

The Lactobacillus plantarum P-8 Probiotic Microcapsule Prevents DSS-Induced Colitis through Improving Intestinal Integrity and Reducing Colonic Inflammation in Mice

Abstract
1. Introduction
2. Materials and Methods
2.1. Microcapsule Preparation Method
2.1.1. Preparation of Bacterial Suspension of L. plantarum P-8
2.1.2. Encapsulation of L. plantarum P-8
2.2. Physical and Chemical Properties of Microcapsules
2.2.1. Survival Rate after Microencapsulation
2.2.2. Characterization of Microcapsules by Scanning Electron Microscopy (SEM)
2.2.3. Survival Rate of Free Probiotic and Probiotic Microcapsules in Simulated Gastrointestinal Conditions
2.2.4. Survival Rate of Free Probiotic and Probiotic Microcapsules in High Temperature Conditions
2.3. Animal Experimental Design and DSS Induced Colitis Model in Mice
2.3.1. Disease Activity Index (DAI)
2.3.2. Histopathology Analysis
2.3.3. Immunofluorescence
2.3.4. Immunohistochemistry
2.3.5. Enzyme-Linked Immunosorbent Assay (ELISA)
2.3.6. Western Blot
2.4. Statistical Analysis
3. Results
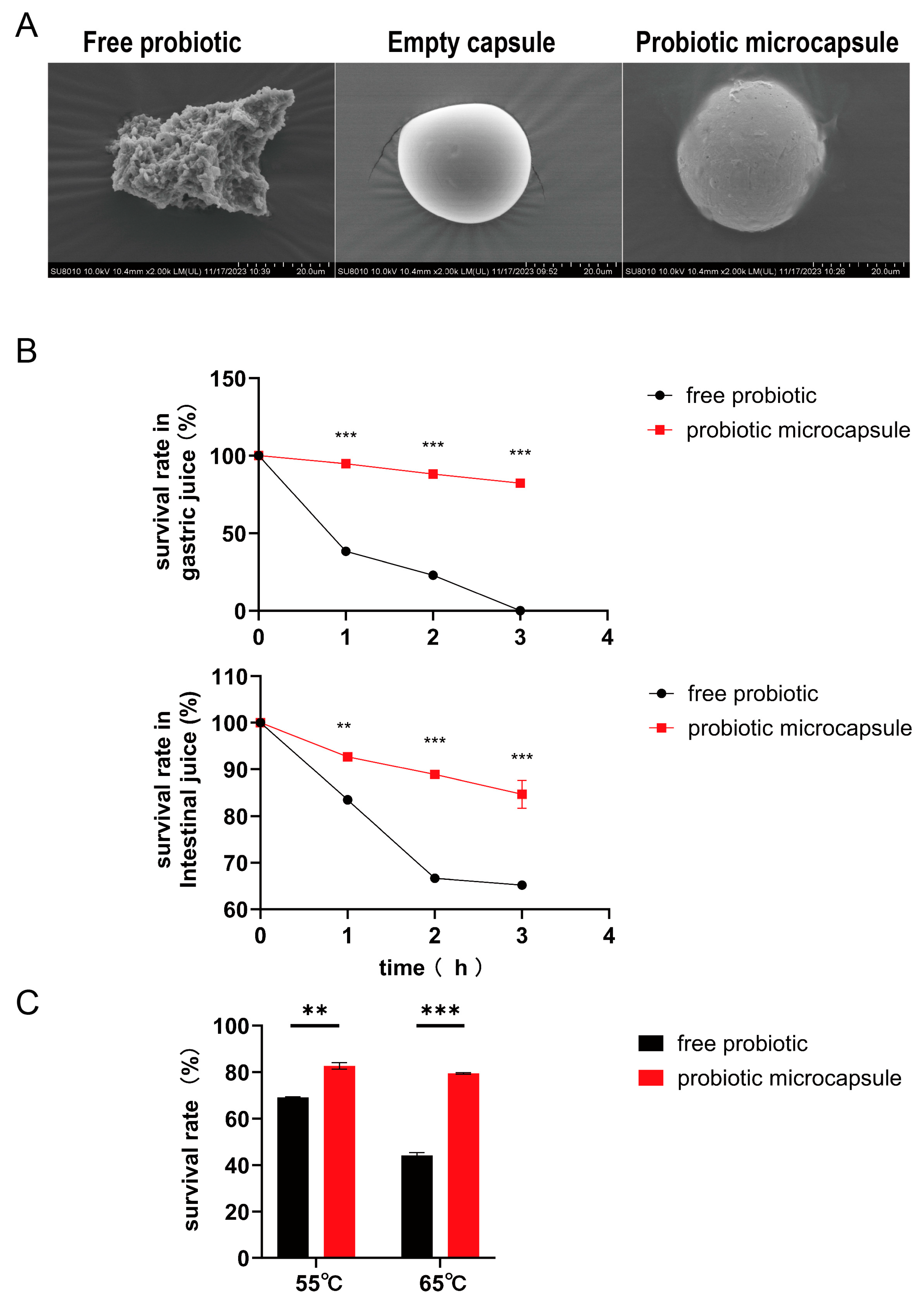
3.1. Morphological Characterisation of Microcapsules and the Survival Rate under Various Conditions
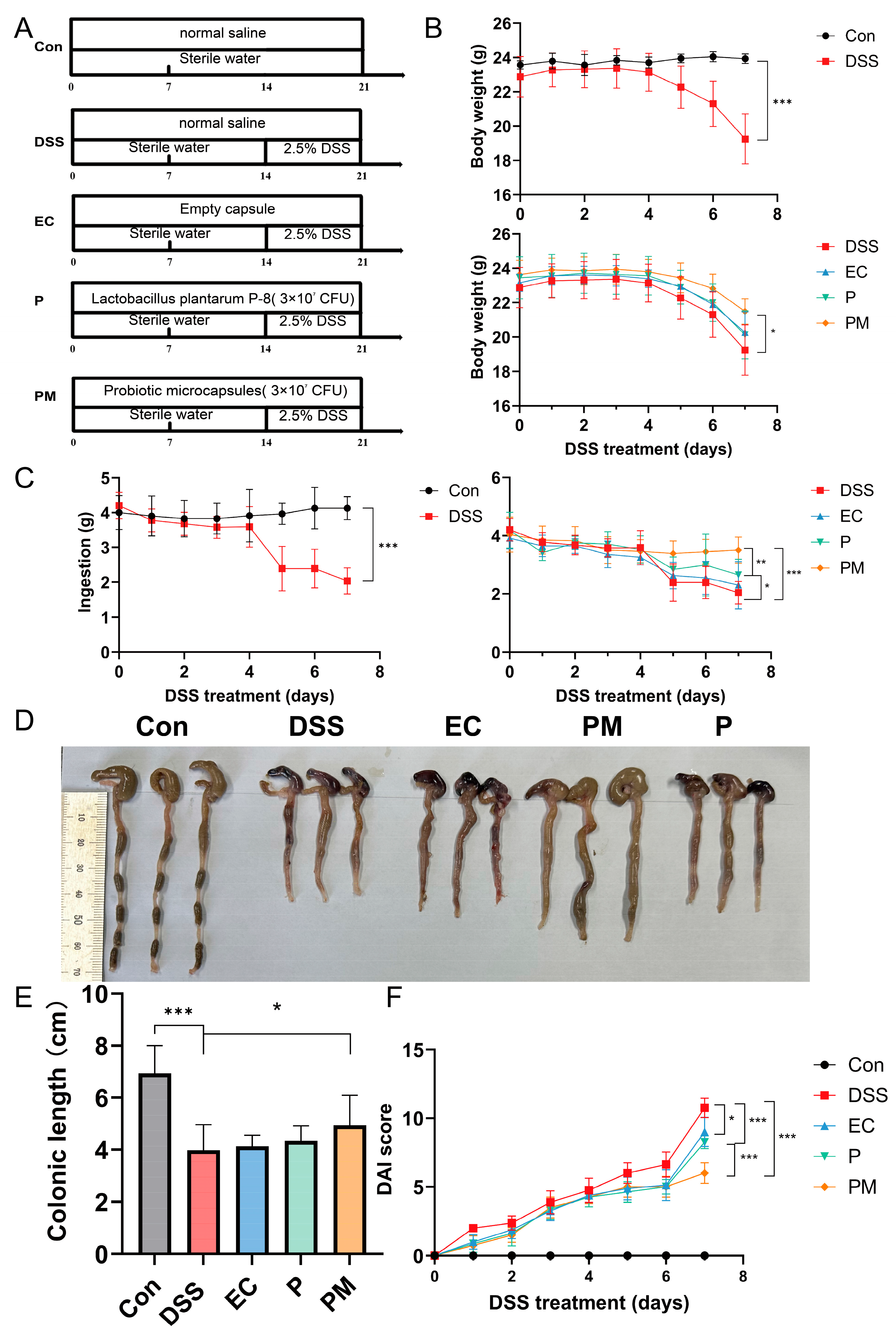
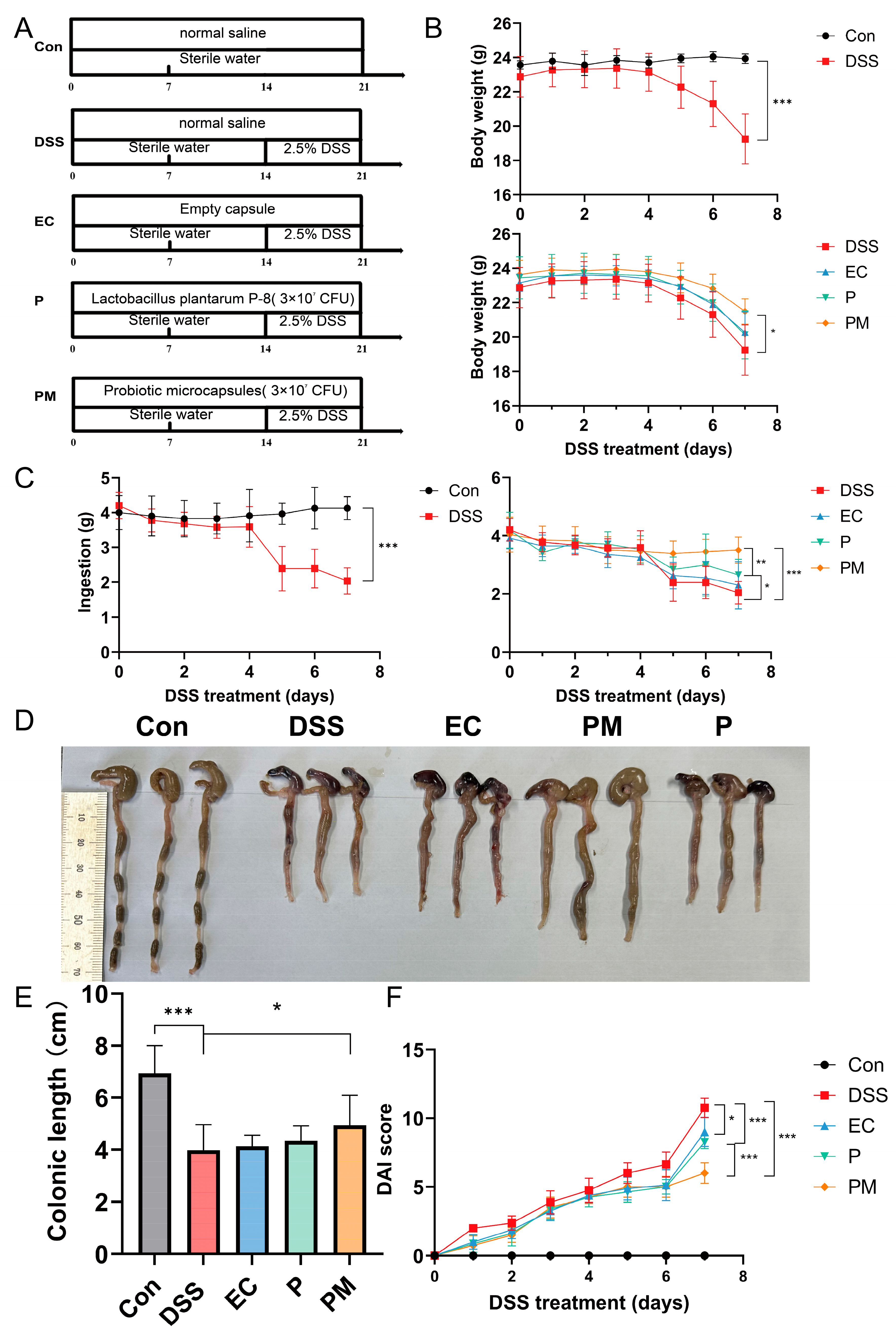
3.2. Probiotic Microcapsule Administration Alleviated Colitis
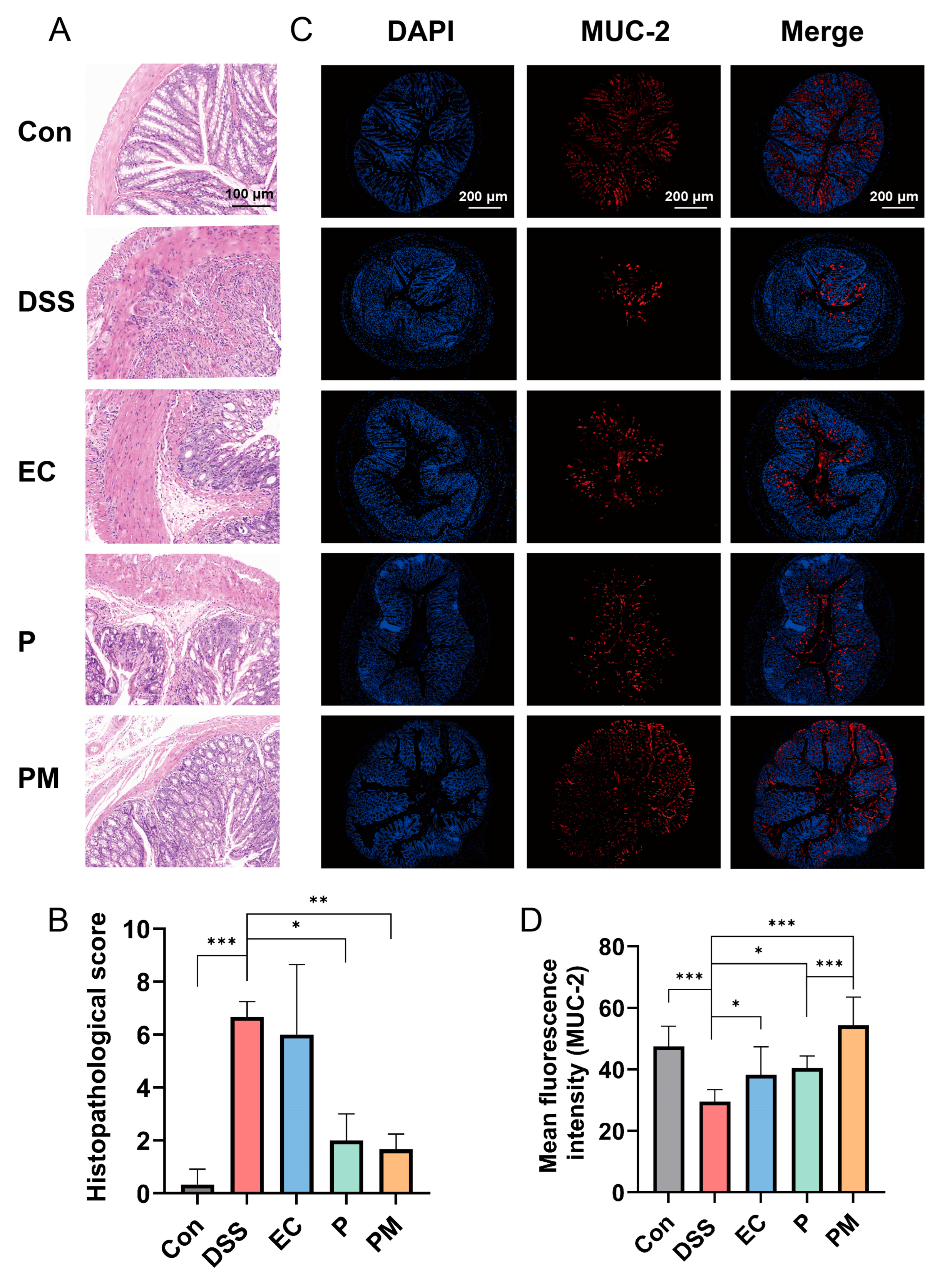
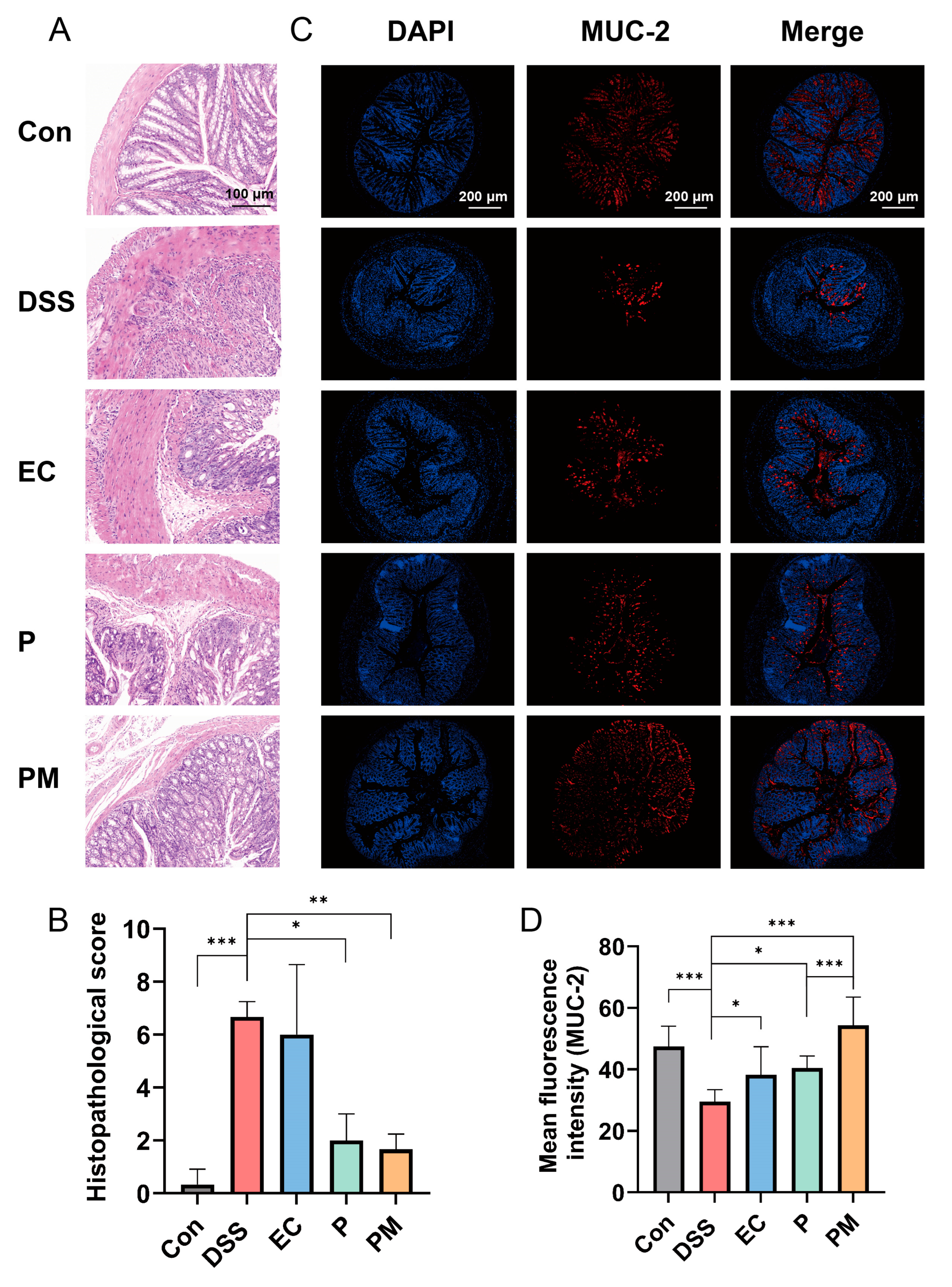
3.3. Probiotic Microcapsules Alleviated Colonic Mucosal Injury
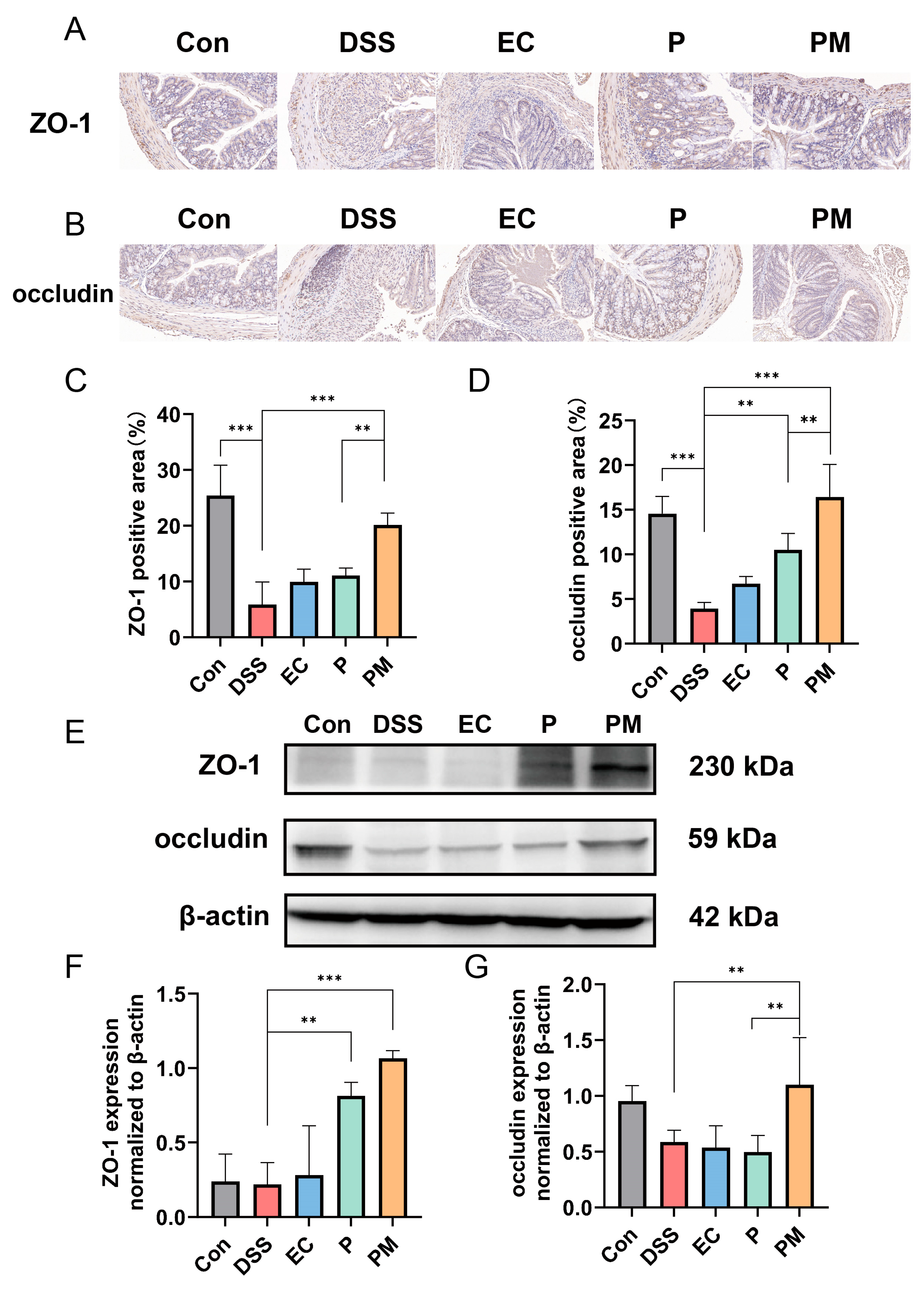
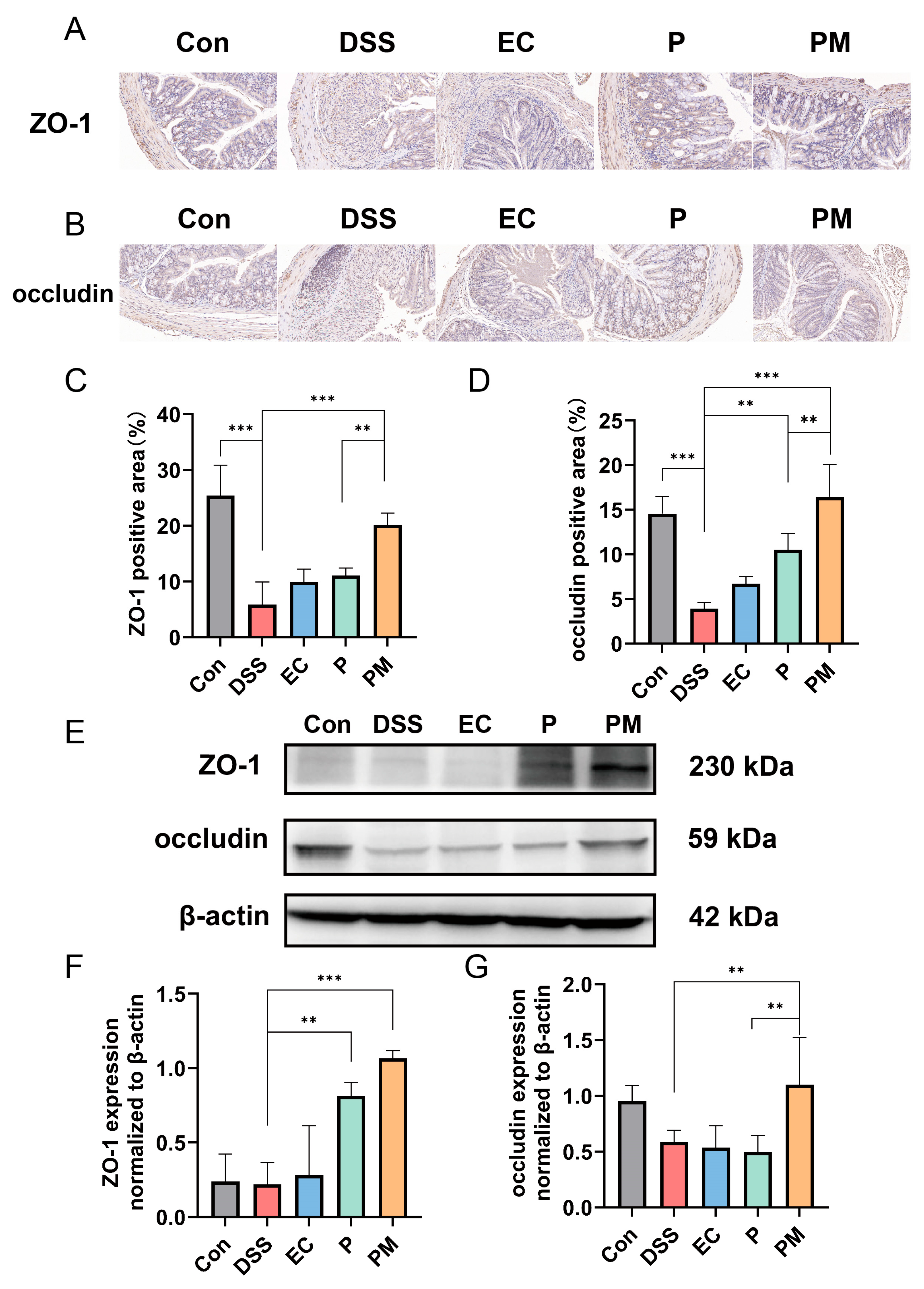
3.4. Probiotic Microcapsules Alleviated Intestinal Epithelial Barrier Injury by Upregulating TJ-Related Proteins
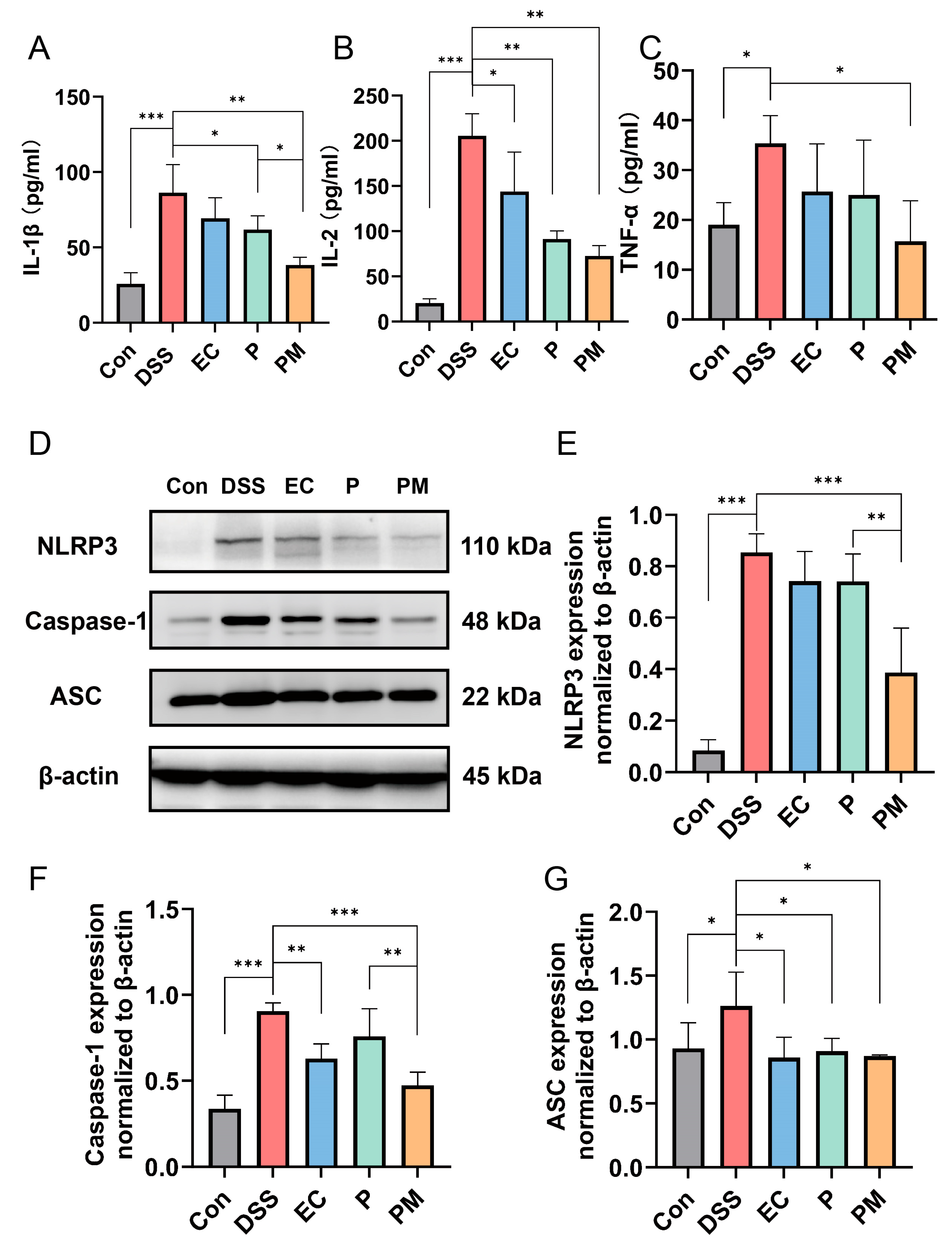
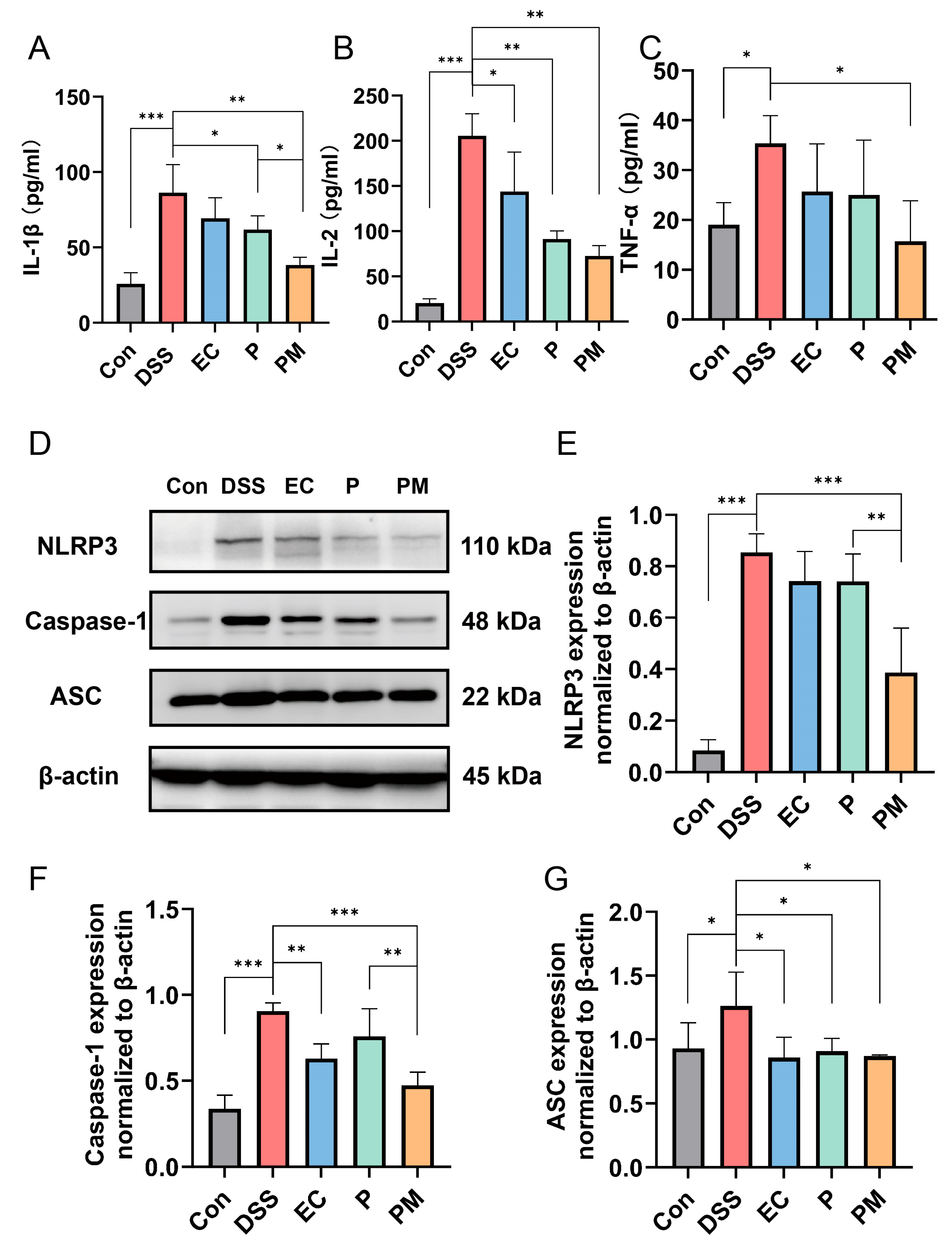
3.5. Probiotic Microcapsules Alleviated Inflammation by Regulating the Production of Inflammatory Cytokines and Suppressing the NLRP3 Inflammasome Signaling Pathway
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhao, B.; Wu, J.; Li, J.; Bai, Y.; Luo, Y.; Ji, B.; Xia, B.; Liu, Z.; Tan, X.; Lv, J.; et al. Lycopene Alleviates DSS-Induced Colitis and Behavioral Disorders via Mediating Microbes-Gut-Brain Axis Balance. J. Agric. Food Chem. 2020, 68, 3963–3975. [Google Scholar] [CrossRef] [PubMed]
- Gros, B.; Kaplan, G.G. Ulcerative Colitis in Adults: A Review. JAMA 2023, 330, 951–965. [Google Scholar] [CrossRef] [PubMed]
- Beamish, L.A.; Osornio-Vargas, A.R.; Wine, E. Air pollution: An environmental factor contributing to intestinal disease. J. Crohns. Colitis 2011, 5, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Samadder, N.J.; Valentine, J.F.; Guthery, S.; Singh, H.; Bernstein, C.N.; Leighton, J.A.; Wan, Y.; Wong, J.; Boucher, K.; Pappas, L.; et al. Family History Associates with Increased Risk of Colorectal Cancer in Patients with Inflammatory Bowel Diseases. Clin. Gastroenterol. Hepatol. 2019, 17, 1807–1813.e1. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.Y.; Liu, X.J.; Hao, J.Y. Gut microbiota in ulcerative colitis: Insights on pathogenesis and treatment. J. Dig. Dis. 2020, 21, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Kaminsky, L.W.; Al-Sadi, R.; Ma, T.Y. IL-1β and the Intestinal Epithelial Tight Junction Barrier. Front. Immunol. 2021, 12, 767456. [Google Scholar] [CrossRef] [PubMed]
- Elkouby-Naor, L.; Ben-Yosef, T. Functions of claudin tight junction proteins and their complex interactions in various physiological systems. Int. Rev. Cell Mol. Biol. 2010, 279, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Mitic, L.L.; Van Itallie, C.M.; Anderson, J.M. Molecular physiology and pathophysiology of tight junctions I. Tight junction structure and function: Lessons from mutant animals and proteins. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 279, G250–G254. [Google Scholar] [CrossRef] [PubMed]
- Odenwald, M.A.; Choi, W.; Kuo, W.T.; Singh, G.; Sailer, A.; Wang, Y.; Shen, L.; Fanning, A.S.; Turner, J.R. The scaffolding protein ZO-1 coordinates actomyosin and epithelial apical specializations in vitro and in vivo. J. Biol. Chem. 2018, 293, 17317–17335. [Google Scholar] [CrossRef]
- Singh, R.; Chandrashekharappa, S.; Bodduluri, S.R.; Baby, B.V.; Hegde, B.; Kotla, N.G.; Hiwale, A.A.; Saiyed, T.; Patel, P.; Vijay-Kumar, M.; et al. Enhancement of the gut barrier integrity by a microbial metabolite through the Nrf2 pathway. Nat. Commun. 2019, 10, 89. [Google Scholar] [CrossRef]
- Dong, Y.; Fan, H.; Zhang, Z.; Jiang, F.; Li, M.; Zhou, H.; Guo, W.; Zhang, Z.; Kang, Z.; Gui, Y.; et al. Berberine ameliorates DSS-induced intestinal mucosal barrier dysfunction through microbiota-dependence and Wnt/β-catenin pathway. Int. J. Biol. Sci. 2022, 18, 1381–1397. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Bai, R.; Zhou, W.; Yao, Z.; Liu, Y.; Tang, S.; Ge, X.; Luo, L.; Luo, C.; Hu, G.F.; et al. Angiogenin maintains gut microbe homeostasis by balancing α-Proteobacteria and Lachnospiraceae. Gut 2021, 70, 666–676. [Google Scholar] [CrossRef] [PubMed]
- Federici, S.; Kredo-Russo, S.; Valdés-Mas, R.; Kviatcovsky, D.; Weinstock, E.; Matiuhin, Y.; Silberberg, Y.; Atarashi, K.; Furuichi, M.; Oka, A.; et al. Targeted suppression of human IBD-associated gut microbiota commensals by phage consortia for treatment of intestinal inflammation. Cell 2022, 185, 2879–2898.e24. [Google Scholar] [CrossRef] [PubMed]
- Nakase, H.; Sato, N.; Mizuno, N.; Ikawa, Y. The influence of cytokines on the complex pathology of ulcerative colitis. Autoimmun. Rev. 2022, 21, 103017. [Google Scholar] [CrossRef] [PubMed]
- Sheng, K.; Xu, Y.; Kong, X.; Wang, J.; Zha, X.; Wang, Y. Probiotic Bacillus cereus Alleviates Dextran Sulfate Sodium-Induced Colitis in Mice through Improvement of the Intestinal Barrier Function, Anti-Inflammation, and Gut Microbiota Modulation. J. Agric. Food Chem. 2021, 69, 14810–14823. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Xu, W.; Zhou, R. NLRP3 inflammasome activation and cell death. Cell Mol. Immunol. 2021, 18, 2114–2127. [Google Scholar] [CrossRef] [PubMed]
- Damião, A.; de Azevedo, M.F.C.; Carlos, A.S.; Wada, M.Y.; Silva, T.V.M.; Feitosa, F.C. Conventional therapy for moderate to severe inflammatory bowel disease: A systematic literature review. World J. Gastroenterol. 2019, 25, 1142–1157. [Google Scholar] [CrossRef] [PubMed]
- Durando, M.; Tiu, H.; Kim, J.S. Sulfasalazine-Induced Crystalluria Causing Severe Acute Kidney Injury. Am. J. Kidney Dis. 2017, 70, 869–873. [Google Scholar] [CrossRef] [PubMed]
- Shin, T.; Okada, H. Infertility in men with inflammatory bowel disease. World J. Gastrointest. Pharmacol. Ther. 2016, 7, 361–369. [Google Scholar] [CrossRef]
- Harmand, P.O.; Solassol, J. Thiopurine Drugs in the Treatment of Ulcerative Colitis: Identification of a Novel Deleterious Mutation in TPMT. Genes 2020, 11, 1212. [Google Scholar] [CrossRef]
- Bao, Y.; Zhang, Y.; Li, H.; Liu, Y.; Wang, S.; Dong, X.; Su, F.; Yao, G.; Sun, T.; Zhang, H. In vitro screen of Lactobacillus plantarum as probiotic bacteria and their fermented characteristics in soymilk. Ann. Microbiol. 2012, 62, 1311–1320. [Google Scholar] [CrossRef]
- Wang, Z.; Bao, Y.; Zhang, Y.; Zhang, J.; Yao, G.; Wang, S.; Zhang, H. Effect of Soymilk Fermented with Lactobacillus plantarum P-8 on Lipid Metabolism and Fecal Microbiota in Experimental Hyperlipidemic Rats. Food Biophys. 2013, 8, 43–49. [Google Scholar] [CrossRef]
- Bao, Y.; Wang, Z.; Zhang, Y.; Zhang, J.; Wang, L.; Dong, X.; Su, F.; Yao, G.; Wang, S.; Zhang, H. Effect of Lactobacillus plantarum P-8 on lipid metabolism in hyperlipidemic rat model. Eur. J. Lipid. Sci. Technol. 2012, 114, 1230–1236. [Google Scholar] [CrossRef]
- Frakolaki, G.; Katsouli, M.; Giannou, V.; Tzia, C. Novel encapsulation approach for Bifidobacterium subsp. lactis (BB-12) viability enhancement through its incorporation into a double emulsion prior to the extrusion process. LWT-Food Sci. Technol. 2020, 130, 109671. [Google Scholar] [CrossRef]
- Zhang, M.; Yin, C.; Qian, J. Microencapsulation of Lactobacillus plantarum LN66 and its survival potential under different packaging conditions. J. Microencapsul. Microcapsules Liposomes Nanoparticles Microcells Microspheres 2022, 39, 601–608. [Google Scholar]
- Rather, S.A.; Akhter, R.; Masoodi, F.A.; Gani, A.; Wani, S.M. Effect of double alginate microencapsulation on in vitro digestibility and thermal tolerance of Lactobacillus plantarum NCDC201 and L. casei NCDC297. LWT Food Sci. Technol. 2017, 83, 50–58. [Google Scholar] [CrossRef]
- Rajam, R.; Kumar, S.B.; Prabhasankar, P.; Anandharamakrishnan, C. Microencapsulation of Lactobacillus plantarum MTCC 5422 in fructooligosaccharide and whey protein wall systems and its impact on noodle quality. J. Food Sci. Technol. 2015, 52, 4029–4041. [Google Scholar] [CrossRef]
- Liu, Y.; Sheng, Y.; Pan, Q.; Xue, Y.; Yu, L.; Tian, F.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. Identification of the key physiological characteristics of Lactobacillus plantarum strains for ulcerative colitis alleviation. Food Funct. 2020, 11, 1279–1291. [Google Scholar] [CrossRef]
- Dan, T.; Chen, H.; Li, T.; Tian, J.; Ren, W.; Zhang, H.; Sun, T. Influence of Lactobacillus plantarum P-8 on Fermented Milk Flavor and Storage Stability. Front. Microbiol. 2019, 9, 423475. [Google Scholar] [CrossRef]
- Wang, Y.; Lv, X.; Li, X.; Zhao, J.; Liu, H. Protective Effect of Lactobacillus plantarum P8 on Growth Performance, Intestinal Health, and Microbiota in Eimeria-Infected Broilers. Front. Microbiol. 2021, 12, 705758. [Google Scholar] [CrossRef]
- Lew, L.C.; Hor, Y.Y.; Yusoff, N.A.A.; Choi, S.B.; Yusoff, M.S.; Roslan, N.S.; Ahmad, A.; Mohammad, J.A.; Abdullah, M.F.I.; Zakaria, N.; et al. Probiotic Lactobacillus plantarum P8 alleviated stress and anxiety while enhancing memory and cognition in stressed adults: A randomised, double-blind, placebo-controlled study. Clin. Nutr. (Edinb. Scotl.) 2018, 38, 2053–2064. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Wang, B.; Wang, W.; Liu, G.; Ge, W.; Zhang, M.; Yue, B.; Kong, M. Microencapsulation of Lactobacillus casei BNCC 134415 under lyophilization enhances cell viability during cold storage and pasteurization, and in simulated gastrointestinal fluids. LWT 2019, 116, 108521. [Google Scholar] [CrossRef]
- Gu, M.; Zhang, Z.; Pan, C.; Goulette, T.R.; Zhang, R.; Hendricks, G.; Mcclements, D.J.; Xiao, H. Encapsulation of Bifidobacterium pseudocatenulatum G7 in gastroprotective microgels: Improvement of the bacterial viability under simulated gastrointestinal conditions. Food Hydrocoll. 2019, 91, 283–289. [Google Scholar] [CrossRef]
- Zhao, M.; Wang, Y.; Huang, X.; Gaenzle, M.; Wu, Z.; Nishinari, K.; Yang, N.; Fang, Y. Ambient storage of microencapsulated Lactobacillus plantarum ST-III by complex coacervation of type-A gelatin and gum arabic. Food Funct. 2018, 9, 1000–1008. [Google Scholar] [CrossRef] [PubMed]
- Chassaing, B.; Aitken, J.D.; Malleshappa, M.; Vijay-Kumar, M. Dextran sulfate sodium (DSS)-induced colitis in mice. Curr. Protoc. Immunol. 2014, 104, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Yao, D.; Dai, W.; Dong, M.; Dai, C.; Wu, S. MUC2 and related bacterial factors: Therapeutic targets for ulcerative colitis. EBioMedicine 2021, 74, 103751. [Google Scholar] [CrossRef]
- Liu, Y.; Yu, X.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. The role of MUC2 mucin in intestinal homeostasis and the impact of dietary components on MUC2 expression. Int. J. Biol. Macromol. 2020, 164, 884–891. [Google Scholar] [CrossRef]
- Gonzalez, A.; Krieg, R.; Massey, H.D.; Carl, D.; Ghosh, S.; Gehr, T.W.B.; Ghosh, S.S. Sodium butyrate ameliorates insulin resistance and renal failure in CKD rats by modulating intestinal permeability and mucin expression. Nephrol. Dial. Transplant. 2019, 34, 783–794. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, T.; Guo, C.; Geng, M.; Gai, S.; Qi, W.; Li, Z.; Song, Y.; Luo, X.; Zhang, T.; et al. Bacillus subtilis RZ001 improves intestinal integrity and alleviates colitis by inhibiting the Notch signalling pathway and activating ATOH-1. Pathog. Dis. 2020, 78, ftaa016. [Google Scholar] [CrossRef]
- Casini-Raggi, V.; Kam, L.; Chong, Y.J.; Fiocchi, C.; Pizarro, T.T.; Cominelli, F. Mucosal imbalance of IL-1 and IL-1 receptor antagonist in inflammatory bowel disease. A novel mechanism of chronic intestinal inflammation. J. Immunol. 1995, 154, 2434–2440. [Google Scholar] [CrossRef]
- Nanthakumar, N.N.; Fusunyan, R.D.; Sanderson, I.; Walker, W.A. Inflammation in the developing human intestine: A possible pathophysiologic contribution to necrotizing enterocolitis. Proc. Natl. Acad. Sci. USA 2000, 97, 6043–6048. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Ma, P.; Zhang, Y.; Mi, Y.; Fan, D. Ginsenoside Rk3 alleviated DSS-induced ulcerative colitis by protecting colon barrier and inhibiting NLRP3 inflammasome pathway. Int. Immunopharmacol. 2020, 85, 106645. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Núñez, G. The NLRP3 inflammasome: Activation and regulation. Trends Biochem. Sci. 2023, 48, 331–344. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Wu, H. Structural Mechanisms of NLRP3 Inflammasome Assembly and Activation. Annu. Rev. Immunol. 2023, 41, 301–316. [Google Scholar] [CrossRef] [PubMed]
- Ahn, H.; Kang, S.G.; Yoon, S.I.; Kim, P.H.; Kim, D.; Lee, G.S. Poly-gamma-glutamic acid from Bacillus subtilis upregulates pro-inflammatory cytokines while inhibiting NLRP3, NLRC4 and AIM2 inflammasome activation. Cell Mol. Immunol. 2018, 15, 111–119. [Google Scholar] [CrossRef]
- Wei, Y.Y.; Fan, Y.M.; Ga, Y.; Zhang, Y.N.; Han, J.C.; Hao, Z.H. Shaoyao decoction attenuates DSS-induced ulcerative colitis, macrophage and NLRP3 inflammasome activation through the MKP1/NF-κB pathway. Phytomedicine 2021, 92, 153743. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Score | Weight Loss | Stool Consistency | Fecal Occult Blood |
|---|---|---|---|
| 0 | 0 | Normal | Normal |
| 1 | 1–5% | - | - |
| 2 | 6–10% | Loose stools | Hemoccult positive |
| 3 | 10–15% | - | - |
| 4 | >15% | Diarrhea | Gross bleeding |
| Score | Inflammation | Mucosal Damage | Crypt Damage |
|---|---|---|---|
| 0 | None | None | None |
| 1 | Mild | Mucous layer | 1/3 |
| 2 | Moderate | Submucosa | 2/3 |
| 3 | Severe | Muscularis and serosa | 100% |
| 4 | - | - | 100% + epithelium loss |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Sun, Y.; Ma, X.; Yang, T.; Wang, F. The Lactobacillus plantarum P-8 Probiotic Microcapsule Prevents DSS-Induced Colitis through Improving Intestinal Integrity and Reducing Colonic Inflammation in Mice. Nutrients 2024, 16, 1055. https://doi.org/10.3390/nu16071055
Wang H, Sun Y, Ma X, Yang T, Wang F. The Lactobacillus plantarum P-8 Probiotic Microcapsule Prevents DSS-Induced Colitis through Improving Intestinal Integrity and Reducing Colonic Inflammation in Mice. Nutrients. 2024; 16(7):1055. https://doi.org/10.3390/nu16071055
Chicago/Turabian StyleWang, Han, Yaxuan Sun, Xuan Ma, Tianyu Yang, and Feng Wang. 2024. "The Lactobacillus plantarum P-8 Probiotic Microcapsule Prevents DSS-Induced Colitis through Improving Intestinal Integrity and Reducing Colonic Inflammation in Mice" Nutrients 16, no. 7: 1055. https://doi.org/10.3390/nu16071055
APA StyleWang, H., Sun, Y., Ma, X., Yang, T., & Wang, F. (2024). The Lactobacillus plantarum P-8 Probiotic Microcapsule Prevents DSS-Induced Colitis through Improving Intestinal Integrity and Reducing Colonic Inflammation in Mice. Nutrients, 16(7), 1055. https://doi.org/10.3390/nu16071055