 days | Decreased plasma levels of hs-CRP, TNF-α, plasma DA0, ET, D-lactic acid, IL-8, and IL-6 and increased CD4/CD8 ratio and CD4+ levels, enhancing the remedying impact in ulcerative colitis patients and regulating T cell frequency, in addition to reducing plasma inflammatory factors. | [79] |
| Bifidobacterium longum Enterococcus faecium, Lactobacillus acidophilus, Lactobacillus plantarum, Bifidobacterium lactis, Streptococcus thermophilus, | ≥18 years | 3 × 109 CFU/g each | 56 days | Decreased expression of serum C-reactive protein (CRP), significantly improved endoscopic and clinical activities, and positive impacts on the acute-phase reactants and endoscopic activity levels. | [80] |
| Lactobacillus rhamnosus NCIMB 30174, Enterococcus faecium NCIMB 30176, Lactobacillus acidophilus NCIMB 30175, Lactobacillus plantarum NCIMB 30173, | 18–70 years | 1 × 1010 CFU/g each | 28 days | Significantly reduced fecal calprotectin levels in ulcerative colitis patients and decreased intestinal inflammation. | [81] |
| Lactobacillus plantarum 299v | ≥18 years | 1 × 1010 CFU/g | 84 days | Reduced enteral nutrition-related gastrointestinal symptoms. Effectively improved the quality of life of cancer patients, nutritional status, and enteral nutrition tolerance. | [82] |
| Probiotics Tone Liver and Annihilate its Ailments | |||||
| Bifidobacterium spp., Lactobacillus spp., Enterococcus spp. | 18–59 years | - | 90 days | Significantly improved aspartate aminotransferase (AST), NAFLD activity score (NAS), total cholesterol (TC), alanine aminotransferase (ALT), glutamine transferase (GGT), triglyceride (TG) levels, and insulin resistance index (HOMA-IR). Improved liver functions, hepatic fatty deposition, and glucose and lipids metabolism in NAFLD patients, enhancing the therapeutic effects. | [83] |
| Pediococcus pentosaceus CBT SL4, Lactobacillus paracasei CBT LPC5, Lactobacillus rhamnosus CBT LR5, Lactobacillus acidophilus CBT LA1, Bifidobacterium breve CBT BR3, Bifidobacterium lactis CBT BL3, | 19–75 years | 1 × 109 CFU/1.4 g each | 84 days | Significantly decreased intrahepatic fat fraction after 12 weeks, along with significant triglyceride reduction. | [84] |
| Bifidobacterium animalis subsp. lactis BB-12 | ≥18 years | 1 × 108 CFU/g | 168 days | Significantly decreased alkaline phosphatase, aspartate aminotransferase, γ-glutamyltransferase, and alanine aminotransferase in serum and reduced NAFLD. | [85] |
| Acetobacter spp., Bifidobacterium spp., Propionibacterium spp., Lactobacillus spp., + Lactococcu spp., | 18–65 years | 6 × 1010, 1 × 1010, 3 × 1010, 1 × 106 CFU/g | 56 days | Significantly reduced the fatty liver index, serum GGT and AST values; diminished chronic systemic inflammatory state; and lowered IL-6 and TNF-α concentrations in NAFLD patients. | [86] |
| Lactobacillus lactis BCMC 12451, Lactobacillus casei BCMC 12313, Lactobacillus acidophilus BCMC 12130, Bifidobacterium longum BCMC 02120, Bifidobacterium infantis BCMC 02129, Bifidobacterium bifidum BCMC 02290 | 18 years and above | 3 × 109 CFU/g | 180 days | Stabilized mucosal immune function, protecting against risen intestinal permeability and playing a complementary role in ministering NAFLD. | [87] |
| Probiotics as Allayers of Gallbladder and Pancreatic Ailments | |||||
| Clostridium butyricum MIYAIRI | 35.5 ± 9.9 | 5 × 109 CFU/g | 180 days | Decreased incidence of gall bladder disease, adverse drug effects, and poor drug compliance rates, confirming the palliative effects of probiotics. | [88] |
| Lactobacillus acidophilus | 48.1 ± 13.8 | 5 × 106 CFU/g | 14 days | Significantly altered serum low-density lipoprotein cholesterol (LDL-C), total cholesterol, total bile acid (TBA), and triglyceride levels. Significantly differed glycoprotein, pH, and free Ca2+ of bile. Altered deoxycholic acid, chenodeoxycholic acid, and cholic acid levels, exhibiting the reverse development of bile composition in patients with cholecystolithiasis taking probiotics, thereby diminishing gallstones. | [89] |
| Enterococcus faecium Bacillus subtilis | 18–75 years | Manufacturer defined recipe | Significantly reduced length of stay (LOS) and shortened abdominal pain relief and oral feeding duration in patients with acute pancreatitis. | [90] | |
| Bifidobacterium infantalis, Bifidobacterium longus, Bifidobacterium bifidum, Lactobacillus acidophilus | 13–79 years | 2.5 × 109 CFU/g | 7 days | Significantly reduced immunoglobulins and C-reactive protein expression. | [91] |
| Acetobacter spp., Lactobacillus + Lactococcus spp., Propionibacterium spp., Bifidobacterium spp., | 18–75 years | 1 × 106, 6 × 1010, 3 × 1010, 1 × 1010 CFU/g | 56 days | Significantly improved β-cell function and reduced fasting glucose and hemoglobin A1C levels. Significantly affected chronic systemic inflammation by decreasing pro-inflammatory cytokines. | [92] |
| Bacillus mesentericus TO-A-, Clostridium butyricum TO-A, Lactobacillus Sporogenes, Streptococcus faecalis T-110 | 18–75 years | 1 × 108, 4 × 106, 2 × 106, 6 × 107 CFU/g | 15 days | Significantly lowered LOS, the duration of antibiotics therapy, and the incidence of postoperative infectious complications in patients with chronic pancreatitis. | [93] |
| Lactobacillus casei, Bifidobacterium bifidum Lactobacillus acidophilus, Lactobacillus rhamnosus | ≥18 years | 1 × 109 CFU/g | 90 days | Significantly reduced bowel frequency and total cholesterol levels. Significantly increased red blood cells, hematocrit, hemoglobin, albumin, serum magnesium, and total lymphocyte count. | [94] |
3. Probiotics Proven to Be Beneficial in Small and Large Intestine Disorders
3.1. Lactobacillus spp. as Small Intestinal Alleviators
3.2. The Palliative Intestinal Permeability of Lactobacillus spp.
3.3. Probiotics as Lenitives against Impaired Nutrient Absorption and Chronic Diarrhea
3.4. Probiotics Modulating Large Intestinal Microflora
3.5. The Alleviating Influence of Lactobacillus spp. on Colitis
3.6. Bifidobacterium, Lactobacillus, and Other Probiotic spp. Modulating Gastrointestinal Cancers
4. Probiotics Tone Liver and Annihilate Its Ailments
Probiotics Rectifying Non-Alcoholic Fatty Liver Disease (NAFLD) and Alcoholic Liver Disease (ALD)
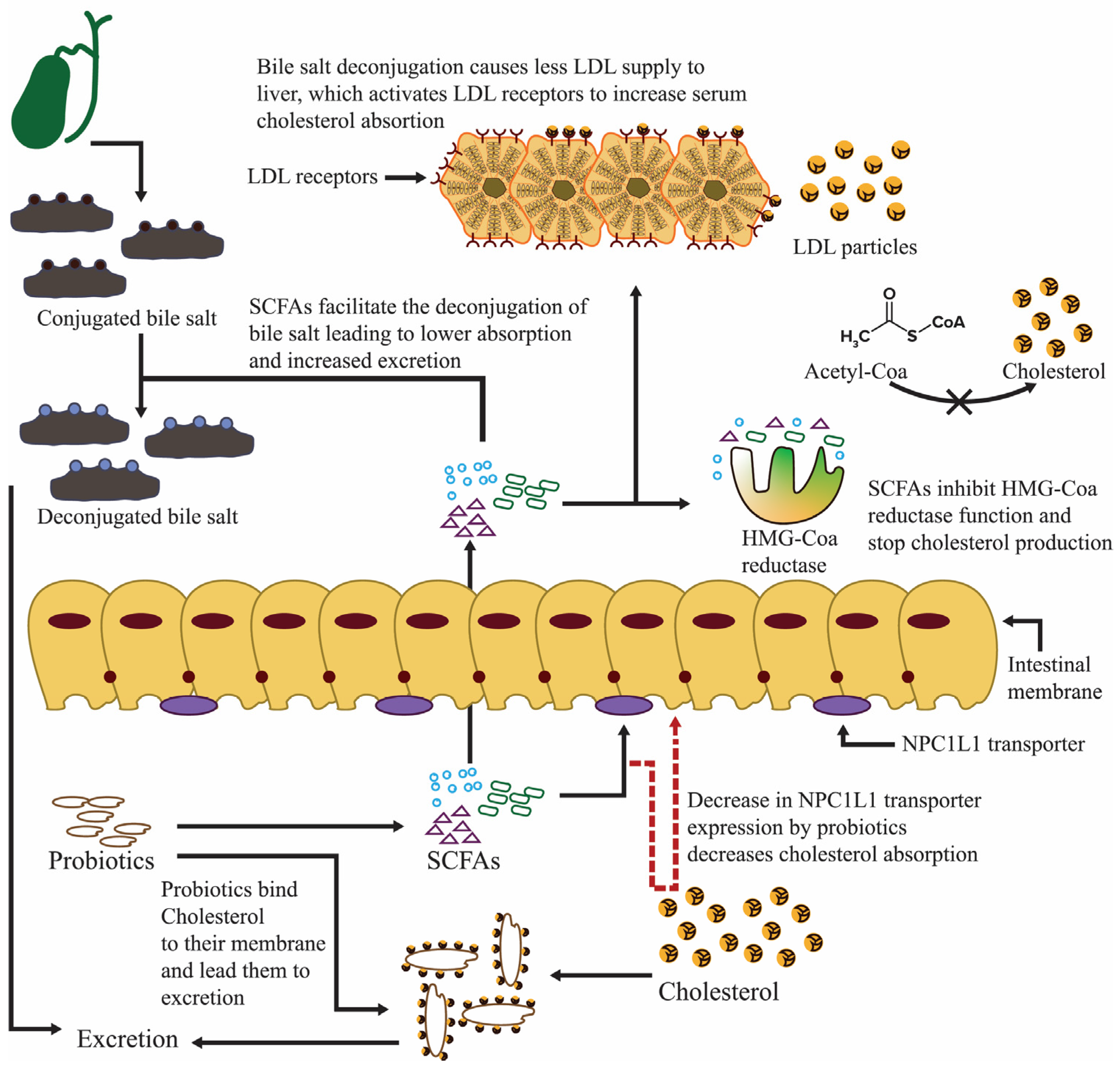
5. Probiotics as Allayers of Gallbladder Ailments
5.1. Probiotics Repress Bile Acid Production and Diminish Gallstones
5.2. Probiotics’ Connection with Bacterial Translocation and Acute and Chronic Pancreatitis
5.3. Probiotics Lower the Risks of Organ Failure and Systemic Inflammatory Response Syndrome
6. Probiotics Fortify the Respiratory Tract and Alleviate Rhinosinusitis and Rhinitis
6.1. Probiotics Have Yielded Encouraging Findings against Asthma
6.2. Probiotics Counter Allergy Illnesses
7. Probiotics Combat Osteoporosis and Build Up the Skeleton
7.1. Probiotics Meliorate Bone Health
7.2. Probiotics Palliate Rheumatoid Arthritis
8. Probiotics Preserve Kidney Integrity and Combat Chronic Kidney Disease (CKD)
Probiotics Conciliate Kidney Ailments during Hemodialysis and Peritoneal Dialysis
9. Probiotics Influence the Microflora of the Male and Female Reproductive Systems
9.1. Probiotics Enhance Cell Membrane Integrity and Functioning
9.2. Probiotics Conserve Male Reproductive Health
9.2.1. Probiotics from the Vagina Protect Spermatozoa
9.2.2. Probiotics Meliorate Reproductive Hormone Release
9.2.3. Probiotics Attenuate DNA Damage, Blood–Testis Barrier, and Spermatozoa Functionality
9.2.4. Probiotics Attenuate Prostatitis and Modulate Lactic Acid Production
9.3. Probiotics Alleviate Bacterial Vaginosis (BV)
9.4. Products from Probiotics Hinder Sexually Transmitted Infections (STIs)
9.5. Probiotics Countermine HIV and Herpes Simplex Virus 2 (HSV-2)
9.6. Probiotics Improve Ovarian Function
9.7. Probiotics Maintain the Steady Provision of Crucial Elements through the Placenta
10. Probiotics Preclude the Prevalence of Cardiovascular Disease (CVD)
10.1. Probiotics Regulate Plasma Glucose and Inulin Levels
10.2. Probiotics Engage as Comforters in Coronary Artery Disease (CAD)
10.3. Probiotics Diminish Inflammation-Associated Ailments
11. Probiotics Alleviate Neurodegenerative and Neurodevelopmental Disorders
11.1. Probiotics Combat Insomnia
11.2. Probiotics Assist in Accommodating for Autism Spectrum Disorder (ASD)
11.3. Probiotics Help in Coping with Attention Deficit Hyperactivity Disorder (ADHD)
11.4. Probiotics Temper Parkinson’s and Alzheimer’s Disease
12. Conclusions and Future Prospects
Author Contributions
Funding
Conflicts of Interest
References
- Hillman, E.T.; Lu, H.; Yao, T.; Nakatsu, C.H. Microbial Ecology along the Gastrointestinal Tract. Microbes Environ. 2017, 32, 300–313. [Google Scholar] [CrossRef]
- Guarner, F.; Malagelada, J.-R. Gut flora in health and disease. Lancet 2003, 361, 512–519. [Google Scholar] [CrossRef]
- Proctor, L.M. The Human Microbiome Project in 2011 and beyond. Cell Host Microbe 2011, 10, 287–291. [Google Scholar] [CrossRef]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the Human Intestinal Microbial Flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef]
- Ley, R.E.; Hamady, M.; Lozupone, C.; Turnbaugh, P.J.; Ramey, R.R.; Bircher, J.S.; Schlegel, M.L.; Tucker, T.A.; Schrenzel, M.D.; Knight, R.; et al. Evolution of mammals and their gut microbes. Science 2008, 320, 1647–1651. [Google Scholar] [CrossRef]
- Ley, R.E.; Bäckhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef]
- Lourens-Hattingh, A.; Viljoen, B.C. Yogurt as probiotic carrier food. Int. Dairy J. 2001, 11, 1–17. [Google Scholar] [CrossRef]
- Kang, J.H.; Yun, S., II; Park, M.H.; Park, J.H.; Jeong, S.Y.; Park, H.O. Anti-Obesity Effect of Lactobacillus gasseri BNR17 in High-Sucrose Diet-Induced Obese Mice. PLoS ONE 2013, 8, e54617. [Google Scholar] [CrossRef] [PubMed]
- Meenakshi, I.K. In-Vitro Characterization of the Anti-Cancer Activity of the Probiotic Bacterium Lactobacillus fermentum NCIMB 5221 and Potential against Colorectal Cancer. J. Cancer Sci. Ther. 2015, 7, 224–235. [Google Scholar] [CrossRef]
- Grover, S.; Rashmi, H.M.; Srivastava, A.K.; Batish, V.K. Probiotics for human health -new innovations and emerging trends. Gut Pathog. 2012, 4, 15. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.-J.; Im, S.-H. Probiotics as an Immune Modulator. J. Nutr. Sci. Vitaminol. 2015, 61 (Suppl. S61), S103–S105. [Google Scholar] [CrossRef]
- Britton, R.A.; Versalovic, J. Probiotics and Gastrointestinal Infections. Interdiscip. Perspect. Infect. Dis. 2008, 2008, 290769. [Google Scholar] [CrossRef]
- Cookson, T.A. Bacterial-Induced Blood Pressure Reduction: Mechanisms for the Treatment of Hypertension via the Gut. Front. Cardiovasc. Med. 2021, 8, 721393. [Google Scholar] [CrossRef]
- Guarino, M.; Altomare, A.; Emerenziani, S.; Di Rosa, C.; Ribolsi, M.; Balestrieri, P.; Iovino, P.; Rocchi, G.; Cicala, M. Mechanisms of Action of Prebiotics and Their Effects on Gastro-Intestinal Disorders in Adults. Nutrients 2020, 12, 1037. [Google Scholar] [CrossRef]
- Markowiak, P.; Śliżewska, K. Effects of Probiotics, Prebiotics, and Synbiotics on Human Health. Nutrients 2017, 9, 1021. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, B.; Delgado, S.; Blanco-Míguez, A.; Lourenço, A.; Gueimonde, M.; Margolles, A. Probiotics, gut microbiota, and their influence on host health and disease. Mol. Nutr. Food Res. 2017, 61, 1600240. [Google Scholar] [CrossRef] [PubMed]
- Mahasneh, S.A.; Mahasneh, A.M. Probiotics: A promising role in dental health. Dent. J. 2017, 5, 26. [Google Scholar] [CrossRef]
- Allaker, R.P.; Stephen, A.S. Use of Probiotics and Oral Health. Curr. Oral Health Rep. 2017, 4, 309–318. [Google Scholar] [CrossRef] [PubMed]
- İnce, G.; Gürsoy, H.; İpçi, Ş.D.; Cakar, G.; Emekli-Alturfan, E.; Yılmaz, S. Clinical and Biochemical Evaluation of Lozenges Containing Lactobacillus reuteri as an Adjunct to Non-Surgical Periodontal Therapy in Chronic Periodontitis. J. Periodontol. 2015, 86, 746–754. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, T.; Kojima, Y.; Seneviratne, C.J.; Maeda, N. Therapeutic application of synbiotics, a fusion of probiotics and prebiotics, and biogenics as a new concept for oral Candida infections: A mini review. Front. Microbiol. 2016, 7, 10. [Google Scholar] [CrossRef] [PubMed]
- Sivamaruthi, B.S.; Kesika, P.; Chaiyasut, C. A Review of the Role of Probiotic Supplementation in Dental Caries. Probiotics Antimicrob. Proteins 2020, 12, 1300–1309. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.X.; Zhong, Y.J.; Dong, Q.Q.; Wong, H.M.; Wen, Y.F. Global, regional, and national burden of severe periodontitis, 1990–2019: An analysis of the Global Burden of Disease Study 2019. J. Clin. Periodontol. 2021, 48, 1165–1188. [Google Scholar] [CrossRef] [PubMed]
- Kuru, B.E.; Laleman, I.; Yalnızoğlu, T.; Kuru, L.; Teughels, W. The Influence of a Bifidobacterium animalis Probiotic on Gingival Health: A Randomized Controlled Clinical Trial. J. Periodontol. 2017, 88, 1115–1123. [Google Scholar] [CrossRef] [PubMed]
- Selwitz, R.H.; Ismail, A.I.; Pitts, N.B. Dental caries. Lancet 2007, 369, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Tao, R.; Jurevic, R.J.; Coulton, K.K.; Tsutsui, M.T.; Roberts, M.C.; Kimball, J.R.; Wells, N.; Berndt, J.; Dale, B.A. Salivary antimicrobial peptide expression and dental caries experience in children. Antimicrob. Agents Chemother. 2005, 49, 3883–3888. [Google Scholar] [CrossRef] [PubMed]
- Shifa, S.; Muthu, M.S.; Amarlal, D.; Rathna Prabhu, V. Quantitative assessment of IgA levels in the unstimulated whole saliva of caries-free and caries-active children. J. Indian Soc. Pedod. Prev. Dent. 2008, 26, 158–161. [Google Scholar]
- Comelli, E.M.; Guggenheim, B.; Stingele, F.; Neeser, J.R. Selection of dairy bacterial strains as probiotics for oral health. Eur. J. Oral Sci. 2002, 110, 218–224. [Google Scholar] [CrossRef]
- Kang, M.S.; Chung, J.; Kim, S.M.; Yang, K.H.; Oh, J.S. Effect of Weissella cibaria isolates on the formation of Streptococcus mutans biofilm. Caries Res. 2006, 40, 418–425. [Google Scholar] [CrossRef]
- Nase, L.; Hatakka, K.; Savilahti, E.; Saxelin, M.; Pönkä, A.; Poussa, T.; Korpela, R.; Meurman, J.H. Effect of Long-Term Consumption of a Probiotic Bacterium, Lactobacillus rhamnosus GG, in Milk on Dental Caries and Caries Risk in Children. Caries Res. 2001, 35, 412–420. [Google Scholar] [CrossRef]
- Çaglar, E.; Kavaloglu, S.C.; Kuscu, O.O.; Sandalli, N.; Holgerson, P.L.; Twetman, S. Effect of chewing gums containing xylitol or probiotic bacteria on salivary mutans streptococci and lactobacilli. Clin. Oral Investig. 2007, 11, 425–429. [Google Scholar] [CrossRef]
- Nikawa, H.; Makihira, S.; Fukushima, H.; Nishimura, H.; Ozaki, Y.; Ishida, K.; Darmawan, S.; Hamada, T.; Hara, K.; Matsumoto, A.; et al. Lactobacillus reuteri in bovine milk fermented decreases the oral carriage of mutans streptococci. Int. J. Food Microbiol. 2004, 95, 219–223. [Google Scholar] [CrossRef]
- Bolla, V.L.; Reddy, M.S.; Srinivas, N.; Reddy, C.S.; Koppolu, P. Investigation and comparison of the effects of two probiotic bacteria, and in reducing mutans streptococci levels in the saliva of children. Ann. Afr. Med. 2022, 21, 395–402. [Google Scholar] [CrossRef]
- Krasse, P.; Carlsson, B.; Dahl, C.; Paulsson, A.; Nilsson, A.; Sinkiewicz, G. Decreased gum bleeding and reduced gingivitis by the probiotic Lactobacillus reuteri. Swed. Dent. J. 2006, 30, 55–60. [Google Scholar] [PubMed]
- Alkaya, B.; Laleman, I.; Keceli, S.; Ozcelik, O.; Cenk Haytac, M.; Teughels, W. Clinical effects of probiotics containing Bacillus species on gingivitis: A pilot randomized controlled trial. J. Periodontal Res. 2017, 52, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Toiviainen, A.; Jalasvuori, H.; Lahti, E.; Gursoy, U.; Salminen, S.; Fontana, M.; Flannagan, S.; Eckert, G.; Kokaras, A.; Paster, B.; et al. Impact of orally administered lozenges with Lactobacillus rhamnosus GG and Bifidobacterium animalis subsp. lactis BB-12 on the number of salivary mutans streptococci, amount of plaque, gingival inflammation and the oral microbiome in healthy adults. Clin. Oral Investig. 2015, 19, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Sabatini, S.; Lauritano, D.; Candotto, V.; Silvestre, F.J.; Nardi, G.M. Oral probiotics in the management of gingivitis in diabetic patients: A double blinded randomized controlled study. J. Biol. Regul. Homeost. Agents 2017, 31, 197–202. [Google Scholar] [PubMed]
- van den Velde, S.; Quirynen, M.; Van Hee, P.; van Steenberghe, D. Halitosis associated volatiles in breath of healthy subjects. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2007, 853, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Campisi, G.; Musciotto, A.; Di Fede, O.; Di Marco, V.; Craxì, A. Halitosis: Could it be more than mere bad breath? Intern. Emerg. Med. 2011, 6, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Scully, C.; Greenman, J. Halitosis (breath odor). Periodontol. 2000 2008, 48, 66–75. [Google Scholar] [CrossRef]
- Hyink, O.; Wescombe, P.A.; Upton, M.; Ragland, N.; Burton, J.P.; Tagg, J.R. Salivaricin A2 and the novel lantibiotic salivaricin B are encoded at adjacent loci on a 190-kilobase transmissible megaplasmid in the oral probiotic strain Streptococcus salivanus K12. Appl. Environ. Microbiol. 2007, 73, 1107–1113. [Google Scholar] [CrossRef]
- Burton, J.P.; Chilcott, C.N.; Moore, C.J.; Speiser, G.; Tagg, J.R. A preliminary study of the effect of probiotic Streptococcus salivarius K12 on oral malodour parameters. J. Appl. Microbiol. 2006, 100, 754–764. [Google Scholar] [CrossRef]
- Burton, J.P.; Chilcott, C.N.; Tagg, J.R. The rationale and potential for the reduction of oral malodour using Streptococcus salivarius probiotics. Oral Dis. 2005, 11, 29–31. [Google Scholar] [CrossRef]
- Allaker, R.P.; Waite, R.D.; Hickling, J.; North, M.; McNab, R.; Bosma, M.P.; Hughes, F.J. Topographic distribution of bacteria associated with oral malodour on the tongue. Arch. Oral Biol. 2008, 53, S8–S12. [Google Scholar] [CrossRef]
- Eren, A.M.; Borisy, G.G.; Huse, S.M.; Mark Welch, J.L. Oligotyping analysis of the human oral microbiome. Proc. Natl. Acad. Sci. USA 2014, 111, E2875–E2884. [Google Scholar] [CrossRef]
- Zaura, E.; Keijser, B.J.; Huse, S.M.; Crielaard, W. Defining the healthy “core microbiome” of oral microbial communities. BMC Microbiol. 2009, 9, 259. [Google Scholar] [CrossRef] [PubMed]
- Kanwal, S.; Vaitla, P. Streptococcus Pyogenes; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Dierksen, K.P.; Moore, C.J.; Inglis, M.; Wescombe, P.A.; Tagg, J.R. The effect of ingestion of milk supplemented with salivaricin A-producing Streptococcus salivarius on the bacteriocin-like inhibitory activity of streptococcal populations on the tongue. FEMS Microbiol. Ecol. 2007, 59, 584–591. [Google Scholar] [CrossRef]
- Nadelman, P.; Magno, M.B.; Masterson, D.; da Cruz, A.G.; Maia, L.C. Are dairy products containing probiotics beneficial for oral health? A systematic review and meta-analysis. Clin. Oral Investig. 2018, 22, 2763–2785. [Google Scholar] [CrossRef]
- Srivastava, S.; Saha, S.; Kumari, M.; Mohd, S. Effect of Probiotic Curd on Salivary pH and Streptococcus mutans: A Double Blind Parallel Randomized Controlled Trial. J. Clin. Diagn. Res. 2016, 10, ZC13–ZC16. [Google Scholar] [CrossRef] [PubMed]
- Sanghvi, U.; Chhabra, T.; Sethuraman, R. Effect of probiotics on the amount and pH of saliva in edentulous patients: A Prospective study. J. Indian Prosthodont. Soc. 2018, 18, 277–281. [Google Scholar] [PubMed]
- Pradeep, K.; Kuttappa, M.A.; Prasana, K.R. Probiotics and oral health: An update. SADJ 2014, 69, 20–24. [Google Scholar]
- Wallace, T.C.; Guarner, F.; Madsen, K.; Cabana, M.D.; Gibson, G.; Hentges, E.; Sanders, M.E. Human gut microbiota and its relationship to health and disease. Nutr. Rev. 2011, 69, 392–403. [Google Scholar] [CrossRef]
- Mack, D.R.; Ahrne, S.; Hyde, L.; Wei, S.; Hollingsworth, M.A. Extracellular MUC3 mucin secretion follows adherence of Lactobacillus strains to intestinal epithelial cells in vitro. Gut 2003, 52, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Hatakka, K.; Ahola, A.J.; Yli-Knuuttila, H.; Richardson, M.; Poussa, T.; Meurman, J.H.; Korpela, R. Probiotics Reduce the Prevalence of Oral Candida in the Elderly—A Randomized Controlled Trial. J. Dent. Res. 2007, 86, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Kusumaningsih, T.; Subijanto, M.S.; Indrawati, R.; Devijanti, R.R. The level of beta defensin-2 in saliva and its expression in parotid gland epithelial cells after probiotic (Lactobacillus reuteri) induction to inhibit Streptococcus mutans in caries. Eur. J. Dent. 2016, 10, 556–560. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lundtorp-Olsen, C.; Enevold, C.; Juel Jensen, C.A.; Stofberg, S.N.; Twetman, S.; Belstrøm, D. Impact of Probiotics on the Salivary Microbiota and Salivary Levels of Inflammation-Related Proteins during Short-Term Sugar Stress: A Randomized Controlled Trial. Pathogens 2021, 10, 392. [Google Scholar] [CrossRef] [PubMed]
- Hallström, H.; Lindgren, S.; Yucel-Lindberg, T.; Dahlén, G.; Renvert, S.; Twetman, S. Effect of probiotic lozenges on inflammatory reactions and oral biofilm during experimental gingivitis. Acta Odontol. Scand. 2013, 71, 828–833. [Google Scholar] [CrossRef] [PubMed]
- Szkaradkiewicz, A.K.; Stopa, J.; Karpiński, T.M. Effect of Oral Administration Involving a Probiotic Strain of Lactobacillus reuteri on Pro-Inflammatory Cytokine Response in Patients with Chronic Periodontitis. Arch. Immunol. Ther. Exp. 2014, 62, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Keller, M.K.; Brandsborg, E.; Holmstrøm, K.; Twetman, S. Effect of tablets containing probiotic candidate strains on gingival inflammation and composition of the salivary microbiome: A randomised controlled trial. Benef. Microbes 2018, 9, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Alanzi, A.; Honkala, S.; Honkala, E.; Varghese, A.; Tolvanen, M.; Söderling, E. Effect of Lactobacillus rhamnosus and Bifidobacterium lactis on gingival health, dental plaque, and periodontopathogens in adolescents: A randomised placebo-controlled clinical trial. Benef. Microbes 2018, 9, 593–602. [Google Scholar] [CrossRef]
- Minić, I.; Pejčić, A.; Bradić-Vasić, M. Effect of the local probiotics in the therapy of periodontitis A randomized prospective study. Int. J. Dent. Hyg. 2022, 20, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Angarita-Díaz, M.P.; Forero-Escobar, D.; Cerón-Bastidas, X.A.; Cisneros-Hidalgo, C.A.; Dávila-Narvaez, F.; Bedoya-Correa, C.M.; Freitas, S.C.; Cabrera-Arango, C.L.; Melo-Colina, R. Effects of a functional food supplemented with probiotics on biological factors related to dental caries in children: A pilot study. Eur. Arch. Paediatr. Dent. 2020, 21, 161–169. [Google Scholar] [CrossRef]
- Ashwin, D.; Ke, V.; Taranath, M.; Ramagoni, N.K.; Nara, A.; Sarpangala, M. Effect of Probiotic Containing Ice-cream on Salivary Mutans Streptococci (SMS) Levels in Children of 6-12 Years of Age: A Randomized Controlled Double Blind Study with Six-months Follow Up. J. Clin. Diagn. Res. 2015, 9, ZC06–ZC09. [Google Scholar] [CrossRef] [PubMed]
- Alamoudi, N.M.; Almabadi, E.S.; El Ashiry, E.A.; El Derwi, D.A. Effect of Probiotic Lactobacillus reuteri on Salivary Cariogenic Bacterial Counts among Groups of Preschool Children in Jeddah, Saudi Arabia: A Randomized Clinical Trial. J. Clin. Pediatr. Dent. 2018, 42, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Di Pierro, F.; Zanvit, A.; Nobili, P.; Risso, P.; Fornaini, C. Cariogram outcome after 90 days of oral treatment with Streptococcus salivarius M18 in children at high risk for dental caries: Results of a randomized, controlled study. Clin. Cosmet. Investig. Dent. 2015, 7, 107–113. [Google Scholar] [CrossRef]
- Suzuki, N.; Yoneda, M.; Tanabe, K.; Fujimoto, A.; Iha, K.; Seno, K.; Yamada, K.; Iwamoto, T.; Masuo, Y.; Hirofuji, T. Lactobacillus salivarius WB21–containing tablets for the treatment of oral malodor: A double-blind, randomized, placebo-controlled crossover trial. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2014, 117, 462–470. [Google Scholar] [CrossRef]
- Penala, S.; Kalakonda, B.; Pathakota, K.R.; Jayakumar, A.; Koppolu, P.; Lakshmi, B.V.; Pandey, R.; Mishra, A. Efficacy of local use of probiotics as an adjunct to scaling and root planing in chronic periodontitis and halitosis: A randomized controlled trial. J. Res. Pharm. Pract. 2016, 5, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Keller, M.K.; Bardow, A.; Jensdottir, T.; Lykkeaa, J.; Twetman, S. Effect of chewing gums containing the probiotic bacterium Lactobacillus reuteri on oral malodour. Acta Odontol. Scand. 2012, 70, 246–250. [Google Scholar] [CrossRef]
- He, L.; Yang, H.; Chen, Z.; Ouyang, X. The Effect of Streptococcus salivarius K12 on Halitosis: A Double-Blind, Randomized, Placebo-Controlled Trial. Probiotics Antimicrob. Proteins 2020, 12, 1321–1329. [Google Scholar] [CrossRef]
- Lee, D.-S.; Lee, S.-A.; Kim, M.; Nam, S.-H.; Kang, M.-S. Reduction of Halitosis by a Tablet Containing Weissella cibaria CMU: A Randomized, Double-Blind, Placebo-Controlled Study. J. Med. Food 2020, 23, 649–657. [Google Scholar] [CrossRef]
- Bonfrate, L.; Di Palo, D.M.; Celano, G.; Albert, A.; Vitellio, P.; De Angelis, M.; Gobbetti, M.; Portincasa, P. Effects of Bifidobacterium longum BB536 and Lactobacillus rhamnosus HN001 in IBS patients. Eur. J. Clin. Investig. 2020, 50, e13201. [Google Scholar] [CrossRef]
- Sadrin, S.; Sennoune, S.; Gout, B.; Marque, S.; Moreau, J.; Zinoune, K.; Grillasca, J.-P.; Pons, O.; Maixent, J.-M. A 2-strain mixture of Lactobacillus acidophilus in the treatment of irritable bowel syndrome: A placebo-controlled randomized clinical trial. Dig. Liver Dis. 2020, 52, 534–540. [Google Scholar] [CrossRef] [PubMed]
- Andresen, V.; Gschossmann, J.; Layer, P. Heat-inactivated Bifidobacterium bifidum MIMBb75 (SYN-HI-001) in the treatment of irritable bowel syndrome: A multicentre, randomised, double-blind, placebo-controlled clinical trial. Lancet Gastroenterol. Hepatol. 2020, 5, 658–666. [Google Scholar] [CrossRef]
- Chaiyasut, C.; Sivamaruthi, B.S.; Lailerd, N.; Sirilun, S.; Khongtan, S.; Fukngoen, P.; Peerajan, S.; Saelee, M.; Chaiyasut, K.; Kesika, P.; et al. Probiotics Supplementation Improves Intestinal Permeability, Obesity Index and Metabolic Biomarkers in Elderly Thai Subjects: A Randomized Controlled Trial. Foods 2022, 11, 268. [Google Scholar] [CrossRef] [PubMed]
- Horvath, A.; Leber, B.; Schmerboeck, B.; Tawdrous, M.; Zettel, G.; Hartl, A.; Madl, T.; Stryeck, S.; Fuchs, D.; Lemesch, S.; et al. Randomised clinical trial: The effects of a multispecies probiotic vs. placebo on innate immune function, bacterial translocation and gut permeability in patients with cirrhosis. Aliment. Pharmacol. Ther. 2016, 44, 926–935. [Google Scholar] [CrossRef]
- Wang, J.; Ke, H.; Liu, K.-X.; Qu, J.-M. Effects of exogenous probiotics on the gut microbiota and clinical outcomes in critically ill patients: A randomized controlled trial. Ann. Palliat. Med. 2021, 10, 1180–1190. [Google Scholar] [CrossRef]
- Stadlbauer, V.; Horvath, A.; Komarova, I.; Schmerboeck, B.; Feldbacher, N.; Klymiuk, I.; Durdevic, M.; Rainer, F.; Blesl, A.; Stiegler, P.; et al. Dysbiosis in early sepsis can be modulated by a multispecies probiotic: A randomised controlled pilot trial. Benef. Microbes 2019, 10, 265–278. [Google Scholar] [CrossRef]
- Li, S.; Yin, Y.; Xiao, D.; Zou, Y. Supplemental bifid triple viable capsule treatment improves inflammatory response and T cell frequency in ulcerative colitis patients. BMC Gastroenterol. 2021, 21, 314. [Google Scholar] [CrossRef] [PubMed]
- Kamarlı Altun, H.; Akal Yıldız, E.; Akın, M. Effects of synbiotic therapy in mild-to-moderately active ulcerative colitis: A randomized placebo-controlled study. Turk. J. Gastroenterol. 2019, 30, 313–320. [Google Scholar] [CrossRef]
- Bjarnason, I.; Sission, G.; Hayee, B. A randomised, double-blind, placebo-controlled trial of a multi-strain probiotic in patients with asymptomatic ulcerative colitis and Crohn’s disease. Inflammopharmacology 2019, 27, 465–473. [Google Scholar] [CrossRef]
- Kaźmierczak-Siedlecka, K.; Folwarski, M.; Skonieczna-Żydecka, K.; Ruszkowski, J.; Makarewicz, W. The use of Lactobacillus plantarum 299v (DSM 9843) in cancer patients receiving home enteral nutrition—Study protocol for a randomized, double-blind, and placebo-controlled trial. Nutr. J. 2020, 19, 98. [Google Scholar] [CrossRef]
- Cai, G.-S.; Su, H.; Zhang, J. Protective effect of probiotics in patients with non-alcoholic fatty liver disease. Medicine 2020, 99, e21464. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.B.; Jun, D.W.; Kang, B.-K.; Lim, J.H.; Lim, S.; Chung, M.-J. Randomized, Double-blind, Placebo-controlled Study of a Multispecies Probiotic Mixture in Nonalcoholic Fatty Liver Disease. Sci. Rep. 2019, 9, 5688. [Google Scholar] [CrossRef] [PubMed]
- Bakhshimoghaddam, F.; Shateri, K.; Sina, M.; Hashemian, M.; Alizadeh, M. Daily Consumption of Synbiotic Yogurt Decreases Liver Steatosis in Patients with Nonalcoholic Fatty Liver Disease: A Randomized Controlled Clinical Trial. J. Nutr. 2018, 148, 1276–1284. [Google Scholar] [CrossRef] [PubMed]
- Kobyliak, N.; Abenavoli, L.; Mykhalchyshyn, G.; Kononenko, L.; Boccuto, L.; Kyriienko, D.; Dynnyk, O. A Multi-strain Probiotic Reduces the Fatty Liver Index, Cytokines and Aminotransferase levels in NAFLD Patients: Evidence from a Randomized Clinical Trial. J. Gastrointestin. Liver Dis. 2018, 27, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Mohamad Nor, M.H.; Ayob, N.; Mokhtar, N.M.; Raja Ali, R.A.; Tan, G.C.; Wong, Z.; Shafiee, N.H.; Wong, Y.P.; Mustangin, M.; Nawawi, K.N.M. The Effect of Probiotics (MCP® BCMC® Strains) on Hepatic Steatosis, Small Intestinal Mucosal Immune Function, and Intestinal Barrier in Patients with Non-Alcoholic Fatty Liver Disease. Nutrients 2021, 13, 3192. [Google Scholar] [CrossRef]
- Han, M.-L.; Lee, M.-H.; Lee, W.-J.; Chen, S.-C.; Almalki, O.M.; Chen, J.-C.; Wu, C.-C. Probiotics for gallstone prevention in patients with bariatric surgery: A prospective randomized trial. Asian J. Surg. 2022, 45, 2664–2669. [Google Scholar] [CrossRef]
- Gao, F.; Guan, D.; Wang, G.; Zhang, L.; He, J.; Lv, W.; Zhang, X.; Tao, W.; Dai, Y.; Xu, S.; et al. Effects of oral tauroursodeoxycholic acid and/or intestinal probiotics on serum biochemical indexes and bile composition in patients with cholecystolithiasis. Front. Pharmacol. 2022, 13, 882764. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.-D.; Zhu, R.-X.; Bian, Z.-Z.; Sun, T.-W. Effect of probiotics on length of hospitalization in mild acute pancreatitis: A randomized, double-blind, placebo-controlled trial. World J. Gastroenterol. 2021, 27, 224–232. [Google Scholar] [CrossRef]
- Sharma, B.; Srivastava, S.; Singh, N.; Sachdev, V.; Kapur, S.; Saraya, A. Role of Probiotics on Gut Permeability and Endotoxemia in Patients With Acute Pancreatitis. J. Clin. Gastroenterol. 2011, 45, 442–448. [Google Scholar] [CrossRef]
- Savytska, M.; Kyriienko, D.; Komisarenko, I.; Kovalchuk, O.; Falalyeyeva, T.; Kobyliak, N. Probiotic for Pancreatic β-Cell Function in Type 2 Diabetes: A Randomized, Double-Blinded, Placebo-Controlled Clinical Trial. Diabetes Ther. 2023, 14, 1915–1931. [Google Scholar] [CrossRef]
- Rammohan, A.; Sathyanesan, J.; Rajendran, K.; Pitchaimuthu, A.; Perumal, S.K.; Balaraman, K.; Ramasamy, R.; Palaniappan, R.; Govindan, M. Synbiotics in Surgery for Chronic Pancreatitis. Ann. Surg. 2015, 262, 31–37. [Google Scholar] [CrossRef]
- dos Santos, P.Q.; Guedes, J.C.; de Jesus, R.P.; dos Santos, R.R.; Fiaconne, R.L. Effects of using symbiotics in the clinical nutritional evolution of patients with chronic pancreatitis: Study prospective, randomized, controlled, double blind. Clin. Nutr. ESPEN 2017, 18, 9–15. [Google Scholar] [CrossRef]
- Braga, V.L.; Rocha, L.P.D.S.; Bernardo, D.D.; Cruz, C.d.O.; Riera, R. What do Cochrane systematic reviews say about probiotics as preventive interventions? Sao Paulo Med. J. 2017, 135, 578–586. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, S.C.; Barbara, G.; Buurman, W.; Ockhuizen, T.; Schulzke, J.-D.; Serino, M.; Tilg, H.; Watson, A.; Wells, J.M. Intestinal permeability--a new target for disease prevention and therapy. BMC Gastroenterol. 2014, 14, 189. [Google Scholar] [CrossRef] [PubMed]
- Ganji-Arjenaki, M.; Rafieian-Kopaei, M. Probiotics are a good choice in remission of inflammatory bowel diseases: A meta analysis and systematic review. J. Cell. Physiol. 2018, 233, 2091–2103. [Google Scholar] [CrossRef] [PubMed]
- Vancamelbeke, M.; Vermeire, S. The intestinal barrier: A fundamental role in health and disease. Expert Rev. Gastroenterol. Hepatol. 2017, 11, 821–834. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Chen, X.; Liu, M.; Guo, X. Effect of three lactobacilli with strain-specific activities on the growth performance, faecal microbiota and ileum mucosa proteomics of piglets. J. Anim. Sci. Biotechnol. 2017, 8, 52. [Google Scholar] [CrossRef]
- Yang, G.-Y.; Yu, J.; Su, J.-H.; Jiao, L.-G.; Liu, X.; Zhu, Y.-H. Oral Administration of Lactobacillus rhamnosus GG Ameliorates Salmonella Infantis-Induced Inflammation in a Pig Model via Activation of the IL-22BP/IL-22/STAT3 Pathway. Front. Cell. Infect. Microbiol. 2017, 7, 323. [Google Scholar] [CrossRef]
- Rao, R.K.; Samak, G. Protection and Restitution of Gut Barrier by Probiotics: Nutritional and Clinical Implications. Curr. Nutr. Food Sci. 2013, 9, 99–107. [Google Scholar]
- Yi, H.; Wang, L.; Xiong, Y.; Wen, X.; Wang, Z.; Yang, X.; Gao, K.; Jiang, Z. Effects of Lactobacillus reuteri LR1 on the growth performance, intestinal morphology, and intestinal barrier function in weaned pigs. J. Anim. Sci. 2018, 96, 2342–2351. [Google Scholar] [CrossRef]
- Cui, Y.; Liu, L.; Dou, X.; Wang, C.; Zhang, W.; Gao, K.; Liu, J.; Wang, H. Lactobacillus reuteri ZJ617 maintains intestinal integrity via regulating tight junction, autophagy and apoptosis in mice challenged with lipopolysaccharide. Oncotarget 2017, 8, 77489–77499. [Google Scholar] [CrossRef]
- Kim, S.H.; Jeung, W.; Choi, I.-D.; Jeong, J.-W.; Lee, D.E.; Huh, C.-S.; Kim, G.-B.; Hong, S.S.; Shim, J.-J.; Lee, J.L.; et al. Lactic Acid Bacteria Improves Peyer’s Patch Cell-Mediated Immunoglobulin A and Tight-Junction Expression in a Destructed Gut Microbial Environment. J. Microbiol. Biotechnol. 2016, 26, 1035–1045. [Google Scholar] [CrossRef]
- Terciolo, C.; Dapoigny, M.; Andre, F. Beneficial effects of Saccharomyces boulardii CNCM I-745 on clinical disorders associated with intestinal barrier disruption. Clin. Exp. Gastroenterol. 2019, 12, 67–82. [Google Scholar] [CrossRef]
- Dukowicz, A.C.; Lacy, B.E.; Levine, G.M. Small intestinal bacterial overgrowth: A comprehensive review. Gastroenterol. Hepatol. (N. Y). 2007, 3, 112–122. [Google Scholar]
- Grace, E.; Shaw, C.; Whelan, K.; Andreyev, H.J.N. Review article: Small intestinal bacterial overgrowth—Prevalence, clinical features, current and developing diagnostic tests, and treatment. Aliment. Pharmacol. Ther. 2013, 38, 674–688. [Google Scholar] [CrossRef]
- Stanghellini, V. Functional Dyspepsia and Irritable Bowel Syndrome: Beyond Rome IV. Dig. Dis. 2017, 35, 14–17. [Google Scholar] [CrossRef]
- Principi, N.; Cozzali, R.; Farinelli, E.; Brusaferro, A.; Esposito, S. Gut dysbiosis and irritable bowel syndrome: The potential role of probiotics. J. Infect. 2018, 76, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Catinean, A.; Neag, A.M.; Nita, A.; Buzea, M.; Buzoianu, A.D. Bacillus spp. Spores—A Promising Treatment Option for Patients with Irritable Bowel Syndrome. Nutrients 2019, 11, 1968. [Google Scholar] [CrossRef] [PubMed]
- Schultz, M.; Veltkamp, C.; Dieleman, L.A.; Grenther, W.B.; Wyrick, P.B.; Tonkonogy, S.L.; Sartor, R.B. Lactobacillus plantarum 299V in the treatment and prevention of spontaneous colitis in interleukin-10-deficient mice. Inflamm. Bowel Dis. 2002, 8, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Fabia, R.; Ar’Rajab, A.; Johansson, M.L.; Willén, R.; Andersson, R.; Molin, G.; Bengmark, S. The effect of exogenous administration of Lactobacillus reuteri R2LC and oat fiber on acetic acid-induced colitis in the rat. Scand. J. Gastroenterol. 1993, 28, 155–162. [Google Scholar] [CrossRef]
- Isolauri, E.; Arvola, T.; Sütas, Y.; Moilanen, E.; Salminen, S. Probiotics in the management of atopic eczema. Clin. Exp. Allergy 2000, 30, 1604–1610. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Song, M.; Lu, X.; Zhu, X.; Deng, J. Gut microbes in gastrointestinal cancers. Semin. Cancer Biol. 2022, 86, 967–975. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-Y.; Chu, S.-H.; Jeon, J.Y.; Lee, M.-K.; Park, J.-H.; Lee, D.-C.; Lee, J.-W.; Kim, N.-K. Effects of 12 weeks of probiotic supplementation on quality of life in colorectal cancer survivors: A double-blind, randomized, placebo-controlled trial. Dig. Liver Dis. 2014, 46, 1126–1132. [Google Scholar] [CrossRef] [PubMed]
- Mörkl, S.; Lackner, S.; Meinitzer, A.; Mangge, H.; Lehofer, M.; Halwachs, B.; Gorkiewicz, G.; Kashofer, K.; Painold, A.; Holl, A.K.; et al. Gut microbiota, dietary intakes and intestinal permeability reflected by serum zonulin in women. Eur. J. Nutr. 2018, 57, 2985–2997. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.-H.; Huang, M.-J.; Zhang, X.-W.; Wang, L.; Huang, N.-Q.; Peng, H.; Lan, P.; Peng, J.-S.; Yang, Z.; Xia, Y.; et al. The effects of perioperative probiotic treatment on serum zonulin concentration and subsequent postoperative infectious complications after colorectal cancer surgery: A double-center and double-blind randomized clinical trial. Am. J. Clin. Nutr. 2013, 97, 117–126. [Google Scholar] [CrossRef]
- Österlund, P.; Ruotsalainen, T.; Korpela, R.; Saxelin, M.; Ollus, A.; Valta, P.; Kouri, M.; Elomaa, I.; Joensuu, H. Lactobacillus supplementation for diarrhoea related to chemotherapy of colorectal cancer: A randomised study. Br. J. Cancer 2007, 97, 1028–1034. [Google Scholar] [CrossRef]
- Aisu, N.; Tanimura, S.; Yamashita, Y.; Yamashita, K.; Maki, K.; Yoshida, Y.; Sasaki, T.; Takeno, S.; Hoshino, S. Impact of perioperative probiotic treatment for surgical site infections in patients with colorectal cancer. Exp. Ther. Med. 2015, 10, 966–972. [Google Scholar] [CrossRef]
- Gao, Z.; Guo, B.; Gao, R.; Zhu, Q.; Wu, W.; Qin, H. Probiotics modify human intestinal mucosa-associated microbiota in patients with colorectal cancer. Mol. Med. Rep. 2015, 12, 6119–6127. [Google Scholar] [CrossRef]
- Kotzampassi, K.; Stavrou, G.; Damoraki, G.; Georgitsi, M.; Basdanis, G.; Tsaousi, G.; Giamarellos-Bourboulis, E.J. A Four-Probiotics Regimen Reduces Postoperative Complications After Colorectal Surgery: A Randomized, Double-Blind, Placebo-Controlled Study. World J. Surg. 2015, 39, 2776–2783. [Google Scholar] [CrossRef]
- Meroni, M.; Longo, M.; Dongiovanni, P. The Role of Probiotics in Nonalcoholic Fatty Liver Disease: A New Insight into Therapeutic Strategies. Nutrients 2019, 11, 2642. [Google Scholar] [CrossRef]
- Vajro, P.; Mandato, C.; Licenziati, M.R.; Franzese, A.; Vitale, D.F.; Lenta, S.; Caropreso, M.; Vallone, G.; Meli, R. Effects of Lactobacillus rhamnosus strain GG in pediatric obesity-related liver disease. J. Pediatr. Gastroenterol. Nutr. 2011, 52, 740–743. [Google Scholar] [CrossRef]
- Abdel Monem, S.M. Probiotic Therapy in Patients with Nonalcoholic Steatohepatitis in Zagazig University Hospitals. Euroasian J. Hepato-Gastroenterol. 2017, 7, 101–106. [Google Scholar] [CrossRef]
- Wong, V.W.S.; Wong, G.L.H.; Chim, A.M.L.; Chu, W.C.W.; Yeung, D.K.W.; Li, K.C.T.; Chan, H.L.Y. Treatment of nonalcoholic steatohepatitis with probiotics. A proof-of-concept study. Ann. Hepatol. 2013, 12, 256–262. [Google Scholar] [CrossRef]
- Ma, Y.-Y.; Li, L.; Yu, C.-H.; Shen, Z.; Chen, L.-H.; Li, Y.-M. Effects of probiotics on nonalcoholic fatty liver disease: A meta-analysis. World J. Gastroenterol. 2013, 19, 6911–6918. [Google Scholar] [CrossRef]
- Gao, X.; Zhu, Y.; Wen, Y.; Liu, G.; Wan, C. Efficacy of probiotics in non-alcoholic fatty liver disease in adult and children: A meta-analysis of randomized controlled trials. Hepatol. Res. 2016, 46, 1226–1233. [Google Scholar] [CrossRef]
- Meroni, M.; Longo, M.; Dongiovanni, P. Alcohol or Gut Microbiota: Who Is the Guilty? Int. J. Mol. Sci. 2019, 20, 4568. [Google Scholar] [CrossRef]
- Loguercio, C.; Federico, A.; Tuccillo, C.; Terracciano, F.; D’Auria, M.V.; De Simone, C.; Blanco, C.D.V. Beneficial Effects of a Probiotic VSL#3 on Parameters of Liver Dysfunction in Chronic Liver Diseases. J. Clin. Gastroenterol. 2005, 39, 540–543. [Google Scholar] [PubMed]
- Malaguarnera, M.; Vacante, M.; Antic, T.; Giordano, M.; Chisari, G.; Acquaviva, R.; Mastrojeni, S.; Malaguarnera, G.; Mistretta, A.; Li Volti, G.; et al. Bifidobacterium longum with Fructo-Oligosaccharides in Patients with Non Alcoholic Steatohepatitis. Dig. Dis. Sci. 2012, 57, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Acalovschi, M. Gallstones in patients with liver cirrhosis: Incidence, etiology, clinical and therapeutical aspects. World J. Gastroenterol. 2014, 20, 7277–7285. [Google Scholar] [CrossRef] [PubMed]
- Castro-Torres, I.G.; Cárdenas-Vázquez, R.d.J.; Velázquez-González, C.; Ventura-Martínez, R.; De la O-Arciniega, M.; Naranjo-Rodríguez, E.B.; Martínez-Vázquez, M. Future therapeutic targets for the treatment and prevention of cholesterol gallstones. Eur. J. Pharmacol. 2015, 765, 366–374. [Google Scholar] [CrossRef]
- Sato, S.; Nagai, H.; Igarashi, Y. Effect of Probiotics on Serum Bile Acids in Patients with Ulcerative Colitis. Hepatogastroenterology 2011, 59, 1804–1808. [Google Scholar]
- Takeda, Y.; Itoh, H.; Kobashi, K. Effect of Clostridium butyricum on the formation and dissolution of gallstones in experimental cholesterol cholelithiasis. Life Sci. 1983, 32, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Hosomi, M.; Tanida, N.; Shimoyama, T. The role of intestinal bacteria in gallstone formation in animal model. A study on biliary lipid composition and bile acid profiles in bile, small intestinal contents and feces of Clostridium butyricum MIYAIRI No. 588 monocontaminated mice. Gastroenterol. Jpn. 1982, 17, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Horáčková, Š.; Plocková, M.; Demnerová, K. Importance of microbial defence systems to bile salts and mechanisms of serum cholesterol reduction. Biotechnol. Adv. 2018, 36, 682–690. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Guo, M.-J.; Gao, Q.; Yang, J.-F.; Yang, L.; Pang, X.-L.; Jiang, X.-J. The effects of probiotics on total cholesterol. Medicine 2018, 97, e9679. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.L.; Martoni, C.J.; Parent, M.; Prakash, S. Cholesterol-lowering efficacy of a microencapsulated bile salt hydrolase-active Lactobacillus reuteri NCIMB 30242 yoghurt formulation in hypercholesterolaemic adults. Br. J. Nutr. 2012, 107, 1505–1513. [Google Scholar] [CrossRef] [PubMed]
- Moschetta, A.; Bookout, A.L.; Mangelsdorf, D.J. Prevention of cholesterol gallstone disease by FXR agonists in a mouse model. Nat. Med. 2004, 10, 1352–1358. [Google Scholar] [CrossRef] [PubMed]
- Wahlström, A.; Kovatcheva-Datchary, P.; Ståhlman, M.; Bäckhed, F.; Marschall, H.-U. Crosstalk between Bile Acids and Gut Microbiota and Its Impact on Farnesoid X Receptor Signalling. Dig. Dis. 2017, 35, 246–250. [Google Scholar] [CrossRef]
- Devkota, S.; Chang, E.B. Interactions between Diet, Bile Acid Metabolism, Gut Microbiota, and Inflammatory Bowel Diseases. Dig. Dis. 2015, 33, 351–356. [Google Scholar] [CrossRef]
- Oh, J.K.; Kim, Y.R.; Lee, B.; Choi, Y.M.; Kim, S.H. Prevention of Cholesterol Gallstone Formation by Lactobacillus acidophilus ATCC 43121 and Lactobacillus fermentum MF27 in Lithogenic Diet-Induced Mice. Food Sci. Anim. Resour. 2021, 41, 343–352. [Google Scholar] [CrossRef]
- Tenner, S.; Baillie, J.; DeWitt, J.; Vege, S.S. American College of Gastroenterology American College of Gastroenterology guideline: Management of acute pancreatitis. Am. J. Gastroenterol. 2013, 108, 1400–1415. [Google Scholar] [CrossRef]
- Ko, J.-S.; Yang, H.-R.; Chang, J.-Y.; Seo, J.-K. Lactobacillus plantarum inhibits epithelial barrier dysfunction and interleukin-8 secretion induced by tumor necrosis factor-alpha. World J. Gastroenterol. 2007, 13, 1962–1965. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.-J.; Gao, C.-F.; Wei, D.; Wang, C.; Ding, S.-Q. Acute pancreatitis: Etiology and common pathogenesis. World J. Gastroenterol. 2009, 15, 1427–1430. [Google Scholar] [CrossRef]
- Dervenis, C.; Hatzitheoklitos, E.; Smailis, D. Bacterial translocation and its prevention in acute pancreatitis. J. Hepatobiliary Pancreat. Surg. 2003, 10, 415–418. [Google Scholar] [CrossRef] [PubMed]
- Peery, A.F.; Dellon, E.S.; Lund, J.; Crockett, S.D.; McGowan, C.E.; Bulsiewicz, W.J.; Gangarosa, L.M.; Thiny, M.T.; Stizenberg, K.; Morgan, D.R.; et al. Burden of Gastrointestinal Disease in the United States: 2012 Update. Gastroenterology 2012, 143, 1179–1187.e3. [Google Scholar] [CrossRef]
- Singhal, B.; Mukherjee, A.; Srivastav, S. Role of Probiotics in Pancreatic Cancer Prevention: The Prospects and Challenges. Adv. Biosci. Biotechnol. 2016, 07, 468–500. [Google Scholar] [CrossRef]
- Muftuoglu, M.A.T.; Isikgor, S.; Tosun, S.; Saglam, A. Effects of probiotics on the severity of experimental acute pancreatitis. Eur. J. Clin. Nutr. 2006, 60, 464–468. [Google Scholar] [CrossRef]
- van Minnen, L.P.; Timmerman, H.M.; Lutgendorff, F.; Verheem, A.; Harmsen, W.; Konstantinov, S.R.; Smidt, H.; Visser, M.R.; Rijkers, G.T.; Gooszen, H.G.; et al. Modification of intestinal flora with multispecies probiotics reduces bacterial translocation and improves clinical course in a rat model of acute pancreatitis. Surgery 2007, 141, 470–480. [Google Scholar] [CrossRef]
- Oláh, A.; Belágyi, T.; Pótó, L.; Romics, L.; Bengmark, S. Synbiotic control of inflammation and infection in severe acute pancreatitis: A prospective, randomized, double blind study. Hepatogastroenterology 2007, 54, 590–594. [Google Scholar]
- Oláh, A.; Romics, L., Jr. Early enteral nutrition in acute pancreatitis—Benefits and limitations. Langenbeck’s Arch. Surg. 2008, 393, 261–269. [Google Scholar] [CrossRef]
- Mangiante, G.; Colucci, G.; Canepari, P.; Bassi, C.; Nicoli, N.; Casaril, A.; Marinello, P.; Signoretto, C.; Bengmark, S. Lactobacillus plantarum reduces infection of pancreatic necrosis in experimental acute pancreatitis. Dig. Surg. 2001, 18, 47–50. [Google Scholar] [CrossRef] [PubMed]
- Lutgendorff, F.; Trulsson, L.M.; van Minnen, L.P.; Rijkers, G.T.; Timmerman, H.M.; Franzén, L.E.; Gooszen, H.G.; Akkermans, L.M.A.; Söderholm, J.D.; Sandström, P.A. Probiotics enhance pancreatic glutathione biosynthesis and reduce oxidative stress in experimental acute pancreatitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 295, G1111–G1121. [Google Scholar] [CrossRef]
- Rohith, G.; Sureshkumar, S.; Anandhi, A.; Kate, V.; Rajesh, B.S.; Abdulbasith, K.M.; Nanda, N.; Palanivel, C.; Vijayakumar, C. Effect of Synbiotics in Reducing the Systemic Inflammatory Response and Septic Complications in Moderately Severe and Severe Acute Pancreatitis: A Prospective Parallel-Arm Double-Blind Randomized Trial. Dig. Dis. Sci. 2023, 68, 969–977. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Zhang, Y.; Yang, Q.; Lee, P.; Windsor, J.A.; Wu, D. An Updated Systematic Review With Meta-analysis: Efficacy of Prebiotic, Probiotic, and Synbiotic Treatment of Patients With Severe Acute Pancreatitis. Pancreas 2021, 50, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Felton, J.S.; Knize, M.G.; Wu, R.W.; Colvin, M.E.; Hatch, F.T.; Malfatti, M.A. Mutagenic potency of food-derived heterocyclic amines. Mutat. Res. Mol. Mech. Mutagen. 2007, 616, 90–94. [Google Scholar] [CrossRef]
- Turesky, R.J. Formation and biochemistry of carcinogenic heterocyclic aromatic amines in cooked meats. Toxicol. Lett. 2007, 168, 219–227. [Google Scholar] [CrossRef]
- Sugimura, T.; Wakabayashi, K.; Nakagama, H.; Nagao, M. Heterocyclic amines: Mutagens/carcinogens produced during cooking of meat and fish. Cancer Sci. 2004, 95, 290–299. [Google Scholar] [CrossRef]
- Kumar, M.; Kumar, A.; Nagpal, R.; Mohania, D.; Behare, P.; Verma, V.; Kumar, P.; Poddar, D.; Aggarwal, P.K.; Henry, C.J.K.; et al. Cancer-preventing attributes of probiotics: An update. Int. J. Food Sci. Nutr. 2010, 61, 473–496. [Google Scholar] [CrossRef]
- Orrhage, K.M.; Annas, A.; Nord, C.E.; Brittebo, E.B.; Rafter, J.J. Effects of Lactic Acid Bacteria on the Uptake and Distribution of the Food Mutagen Trp-P-2 in Mice. Scand. J. Gastroenterol. 2002, 37, 215–221. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, L.; Zhang, J.; Li, Y.; He, Q.; Li, H.; Guo, X.; Guo, J.; Zhang, H. Probiotic Lactobacillus casei Zhang ameliorates high-fructose-induced impaired glucose tolerance in hyperinsulinemia rats. Eur. J. Nutr. 2014, 53, 221–232. [Google Scholar] [CrossRef]
- Rhee, C.-H.; Park, H.-D. Three Glycoproteins with Antimutagenic Activity Identified in Lactobacillus plantarum KLAB21. Appl. Environ. Microbiol. 2001, 67, 3445–3449. [Google Scholar] [CrossRef] [PubMed]
- Niderkorn, V.; Boudra, H.; Morgavi, D.P. Binding of Fusarium mycotoxins by fermentative bacteria in vitro. J. Appl. Microbiol. 2006, 101, 849–856. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, F.; Halttunen, T.; Tahvonen, R.; Salminen, S. Probiotic bacteria as potential detoxification tools: Assessing their heavy metal binding isotherms. Can. J. Microbiol. 2006, 52, 877–885. [Google Scholar] [CrossRef]
- Halttunen, T.; Collado, M.C.; El-Nezami, H.; Meriluoto, J.; Salminen, S. Combining strains of lactic acid bacteria may reduce their toxin and heavy metal removal efficiency from aqueous solution. Lett. Appl. Microbiol. 2007, 46, 160–165. [Google Scholar] [CrossRef]
- El-Nezami, H.S.; Chrevatidis, A.; Auriola, S.; Salminen, S.; Mykkänen, H. Removal of common Fusarium toxins in vitro by strains of Lactobacillus and Propionibacterium. Food Addit. Contam. 2002, 19, 680–686. [Google Scholar] [CrossRef] [PubMed]
- Lahtinen, S.J.; Haskard, C.A.; Ouwehand, A.C.; Salminen, S.J.; Ahokas, J.T. Binding of aflatoxin B 1 to cell wall components of Lactobacillus rhamnosus strain GG. Food Addit. Contam. 2004, 21, 158–164. [Google Scholar] [CrossRef]
- Bousquet, J.; Khaltaev, N.; Cruz, A.A.; Denburg, J.; Fokkens, W.J.; Togias, A.; Zuberbier, T.; Baena-Cagnani, C.E.; Canonica, G.W.; van Weel, C.; et al. Allergic Rhinitis and its Impact on Asthma (ARIA) 2008 update (in collaboration with the World Health Organization, GA(2)LEN and AllerGen). Allergy 2008, 63 (Suppl. S8), 8–160. [Google Scholar] [CrossRef]
- Fokkens, W.J.; Lund, V.J.; Hopkins, C.; Hellings, P.W.; Kern, R.; Reitsma, S.; Toppila-Salmi, S.; Bernal-Sprekelsen, M.; Mullol, J.; Alobid, I.; et al. European Position Paper on Rhinosinusitis and Nasal Polyps 2020. Rhinology 2020, 58, 1–464. [Google Scholar] [CrossRef]
- De Boeck, I.; van den Broek, M.F.L.; Allonsius, C.N.; Spacova, I.; Wittouck, S.; Martens, K.; Wuyts, S.; Cauwenberghs, E.; Jokicevic, K.; Vandenheuvel, D.; et al. Lactobacilli Have a Niche in the Human Nose. Cell Rep. 2020, 31, 107674. [Google Scholar] [CrossRef]
- Gan, W.; Yang, F.; Tang, Y.; Zhou, D.; Qing, D.; Hu, J.; Liu, S.; Liu, F.; Meng, J. The difference in nasal bacterial microbiome diversity between chronic rhinosinusitis patients with polyps and a control population. Int. Forum Allergy Rhinol. 2019, 9, 582–592. [Google Scholar] [CrossRef]
- Stearns, J.C.; Davidson, C.J.; McKeon, S.; Whelan, F.J.; Fontes, M.E.; Schryvers, A.B.; Bowdish, D.M.E.; Kellner, J.D.; Surette, M.G. Culture and molecular-based profiles show shifts in bacterial communities of the upper respiratory tract that occur with age. ISME J. 2015, 9, 1246–1259. [Google Scholar] [CrossRef]
- Ling, Z.; Liu, X.; Luo, Y.; Yuan, L.; Nelson, K.E.; Wang, Y.; Xiang, C.; Li, L. Pyrosequencing analysis of the human microbiota of healthy Chinese undergraduates. BMC Genom. 2013, 14, 390. [Google Scholar] [CrossRef]
- Jensen, A.; Fagö-Olsen, H.; Sørensen, C.H.; Kilian, M. Molecular mapping to species level of the tonsillar crypt microbiota associated with health and recurrent tonsillitis. PLoS ONE 2013, 8, e56418. [Google Scholar] [CrossRef]
- Abreu, N.A.; Nagalingam, N.A.; Song, Y.; Roediger, F.C.; Pletcher, S.D.; Goldberg, A.N.; Lynch, S. V Sinus microbiome diversity depletion and Corynebacterium tuberculostearicum enrichment mediates rhinosinusitis. Sci. Transl. Med. 2012, 4, 151ra124. [Google Scholar] [CrossRef]
- Hasegawa, K.; Linnemann, R.W.; Mansbach, J.M.; Ajami, N.J.; Espinola, J.A.; Petrosino, J.F.; Piedra, P.A.; Stevenson, M.D.; Sullivan, A.F.; Thompson, A.D.; et al. Nasal Airway Microbiota Profile and Severe Bronchiolitis in Infants: A Case-control Study. Pediatr. Infect. Dis. J. 2017, 36, 1044–1051. [Google Scholar] [CrossRef]
- Biesbroek, G.; Tsivtsivadze, E.; Sanders, E.A.M.; Montijn, R.; Veenhoven, R.H.; Keijser, B.J.F.; Bogaert, D. Early respiratory microbiota composition determines bacterial succession patterns and respiratory health in children. Am. J. Respir. Crit. Care Med. 2014, 190, 1283–1292. [Google Scholar] [CrossRef] [PubMed]
- De Boeck, I.; Wittouck, S.; Martens, K.; Claes, J.; Jorissen, M.; Steelant, B.; van den Broek, M.F.L.; Seys, S.F.; Hellings, P.W.; Vanderveken, O.M.; et al. Anterior Nares Diversity and Pathobionts Represent Sinus Microbiome in Chronic Rhinosinusitis. mSphere 2019, 4, e00532-19. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-S.; Jan, R.-L.; Lin, Y.-L.; Chen, H.-H.; Wang, J.-Y. Randomized placebo-controlled trial of Lactobacillus on asthmatic children with allergic rhinitis. Pediatr. Pulmonol. 2010, 45, 1111–1120. [Google Scholar] [CrossRef] [PubMed]
- Gutkowski, P.; Madaliński, K.; Grek, M.; Dmeńska, H.; Syczewska, M.; Michałkiewicz, J. Clinical immunology Effect of orally administered probiotic strains Lactobacillus and Bifidobacterium in children with atopic asthma. Cent. Eur. J. Immunol. 2010, 35, 233–238. [Google Scholar]
- van de Pol, M.A.; Lutter, R.; Smids, B.S.; Weersink, E.J.M.; van der Zee, J.S. Synbiotics reduce allergen-induced T-helper 2 response and improve peak expiratory flow in allergic asthmatics. Allergy 2011, 66, 39–47. [Google Scholar] [CrossRef]
- Dehnavi, S.; Azad, F.J.; Hoseini, R.F.; Moazzen, N.; Tavakkol-Afshari, J.; Nikpoor, A.R.; Salmani, A.A.; Ahanchian, H.; Mohammadi, M. A significant decrease in the gene expression of interleukin-17 following the administration of synbiotic in patients with allergic rhinitis who underwent immunotherapy: A placebo-controlled clinical trial. J. Res. Med. Sci. 2019, 24, 51. [Google Scholar] [CrossRef]
- Miraglia Del Giudice, M.; Indolfi, C.; Capasso, M.; Maiello, N.; Decimo, F.; Ciprandi, G. Bifidobacterium mixture (B longum BB536, B infantis M-63, B breve M-16V) treatment in children with seasonal allergic rhinitis and intermittent asthma. Ital. J. Pediatr. 2017, 43, 25. [Google Scholar] [CrossRef]
- Wardani, A.T.W.; Wiratno; Fatmawati, D. Potential Immuno-modulatory activity of Probiotics containing Lactobacillusacidophilus and Lacobacilluscasei to increase the ratio of IFN γ/IL-4 in patients with Allergic Rhinitis. Bangladesh J. Med. Sci. 2018, 18, 42–45. [Google Scholar] [CrossRef]
- Singh, A.; Hacini-Rachinel, F.; Gosoniu, M.L.; Bourdeau, T.; Holvoet, S.; Doucet-Ladeveze, R.; Beaumont, M.; Mercenier, A.; Nutten, S. Immune-modulatory effect of probiotic Bifidobacterium lactis NCC2818 in individuals suffering from seasonal allergic rhinitis to grass pollen: An exploratory, randomized, placebo-controlled clinical trial. Eur. J. Clin. Nutr. 2013, 67, 161–167. [Google Scholar] [CrossRef]
- Ahmed, M.; Billoo, A.G.; Iqbal, K. Efficacy of probiotic in perennial allergic rhinitis under five year children: A randomized controlled trial. Pak. J. Med. Sci. 2019, 35, 1538–1543. [Google Scholar] [CrossRef] [PubMed]
- Drago, L.; Cioffi, L.; Giuliano, M.; Pane, M.; Amoruso, A.; Schiavetti, I.; Reid, G.; Ciprandi, G.; PROPAM Study Group. The Probiotics in Pediatric Asthma Management (PROPAM) Study in the Primary Care Setting: A Randomized, Controlled, Double-Blind Trial with Ligilactobacillus salivarius LS01 (DSM 22775) and Bifidobacterium breve B632 (DSM 24706). J. Immunol. Res. 2022, 2022, 3837418. [Google Scholar] [CrossRef] [PubMed]
- Sadrifar, S.; Abbasi-Dokht, T.; Forouzandeh, S.; Malek, F.; Yousefi, B.; Salek Farrokhi, A.; Karami, J.; Baharlou, R. Immunomodulatory effects of probiotic supplementation in patients with asthma: A randomized, double-blind, placebo-controlled trial. Allergy Asthma Clin. Immunol. 2023, 19, 1. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-C.; Yang, Y.-H.; Chuang, S.-Y.; Huang, S.-Y.; Pan, W.-H. Reduced medication use and improved pulmonary function with supplements containing vegetable and fruit concentrate, fish oil and probiotics in asthmatic school children: A randomised controlled trial. Br. J. Nutr. 2013, 110, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Ahanchian, H.; Jafari, S.A.; Ansari, E.; Ganji, T.; Kiani, M.A.; Khalesi, M.; Momen, T.; Kianifar, H. A multi-strain Synbiotic may reduce viral respiratory infections in asthmatic children: A randomized controlled trial. Electron. Physician 2016, 8, 2833–2839. [Google Scholar] [CrossRef] [PubMed]
- Hassanzad, M.; Maleki Mostashari, K.; Ghaffaripour, H.; Emami, H.; Rahimi Limouei, S.; Velayati, A.A. Synbiotics and Treatment of Asthma: A Double-Blinded, Randomized, Placebo-Controlled Clinical Trial. Galen Med. J. 2019, 8, e1350. [Google Scholar] [CrossRef] [PubMed]
- Takimoto, T.; Hatanaka, M.; Hoshino, T.; Takara, T.; Tanaka, K.; Shimizu, A.; Morita, H.; Nakamura, T. Effect of Bacillus subtilis C-3102 on bone mineral density in healthy postmenopausal Japanese women: A randomized, placebo-controlled, double-blind clinical trial. Biosci. Microbiota Food Health 2018, 37, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Jafarnejad, S.; Djafarian, K.; Fazeli, M.R.; Yekaninejad, M.S.; Rostamian, A.; Keshavarz, S.A. Effects of a Multispecies Probiotic Supplement on Bone Health in Osteopenic Postmenopausal Women: A Randomized, Double-blind, Controlled Trial. J. Am. Coll. Nutr. 2017, 36, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Jansson, P.-A.; Curiac, D.; Lazou Ahrén, I.; Hansson, F.; Martinsson Niskanen, T.; Sjögren, K.; Ohlsson, C. Probiotic treatment using a mix of three Lactobacillus strains for lumbar spine bone loss in postmenopausal women: A randomised, double-blind, placebo-controlled, multicentre trial. Lancet Rheumatol. 2019, 1, e154–e162. [Google Scholar] [CrossRef] [PubMed]
- de Araújo, É.M.R.; Meneses, G.C.; Carioca, A.A.F.; Martins, A.M.C.; Daher, E.D.F.; da Silva Junior, G.B. Use of probiotics in patients with chronic kidney disease on hemodialysis: A randomized clinical trial. Braz. J. Nephrol. 2023, 45, 152–161. [Google Scholar] [CrossRef]
- Lim, P.S.; Wang, H.F.; Lee, M.C.; Chiu, L.-S.; Wu, M.-Y.; Chang, W.-C.; Wu, T.K. The Efficacy of Lactobacillus-Containing Probiotic Supplementation in Hemodialysis Patients: A Randomized, Double-Blind, Placebo-Controlled Trial. J. Ren. Nutr. 2021, 31, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Chávez-Íñiguez, J.S.; Ibarra-Estrada, M.; Gallardo-González, A.M.; Cisneros-Hernández, A.; Granado, R.C.-D.; Chávez-Alonso, G.; Hernández-Barajas, E.M.; Romero-Muñoz, A.C.; Ramos-Avellaneda, F.; Prieto-Magallanes, M.L.; et al. Probiotics in septic acute kidney injury, a double blind, randomized control trial. Ren. Fail. 2023, 45, 2260003. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Yang, L.; Dai, B.; Lin, B.; Lin, S.; Lin, E. Effects of Probiotics on Malnutrition and Health-Related Quality of Life in Patients Undergoing Peritoneal Dialysis: A Randomized Controlled Trial. J. Ren. Nutr. 2021, 31, 199–205. [Google Scholar] [CrossRef]
- Simeoni, M.; Citraro, M.L.; Cerantonio, A.; Deodato, F.; Provenzano, M.; Cianfrone, P.; Capria, M.; Corrado, S.; Libri, E.; Comi, A.; et al. An open-label, randomized, placebo-controlled study on the effectiveness of a novel probiotics administration protocol (ProbiotiCKD) in patients with mild renal insufficiency (stage 3a of CKD). Eur. J. Nutr. 2019, 58, 2145–2156. [Google Scholar] [CrossRef]
- Mitrović, M.; Stanković-Popović, V.; Tolinački, M.; Golić, N.; Soković Bajić, S.; Veljović, K.; Nastasijević, B.; Soldatović, I.; Svorcan, P.; Dimković, N. The Impact of Synbiotic Treatment on the Levels of Gut-Derived Uremic Toxins, Inflammation, and Gut Microbiome of Chronic Kidney Disease Patients—A Randomized Trial. J. Ren. Nutr. 2023, 33, 278–288. [Google Scholar] [CrossRef]
- Treewatchareekorn, S.; Tungsanga, S. WCN23-0326 effect of Lactobacillus rhamnosus GG on gut-derived uremic toxin and gut microbiome in non-dialysis chronic kidney disease patients: A randomized controlled trial. Kidney Int. Rep. 2023, 8, S211–S212. [Google Scholar] [CrossRef]
- Jiang, H.; Zhang, Y.; Xu, D.; Wang, Q. Probiotics ameliorates glycemic control of patients with diabetic nephropathy: A randomized clinical study. J. Clin. Lab. Anal. 2021, 35, e23650. [Google Scholar] [CrossRef] [PubMed]
- Haghighat, N.; Mohammadshahi, M.; Shayanpour, S.; Haghighizadeh, M.H.; Rahmdel, S.; Rajaei, M. The Effect of Synbiotic and Probiotic Supplementation on Mental Health Parameters in Patients Undergoing Hemodialysis: A Double-blind, Randomized, Placebo-controlled Trial. Indian J. Nephrol. 2021, 31, 149–156. [Google Scholar]
- Liu, S.; Liu, H.; Chen, L.; Liang, S.-S.; Shi, K.; Meng, W.; Xue, J.; He, Q.; Jiang, H. Effect of probiotics on the intestinal microbiota of hemodialysis patients: A randomized trial. Eur. J. Nutr. 2020, 59, 3755–3766. [Google Scholar] [CrossRef]
- Haghighat, N.; Mohammadshahi, M.; Shayanpour, S.; Haghighizadeh, M.H. Effects of Synbiotics and Probiotics Supplementation on Serum Levels of Endotoxin, Heat Shock Protein 70 Antibodies and Inflammatory Markers in Hemodialysis Patients: A Randomized Double-Blinded Controlled Trial. Probiotics Antimicrob. Proteins 2020, 12, 144–151. [Google Scholar] [CrossRef]
- Moludi, J.; Saiedi, S.; Ebrahimi, B.; Alizadeh, M.; Khajebishak, Y.; Ghadimi, S.S. Probiotics Supplementation on Cardiac Remodeling Following Myocardial Infarction: A Single-Center Double-Blind Clinical Study. J. Cardiovasc. Transl. Res. 2021, 14, 299–307. [Google Scholar] [CrossRef]
- Pourrajab, B.; Naderi, N.; Janani, L.; Mofid, V.; Hajahmadi, M.; Dehnad, A.; Shidfar, F. Comparison of probiotic yogurt and ordinary yogurt consumption on serum Pentraxin3, NT-proBNP, oxLDL, and ApoB100 in patients with chronic heart failure: A randomized, triple-blind, controlled trial. Food Funct. 2020, 11, 10000–10010. [Google Scholar] [CrossRef]
- Moludi, J.; Alizadeh, M.; Behrooz, M.; Maleki, V.; Seyed Mohammadzad, M.H.; Golmohammadi, A. Interactive Effect of Probiotics Supplementation and Weight Loss Diet on Metabolic Syndrome Features in Patients With Coronary Artery Diseases: A Double-Blind, Placebo-Controlled, Randomized Clinical Trial. Am. J. Lifestyle Med. 2021, 15, 653–663. [Google Scholar] [CrossRef]
- Shoaei Matin, S.; Shidfar, F.; Naderi, N.; Amin, A.; Hosseini-Baharanchi, F.S.; Dehnad, A. The Effect of Synbiotic Consumption on Serum NTproBNP, hsCRP and Blood Pressure in Patients with Chronic Heart Failure: A Randomized, Triple-Blind, Controlled Trial. Front. Nutr. 2022, 8, 822498. [Google Scholar] [CrossRef]
- Moludi, J.; Khedmatgozar, H.; Nachvak, S.M.; Abdollahzad, H.; Moradinazar, M.; Sadeghpour Tabaei, A. The effects of co-administration of probiotics and prebiotics on chronic inflammation, and depression symptoms in patients with coronary artery diseases: A randomized clinical trial. Nutr. Neurosci. 2022, 25, 1659–1668. [Google Scholar] [CrossRef] [PubMed]
- Romão da Silva, L.d.F.; de Oliveira, Y.; de Souza, E.L.; de Luna Freire, M.O.; Braga, V. de A.; Magnani, M.; de Brito Alves, J.L. Effects of probiotic therapy on cardio-metabolic parameters and autonomic modulation in hypertensive women: A randomized, triple-blind, placebo-controlled trial. Food Funct. 2020, 11, 7152–7163. [Google Scholar] [CrossRef]
- Moludi, J.; Kafil, H.S.; Qaisar, S.A.; Gholizadeh, P.; Alizadeh, M.; Vayghyan, H.J. Effect of probiotic supplementation along with calorie restriction on metabolic endotoxemia, and inflammation markers in coronary artery disease patients: A double blind placebo controlled randomized clinical trial. Nutr. J. 2021, 20, 47. [Google Scholar] [CrossRef] [PubMed]
- Cai, T.; Gallelli, L.; Cione, E.; Perletti, G.; Ciarleglio, F.; Malossini, G.; De Pretis, G.; Palmieri, A.; Mirone, V.; Bartoletti, R.; et al. The use of Lactobacillus casei DG® prevents symptomatic episodes and reduces the antibiotic use in patients affected by chronic bacterial prostatitis: Results from a phase IV study. World J. Urol. 2021, 39, 3433–3440. [Google Scholar] [CrossRef] [PubMed]
- Manfredi, C.; Calace, F.P.; Fusco, F.; Quattrone, C.; Giordano, D.; Crocetto, F.; Creta, M.; De Sio, M.; Arcaniolo, D. Escherichia coli Nissle 1917 as adjuvant therapy in patients with chronic bacterial prostatitis: A non-blinded, randomized, controlled trial. World J. Urol. 2021, 39, 4373–4379. [Google Scholar] [CrossRef]
- Abbasi, B.; Abbasi, H.; Niroumand, H. Synbiotic (FamiLact) administration in idiopathic male infertility enhances sperm quality, DNA integrity, and chromatin status: A triple-blinded randomized clinical trial. Int. J. Reprod. Biomed. 2021, 19, 235–244. [Google Scholar] [CrossRef]
- Asadi, M.; Gholipour, F.; Rahavian, A.; Javanbakht, M. Effects of probiotic supplementation on semen parameters after varicocelectomy: A randomized controlled trial. J. Res. Med. Sci. 2023, 28, 74. [Google Scholar]
- Park, S.-H.; Lee, E.S.; Park, S.T.; Jeong, S.Y.; Yun, Y.; Kim, Y.; Jeong, Y.; Kang, C.-H.; Choi, H.J. Efficacy and Safety of MED-01 Probiotics on Vaginal Health: A 12-Week, Multicenter, Randomized, Double-Blind, Placebo-Controlled Clinical Trial. Nutrients 2023, 15, 331. [Google Scholar] [CrossRef]
- Mollazadeh-Narestan, Z.; Yavarikia, P.; Homayouni-Rad, A.; Samadi Kafil, H.; Mohammad-Alizadeh-Charandabi, S.; Gholizadeh, P.; Mirghafourvand, M. Comparing the Effect of Probiotic and Fluconazole on Treatment and Recurrence of Vulvovaginal Candidiasis: A Triple-Blinded Randomized Controlled Trial. Probiotics Antimicrob. Proteins 2023, 15, 1436–1446. [Google Scholar] [CrossRef]
- Ang, X.-Y.; Chung, F.-Y.-L.; Lee, B.-K.; Azhar, S.N.A.; Sany, S.; Roslan, N.S.; Ahmad, N.; Yusof, S.M.; Abdullah, N.; Nik Ab Rahman, N.N.; et al. Lactobacilli reduce recurrences of vaginal candidiasis in pregnant women: A randomized, double-blind, placebo-controlled study. J. Appl. Microbiol. 2022, 132, 3168–3180. [Google Scholar] [CrossRef] [PubMed]
- Mändar, R.; Sõerunurk, G.; Štšepetova, J.; Smidt, I.; Rööp, T.; Kõljalg, S.; Saare, M.; Ausmees, K.; Le, D.D.; Jaagura, M.; et al. Impact of Lactobacillus crispatus-containing oral and vaginal probiotics on vaginal health: A randomised double-blind placebo controlled clinical trial. Benef. Microbes 2023, 14, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Lew, L.-C.; Hor, Y.-Y.; Yusoff, N.A.A.; Choi, S.-B.; Yusoff, M.S.B.; Roslan, N.S.; Ahmad, A.; Mohammad, J.A.M.; Abdullah, M.F.I.L.; Zakaria, N.; et al. Probiotic Lactobacillus plantarum P8 alleviated stress and anxiety while enhancing memory and cognition in stressed adults: A randomised, double-blind, placebo-controlled study. Clin. Nutr. 2019, 38, 2053–2064. [Google Scholar] [CrossRef]
- Akhgarjand, C.; Vahabi, Z.; Shab-Bidar, S.; Etesam, F.; Djafarian, K. Effects of probiotic supplements on cognition, anxiety, and physical activity in subjects with mild and moderate Alzheimer’s disease: A randomized, double-blind, and placebo-controlled study. Front. Aging Neurosci. 2022, 14, 1032494. [Google Scholar] [CrossRef] [PubMed]
- Arnold, L.E.; Luna, R.A.; Williams, K.; Chan, J.; Parker, R.A.; Wu, Q.; Hollway, J.A.; Jeffs, A.; Lu, F.; Coury, D.L.; et al. Probiotics for Gastrointestinal Symptoms and Quality of Life in Autism: A Placebo-Controlled Pilot Trial. J. Child Adolesc. Psychopharmacol. 2019, 29, 659–669. [Google Scholar] [CrossRef] [PubMed]
- Papalini, S.; Michels, F.; Kohn, N.; Wegman, J.; van Hemert, S.; Roelofs, K.; Arias-Vasquez, A.; Aarts, E. Stress matters: Randomized controlled trial on the effect of probiotics on neurocognition. Neurobiol. Stress 2019, 10, 100141. [Google Scholar] [CrossRef] [PubMed]
- Haghighat, N.; Rajabi, S.; Mohammadshahi, M. Effect of synbiotic and probiotic supplementation on serum brain-derived neurotrophic factor level, depression and anxiety symptoms in hemodialysis patients: A randomized, double-blinded, clinical trial. Nutr. Neurosci. 2021, 24, 490–499. [Google Scholar] [CrossRef] [PubMed]
- Salleh, R.M.; Kuan, G.; Aziz, M.N.A.; Rahim, M.R.A.; Rahayu, T.; Sulaiman, S.; Kusuma, D.W.Y.; Adikari, A.M.G.C.P.; Razam, M.S.M.; Radhakrishnan, A.K.; et al. Effects of Probiotics on Anxiety, Stress, Mood and Fitness of Badminton Players. Nutrients 2021, 13, 1783. [Google Scholar] [CrossRef] [PubMed]
- Asher, M.I.; Montefort, S.; Björkstén, B.; Lai, C.K.W.; Strachan, D.P.; Weiland, S.K.; Williams, H.; ISAAC Phase Three Study Group. Worldwide time trends in the prevalence of symptoms of asthma, allergic rhinoconjunctivitis, and eczema in childhood: ISAAC Phases One and Three repeat multicountry cross-sectional surveys. Lancet 2006, 368, 733–743. [Google Scholar] [CrossRef] [PubMed]
- Sicherer, S.H.; Wood, R.A.; Stablein, D.; Lindblad, R.; Burks, A.W.; Liu, A.H.; Jones, S.M.; Fleischer, D.M.; Leung, D.Y.M.; Sampson, H.A. Maternal consumption of peanut during pregnancy is associated with peanut sensitization in atopic infants. J. Allergy Clin. Immunol. 2010, 126, 1191–1197. [Google Scholar] [CrossRef] [PubMed]
- Kalliomäki, M.; Salminen, S.; Arvilommi, H.; Kero, P.; Koskinen, P.; Isolauri, E. Probiotics in primary prevention of atopic disease: A randomised placebo-controlled trial. Lancet 2001, 357, 1076–1079. [Google Scholar] [CrossRef]
- West, C.E.; Hammarström, M.-L.; Hernell, O. Probiotics during weaning reduce the incidence of eczema. Pediatr. Allergy Immunol. 2009, 20, 430–437. [Google Scholar] [CrossRef]
- Hougee, S.; Vriesema, A.J.M.; Wijering, S.C.; Knippels, L.M.J.; Folkerts, G.; Nijkamp, F.P.; Knol, J.; Garssen, J. Oral treatment with probiotics reduces allergic symptoms in ovalbumin-sensitized mice: A bacterial strain comparative study. Int. Arch. Allergy Immunol. 2010, 151, 107–117. [Google Scholar] [CrossRef]
- Feleszko, W.; Jaworska, J.; Rha, R.-D.; Steinhausen, S.; Avagyan, A.; Jaudszus, A.; Ahrens, B.; Groneberg, D.A.; Wahn, U.; Hamelmann, E. Probiotic-induced suppression of allergic sensitization and airway inflammation is associated with an increase of T regulatory-dependent mechanisms in a murine model of asthma. Clin. Exp. Allergy 2007, 37, 498–505. [Google Scholar] [CrossRef]
- Zhang, B.; An, J.; Shimada, T.; Liu, S.; Maeyama, K. Oral administration of Enterococcus faecalis FK-23 suppresses Th17 cell development and attenuates allergic airway responses in mice. Int. J. Mol. Med. 2012, 30, 248–254. [Google Scholar] [CrossRef]
- Kanis, J.A.; McCloskey, E.V.; Johansson, H.; Cooper, C.; Rizzoli, R.; Reginster, J.-Y.; Scientific Advisory Board of the European Society for Clinical and Economic Aspects of Osteoporosis and Osteoarthritis (ESCEO) and the Committee of Scientific Advisors of the International Osteoporosis Foundation (IOF). European guidance for the diagnosis and management of osteoporosis in postmenopausal women. Osteoporos. Int. 2013, 24, 23–57. [Google Scholar] [CrossRef]
- Collins, F.L.; Rios-Arce, N.D.; Schepper, J.D.; Parameswaran, N.; McCabe, L.R. The Potential of Probiotics as a Therapy for Osteoporosis. Microbiol. Spectr. 2017, 5, 213–233. [Google Scholar] [CrossRef] [PubMed]
- Schepper, J.D.; Irwin, R.; Kang, J.; Dagenais, K.; Lemon, T.; Shinouskis, A.; Parameswaran, N.; McCabe, L.R. Probiotics in Gut-Bone Signaling. Adv. Exp. Med. Biol. 2017, 1033, 225–247. [Google Scholar] [PubMed]
- Ohlsson, C.; Engdahl, C.; Fåk, F.; Andersson, A.; Windahl, S.H.; Farman, H.H.; Movérare-Skrtic, S.; Islander, U.; Sjögren, K. Probiotics protect mice from ovariectomy-induced cortical bone loss. PLoS ONE 2014, 9, e92368. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, F.C.; Castro, A.S.B.; Rodrigues, V.C.; Fernandes, S.A.; Fontes, E.A.F.; de Oliveira, T.T.; Martino, H.S.D.; de Luces Fortes Ferreira, C.L. Yacon flour and Bifidobacterium longum modulate bone health in rats. J. Med. Food 2012, 15, 664–670. [Google Scholar] [CrossRef] [PubMed]
- Britton, R.A.; Irwin, R.; Quach, D.; Schaefer, L.; Zhang, J.; Lee, T.; Parameswaran, N.; McCabe, L.R. Probiotic L. reuteri treatment prevents bone loss in a menopausal ovariectomized mouse model. J. Cell. Physiol. 2014, 229, 1822–1830. [Google Scholar] [CrossRef] [PubMed]
- Chiang, S.-S.; Pan, T.-M. Antiosteoporotic effects of Lactobacillus-fermented soy skim milk on bone mineral density and the microstructure of femoral bone in ovariectomized mice. J. Agric. Food Chem. 2011, 59, 7734–7742. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Motyl, K.J.; Irwin, R.; MacDougald, O.A.; Britton, R.A.; McCabe, L.R. Loss of Bone and Wnt10b Expression in Male Type 1 Diabetic Mice Is Blocked by the Probiotic Lactobacillus reuteri. Endocrinology 2015, 156, 3169–3182. [Google Scholar] [CrossRef]
- McCabe, L.R.; Irwin, R.; Schaefer, L.; Britton, R.A. Probiotic use decreases intestinal inflammation and increases bone density in healthy male but not female mice. J. Cell. Physiol. 2013, 228, 1793–1798. [Google Scholar] [CrossRef] [PubMed]
- Collins, F.L.; Irwin, R.; Bierhalter, H.; Schepper, J.; Britton, R.A.; Parameswaran, N.; McCabe, L.R. Lactobacillus reuteri 6475 Increases Bone Density in Intact Females Only under an Inflammatory Setting. PLoS ONE 2016, 11, e0153180. [Google Scholar] [CrossRef]
- Kruger, M.C.; Fear, A.; Chua, W.-H.; Plimmer, G.G.; Schollum, L.M. The effect of Lactobacillus rhamnosus HN001 on mineral absorption and bone health in growing male and ovariectomised female rats. Dairy Sci. Technol. 2009, 89, 219–231. [Google Scholar] [CrossRef]
- Tomofuji, T.; Ekuni, D.; Azuma, T.; Irie, K.; Endo, Y.; Yamamoto, T.; Ishikado, A.; Sato, T.; Harada, K.; Suido, H.; et al. Supplementation of broccoli or Bifidobacterium longum-fermented broccoli suppresses serum lipid peroxidation and osteoclast differentiation on alveolar bone surface in rats fed a high-cholesterol diet. Nutr. Res. 2012, 32, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-Y.; Chassaing, B.; Tyagi, A.M.; Vaccaro, C.; Luo, T.; Adams, J.; Darby, T.M.; Weitzmann, M.N.; Mulle, J.G.; Gewirtz, A.T.; et al. Sex steroid deficiency-associated bone loss is microbiota dependent and prevented by probiotics. J. Clin. Investig. 2016, 126, 2049–2063. [Google Scholar] [CrossRef] [PubMed]
- Parvaneh, K.; Ebrahimi, M.; Sabran, M.R.; Karimi, G.; Hwei, A.N.M.; Abdul-Majeed, S.; Ahmad, Z.; Ibrahim, Z.; Jamaluddin, R. Probiotics (Bifidobacterium longum) Increase Bone Mass Density and Upregulate Sparc and Bmp-2 Genes in Rats with Bone Loss Resulting from Ovariectomy. BioMed Res. Int. 2015, 2015, 897639. [Google Scholar] [CrossRef]
- Wang, W.; Zhou, H.; Liu, L. Side effects of methotrexate therapy for rheumatoid arthritis: A systematic review. Eur. J. Med. Chem. 2018, 158, 502–516. [Google Scholar] [CrossRef]
- Tilstra, J.S.; Lienesch, D.W. Rheumatoid Nodules. Dermatol. Clin. 2015, 33, 361–371. [Google Scholar] [CrossRef]
- Sepriano, A.; Kerschbaumer, A.; Smolen, J.S.; van der Heijde, D.; Dougados, M.; van Vollenhoven, R.; McInnes, I.B.; Bijlsma, J.W.; Burmester, G.R.; de Wit, M.; et al. Safety of synthetic and biological DMARDs: A systematic literature review informing the 2019 update of the EULAR recommendations for the management of rheumatoid arthritis. Ann. Rheum. Dis. 2020, 79, 760–770. [Google Scholar] [CrossRef]
- Schrezenmeier, E.; Dörner, T. Mechanisms of action of hydroxychloroquine and chloroquine: Implications for rheumatology. Nat. Rev. Rheumatol. 2020, 16, 155–166. [Google Scholar] [CrossRef]
- Dos Reis Neto, E.T.; Kakehasi, A.M.; de Medeiros Pinheiro, M.; Ferreira, G.A.; Marques, C.D.L.; da Mota, L.M.H.; Dos Santos Paiva, E.; Pileggi, G.C.S.; Sato, E.I.; Reis, A.P.M.G.; et al. Revisiting hydroxychloroquine and chloroquine for patients with chronic immunity-mediated inflammatory rheumatic diseases. Adv. Rheumatol. 2020, 60, 32. [Google Scholar] [CrossRef] [PubMed]
- Wells, P.M.; Adebayo, A.S.; Bowyer, R.C.E.; Freidin, M.B.; Finckh, A.; Strowig, T.; Lesker, T.R.; Alpizar-Rodriguez, D.; Gilbert, B.; Kirkham, B.; et al. Associations between gut microbiota and genetic risk for rheumatoid arthritis in the absence of disease: A cross-sectional study. Lancet Rheumatol. 2020, 2, e418–e427. [Google Scholar] [CrossRef] [PubMed]
- Pianta, A.; Arvikar, S.; Strle, K.; Drouin, E.E.; Wang, Q.; Costello, C.E.; Steere, A.C. Evidence of the Immune Relevance of Prevotella copri, a Gut Microbe, in Patients With Rheumatoid Arthritis. Arthritis Rheumatol. 2017, 69, 964–975. [Google Scholar] [CrossRef] [PubMed]
- Balakrishnan, B.; Luckey, D.; Taneja, V. Autoimmunity-Associated Gut Commensals Modulate Gut Permeability and Immunity in Humanized Mice. Mil. Med. 2019, 184, 529–536. [Google Scholar] [CrossRef]
- Ferreira-Halder, C.V.; Faria, A.V.d.S.; Andrade, S.S. Action and function of Faecalibacterium prausnitzii in health and disease. Best Pract. Res. Clin. Gastroenterol. 2017, 31, 643–648. [Google Scholar] [CrossRef]
- Chu, X.-J.; Cao, N.-W.; Zhou, H.-Y.; Meng, X.; Guo, B.; Zhang, H.-Y.; Li, B.-Z. The oral and gut microbiome in rheumatoid arthritis patients: A systematic review. Rheumatology 2021, 60, 1054–1066. [Google Scholar] [CrossRef]
- Foreman, K.J.; Marquez, N.; Dolgert, A.; Fukutaki, K.; Fullman, N.; McGaughey, M.; Pletcher, M.A.; Smith, A.E.; Tang, K.; Yuan, C.-W.; et al. Forecasting life expectancy, years of life lost, and all-cause and cause-specific mortality for 250 causes of death: Reference and alternative scenarios for 2016-40 for 195 countries and territories. Lancet 2018, 392, 2052–2090. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, A.; Ferro, C.J.; Balafa, O.; Burnier, M.; Ekart, R.; Halimi, J.-M.; Kreutz, R.; Mark, P.B.; Persu, A.; Rossignol, P.; et al. Mineralocorticoid receptor antagonists for nephroprotection and cardioprotection in patients with diabetes mellitus and chronic kidney disease. Nephrol. Dial. Transplant. 2023, 38, 10–25. [Google Scholar] [CrossRef]
- Ortiz, A.; Fernandez-Fernandez, B. Atrasentan: The Difficult Task of Integrating Endothelin A Receptor Antagonists into Current Treatment Paradigm for Diabetic Kidney Disease. Clin. J. Am. Soc. Nephrol. 2021, 16, 1775–1778. [Google Scholar] [CrossRef]
- Rovin, B.H.; Adler, S.G.; Barratt, J.; Bridoux, F.; Burdge, K.A.; Chan, T.M.; Cook, H.T.; Fervenza, F.C.; Gibson, K.L.; Glassock, R.J.; et al. Executive summary of the KDIGO 2021 Guideline for the Management of Glomerular Diseases. Kidney Int. 2021, 100, 753–779. [Google Scholar] [CrossRef]
- Makkapati, S.; D’Agati, V.D.; Balsam, L. “Green Smoothie Cleanse” Causing Acute Oxalate Nephropathy. Am. J. Kidney Dis. 2018, 71, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Carriazo, S.; Perez-Gomez, M.V.; Cordido, A.; García-González, M.A.; Sanz, A.B.; Ortiz, A.; Sanchez-Niño, M.D. Dietary Care for ADPKD Patients: Current Status and Future Directions. Nutrients 2019, 11, 1576. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Cao, C.; Wu, Z.; Zhang, H.; Sun, Z.; Wang, M.; Xu, H.; Zhao, Z.; Wang, Y.; Pei, G.; et al. The probiotic L. casei Zhang slows the progression of acute and chronic kidney disease. Cell Metab. 2021, 33, 1926–1942.e8. [Google Scholar] [CrossRef] [PubMed]
- Andrade-Oliveira, V.; Amano, M.T.; Correa-Costa, M.; Castoldi, A.; Felizardo, R.J.F.; de Almeida, D.C.; Bassi, E.J.; Moraes-Vieira, P.M.; Hiyane, M.I.; Rodas, A.C.D.; et al. Gut Bacteria Products Prevent AKI Induced by Ischemia-Reperfusion. J. Am. Soc. Nephrol. 2015, 26, 1877–1888. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Y.J.; Loh, Y.W.; Singer, J.; Zhu, W.; Macia, L.; Mackay, C.R.; Wang, W.; Chadban, S.J.; Wu, H. Fiber Derived Microbial Metabolites Prevent Acute Kidney Injury Through G-Protein Coupled Receptors and HDAC Inhibition. Front. Cell Dev. Biol. 2021, 9, 648639. [Google Scholar] [CrossRef]
- Miranda Alatriste, P.V.; Urbina Arronte, R.; Gómez Espinosa, C.O.; Espinosa Cuevas, M. de los Á. Effect of probiotics on human blood urea levels in patients with chronic renal failure. Nutr. Hosp. 2014, 29, 582–590. [Google Scholar]
- Jiang, S.; Xie, S.; Lv, D.; Zhang, Y.; Deng, J.; Zeng, L.; Chen, Y. A reduction in the butyrate producing species Roseburia spp. and Faecalibacterium prausnitzii is associated with chronic kidney disease progression. Antonie Van Leeuwenhoek 2016, 109, 1389–1396. [Google Scholar] [CrossRef]
- Miraghajani, M.; Zaghian, N.; Mirlohi, M.; Feizi, A.; Ghiasvand, R. The Impact of Probiotic Soy Milk Consumption on Oxidative Stress Among Type 2 Diabetic Kidney Disease Patients: A Randomized Controlled Clinical Trial. J. Ren. Nutr. 2017, 27, 317–324. [Google Scholar] [CrossRef]
- Guida, B.; Germanò, R.; Trio, R.; Russo, D.; Memoli, B.; Grumetto, L.; Barbato, F.; Cataldi, M. Effect of short-term synbiotic treatment on plasma p-cresol levels in patients with chronic renal failure: A randomized clinical trial. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 1043–1049. [Google Scholar] [CrossRef] [PubMed]
- Rossi, M.; Johnson, D.W.; Morrison, M.; Pascoe, E.M.; Coombes, J.S.; Forbes, J.M.; Szeto, C.-C.; McWhinney, B.C.; Ungerer, J.P.J.; Campbell, K.L. Synbiotics Easing Renal Failure by Improving Gut Microbiology (SYNERGY): A Randomized Trial. Clin. J. Am. Soc. Nephrol. 2016, 11, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Ranganathan, N.; Ranganathan, P.; Friedman, E.A.; Joseph, A.; Delano, B.; Goldfarb, D.S.; Tam, P.; Rao, A.V.; Anteyi, E.; Musso, C.G. Pilot study of probiotic dietary supplementation for promoting healthy kidney function in patients with chronic kidney disease. Adv. Ther. 2010, 27, 634–647. [Google Scholar] [CrossRef] [PubMed]
- Wang, I.-K.; Yen, T.-H.; Hsieh, P.-S.; Ho, H.-H.; Kuo, Y.-W.; Huang, Y.-Y.; Kuo, Y.-L.; Li, C.-Y.; Lin, H.-C.; Wang, J.-Y. Effect of a Probiotic Combination in an Experimental Mouse Model and Clinical Patients With Chronic Kidney Disease: A Pilot Study. Front. Nutr. 2021, 8, 661794. [Google Scholar] [CrossRef] [PubMed]
- Pavan, M. Influence of prebiotic and probiotic supplementation on the progression of chronic kidney disease. Minerva Urol. Nefrol. 2016, 68, 222–226. [Google Scholar]
- Yang, C.-Y.; Chen, T.-W.; Lu, W.-L.; Liang, S.-S.; Huang, H.-D.; Tseng, C.-P.; Tarng, D.-C. Synbiotics Alleviate the Gut Indole Load and Dysbiosis in Chronic Kidney Disease. Cells 2021, 10, 114. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, R.; Pechenyak, B.; Vyas, U.; Ranganathan, P.; Weinberg, A.; Liang, P.; Mallappallil, M.C.; Norin, A.J.; Friedman, E.A.; Saggi, S.J. Randomized controlled trial of strain-specific probiotic formulation (Renadyl) in dialysis patients. BioMed Res. Int. 2014, 2014, 568571. [Google Scholar] [CrossRef] [PubMed]
- Meijers, B.K.I.; De Preter, V.; Verbeke, K.; Vanrenterghem, Y.; Evenepoel, P. p-Cresyl sulfate serum concentrations in haemodialysis patients are reduced by the prebiotic oligofructose-enriched inulin. Nephrol. Dial. Transplant. 2010, 25, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Sirich, T.L.; Plummer, N.S.; Gardner, C.D.; Hostetter, T.H.; Meyer, T.W. Effect of increasing dietary fiber on plasma levels of colon-derived solutes in hemodialysis patients. Clin. J. Am. Soc. Nephrol. 2014, 9, 1603–1610. [Google Scholar] [CrossRef]
- Wang, I.-K.; Wu, Y.-Y.; Yang, Y.-F.; Ting, I.-W.; Lin, C.-C.; Yen, T.-H.; Chen, J.-H.; Wang, C.-H.; Huang, C.-C.; Lin, H.-C. The effect of probiotics on serum levels of cytokine and endotoxin in peritoneal dialysis patients: A randomised, double-blind, placebo-controlled trial. Benef. Microbes 2015, 6, 423–430. [Google Scholar] [CrossRef]
- Lundy, S.D.; Sangwan, N.; Parekh, N.V.; Selvam, M.K.P.; Gupta, S.; McCaffrey, P.; Bessoff, K.; Vala, A.; Agarwal, A.; Sabanegh, E.S.; et al. Functional and Taxonomic Dysbiosis of the Gut, Urine, and Semen Microbiomes in Male Infertility. Eur. Urol. 2021, 79, 826–836. [Google Scholar] [CrossRef]
- Koedooder, R.; Mackens, S.; Budding, A.; Fares, D.; Blockeel, C.; Laven, J.; Schoenmakers, S. Identification and evaluation of the microbiome in the female and male reproductive tracts. Hum. Reprod. Update 2019, 25, 298–325. [Google Scholar] [CrossRef]
- Rowe, M.; Veerus, L.; Trosvik, P.; Buckling, A.; Pizzari, T. The Reproductive Microbiome: An Emerging Driver of Sexual Selection, Sexual Conflict, Mating Systems, and Reproductive Isolation. Trends Ecol. Evol. 2020, 35, 220–234. [Google Scholar] [CrossRef] [PubMed]
- Schoenmakers, S.; Steegers-Theunissen, R.; Faas, M. The matter of the reproductive microbiome. Obstet. Med. 2019, 12, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Reid, J.N.S.; Bisanz, J.E.; Monachese, M.; Burton, J.P.; Reid, G. The Rationale for Probiotics Improving Reproductive Health and Pregnancy Outcome. Am. J. Reprod. Immunol. 2013, 69, 558–566. [Google Scholar] [CrossRef] [PubMed]
- Sanz, Y. Gut microbiota and probiotics in maternal and infant health. Am. J. Clin. Nutr. 2011, 94, S2000–S2005. [Google Scholar] [CrossRef] [PubMed]
- Weng, S.-L.; Chiu, C.-M.; Lin, F.-M.; Huang, W.-C.; Liang, C.; Yang, T.; Yang, T.-L.; Liu, C.-Y.; Wu, W.-Y.; Chang, Y.-A.; et al. Bacterial Communities in Semen from Men of Infertile Couples: Metagenomic Sequencing Reveals Relationships of Seminal Microbiota to Semen Quality. PLoS ONE 2014, 9, e110152. [Google Scholar] [CrossRef]
- Tomaiuolo, R.; Veneruso, I.; Cariati, F.; D’Argenio, V. Microbiota and Human Reproduction: The Case of Male Infertility. High-Throughput 2020, 9, 10. [Google Scholar] [CrossRef]
- Barbonetti, A.; Cinque, B.; Vassallo, M.R.C.; Mineo, S.; Francavilla, S.; Cifone, M.G.; Francavilla, F. Effect of vaginal probiotic lactobacilli on in vitro–induced sperm lipid peroxidation and its impact on sperm motility and viability. Fertil. Steril. 2011, 95, 2485–2488. [Google Scholar] [CrossRef]
- Aitken, R.J.; Harkiss, D.; Buckingham, D.W. Analysis of lipid peroxidation mechanisms in human spermatozoa. Mol. Reprod. Dev. 1993, 35, 302–315. [Google Scholar] [CrossRef]
- Dardmeh, F.; Alipour, H.; Gazerani, P.; van der Horst, G.; Brandsborg, E.; Nielsen, H.I. Lactobacillus rhamnosus PB01 (DSM 14870) supplementation affects markers of sperm kinematic parameters in a diet-induced obesity mice model. PLoS ONE 2017, 12, e0185964. [Google Scholar] [CrossRef]
- Poutahidis, T.; Springer, A.; Levkovich, T.; Qi, P.; Varian, B.J.; Lakritz, J.R.; Ibrahim, Y.M.; Chatzigiagkos, A.; Alm, E.J.; Erdman, S.E. Probiotic Microbes Sustain Youthful Serum Testosterone Levels and Testicular Size in Aging Mice. PLoS ONE 2014, 9, e84877. [Google Scholar] [CrossRef] [PubMed]
- Valcarce, D.G.; Genovés, S.; Riesco, M.F.; Martorell, P.; Herráez, M.P.; Ramón, D.; Robles, V. Probiotic administration improves sperm quality in asthenozoospermic human donors. Benef. Microbes 2017, 8, 193–206. [Google Scholar] [CrossRef] [PubMed]
- Attia, Y.A.; Abd El Hamid, E.A.; Ismaiel, A.M.; El-Nagar, A. The detoxication of nitrate by two antioxidants or a probiotic, and the effects on blood and seminal plasma profiles and reproductive function of New Zealand White rabbit bucks. Animal 2013, 7, 591–601. [Google Scholar] [CrossRef] [PubMed]
- Al-Asmakh, M.; Stukenborg, J.-B.; Reda, A.; Anuar, F.; Strand, M.-L.; Hedin, L.; Pettersson, S.; Söder, O. The Gut Microbiota and Developmental Programming of the Testis in Mice. PLoS ONE 2014, 9, e103809. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, H.; Yang, Q.; Li, P.; Wen, Y.; Han, X.; Li, B.; Jiang, H.; Li, X. Genomic Sequencing Reveals the Diversity of Seminal Bacteria and Relationships to Reproductive Potential in Boar Sperm. Front. Microbiol. 2020, 11, 1873. [Google Scholar] [CrossRef]
- Tian, X.; Yu, Z.; Feng, P.; Ye, Z.; Li, R.; Liu, J.; Hu, J.; Kakade, A.; Liu, P.; Li, X. Lactobacillus plantarum TW1-1 Alleviates Diethylhexylphthalate-Induced Testicular Damage in Mice by Modulating Gut Microbiota and Decreasing Inflammation. Front. Cell. Infect. Microbiol. 2019, 9, 221. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J.; Smith, T.B.; Jobling, M.S.; Baker, M.A.; De Iuliis, G.N. Oxidative stress and male reproductive health. Asian J. Androl. 2014, 16, 31–38. [Google Scholar] [CrossRef]
- Rosa, L.S.; Santos, M.L.; Abreu, J.P.; Balthazar, C.F.; Rocha, R.S.; Silva, H.L.A.; Esmerino, E.A.; Duarte, M.C.K.H.; Pimentel, T.C.; Freitas, M.Q.; et al. Antiproliferative and apoptotic effects of probiotic whey dairy beverages in human prostate cell lines. Food Res. Int. 2020, 137, 109450. [Google Scholar] [CrossRef]
- Celebioglu, H.U. Effects of potential synbiotic interaction between Lactobacillus rhamnosus GG and salicylic acid on human colon and prostate cancer cells. Arch. Microbiol. 2021, 203, 1221–1229. [Google Scholar] [CrossRef]
- Chiancone, F.; Carrino, M.; Meccariello, C.; Pucci, L.; Fedelini, M.; Fedelini, P. The Use of a Combination of Vaccinium Macracarpon, Lycium barbarum L. and Probiotics (Bifiprost®) for the Prevention of Chronic Bacterial Prostatitis: A Double-Blind Randomized Study. Urol. Int. 2019, 103, 423–426. [Google Scholar] [CrossRef] [PubMed]
- Pacifici, L.; Santacroce, L.; Dipalma, G.; Haxhirexha, K.; Topi, S.; Cantore, S.; Altini, V.; Pacifici, A.; De Vito, D.; Pettini, F. Gender medicine: The impact of probiotics on male patients. Clin. Ter. 2021, 172, 8–15. [Google Scholar]
- Ravel, J.; Gajer, P.; Abdo, Z.; Schneider, G.M.; Koenig, S.S.K.; McCulle, S.L.; Karlebach, S.; Gorle, R.; Russell, J.; Tacket, C.O.; et al. Vaginal microbiome of reproductive-age women. Proc. Natl. Acad. Sci. USA 2011, 108, 4680–4687. [Google Scholar] [CrossRef]
- Pelzer, E.S.; Allan, J.A.; Waterhouse, M.A.; Ross, T.; Beagley, K.W.; Knox, C.L. Microorganisms within Human Follicular Fluid: Effects on IVF. PLoS ONE 2013, 8, e59062. [Google Scholar] [CrossRef]
- Tachedjian, G.; Aldunate, M.; Bradshaw, C.S.; Cone, R.A. The role of lactic acid production by probiotic Lactobacillus species in vaginal health. Res. Microbiol. 2017, 168, 782–792. [Google Scholar] [CrossRef]
- Fredricks, D.N.; Fiedler, T.L.; Marrazzo, J.M. Molecular Identification of Bacteria Associated with Bacterial Vaginosis. N. Engl. J. Med. 2005, 353, 1899–1911. [Google Scholar] [CrossRef]
- Verhelst, R.; Verstraelen, H.; Claeys, G.; Verschraegen, G.; Delanghe, J.; Van Simaey, L.; De Ganck, C.; Temmerman, M.; Vaneechoutte, M. Cloning of 16S rRNA genes amplified from normal and disturbed vaginal microflora suggests a strong association between Atopobium vaginae, Gardnerella vaginalis and bacterial vaginosis. BMC Microbiol. 2004, 4, 16. [Google Scholar] [CrossRef] [PubMed]
- O’Hanlon, D.E.; Moench, T.R.; Cone, R.A. Vaginal pH and Microbicidal Lactic Acid When Lactobacilli Dominate the Microbiota. PLoS ONE 2013, 8, e80074. [Google Scholar] [CrossRef] [PubMed]
- Quaranta, G.; Sanguinetti, M.; Masucci, L. Fecal Microbiota Transplantation: A Potential Tool for Treatment of Human Female Reproductive Tract Diseases. Front. Immunol. 2019, 10, 2653. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.M.; Chase, D.M.; Herbst-Kralovetz, M.M. Uterine microbiota: Residents, tourists, or invaders? Front. Immunol. 2018, 9, 208. [Google Scholar] [CrossRef] [PubMed]
- Heil, B.A.; Paccamonti, D.L.; Sones, J.L. Role for the mammalian female reproductive tract microbiome in pregnancy outcomes. Physiol. Genom. 2019, 51, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Brotman, R.M. Vaginal microbiome and sexually transmitted infections: An epidemiologic perspective. J. Clin. Investig. 2011, 121, 4610–4617. [Google Scholar] [CrossRef]
- MacPhee, R.A.; Hummelen, R.; Bisanz, J.E.; Miller, W.L.; Reid, G. Probiotic strategies for the treatment and prevention of bacterial vaginosis. Expert Opin. Pharmacother. 2010, 11, 2985–2995. [Google Scholar] [CrossRef]
- Hütt, P.; Lapp, E.; Štšepetova, J.; Smidt, I.; Taelma, H.; Borovkova, N.; Oopkaup, H.; Ahelik, A.; Rööp, T.; Hoidmets, D.; et al. Characterisation of probiotic properties in human vaginal lactobacilli strains. Microb. Ecol. Health Dis. 2016, 27, 30484. [Google Scholar] [CrossRef]
- Borges, S.; Silva, J.; Teixeira, P. The role of lactobacilli and probiotics in maintaining vaginal health. Arch. Gynecol. Obstet. 2014, 289, 479–489. [Google Scholar] [CrossRef]
- Santos, C.M.A.; Pires, M.C.V.; Leão, T.L.; Hernández, Z.P.; Rodriguez, M.L.; Martins, A.K.S.; Miranda, L.S.; Martins, F.S.; Nicoli, J.R. Selection of Lactobacillus strains as potential probiotics for vaginitis treatment. Microbiology 2016, 162, 1195–1207. [Google Scholar] [CrossRef]
- Senok, A.C.; Verstraelen, H.; Temmerman, M.; Botta, G.A. Probiotics for the treatment of bacterial vaginosis. In Cochrane Database of Systematic Reviews; Senok, A.C., Ed.; John Wiley & Sons, Ltd.: Chichester, UK, 2006. [Google Scholar]
- Andersch, B.; Lindell, D.; Dahlén, I.; Brandberg, Å. Bacterial Vaginosis and the Effect of Intermittent Prophylactic Treatment with an Acid Lactate Gel. Gynecol. Obstet. Investig. 1990, 30, 114–119. [Google Scholar] [CrossRef]
- Decena, D.C.D.; Co, J.T.; Manalastas, R.M.; Palaypayon, E.P.; Padolina, C.S.; Sison, J.M.; Dancel, L.A.; Lelis, M.A. Metronidazole with Lactacyd vaginal gel in bacterial vaginosis. J. Obstet. Gynaecol. Res. 2006, 32, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Valeria Bahamondes, M.; Portugal, P.M.; Brolazo, E.M.; Simões, J.A.; Bahamondes, L. Use of a lactic acid plus lactoserum intimate liquid soap for external hygiene in the prevention of bacterial vaginosis recurrence after metronidazole oral treatment. Rev. Assoc. Med. Bras. 2011, 57, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Valore, E.V.; Park, C.H.; Igreti, S.L.; Ganz, T. Antimicrobial components of vaginal fluid. Am. J. Obstet. Gynecol. 2002, 187, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Graver, M.A.; Wade, J.J. The role of acidification in the inhibition of Neisseria gonorrhoeae by vaginal lactobacilli during anaerobic growth. Ann. Clin. Microbiol. Antimicrob. 2011, 10, 8. [Google Scholar] [CrossRef] [PubMed]
- Nardini, P.; Ñahui Palomino, R.A.; Parolin, C.; Laghi, L.; Foschi, C.; Cevenini, R.; Vitali, B.; Marangoni, A. Lactobacillus crispatus inhibits the infectivity of Chlamydia trachomatis elementary bodies, in vitro study. Sci. Rep. 2016, 6, 29024. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Luna, Y.; Yu, P.; Fan, H. Lactobacilli Inactivate Chlamydia trachomatis through Lactic Acid but Not H2O2. PLoS ONE 2014, 9, e107758. [Google Scholar] [CrossRef] [PubMed]
- Breshears, L.M.; Edwards, V.L.; Ravel, J.; Peterson, M.L. Lactobacillus crispatus inhibits growth of Gardnerella vaginalis and Neisseria gonorrhoeae on a porcine vaginal mucosa model. BMC Microbiol. 2015, 15, 276. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, C.; Balkus, J.E.; Fredricks, D.; Liu, C.; McKernan-Mullin, J.; Frenkel, L.M.; Mwachari, C.; Luque, A.; Cohn, S.E.; Cohen, C.R.; et al. Interaction Between Lactobacilli, Bacterial Vaginosis-Associated Bacteria, and HIV Type 1 RNA and DNA Genital Shedding in U.S. and Kenyan Women. AIDS Res. Hum. Retroviruses 2013, 29, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Borgdorff, H.; Tsivtsivadze, E.; Verhelst, R.; Marzorati, M.; Jurriaans, S.; Ndayisaba, G.F.; Schuren, F.H.; van de Wijgert, J.H. Lactobacillus-dominated cervicovaginal microbiota associated with reduced HIV/STI prevalence and genital HIV viral load in African women. ISME J. 2014, 8, 1781–1793. [Google Scholar] [CrossRef] [PubMed]
- Aldunate, M.; Tyssen, D.; Johnson, A.; Zakir, T.; Sonza, S.; Moench, T.; Cone, R.; Tachedjian, G. Vaginal concentrations of lactic acid potently inactivate HIV. J. Antimicrob. Chemother. 2013, 68, 2015–2025. [Google Scholar] [CrossRef]
- Dimitonova, S.P.; Danova, S.T.; Serkedjieva, J.P.; Bakalov, B.V. Antimicrobial activity and protective properties of vaginal lactobacilli from healthy Bulgarian women. Anaerobe 2007, 13, 178–184. [Google Scholar] [CrossRef]
- Isaacs, C.E.; Xu, W. Theaflavin-3,3′-Digallate and Lactic Acid Combinations Reduce Herpes Simplex Virus Infectivity. Antimicrob. Agents Chemother. 2013, 57, 3806–3814. [Google Scholar] [CrossRef]
- Conti, C.; Malacrino, C.; Mastromarino, P. Inhibition of herpes simplex virus type 2 by vaginal lactobacilli. J. Physiol. Pharmacol. 2009, 60, 19–26. [Google Scholar]
- Chen, Q.; Wang, B.; Wang, S.; Qian, X.; Li, X.; Zhao, J.; Zhang, H.; Chen, W.; Wang, G. Modulation of the Gut Microbiota Structure with Probiotics and Isoflavone Alleviates Metabolic Disorder in Ovariectomized Mice. Nutrients 2021, 13, 1793. [Google Scholar] [CrossRef]
- Szydłowska, I.; Marciniak, A.; Brodowska, A.; Loj, B.; Ciećwież, S.; Skonieczna-Żydecka, K.; Palma, J.; Łoniewski, I.; Stachowska, E. Effects of probiotics supplementation on the hormone and body mass index in perimenopausal and postmenopausal women using the standardized diet. A 5-week double-blind, placebo-controlled, and randomized clinical study. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 3859–3867. [Google Scholar] [PubMed]
- Lei, K.; Li, Y.L.; Yu, D.Y.; Rajput, I.R.; Li, W.F. Influence of dietary inclusion of Bacillus licheniformis on laying performance, egg quality, antioxidant enzyme activities, and intestinal barrier function of laying hens. Poult. Sci. 2013, 92, 2389–2395. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, S.; Pang, Q.; Miao, Z. Bacillus amyloliquefaciens BLCC1-0238 Can Effectively Improve Laying Performance and Egg Quality Via Enhancing Immunity and Regulating Reproductive Hormones of Laying Hens. Probiotics Antimicrob. Proteins 2020, 12, 246–252. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Zhang, K.; Ding, X.; Celi, P.; Yan, L.; Bai, S.; Zeng, Q.; Mao, X.; Xu, S.; Wang, J. The impact of dietary supplementation of different feed additives on performances of broiler breeders characterized by different egg-laying rate. Poult. Sci. 2019, 98, 6091–6099. [Google Scholar] [CrossRef] [PubMed]
- Gioacchini, G.; Maradonna, F.; Lombardo, F.; Bizzaro, D.; Olivotto, I.; Carnevali, O. Increase of fecundity by probiotic administration in zebrafish (Danio rerio). Reproduction 2010, 140, 953–959. [Google Scholar] [CrossRef]
- Sun, C.; Groom, K.M.; Oyston, C.; Chamley, L.W.; Clark, A.R.; James, J.L. The placenta in fetal growth restriction: What is going wrong? Placenta 2020, 96, 10–18. [Google Scholar] [CrossRef]
- Jiménez, E.; Fernández, L.; Marín, M.L.; Martín, R.; Odriozola, J.M.; Nueno-Palop, C.; Narbad, A.; Olivares, M.; Xaus, J.; Rodríguez, J.M. Isolation of Commensal Bacteria from Umbilical Cord Blood of Healthy Neonates Born by Cesarean Section. Curr. Microbiol. 2005, 51, 270–274. [Google Scholar] [CrossRef]
- Rautava, S.; Collado, M.C.; Salminen, S.; Isolauri, E. Probiotics Modulate Host-Microbe Interaction in the Placenta and Fetal Gut: A Randomized, Double-Blind, Placebo-Controlled Trial. Neonatology 2012, 102, 178–184. [Google Scholar] [CrossRef]
- Yang, P.; Li, Z.; Tye, K.D.; Chen, Y.; Lu, T.; He, Z.; Zhou, J.; Xiao, X. Effects of an orally supplemented probiotic on the autophagy protein LC3 and Beclin1 in placentas undergoing spontaneous delivery during normal pregnancy. BMC Pregnancy Childbirth 2020, 20, 216. [Google Scholar] [CrossRef]
- Yeganegi, M.; Watson, C.S.; Martins, A.; Kim, S.O.; Reid, G.; Challis, J.R.G.; Bocking, A.D. Effect of Lactobacillus rhamnosus GR-1 supernatant and fetal sex on lipopolysaccharide-induced cytokine and prostaglandin-regulating enzymes in human placental trophoblast cells: Implications for treatment of bacterial vaginosis and prevention of preterm. Am. J. Obstet. Gynecol. 2009, 200, e1–e532. [Google Scholar] [CrossRef]
- Brantsaeter, A.L.; Myhre, R.; Haugen, M.; Myking, S.; Sengpiel, V.; Magnus, P.; Jacobsson, B.; Meltzer, H.M. Intake of Probiotic Food and Risk of Preeclampsia in Primiparous Women: The Norwegian Mother and Child Cohort Study. Am. J. Epidemiol. 2011, 174, 807–815. [Google Scholar] [CrossRef]
- Gu, X.L.; Li, H.; Song, Z.H.; Ding, Y.N.; He, X.; Fan, Z.Y. Effects of isomaltooligosaccharide and Bacillus supplementation on sow performance, serum metabolites, and serum and placental oxidative status. Anim. Reprod. Sci. 2019, 207, 52–60. [Google Scholar] [CrossRef]
- Jarrett, P.; Meczner, A.; Costeloe, K.; Fleming, P. Historical aspects of probiotic use to prevent necrotising enterocolitis in preterm babies. Early Hum. Dev. 2019, 135, 51–57. [Google Scholar] [CrossRef]
- Mirpuri, J.; Neu, J. Maternal microbial factors that affect the fetus and subsequent offspring. Semin. Perinatol. 2021, 45, 151449. [Google Scholar] [CrossRef]
- Rasool, A.; Alvarado-Flores, F.; O’Tierney-Ginn, P. Placental Impact of Dietary Supplements: More Than Micronutrients. Clin. Ther. 2021, 43, 226–245. [Google Scholar] [CrossRef]
- Böhmer, B.M.; Kramer, W.; Roth-Maier, D.A. Dietary probiotic supplementation and resulting effects on performance, health status, and microbial characteristics of primiparous sows. J. Anim. Physiol. Anim. Nutr. 2006, 90, 309–315. [Google Scholar] [CrossRef]
- Hayakawa, T.; Masuda, T.; Kurosawa, D.; Tsukahara, T. Dietary administration of probiotics to sows and/or their neonates improves the reproductive performance, incidence of post-weaning diarrhea and histopathological parameters in the intestine of weaned piglets. Anim. Sci. J. 2016, 87, 1501–1510. [Google Scholar] [CrossRef] [PubMed]
- Romano-Keeler, J.; Weitkamp, J.-H. Maternal influences on fetal microbial colonization and immune development. Pediatr. Res. 2015, 77, 189–195. [Google Scholar] [CrossRef]
- Saxelby, C. Cardiovascular Disease: Diet, Nutrition and Emerging Risk Factors. Nutr. Diet. 2006, 63, 189–190. [Google Scholar] [CrossRef]
- Frostegård, J. Immunity, atherosclerosis and cardiovascular disease. BMC Med. 2013, 11, 117. [Google Scholar] [CrossRef] [PubMed]
- Asada, Y.; Yamashita, A.; Sato, Y.; Hatakeyama, K. Thrombus Formation and Propagation in the Onset of Cardiovascular Events. J. Atheroscler. Thromb. 2018, 25, 653–664. [Google Scholar] [CrossRef] [PubMed]
- Paganelli, F.; Mottola, G.; Fromonot, J.; Marlinge, M.; Deharo, P.; Guieu, R.; Ruf, J. Hyperhomocysteinemia and Cardiovascular Disease: Is the Adenosinergic System the Missing Link? Int. J. Mol. Sci. 2021, 22, 1690. [Google Scholar] [CrossRef] [PubMed]
- Raygan, F.; Rezavandi, Z.; Bahmani, F.; Ostadmohammadi, V.; Mansournia, M.A.; Tajabadi-Ebrahimi, M.; Borzabadi, S.; Asemi, Z. The effects of probiotic supplementation on metabolic status in type 2 diabetic patients with coronary heart disease. Diabetol. Metab. Syndr. 2018, 10, 51. [Google Scholar] [CrossRef] [PubMed]
- Raygan, F.; Ostadmohammadi, V.; Bahmani, F.; Asemi, Z. The effects of vitamin D and probiotic co-supplementation on mental health parameters and metabolic status in type 2 diabetic patients with coronary heart disease: A randomized, double-blind, placebo-controlled trial. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2018, 84, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Raygan, F.; Ostadmohammadi, V.; Asemi, Z. The effects of probiotic and selenium co-supplementation on mental health parameters and metabolic profiles in type 2 diabetic patients with coronary heart disease: A randomized, double-blind, placebo-controlled trial. Clin. Nutr. 2019, 38, 1594–1598. [Google Scholar] [CrossRef] [PubMed]
- Tajabadi-Ebrahimi, M.; Sharifi, N.; Farrokhian, A.; Raygan, F.; Karamali, F.; Razzaghi, R.; Taheri, S.; Asemi, Z. A Randomized Controlled Clinical Trial Investigating the Effect of Synbiotic Administration on Markers of Insulin Metabolism and Lipid Profiles in Overweight Type 2 Diabetic Patients with Coronary Heart Disease. Exp. Clin. Endocrinol. Diabetes 2017, 125, 21–27. [Google Scholar] [CrossRef]
- Malik, M.; Suboc, T.M.; Tyagi, S.; Salzman, N.; Wang, J.; Ying, R.; Tanner, M.J.; Kakarla, M.; Baker, J.E.; Widlansky, M.E. Lactobacillus plantarum 299v Supplementation Improves Vascular Endothelial Function and Reduces Inflammatory Biomarkers in Men With Stable Coronary Artery Disease. Circ. Res. 2018, 123, 1091–1102. [Google Scholar] [CrossRef]
- Koppinger, M.P.; Lopez-Pier, M.A.; Skaria, R.; Harris, P.R.; Konhilas, J.P. Lactobacillus reuteri attenuates cardiac injury without lowering cholesterol in low-density lipoprotein receptor-deficient mice fed standard chow. Am. J. Physiol. Heart Circ. Physiol. 2020, 319, H32–H41. [Google Scholar] [CrossRef]
- Mengheri, E. Health, probiotics, and inflammation. J. Clin. Gastroenterol. 2008, 42 (Suppl. S3), S177–S178. [Google Scholar] [CrossRef]
- Smelt, M.J.; de Haan, B.J.; Bron, P.A.; van Swam, I.; Meijerink, M.; Wells, J.M.; Faas, M.M.; de Vos, P. Probiotics can generate FoxP3 T-cell responses in the small intestine and simultaneously inducing CD4 and CD8 T cell activation in the large intestine. PLoS ONE 2013, 8, e68952. [Google Scholar] [CrossRef]
- Okamoto, N.; Noma, T.; Ishihara, Y.; Miyauchi, Y.; Takabatake, W.; Oomizu, S.; Yamaoka, G.; Ishizawa, M.; Namba, T.; Murakami, K.; et al. Prognostic value of circulating regulatory T cells for worsening heart failure in heart failure patients with reduced ejection fraction. Int. Heart J. 2014, 55, 271–277. [Google Scholar] [CrossRef]
- Danilo, C.A.; Constantopoulos, E.; McKee, L.A.; Chen, H.; Regan, J.A.; Lipovka, Y.; Lahtinen, S.; Stenman, L.K.; Nguyen, T.-V.V.; Doyle, K.P.; et al. Bifidobacterium animalis subsp. lactis 420 mitigates the pathological impact of myocardial infarction in the mouse. Benef. Microbes 2017, 8, 257–269. [Google Scholar] [CrossRef]
- Arpaia, N.; Campbell, C.; Fan, X.; Dikiy, S.; van der Veeken, J.; DeRoos, P.; Liu, H.; Cross, J.R.; Pfeffer, K.; Coffer, P.J.; et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature 2013, 504, 451–455. [Google Scholar] [CrossRef]
- Wang, L.; de Zoeten, E.F.; Greene, M.I.; Hancock, W.W. Immunomodulatory effects of deacetylase inhibitors: Therapeutic targeting of FOXP3+ regulatory T cells. Nat. Rev. Drug Discov. 2009, 8, 969–981. [Google Scholar] [CrossRef] [PubMed]
- Gan, X.T.; Ettinger, G.; Huang, C.X.; Burton, J.P.; Haist, J.V.; Rajapurohitam, V.; Sidaway, J.E.; Martin, G.; Gloor, G.B.; Swann, J.R.; et al. Probiotic administration attenuates myocardial hypertrophy and heart failure after myocardial infarction in the rat. Circ. Heart Fail. 2014, 7, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Gupta, D.; Mehrotra, R.; Mago, P. Psychobiotics: The Next-Generation Probiotics for the Brain. Curr. Microbiol. 2021, 78, 449–463. [Google Scholar] [CrossRef] [PubMed]
- Aizawa, E.; Tsuji, H.; Asahara, T.; Takahashi, T.; Teraishi, T.; Yoshida, S.; Ota, M.; Koga, N.; Hattori, K.; Kunugi, H. Possible association of Bifidobacterium and Lactobacillus in the gut microbiota of patients with major depressive disorder. J. Affect. Disord. 2016, 202, 254–257. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Wang, K.; Hu, J. Effect of Probiotics on Depression: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Nutrients 2016, 8, 483. [Google Scholar] [CrossRef] [PubMed]
- Kelly, J.R.; Allen, A.P.; Temko, A.; Hutch, W.; Kennedy, P.J.; Farid, N.; Murphy, E.; Boylan, G.; Bienenstock, J.; Cryan, J.F.; et al. Lost in translation? The potential psychobiotic Lactobacillus rhamnosus (JB-1) fails to modulate stress or cognitive performance in healthy male subjects. Brain Behav. Immun. 2017, 61, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Allen, A.P.; Hutch, W.; Borre, Y.E.; Kennedy, P.J.; Temko, A.; Boylan, G.; Murphy, E.; Cryan, J.F.; Dinan, T.G.; Clarke, G. Bifidobacterium longum 1714 as a translational psychobiotic: Modulation of stress, electrophysiology and neurocognition in healthy volunteers. Transl. Psychiatry 2016, 6, e939. [Google Scholar] [CrossRef] [PubMed]
- Savignac, H.M.; Kiely, B.; Dinan, T.G.; Cryan, J.F. Bifidobacteria exert strain-specific effects on stress-related behavior and physiology in BALB/c mice. Neurogastroenterol. Motil. 2014, 26, 1615–1627. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, K.; Itoh, N.; Yamamoto, S.; Higo-Yamamoto, S.; Nakakita, Y.; Kaneda, H.; Shigyo, T.; Oishi, K. Dietary heat-killed Lactobacillus brevis SBC8803 promotes voluntary wheel-running and affects sleep rhythms in mice. Life Sci. 2014, 111, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Yamamura, S.; Morishima, H.; Kumano-go, T.; Suganuma, N.; Matsumoto, H.; Adachi, H.; Sigedo, Y.; Mikami, A.; Kai, T.; Masuyama, A.; et al. The effect of Lactobacillus helveticus fermented milk on sleep and health perception in elderly subjects. Eur. J. Clin. Nutr. 2009, 63, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.-W.; Park, J.G.; Ilhan, Z.E.; Wallstrom, G.; Labaer, J.; Adams, J.B.; Krajmalnik-Brown, R. Reduced incidence of Prevotella and other fermenters in intestinal microflora of autistic children. PLoS ONE 2013, 8, e68322. [Google Scholar] [CrossRef] [PubMed]
- Shaaban, S.Y.; El Gendy, Y.G.; Mehanna, N.S.; El-Senousy, W.M.; El-Feki, H.S.A.; Saad, K.; El-Asheer, O.M. The role of probiotics in children with autism spectrum disorder: A prospective, open-label study. Nutr. Neurosci. 2018, 21, 676–681. [Google Scholar] [CrossRef] [PubMed]
- Cenit, M.C.; Sanz, Y.; Codoñer-Franch, P. Influence of gut microbiota on neuropsychiatric disorders. World J. Gastroenterol. 2017, 23, 5486–5498. [Google Scholar] [CrossRef] [PubMed]
- Dinan, T.G.; Borre, Y.E.; Cryan, J.F. Genomics of schizophrenia: Time to consider the gut microbiome? Mol. Psychiatry 2014, 19, 1252–1257. [Google Scholar] [CrossRef]
- Cenit, M.C.; Nuevo, I.C.; Codoñer-Franch, P.; Dinan, T.G.; Sanz, Y. Gut microbiota and attention deficit hyperactivity disorder: New perspectives for a challenging condition. Eur. Child Adolesc. Psychiatry 2017, 26, 1081–1092. [Google Scholar] [CrossRef]
- Sampson, T.R.; Debelius, J.W.; Thron, T.; Janssen, S.; Shastri, G.G.; Ilhan, Z.E.; Challis, C.; Schretter, C.E.; Rocha, S.; Gradinaru, V.; et al. Gut Microbiota Regulate Motor Deficits and Neuroinflammation in a Model of Parkinson’s Disease. Cell 2016, 167, 1469–1480.e12. [Google Scholar] [CrossRef]
- Everard, A.; Cani, P.D. Diabetes, obesity and gut microbiota. Best Pract. Res. Clin. Gastroenterol. 2013, 27, 73–83. [Google Scholar] [CrossRef]
- Harach, T.; Jammes, F.; Muller, C.; Duthilleul, N.; Cheatham, V.; Zufferey, V.; Cheatham, D.; Lukasheva, Y.A.; Lasser, T.; Bolmont, T. Administrations of human adult ischemia-tolerant mesenchymal stem cells and factors reduce amyloid beta pathology in a mouse model of Alzheimer’s disease. Neurobiol. Aging 2017, 51, 83–96. [Google Scholar] [CrossRef] [PubMed]






| Name of the Probiotic Strains | Age of the Participants | Dose of the Probiotics | Duration of Study | Outcomes of the Study | References |
|---|---|---|---|---|---|
| Probiotics Fortify the Respiratory Tract | |||||
| Lactobacillus acidophilus, Lactobacillus casei, Lactobacillus bulgaricus, Lactobacillus rhamnosus, Streptococcus thermophilus Bifidobacterium longum, Bifidobacterium breve, | 9–53 years | 1 × 109 CFU/g | 60 days | Slightly increased expression of forkhead box P3 (FoxP3); transforming growth factor-β (TGF-β) and interferon-gamma (IFN-γ); and decreased interleukin (IL)-10 and IL-4 expression. Significantly decreased IL-17 values after synbiotic intake, showing controlled immunopathogenesis of allergic rhinitis. | [183] |
| Bifidobacterium breve M-16, Bifidobacterium infantis M-63, Bifidobacterium longum BB536 | 4–17 years | 1 × 109, 1 × 109, 3 × 109 CFU/g | 56 days | Significantly elevated quality of life (QoL) and alleviated nasal symptoms in children with allergic rhinitis. | [184] |
| Lactobacillus acidophilus, Lactobacillus casei | 18–21 years | - | 28 days | Significantly elevated IFN-γ levels and decreased IL-4 levels, and significant difference among the IFN-γ:IL-4 exhibiting improved IFN-γ/IL-4 levels in patients with allergic rhinitis. | [185] |
| Bifidobacterium lactis NCC2818 | 20–65 years | 2 × 109 CFU/g | 56 days | Significantly lowered Th-2 cytokine concentrations (IL-3 and IL-5), nasal symptom scores, and activated-CD63-expressing basophils, exhibiting the capacity of NCC2818 to mitigate allergic symptoms and immune parameters. | [186] |
| Lactobacillus Paracasei 33 | 0.5–5 years | 2 × 109 CFU/g | 42 days | Significantly improved nasal blocking, sneezing, rhinorrhea, feeding and sleeping difficulties, and coughing, showing the equal effectiveness of Lactobacillus paracasei 33 as cetirizine in children with perennial allergic rhinitis without any significant side effects. | [187] |
| Bifidobacterium breve B632 Ligilactobacillus salivarius LS01 | 3–14 years | 1 × 109 CFU/g each | 112 days | The significant reduction in asthmatic exacerbations demonstrates the effectiveness and safety of Bifidobacterium breve B632 and Ligilactobacillus salivarius LS01, proving that they are auxiliary remedies. | [188] |
| Lactobacillus acidophilus, Lactobacillus rhamnosus, Lactobacillus bulgaricus, Lactobacillus casei, Streptococcus thermophiles, Bifidobacterium longum, Bifidobacterium breve, | 38.62 ± 10.49 years | 3 × 109, 7 × 109, 5 × 108, 3 × 109, 3 × 108, 1 × 109, and 2 × 1010 CFU/g | 60 days | Significant reduction in IL-4, miR146-a, and miR-16 levels and elevation in miR-133b level displayed significantly improved forced vital capacity (FVC) and forced expiratory volume for 1s (FEV1). | [189] |
| Lactobacillus salivarius PM-A0006 | 10–12 years | 2 × 109 CFU/g | 112 days | Significantly improved pulmonary functioning parameters, such as FVC, FEV1, and FEV1:FVC ratio, and significantly decreased use the of inhaled corticosteroids and short-acting inhaled bronchodilators, as well as the diminished use of medicine in asthmatic children. | [190] |
| Streptococcus thermophilus, Lactobacillus bulgaricus, Lactobacillus acidophilus, Lactobacillus rhamnosus, Lactobacillus casei, Bifidobacterium infantis, Bifidobacterium breve | 6–12 years | 1 × 109 CFU/g each | 60 days | Significantly lowered the number of viral respiratory infections and decreased the use of Salbutamol. | [191] |
| Lactobacillus bulgaris, Lactobacillus Casei, Streptococcus thermophiles, Lactobacillus acidophilus, Bifidobacterium breve, Lactobacillus rhamnosus, Bifidobacterium infantis, | ≤12 years | Manufacturer defined recipe | 180 days | Significantly alleviated asthma symptoms and decreased outpatient visits with rare side effects, exhibiting positive effects on the QoL of asthmatic patients. | [192] |
| Probiotics Meliorate Bone Health | |||||
| Bacillus subtilis C-3102 | 50–69 years | 3.4 × 109 CFU/g | 168 days | Significant increase in the Bifidobacterium genus and a decrease in the Fusobacterium genus. Significant increase in hip BMD and decrease in bone resorption markers such as urinary type I collagen cross-linked N-telopeptide (uNTx) and tartrate-resistant acid phosphatase isoform 5b (TRACP-5b), suggesting improved BMD by gut microbiota modulation and bone resorption inhibition in postmenopausal women. | [193] |
| Streptococcus thermophiles, Bifidobacterium breve, Bifidobacterium longum, Lactobacillus acidophilus, Lactobacillus bulgaricus, Lactobacillus rhamnosus, Lactobacillus casei, | 50–72 years | 3 × 108, 2 × 1010, 1 × 1011, 3 × 1010, 5 × 108, 7 × 109, and 2.6 × 1010 CFU/g | 180 days | Significant decrease in cross-linked C-telopeptide (CTX) and bone-specific alkaline phosphatase (BALP), TNF-α, and serum parathyroid hormone (PTH) levels. | [194] |
| Lactobacillus plantarum DSM 15313 and DSM 15312, Lactobacillus paracasei DSM 13434 | 59.1 ± 3.8 years | 1 × 1010 CFU/capsule | 365 days | Significantly reduced lumbar spine bone mineral density (LS-BMD) losses. | [195] |
| Probiotics Preserve Kidney Integrity | |||||
| Bifidobacterium longum A101 Bifidobacterium bifidum A218 Lactobacillus rhamnosus, Lactobacillus Plantarum A87 | 22–69 years | 1 × 109 CFU/g each | 90 days | Significantly decreased serum syndecan-1 and blood glucose levels, indicating improved metabolism and systemic inflammation reduction in chronic kidney disease patients. | [196] |
| Lactobaccillus pentosus LPE588, Lactobaccillus salivarius LS159, Lactococcus lactis subsp. lactis LL358 | 39–75 years | 1 × 1011 CFU/g | 180 days | Significantly decreased indoxyl sulfate levels in serum in patients on hemodialysis (HD), with slight changes in serum p-cresyl sulfate, blood urea nitrogen, hemoglobin levels, blood glucose, microbial and inflammatory translocation markers. | [197] |
| Lactobacillus acidophilus, Streptococcus thermophilus, Bifidobacterium longum | ≥18 years | 9 × 1010 CFU/capsule | 7 days | Significantly decreased blood urea values. | [198] |
| Streptococcus thermophilus, Lactobacillus bulgaricus, Bifidobacterium longum | 18–75 years | 1 × 109 CFU/g | 60 days | Significantly decreased high-sensitivity C-reactive protein (hs-CRP) and IL-6 levels and increased triceps skinfold thickness, upper arm circumference, and serum albumin levels in peritoneal dialysis patients, leading to higher social and physical functioning, as well as improved QoL and malnutrition. | [199] |
| Lactobacillus rhamnosus HN-001 and LR-32, Enterococcus faecium UBEF-41, Saccharomyces cerevisiae subspecies Boulardii MTCC-5375, Lactobacillus acidophilus LA-14, Bifidobacterium longum BL05, Bifidobacterium bifidum BB06, Bifidobacterium brevis BB03 | >18 years | Manufacturer’s recipe | 111 days | Significantly elevated fecal Bifidobacteria and Lactobacillales concentrations and diminished 3-methyl-indole (3-MI) and urinary indican levels. Significantly improved levels of serum calcium, ferritin, iron, C-reactive protein, transferrin saturation, serum intact parathormone (iPTH), and β2-microglobulin. | [200] |
| Lactobacillus casei, Bifidobacterium lactis, Lactobacillus acidophilus, | ≥18 years | 3.2 × 1010 CFU/g | 84 days | Significantly improved Subdoligranulum, Lactobacillus, and Bifidobacteria genera and glomerular filtration rate. Decreased serum IS and hr-CRP levels, exhibiting decreased microinflammation and uremic toxins levels in patients with chronic kidney disease. | [201] |
| Lactobacillus rhamnosus GG | 70.15 ± 12.3 years | 3.5 × 1011 CFU/g | 56 days | Significantly lowered the serum P-cresol sulfate (PCS) and albuminuria and improved estimated glomerular filtration rate (eGFR), serum creatinine (SCr), blood urea nitrogen (BUN), and proteinuria. Significantly reduced pathogenic Proteobacteria and elevated Firmicutes and Actinobacteriota count, suggesting gut microbiota melioration. | [202] |
| Streptococcus thermophilus Lactobacillus acidophilus, Bifidobacterium bifidum, | 18–75 years | 4.3 × 109, 4.2 × 109, and 1.2 × 109 CFU/g | 84 days | Significantly reduced fasting blood glucose, mAlb/Cr, and HbA1c and slightly decreased 2h postprandial blood glucose level and eGFR while somewhat elevating mAlb/Cr levels, suggesting ameliorated glycemic control in diabetic nephropathy patients. | [203] |
| Bifidobacterium bifidum, Bifidobacterium longum Bifidobacterium lactis, Lactobacillus acidophilus, | 30–65 years | 2.7 × 107 CFU/g each | 84 days | Significantly decreased beck depression inventory (BDI) and beck anxiety inventory (BAI) and significantly increased serum hemoglobin (Hb) levels. | [204] |
| Enterococcus faecalis YIT0072 Lactobacillus acidophilus YIT2004, Bifidobacterium longum, | 18–70 years | 1.1 × 109, 0.53 × 109, and 2.2 × 109 CFU/g | 180 days | Significantly restored Bacteroidaceae and Enterococcaceae, and reduced Clostridiales Family XIII. Incertae Sedis, Erysipelotrichaceae, Peptostreptococcaceae, Ruminococcaceae, and Halomonadaceae in non-diabetic hemodialysis patients. Significantly reduced uremic retention solutes, including 1-methylinosine, 3-guanidinopropionic acid, and indole-3-acetic acid-O-glucuronide, in feces or serum. | [205] |
| Bifidobacterium lactis BIA-6, Bifidobacterium longum LAF-5, Lactobacillus acidophilus T16, Bifidobacterium bifidum BIA-6 | 30–65 years | 2.7 × 107 CFU/g each | 84 days | Significantly altered IL-6 and hs-CRP and significantly changed anti-HSP70 after synbiotic intake, exhibiting improved anti-HSP70 serum levels, endotoxin, and inflammatory markers. | [206] |
| Probiotics Preclude the Prevalence of Cardiovascular Diseases | |||||
| Lactobacillus rhamnosus GG | 56.70 ± 9.10 years | 1.6 × 109 CFU/g | 90 days | Significantly decreased serum TGF-β and trimethylamine N-oxide (TMAO) levels. Sightly differed matrix metalloproteinase-9 (MMP-9) and procollagen III levels and improved echocardiographic indices. Left ventricular ejection fraction (LVEF) and variation in procollagen III predicted 62% final LVEF levels, exhibiting beneficial effects on cardiac remodeling in myocardial infarction patients. | [207] |
| Bifidobacterium lactis, Bb12, Lactobacillus acidophilus La-5 | 30–70 years | 1 × 107 CFU/g each | 70 days | Significantly decreased oxidized low-density lipoprotein (ox-LDL) and apolipoprotein B100 (ApoB100) and slightly changed N-terminal pro-brain natriuretic peptide (NT-proBNP) and pentraxin3 (PTX3), suggesting improvements in the oxidative status of congestive heart failure (CHF) patients. | [208] |
| Lactobacillus rhamnosus GG | 30–70 years | 1.6 × 109 CFU/g | 84 days | Significantly decreased low-density lipoprotein cholesterol and total cholesterol, with slight differences in blood pressure and MetS feature indices suggesting decreased cardiovascular risk factors. | [209] |
| Streptococcus thermophiles, Bifidobacterium longum, Lactobacillus acidophilus, Lactobacillus casei, Lactobacillus bulgaricus, Bifidobacterium breve, Lactobacillus rhamnosus, | 30–70 years | 1.5 × 108, 5 × 108, 1.5 × 1010, 1 × 109, 2.5 × 108, 1 × 1010, and 3.5 × 109 CFU/g | 70 days | Significantly decreased NT-proBNP levels and hindered the elevation of hs-CRP, exhibiting the advantageous effects of synbiotics on inflammatory status. | [210] |
| Lactobacillus rhamnosus GG | 8–85 years | 1.9 × 109 CFU/g | 60 days | Significant reductions in BDI, Spielberger state–trait anxiety inventory (STAI) state, and STAI-trait scores, lipopolysaccharide (LPS), TNF-α, and hs-CRP, suggesting beneficial health impacts on inflammatory biomarkers, anxiety, and depression. | [211] |
| Lactobacillus paracasei LPC-37, Bifidobacterium lactis HN019, Lactobacillus acidophilus NCFM, Lactobacillus rhamnosus HN001 | 20–50 years | 1 × 109 CFU/g each | 56 days | Significant reduction in cholesterol and fasting glucose levels while elevating HDL-cholesterol. Slightly lowered systolic BP and diastolic BP and reduced low-frequency (LF) oscillation and LF/high-frequency (HF) ratio, suggesting improved autonomic modulation and lipid profiles in hypertensive women. | [212] |
| Lactobacillus rhamnosus GG | 56.70 ± 9.10 years | 1.6 × 109 CFU/g | 84 days | Significantly decreased IL1-Beta and LPS levels and significantly meliorated cardiovascular-related factors, suggesting advantageous impacts on mega inflammation and metabolic endotoxemia in coronary artery disease patients. | [213] |
| Probiotics Influence the Male and Female Reproductive Systems | |||||
| Lactobacillus casei DG | 18–45 years | 2.4 × 1010 CFU/g | 90 days | Significantly changed QoL, International Prostate Symptom Score (IPSS), and NIH Chronic Prostatitis Symptom Index (NIH-CPSI) and significantly decreased antibiotic use and symptomatic recurrence. | [214] |
| Escherichia coli Nissle 1917 | ≥ 18 year | 2.5–25 × 109 CFU/g | 84 days | Significantly lowered biological recurrence rate and NIH-CPSI score, exhibiting efficiently controlled and diminished biological recurrences in chronic bacterial prostatitis patients. | [215] |
| Lactobacillus acidophilus, Streptococcus thermophiles, Lactobacillus casei, Bifidobacterium longum, Lactobacillus bulgaricus, Bifidobacterium breve, Lactobacillus rhamnosus, | 34.5 years (mean) | 1 × 109 CFU/g | 80 days | Significantly meliorated normal morphology, motility, sperm concentration, DNA fragmentation, and sperm lipid peroxidation. | [216] |
| Bifidobacterium longum, Streptococcus thermophilus, Lactobacillus rhamnosus, Lactobacillus acidophilus, Bifidobacterium breve, Lactobacillus casei, Lactobacillus bulgaricus, | ≥ 18 years | 1 × 109 CFU/capsule | 90 days | Significantly improved sperm concentration and normal morphology and slightly improved sperm motility and volume after varicocelectomy, demonstrating the benefits of probiotics in improving semen parameters. | [217] |
| Lactobacillus rhamnosus MG4288, Lactobacillus fermentum MG901, Lactobacillus paracasei MG4272, Lactobacillus plantarum MG989, Lactobacillus salivarius MG242, | 19–50 years | 1.0 × 109 CFU/g each | 84 days | Significant reduction in Nugent score, a substantial increase in Lactobacillus plantarum in the vagina, and suppression in pathogenic bacteria such as Atopobium vaginae, Gardnerella vaginalis, and Mobiluncus spp., exhibiting bacterial vaginosis (BV) alleviation. | [218] |
| Lactobacillus acidophilus LA-5 | 18–49 years | 1 × 109 CFU/g | 65 days | Lactobacillusacidophilus supplementation in treating vulvovaginal candidiasis (VVC) and decreasing negative culture, vulvovaginal erythema, and abnormal discharge, dyspareunia, dysuria, frequent urination, and burning is enormously comparable to fluconazole. | [219] |
| Lactobacillus fermentum LF26, Lactobacillus delbrueckii subsp. lactis LDL114, Lactobacillus rhamnosus LRH10, Lactobacillus plantarum LP115, Lactobacillus. paracasei LPC12, Lactobacillus. helveticus LA25 | 28.95 ± 0.70 years | 3.2 × 109 CFU | 56 days | Significantly reduced discharge, burning, and irritation, along with reduced vulvovaginal symptoms, recurrences of VC, recurrences of social and emotional stress, exhibiting the alleviation of disease and increased defecation times per week, showing a reduced risk of pregnancy-induced constipation. | [220] |
| Lactobacillus crispatus DSM32716, DSM32717, DSM32718, DSM32720, | 18–50 years | 3 × 1010 CFU/capsule | 90 days | Significantly reduced symptoms of BV and VVC, remarkable improvements in the smell and amount of discharge, Nugent score, and irritation/itching. Significantly improved vaginal lactobacilli counts and decreased BV-related bacteria. | [221] |
| Probiotics Alleviate Neurodegenerative and Neurodevelopmental Disorders | |||||
| Lactobacillus plantarum P8 | 31.7 ± 11.1 years | 1 × 1010 CFU/g | 84 days | Significantly reduced stress and anxiety. Slightly altered plasma cortisol levels and significantly reduced TNF-α and IFN-γ. Improved cognitive and memory traits, including verbal learning and memory and social-emotional cognition, in stressed adults. | [222] |
| Bifidobacterium longum R0175 or Lactobacillus rhamnosus HA-114 | 50–90 years | 1 × 105 CFU/g | 84 days | Significant improvement in cognition in patients with Alzheimer’s disease. | [223] |
| Bifidobacterium breve, Bifidobacterium infantis, Lactobacillus plantarum, Lactobacillus acidophilus, Streptococcus thermophilus, Lactobacillus casei, Bifidobacterium longum, Lactobacillus delbrueckii subsp. bulgaricus | 3–12 years | 9 × 109 CFU/g | 56 days | Significantly improved pediatric quality of life inventory (PedsQL) and GI complaints in children on the autism spectrum. | [224] |
| Lactococcus lactis W58, Lactococcus lactis W19, Bifidobacterium lactis W51 and W52, Bifidobacterium bifidum W23, Lactobacillus brevis W63, Lactobacillus acidophilus W37, Lactobacillus salivarius W24, Lactobacillus casei W5 | ~21 years | 2.5 × 109 CFU/g | 28 days | Significant improvement in memory performance, attributed to the neural changes in the frontal cortex occurred during cognitive control interventions. Implemented measures to mitigate the adverse impacts of stress on cognitive function. | [225] |
| Bifidobacterium longum BIA-8, Bifidobacterium lactis BIA-7, Bifidobacterium bifidum BIA-6, Lactobacillus acidophilus T16 | 6.64 ± 10.69 years | 2.7 × 107 CFU/g each | 84 days | Significantly decreased hospital anxiety and depression scale (HADS) depression scores, along with significant changes in HADS-ANX scores. Significantly increased the serum brain-derived neurotrophic factor in patients with depression. | [226] |
| Lactobacillus casei Shirota | 19–22 years | 3 × 1010 CFU/g | 42 days | Significant decrease in stress and anxiety levels of participants and a significant improvement in their aerobic capacities. | [227] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Virk, M.S.; Virk, M.A.; He, Y.; Tufail, T.; Gul, M.; Qayum, A.; Rehman, A.; Rashid, A.; Ekumah, J.-N.; Han, X.; et al. The Anti-Inflammatory and Curative Exponent of Probiotics: A Comprehensive and Authentic Ingredient for the Sustained Functioning of Major Human Organs. Nutrients 2024, 16, 546. https://doi.org/10.3390/nu16040546
Virk MS, Virk MA, He Y, Tufail T, Gul M, Qayum A, Rehman A, Rashid A, Ekumah J-N, Han X, et al. The Anti-Inflammatory and Curative Exponent of Probiotics: A Comprehensive and Authentic Ingredient for the Sustained Functioning of Major Human Organs. Nutrients. 2024; 16(4):546. https://doi.org/10.3390/nu16040546
Chicago/Turabian StyleVirk, Muhammad Safiullah, Muhammad Abdulrehman Virk, Yufeng He, Tabussam Tufail, Mehak Gul, Abdul Qayum, Abdur Rehman, Arif Rashid, John-Nelson Ekumah, Xu Han, and et al. 2024. "The Anti-Inflammatory and Curative Exponent of Probiotics: A Comprehensive and Authentic Ingredient for the Sustained Functioning of Major Human Organs" Nutrients 16, no. 4: 546. https://doi.org/10.3390/nu16040546
APA StyleVirk, M. S., Virk, M. A., He, Y., Tufail, T., Gul, M., Qayum, A., Rehman, A., Rashid, A., Ekumah, J.-N., Han, X., Wang, J., & Ren, X. (2024). The Anti-Inflammatory and Curative Exponent of Probiotics: A Comprehensive and Authentic Ingredient for the Sustained Functioning of Major Human Organs. Nutrients, 16(4), 546. https://doi.org/10.3390/nu16040546