

Ingestion of Bean Leaves Reduces Metabolic Complications and Restores Intestinal Integrity in C57BL/6 Mice with Obesity Induced by a High-Fat and High-Fructose Diet
 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Bean Leaves
2.2. Bromathological and Phytochemical Characterization of the Common Bean Leaves
2.2.1. Bromatological Characterization of the Common Bean Leaves
2.2.2. Extraction of Phenolic Compounds
2.2.3. Determination of Total Phenols
2.2.4. Determination of Total Flavonoids
2.2.5. Quantification of Condensed Tannins
2.2.6. Quantification of Total Saponins
2.2.7. Identification and Quantification of Phenolic and Flavonoids in Bean Leaves by UPLC-ESI-MS/MS
2.3. Experimental Design
2.3.1. Experimental Animals
2.3.2. Design of Diets for Experimental Animals
2.3.3. Energy and Nutritional Content of Diets
2.3.4. Experimental Design
2.4. Changes in Body Composition from the Consumption of Bean Leaves and the Change to a Healthy Diet
2.4.1. Body Composition
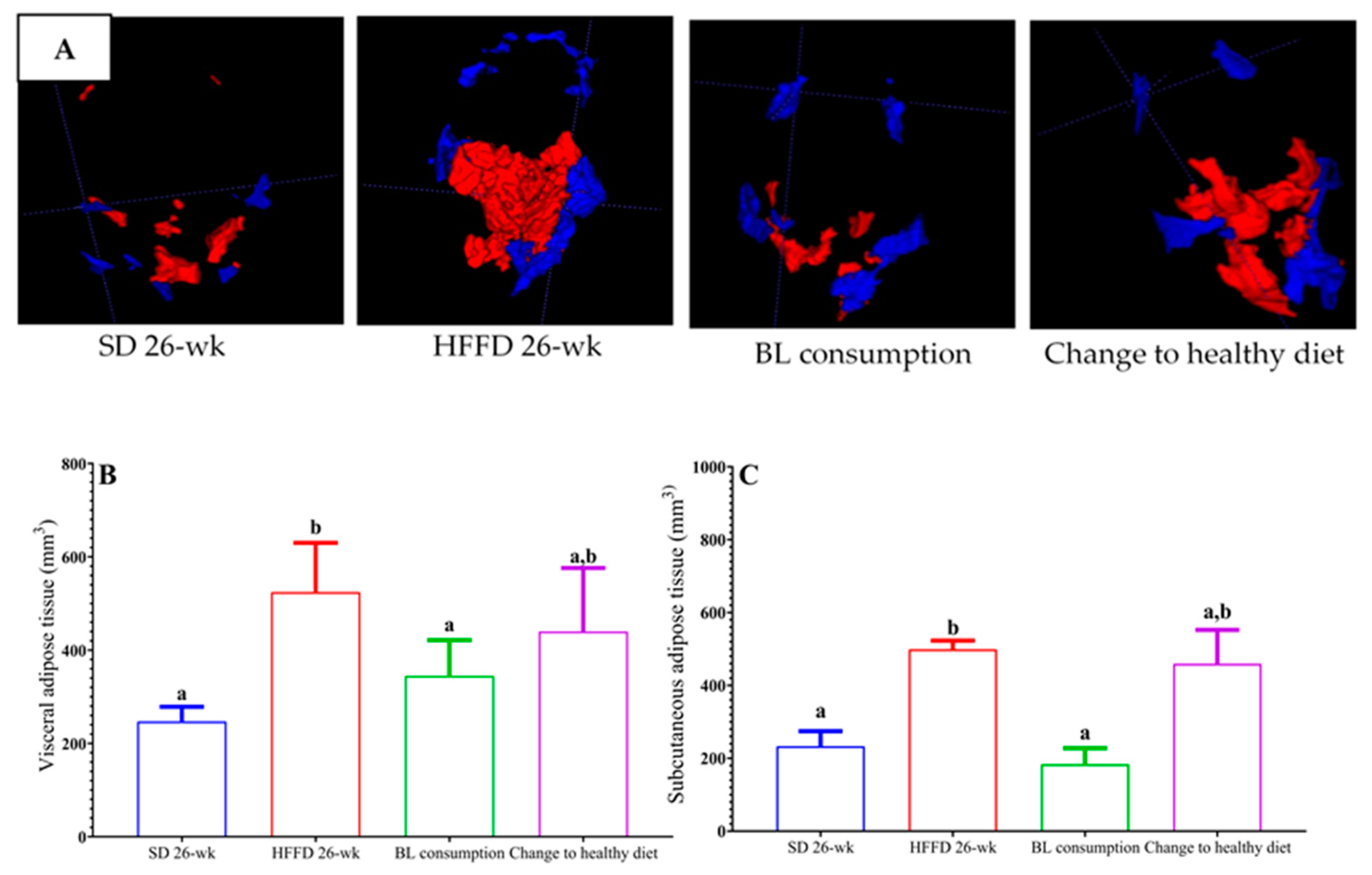
2.4.2. Quantification of Adipose Tissue
2.5. Role of Bean Leaves Consumption and the Change to a Healthy Diet in Lipid Metabolism
2.5.1. Determination of the Content of Total Cholesterol and Triglycerides in Serum
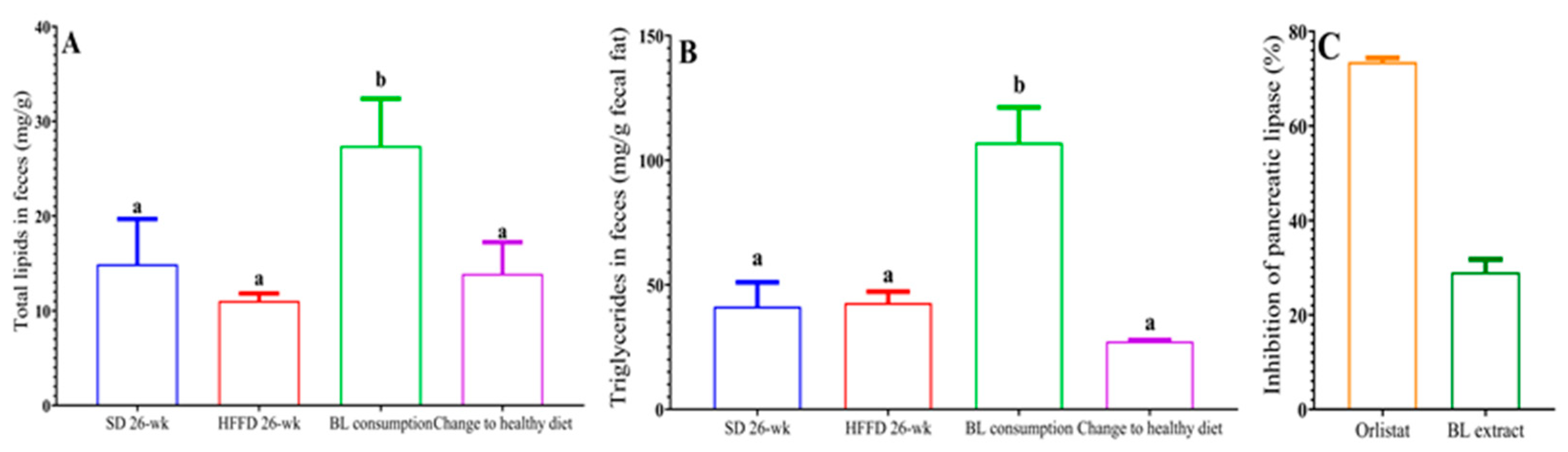
2.5.2. Total Lipids and Triglycerides in Feces
2.5.3. Pancreatic Lipase Activity
2.6. Euthanasia
2.7. Impact of Bean Leaves Consumption and Healthy Diet Change on the Intestinal Integrity and Metabolic Endotoxemia
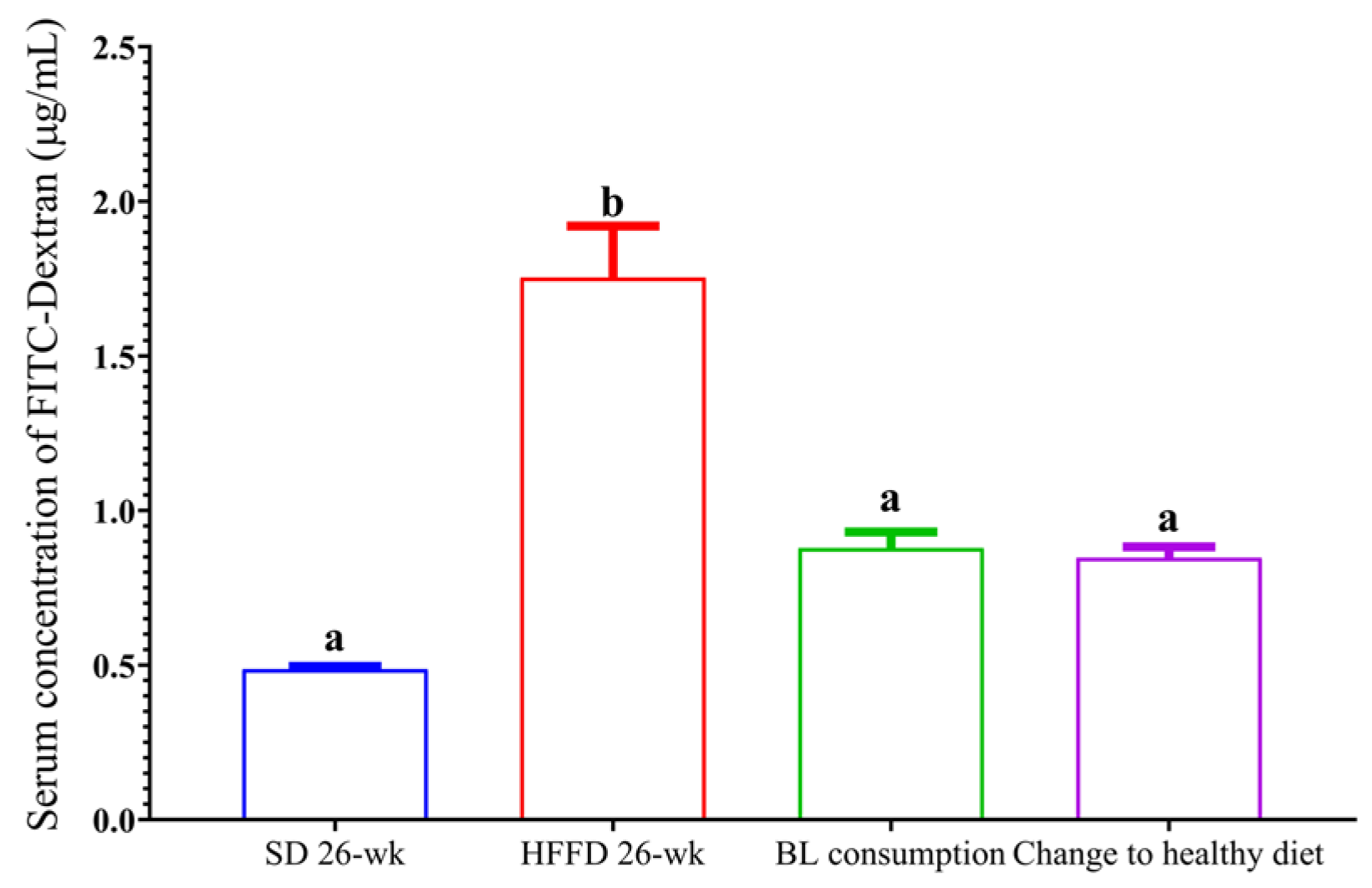
2.7.1. Intestinal Permeability by the FITC–Dextran Assay
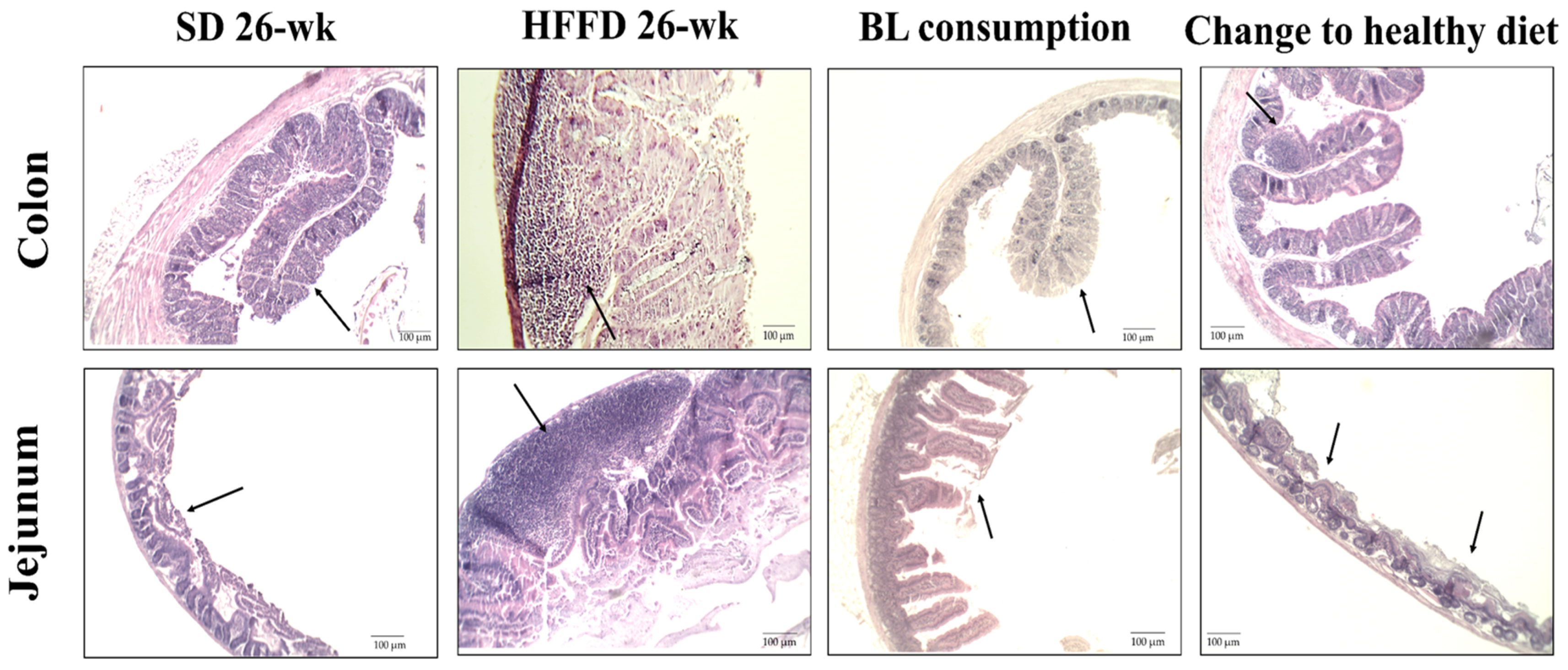
2.7.2. Histopathological and Morphometric Analysis of the Jejunum and Proximal Colon
2.7.3. Determination of LPS and IL-6 Levels in Serum
2.8. Statistical Analysis
3. Results
3.1. Quantification of Dietary Fiber and Phytochemical Compounds
3.2. Effect of Bean Leaves on Energy Intake
3.3. Treatment with Bean Leaves and Its Effect on Body Composition
3.3.1. Body Weight and the Increase in the Abdominal Circumference
3.3.2. Determination of Volume Adipose Tissue
3.4. Impact of the Consumption of Different Diets on the Lipid Profile
3.4.1. Serum Cholesterol and Triglycerides Concentration
3.4.2. Lipid Excretion
3.4.3. Inhibition of Pancreatic Lipase In Vitro
3.5. Changes in Morphometry and Histopathology in the Intestine and colon
3.5.1. Relative Weight and Length of the Small Intestine and Colon
3.5.2. Histological Change in Intestine and Colon
3.6. Intestinal Permeability and Metabolic Endotoxemia
3.6.1. Intestinal Permeability
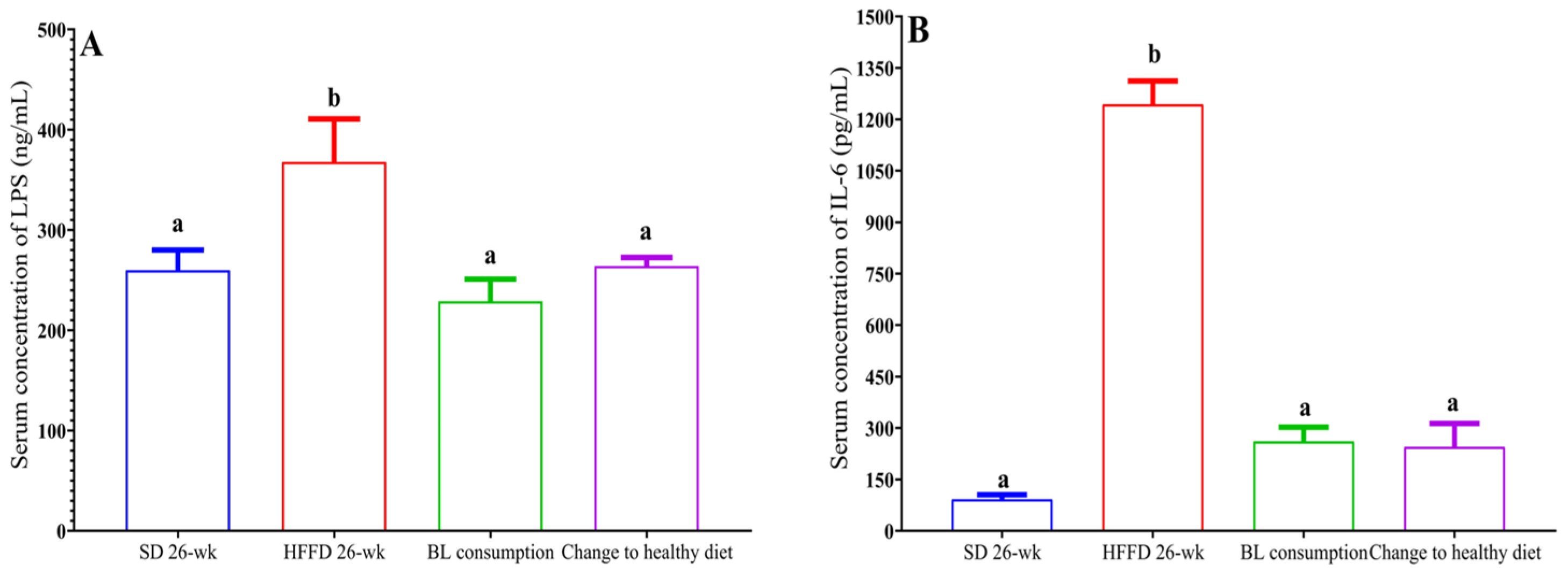
3.6.2. Metabolic Endotoxemia and Inflammation
3.6.3. Intake of Dietary Fiber and Total Phenols
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, S.J.; Kim, S.-E.; Kim, A.-R.; Kang, S.; Park, M.-Y.; Sung, M.-K. Dietary fat intake and age modulate the composition of the gut microbiota and colonic inflammation in C57BL/6J mice. BMC Microbiol. 2019, 19, 193. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Campos Mayoral, L.; Mayoral Andrade, G.; Pérez-Campos Mayoral, E.; Hernandez Huerta, T.; Pia Canseco, S.; Rodal Canales, F.J.; Cabrera-Fuentes, H.A.; Martinez Cruz, M.; Pérez Santiago, A.D.; Alpuche, J.J.; et al. Obesity subtypes, related biomarkers & heterogeneity. Indian. J. Med. Res. 2020, 151, 11–21. [Google Scholar] [CrossRef]
- Agustí, A.; García-Pardo, M.P.; López-Almela, I.; Campillo, I.; Maes, M.; Romani-Perez, M.; Sanz, Y. Interplay between the gut-brain axis, obesity and cognitive function. Front. Neurosci. 2018, 12, 155. [Google Scholar] [CrossRef]
- Horne, R.G.; Yu, Y.; Zhang, R.; Abdalqadir, N.; Rossi, L.; Surette, M.; Sherman, P.M.; Adeli, K. High fat-high fructose diet-induced changes in the gut microbiota associated with dyslipidemia in Syrian hamsters. Nutrients 2020, 12, 3557. [Google Scholar] [CrossRef] [PubMed]
- Soares, A.; Beraldi, E.J.; Ferreira, P.E.; Bazotte, R.B.; Buttow, N.C. Intestinal and neuronal myenteric adaptations in the small intestine induced by a high-fat diet in mice. BMC Gastroenterol. 2015, 15, 3. [Google Scholar] [CrossRef] [PubMed]
- Dailey, M.J. Nutrient-induced intestinal adaption and its effect in obesity. Physiol. Behav. 2014, 136, 74–78. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, J.; Fernández Real, J.M.; Guarner, F.; Guelmonde, M.; Rodriguez, J.M.; Saens de Pipaon, M.; Sanz, Y. Gut microbes and health. Gastroenterol. Hepatol. 2021, 44, 519–535. [Google Scholar] [CrossRef]
- Organización Mundial de la Salud (OMS). Dieta Saludable. Página Web. Available online: https://www.who.int/es/news-room/fact-sheets/detail/healthy-diet (accessed on 12 October 2022).
- SAGARPA. Planeación Agrícola Nacional 2017–2030. Frijol Mexicano. Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación. 2017. México. Available online: https://docplayer.es/83182578-Planeacion-agricola-nacional-frijol.html (accessed on 4 June 2022).
- Mateos-Maces, L.; Chávez-Servia, J.L.; Vera-Guzmán, A.M.; Aquino-Bolaños, E.N.; Alba-Jiménez, J.E.; Villagómez-González, B.B. Edible leafy plants from Mexico as sources of antioxidant compounds, and their nutritional, nutraceutical and antimicrobial potential: A review. Antioxidants 2020, 9, 541. [Google Scholar] [CrossRef]
- Becerril-Campos, A.A.; Ocampo-Anguiano, P.V.; Mondragón-Jacobo, C.; Escobar-García, K.; Camacho-Barrón, M.; Anaya-Loyola, M.A.; Feregrino-Pérez, A.A.; García-Gasca, G.; Ahumada-Solórzano, S.M. Phaseolus vulgaris L. leaves increase short-chain fatty acid (SCFA) production, ameliorating early metabolic alterations. Plant Foods Hum. Nutr. 2022, 77, 421–426. [Google Scholar] [CrossRef]
- Martinez-Zavala, M.; Mora-Avilés, M.A.; Anaya-Loyola, M.A.; Guzmán-Maldonado, O.; Aguilera-Barreyro, A.; Blanco-Labra, A.; García-Gasca, T. Common bean leaves as a source of dietary iron. Plant Foods Hum. Nutr. 2016, 71, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Becerril-Campos, A.A.; Ramos-Gómez, M.; de los Ríos-Arellano, E.A.; Ocampo-Anguiano, P.V.; Gónzalez-Gallardo, A.; Macotela, Y.; García-Gasca, T.; Ahumada-Solórzano, S.M. Bean leaves ameliorate lipotoxicity in fatty liver disease. Nutrients 2023, 15, 2928. [Google Scholar] [CrossRef] [PubMed]
- Heredia-Ortega, J.; Aguilera, T.; Resis, M.; Bernal, K. Fibra dietética total: Métodos de determinación en pastas de oleaginosas. In Facultad de Ciencias Naturales; Universidad Autonoma de Queretaro: Queretaro, México, 2020; pp. 91–102. [Google Scholar]
- Herrera-Rocha, K.M.; Rocha-Guzmán, N.E.; Gallegos-Infante, J.A.; González-Laredo, R.F.; Larrosa-Pérez, M.; Moreno-Jiménez, M.R. Phenolic acids and flavonoids in acetonic extract from quince (Cydonia oblonga Mill.): Nutraceuticals with antioxidant and anti-inflammatory potential. Molecules 2022, 27, 2462. [Google Scholar] [CrossRef]
- Baek, S.; Choi, N.H.; Lee, K.P.; Jhun, H.; Kim, J. Smallanthus sonchifolius leaf attenuates neuroinflammation. J. Exerc. Nutr. Biochem. 2018, 22, 31–35. [Google Scholar] [CrossRef]
- Singleton, V.; Rosi, J. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 58–144. [Google Scholar] [CrossRef]
- Campos-Vega, R.; Reynoso-Camacho, R.; Pedraza-Aboytes, G.; Acosta Gallegos, J.A.; Guzmán-Maldonado, S.H.; Paredes López, O.; Oomah, B.D.; Loarca-Piña, G. Chemical composition and in vitro polysaccharide fermentation of different beans (Phaseolus vulgaris L.). J. Food Sci. 2009, 74, 59–65. [Google Scholar] [CrossRef]
- Feregrino-Pérez, A.A.; Berumen, L.C.; García-Alcocer, G.; Guevara-Gonzalez, R.G.; Ramos-Gomez, M.; Reynoso-Camacho, R.; Acosta-Gallegos, J.A.; Loarca-Piña, G. Composition and chemopreventive effect of polysaccharides from common beans (Phaseolus vulgaris L.) on azoxymethane-induced colon cancer. J. Agric. Food Chem. 2008, 56, 8737–8744. [Google Scholar] [CrossRef] [PubMed]
- Dini, I.; Tenore, G.C.; Dini, A. Saponins in ipomoea batatas tubers: Isolation, characterization, quantification and antioxidant properties. Food Chem. 2009, 113, 411–419. [Google Scholar] [CrossRef]
- García-García, J.A.; Reding-Bernal, A.; López-Alvarenga, J.C. Cálculo del tamaño de la muestra en investigación en educación médica. Investig. En Educ. Médica 2013, 2, 217–224. [Google Scholar] [CrossRef]
- Martínez-Orozco, H.; Reyes-Castro, L.A.; Lomas-Soria, C.; Sandoval-Salazar, C.; Ramírez-Emiliano, J.; Díaz-Cintra, S.; Solís-Ortiz, S. High-fat and combined high-fat–high-fructose diets impair episodic-like memory and decrease glutamate and glutamine in the hippocampus of adult mice. Nutr. Neurosci. 2022, 25, 2479–2489. [Google Scholar] [CrossRef]
- Kraus, D.; Yang, Q.; Kahn, B. Lipid Extraction from Mouse Feces. BIO-PROTOCOL 2015, 5, e1375. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Ramírez, I.F.; González-Dávalos, M.L.; Mora, O.; Gallegos-Corona, M.A.; Reynoso-Camacho, R. Effect of Ocimum sanctum and Crataegus pubescens aqueous extracts on obesity, inflammation, and glucose metabolism. J. Funct. Foods 2017, 35, 24–31. [Google Scholar] [CrossRef]
- Shi, H.; Wang, Q.; Zheng, M.; Hao, S.; Lum, J.S.; Chen, X.; Huang, X.F.; Yu, Y.; Zheng, K. Supplement of microbiota-accessible carbohydrates prevents neuroinflammation and cognitive decline by improving the gut microbiota-brain axis in diet-induced obese mice. J. Neuroinflamm. 2020, 17, 77. [Google Scholar] [CrossRef] [PubMed]
- van de Wouw, M.; Boehme, M.; Lyte, J.M.; Wiley, N.; Strain, C.; O’Sullivan, O.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Short-chain fatty acids: Microbial metabolites that alleviate stress-induced brain–gut axis alterations. J. Physiol. 2018, 596, 4923–4944. [Google Scholar] [CrossRef] [PubMed]
- Quesenberry, K.E.; Carpenter, J.W. Rats and mice. In Ferrets, Rabbits and Todents: Clinical Medicine and Surgery, 4th ed.; Elsevier: Amsterdam, The Netherlands, 2012; pp. 345–365. [Google Scholar]
- Zhao, Q.; Hou, D.; Fu, Y.; Xue, Y.; Guan, X.; Shen, Q. Adzuki bean alleviates obesity and insulin resistance induced by a high-fat diet and modulates gut microbiota in mice. Nutrients 2021, 13, 3240. [Google Scholar] [CrossRef]
- Martinez, K.B.; Mackert, J.D.; McIntosh, M.K. Chapter 18: Polyphenols and Intestinal Health; Watson, R.R., Ed.; Academic Press: Cambridge, MA, USA, 2017; pp. 191–210. [Google Scholar] [CrossRef]
- Yoon, D.S.; Cho, S.Y.; Yoon, H.J.; Kim, S.R.; Jung, U.J. Protective effects of p-coumaric acid against high-fat diet-induced metabolic dysregulation in mice. Biomed. Pharmacother. 2021, 142, 111969. [Google Scholar] [CrossRef] [PubMed]
- Jihyeon, Y.; Chu-Sook, K.; Thai Hien, T.; Min-Seon, K.; Tsuyoshi, G.; Teruo, K.; Myug-Sook, C.; Taesun, P.; Mi-Kyung, S.; Jong Won, Y.; et al. Quercetin protects obesity-induced hypothalamic inflammation by reducing microglia-mediated inflammatory responses via HO-1 induction. Nutrients 2017, 9, 650. [Google Scholar] [CrossRef]
- Wang, T.; Wu, Q.; Zhao, T. Preventive effects of kaempferol on high-fat diet-induced obesity complications in C57BL/6 Mice. BioMed Res. Int. 2020, 2020, 4532482. [Google Scholar] [CrossRef] [PubMed]
- Bian, Y.; Lei, J.; Zhong, J.; Wang, B.; Wan, Y.; Li, J.; Liao, C.; He, W.; Liu, Z.; Ito, K.; et al. Kaempferol reduces obesity, prevents intestinal inflammation, and modulates gut microbiota in high-fat diet mice. J. Nutr. Biochem. 2022, 99, 108840. [Google Scholar] [CrossRef] [PubMed]
- Santiago-Saenz, Y.O.; Hernández-Fuentes, A.D.; López-Palestina, C.U.; Garrido-Cauich, J.H.; Alatorre-Cruz, L.M.; Monroy-Torres, R. Nutritional importance and biological activity of compounds from quelites consumed in Mexico. Rev. Chil. Nutr. 2019, 46, 593–605. [Google Scholar] [CrossRef]
- Sallam, I.E.; Abdelwareth, A.; Attia, H.; Aziz, R.K.; Homsi, M.N.; von Bergen, M.; Farag, M.A. Effect of gut microbiota biotransformation on dietary tannins and human health implications. Microorganisms 2021, 9, 965. [Google Scholar] [CrossRef]
- Toney, A.M.; Albusharif, M.; Works, D.; Polenz, L.; Schlange, S.; Chaidez, V.; Ramer-Tait, A.E.; Chung, S. Differential effects of whole red raspberry polyphenols and their gut metabolite urolithin a on neuroinflammation in BV-2 microglia. Int. J. Environ. Res. Public Health 2021, 18, 68. [Google Scholar] [CrossRef]
- Do, M.H.; Lee, E.; Oh, M.J.; Kim, Y.; Park, H.Y. High-glucose or-fructose diet cause changes of the gut microbiota and metabolic disorders in mice without body weight change. Nutrients 2018, 10, 761. [Google Scholar] [CrossRef] [PubMed]
- Lutsiv, T.; Weir, T.L.; McGinley, J.N.; Neil, E.S.; Wei, Y.; Thompson, H.J. Compositional changes of the high-fat diet-induced gut microbiota upon consumption of common pulses. Nutrients 2021, 13, 3992. [Google Scholar] [CrossRef] [PubMed]
- Tan, R.; Dong, H.; Chen, Z.; Jin, M.; Yin, J.; Li, H.; Shi, D.; Shao, Y.; Wang, H.; Chen, T.; et al. Intestinal microbioa mediate high-fructose and high-fat diets to induce chronic intestinal inflammation. Front. Cell Infect. Microbiol. 2021, 11, 654074. [Google Scholar] [CrossRef] [PubMed]
- Shamah-Levy, T.; Vielma-Orozco, E.; Heredia-Hernández, O. Encuesta Nacional de Salud y Nutrición 2018-19 Resultados Nacionales. 2020. Available online: https://ensanut.insp.mx/encuestas/ensanut2018/doctos/informes/ensanut_2018_informe_final.pdf (accessed on 4 December 2022).
- Bala, C.; Craciun, A.E.; Hancu, N. Updating the concept of metabolically healthy obesity. Acta Endocrinol. 2016, 12, 197–205. [Google Scholar] [CrossRef]
- Zou, Y.; Sheng, G.; Yu, M.; Xie, G. The association between triglycerides and ectopic fat obesity: An inverted U-shaped curve. PLoS ONE 2020, 15, e0243068. [Google Scholar] [CrossRef]
- Obonyo, H.R.; Selvi, S.V. Evaluation of hydrated extract of Phaseolus vulgaris L. (bean plant) on hypoglycemia and hypolipidemic in streptozotocin-induced diabetic albino wistar rats. Int. J. Res. Pharm. Sci. 2019, 10, 3704–3710. [Google Scholar] [CrossRef]
- Thompson, H.J.; McGinley, J.N.; Neil, E.S.; Brick, M.A. Beneficial effects of common bean on adiposity and lipid metabolism. Nutrients 2017, 9, 998. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Liu, X.; Zhang, J.; Yan-Zhi, L.; Lu, L.; Mi, J.; You-Long, C.; Ya-Mei, Y.; Lin-Wu, R. Preventive effects of anthocyanins from lycium ruthenicum murray in high-fat diet-induced obese mice are related to the regulation of intestinal microbiota and inhibition of pancreatic lipase activity. Molecules 2022, 27, 2141. [Google Scholar] [CrossRef]
- Escobar-Ortiz, A.; Hernández-Saavedra, D.; Lizardi-Mendoza, J.; Pérez-Ramírez, I.F.; Mora-Izaguirre, O.; Ramos-Gómez, M.; Reynoso-Camacho, R. Consumption of cricket (Acheta domesticus) flour decreases insulin resistance and fat accumulation in rats fed with high-fat and -fructose diet. J. Food Biochem. 2022, 46, e14269. [Google Scholar] [CrossRef]
- Jang, C.; Hui, S.; Lu, W.; Cowan, A.J.; Morscher, R.J.; Lee, G.; Liu, W.; Tesz, G.J.; Birnbaun, M.J.; Rabinowitz, J.D. The small intestine converts dietary fructose into glucose and organic acids. Cell Metab. 2018, 27, 351–361. [Google Scholar] [CrossRef]
- Costes, L.M.M.; Lindenbergh-Kortleve, D.J.; van Berkel, L.A.; Veenbergen, S.; Raatgeep, H.C.; Simons-Oosterhuis, Y.; van Haaften, D.H.; Karrich, J.J.; Escher, J.C.; Groeneweg, M.; et al. IL-10 signaling prevents gluten-dependent intraepithelial CD4+ cytotoxic T lymphocyte infiltration and epithelial damage in the small intestine. Mucosal Immunol. 2019, 12, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Liu, Y.; Hu, J.; Gao, Y.; Ma, Y.; Wen, D. Chlorogenic acid-induced gut microbiota improves metabolic endotoxemia. Front Endocrinol. 2021, 12, 762691. [Google Scholar] [CrossRef] [PubMed]
- McGinley, J.N.; Fitzgerald, V.K.; Neil, E.S.; Omerigic, H.M.; Heuberger, A.L.; Weir, T.L.; McGee, R.; Vandemark, G.; Thompson, H.J. Pulse crop effects on gut microbial populations, intestinal function, and adiposity in a mouse model of diet-induced obesity. Nutrients 2020, 12, 593. [Google Scholar] [CrossRef] [PubMed]
- Di Tommaso, N.; Gasbarrini, A.; Ponziani, F.R. Intestinal barrier in human health and disease. Int. J. Environ. Res. Public Health 2021, 18, 12836. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Carbohydrates | Total Lipids | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Energy (kcal/g) | Proteins (%) | (%) | Of Which Fructose (%) | (%) | Of Which Lard (%) | Crude Fiber (g/100 g) | Total Dietary Fiber (g/100 g) | Total Phenolic Compounds (mg/g) | |
| SD | 3.40 | 28.54 | 58.15 | 0.00 | 13.43 | 0.00 | 5.10 | 36.92 | 2.53 |
| HFFD | 4.70 | 14.28 | 40.90 | 19.98 | 44.81 | 40.17 | 3.07 | 17.31 | 1.55 |
| BL consumption | 4.70 | 13.79 | 41.20 | 20.26 | 45.02 | 40.73 | 3.16 | 21.00 | 3.26 |
| Group | Diet |
|---|---|
| SD 26-week | Standard diet by 26 weeks |
| HFFD 26-week | HFFD by 26 weeks |
| BL consumption | HFFD + 10% BL by 12 weeks |
| Change healthy diet | Standard diet by 12 week |
| Compounds | µg/g |
|---|---|
| Phenolic acids | |
| Quinic acid | 97.44 ± 7.08 |
| Caffeic acid (derivative) * | 57.14 ± 3.95 |
| Cinnamic acid (derivative) * | 72.41 ± 13.00 |
| Protocatechoic acid | 8.31 ± 2.17 |
| Caftaric acid | 207.21 ± 22.20 |
| 4-hydroxybenzoic acid | 106.59 ± 2.18 |
| Coumaric acid (derivative) * | 1283.81 ± 110.87 |
| Ferulic acid (derivative) * | 95.53 ± 11.82 |
| 2-hydroxybenzoic acid | 96.76 ± 10.79 |
| Benzoic acid | 49.55 ± 0.54 |
| Flavonoids | |
| Rutin | 971.18 ± 73.19 |
| Quercetin 3-O-ß-glucuronide | 6892.20 ± 99.42 |
| Quercetin 3-O-glucoside | 73.05 ± 7.53 |
| Kaempferol 3-O-glucoside | 11.24 ± 0.10 |
| Naringin | 1.01 ± 0.36 |
| Naringenin | 2.29 ± 0.61 |
| Obesity Induction Phase | Experimental Treatment Phase | |||||
|---|---|---|---|---|---|---|
| SD 14-Week | HFFD 14-Week | SD 26-Week | HFFD 26-Week | BL Consumption | Change to Healthy Diet | |
| Food intake (g/day) | 4.59 ± 0.50 | 4.88 ± 0.30 | 3.79 ± 0.40 a | 5.13 ± 0.80 a | 4.54 ± 0.60 a | 5.09 ± 0.50 a |
| Energy intake (Kcal/day) | 15.75 ± 1.50 | 22.94 ± 1.30 * | 12.89 ± 1.50 a | 23.48 ± 3.80 b | 20.80 ± 2.70 b | 17.11 ± 1.50 a |
| Water intake (mL/day) | 4.62 ± 0.50 | 4.92 ± 0.40 | 5.05 ± 0.40 a | 5.61 ± 0.50 a | 4.90 ± 0.70 a | 6.10 ± 0.80 a |
| Obesity Induction Phase | Experimental Treatment Phase | |||||
|---|---|---|---|---|---|---|
| SD 14-Week | HFFD 14-Week | SD 26-Week | HFFD 26-Week | BL Consumption | Change to Healthy Diet | |
| Body weight (g) | 25.63 ± 0.57 | 33.47 ± 0.72 * | 27.96 ± 0.65 a | 30.50 ± 0.52 b | 28.09 ± 0.84 ab | 29.47 ± 0.42 a |
| Abdominal circumference (cm) | 7.50 ± 0.46 | 10.40 ± 0.43 * | 7.79 ± 0.29 a | 8.33 ± 0.28 a | 8.50 ± 0.39 a | 6.83 ± 0.17 b |
| Thoracic circumference (cm) | 7.13 ± 0.43 | 9.2 ± 0.73 | 7.36 ± 0.26 a | 8.17 ± 0.25 a | 8.50 ± 0.55 a | 7.33 ± 0.33 a |
| Serum cholesterol (mg/dL) | 92.60 ± 8.60 | 122.80 ± 11.90 * | 98.30 ± 0.90 a | 119.70 ± 3.40 b | 114.40 ± 3.50 b | 104.90 ± 4.20 b |
| Serum triglycerides (mg/dL) | 77.70 ± 7.60 | 137.30 ± 17.50 * | 99.50 ± 3.90 a | 141.10 ± 4.20 b | 110.70 ± 7.30 a | 106.80 ± 7.70 a |
| Obesity Induction Phase | Experimental Treatment Phase | |||||
|---|---|---|---|---|---|---|
| Length (cm) | SD 14-Week | HFFD 14-Week | SD 26-Week | HFFD 26-Week | BL Consumption | Change to Healthy Diet |
| Small intestine | 44.00 ± 2.91 | 36.63 ± 1.58 * | 39.85 ± 1.72 a | 34.31 ± 0.97 a | 37.95 ± 1.04 a | 37.24 ± 1.67 a |
| Total colon | 8.86 ± 0.50 | 6.98 ± 0.50 | 7.28 ± 0.45 a | 6.58 ± 0.52 a | 7.41 ± 0.48 a | 5.43 ± 0.53 b |
| Percentage (%) | SD 26-Week | HFFD 26-Week | BL Consumption | Change to Healthy Diet |
|---|---|---|---|---|
| Colon | 1.82 ± 0.06 a | 1.23 ± 0.03 b | 1.24 ± 0.02 b | 1.31 ± 0.09 b |
| Small intestine | 7.06 ± 0.28 a | 5.77 ± 0.27 b | 6.25 ± 0.14 ab | 6.09 ± 0.14 b |
| Liver | 4.81 ± 0.14 a | 5.93 ± 0.28 ab | 5.58 ± 0.59 a | 4.68 ± 0.21 ac |
| Intake (g/day) | SD 26-Week | HFFD 26-Week | BL Consumption | Change to Healthy Diet |
|---|---|---|---|---|
| Soluble fiber | 0.21 ± 0.02 a | 0.08 ± 0.01 b | 0.24 ± 0.03 a | 0.33 ± 0.04 a |
| Insoluble fiber | 0.81 ± 0.08 a | 0.23 ± 0.02 b | 0.70 ± 0.09 a | 1.08 ± 0.13 a |
| Total dietary fiber | 1.20 ± 0.10 a | 0.27 ± 0.02 b | 0.94 ± 0.02 c | 1.40 ± 0.16 d |
| Total phenolic compounds | 9.59 ± 1.02 a | 7.95 ± 1.24 a | 14.8 ± 2.72 b | 12.87 ± 1.27 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ocampo-Anguiano, P.V.; Victoria-Ruiz, L.L.; Reynoso-Camacho, R.; Olvera-Ramírez, A.M.; Rocha-Guzmán, N.E.; Ramos-Gómez, M.; Ahumada-Solórzano, S.M. Ingestion of Bean Leaves Reduces Metabolic Complications and Restores Intestinal Integrity in C57BL/6 Mice with Obesity Induced by a High-Fat and High-Fructose Diet. Nutrients 2024, 16, 367. https://doi.org/10.3390/nu16030367
Ocampo-Anguiano PV, Victoria-Ruiz LL, Reynoso-Camacho R, Olvera-Ramírez AM, Rocha-Guzmán NE, Ramos-Gómez M, Ahumada-Solórzano SM. Ingestion of Bean Leaves Reduces Metabolic Complications and Restores Intestinal Integrity in C57BL/6 Mice with Obesity Induced by a High-Fat and High-Fructose Diet. Nutrients. 2024; 16(3):367. https://doi.org/10.3390/nu16030367
Chicago/Turabian StyleOcampo-Anguiano, Perla Viridiana, Laura Lizeth Victoria-Ruiz, Rosalía Reynoso-Camacho, Andrea Margarita Olvera-Ramírez, Nuria Elizabeth Rocha-Guzmán, Minerva Ramos-Gómez, and Santiaga Marisela Ahumada-Solórzano. 2024. "Ingestion of Bean Leaves Reduces Metabolic Complications and Restores Intestinal Integrity in C57BL/6 Mice with Obesity Induced by a High-Fat and High-Fructose Diet" Nutrients 16, no. 3: 367. https://doi.org/10.3390/nu16030367
APA StyleOcampo-Anguiano, P. V., Victoria-Ruiz, L. L., Reynoso-Camacho, R., Olvera-Ramírez, A. M., Rocha-Guzmán, N. E., Ramos-Gómez, M., & Ahumada-Solórzano, S. M. (2024). Ingestion of Bean Leaves Reduces Metabolic Complications and Restores Intestinal Integrity in C57BL/6 Mice with Obesity Induced by a High-Fat and High-Fructose Diet. Nutrients, 16(3), 367. https://doi.org/10.3390/nu16030367