
The Umbilical Cord Creatine Flux and Time Course of Human Milk Creatine across Lactation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
- To estimate the placental Cr flux at the end of pregnancy by measuring umbilical arterial and venous cord blood Cr concentrations immediately postpartum after caesarian section.
- To analyze whether the Cr concentration in breast milk is constant during a single breast meal (intraindividual variability).
- To analyze the time course of the association between HM Cr and HM Crn, fat, carbohydrates, and especially protein. It is unknown whether Cr is independently secreted into HM or whether it just parallels, e.g., HM protein or Crn concentration.
- To analyze human breast milk Cr concentration longitudinally, starting with colostrum throughout the first six months of breastfeeding, to study if colostrum has higher Cr levels, suggesting increased immediate postnatal needs.
2. Materials and Methods
2.1. Umbilical Cord Blood Samples
2.2. Collection of Breast Milk
2.3. Longitudinal Study on Breast Milk Creatine in Omnivore Mothers
2.4. Preparation and Analysis of Blood Samples
2.5. Preparation and Analysis of Human Milk Samples
2.6. Statistics
3. Results
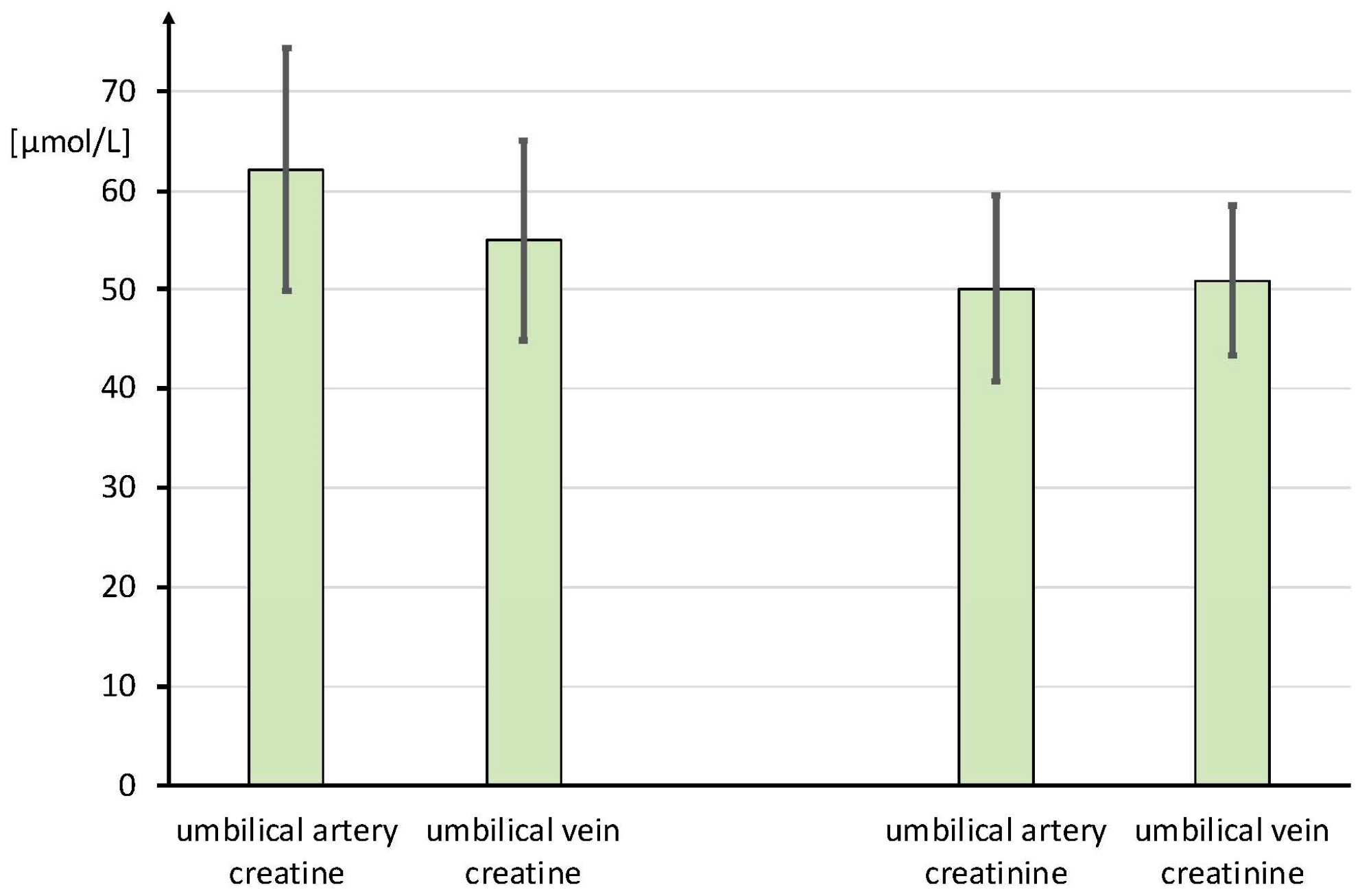
3.1. Umbilical Creatine Flux
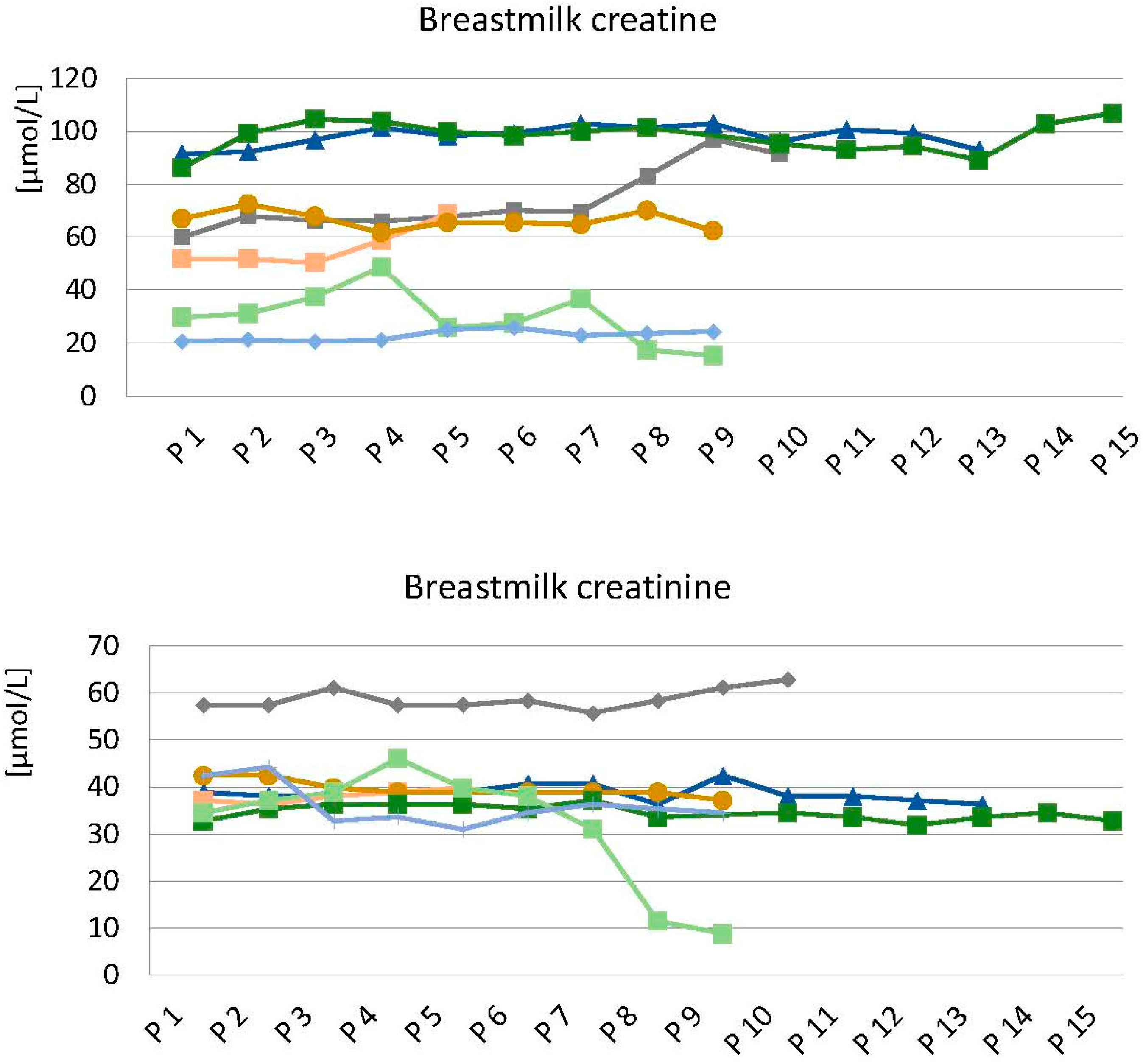
3.2. Intra- and Interindividual Variability in Breast Milk Creatine and Creatinine
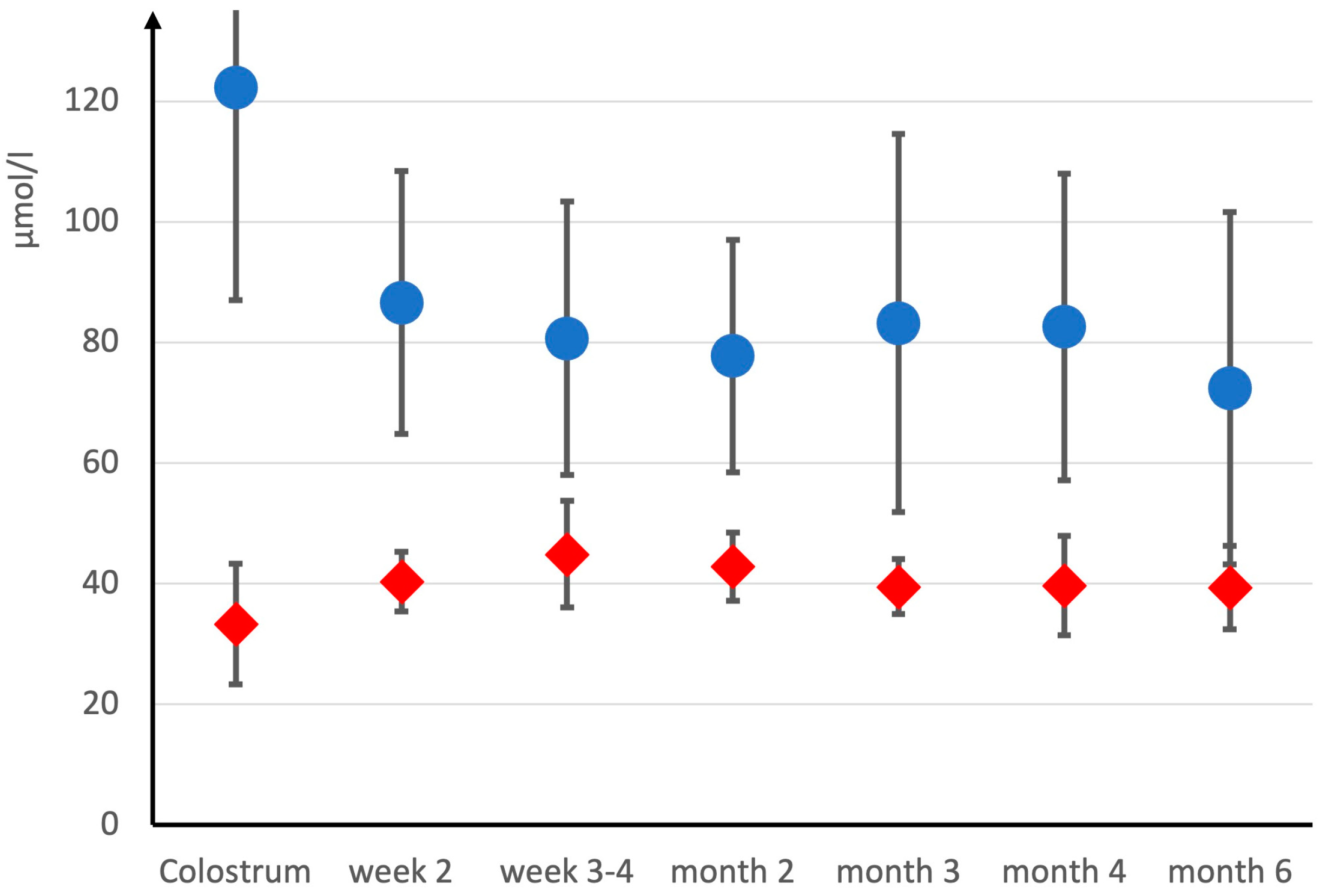
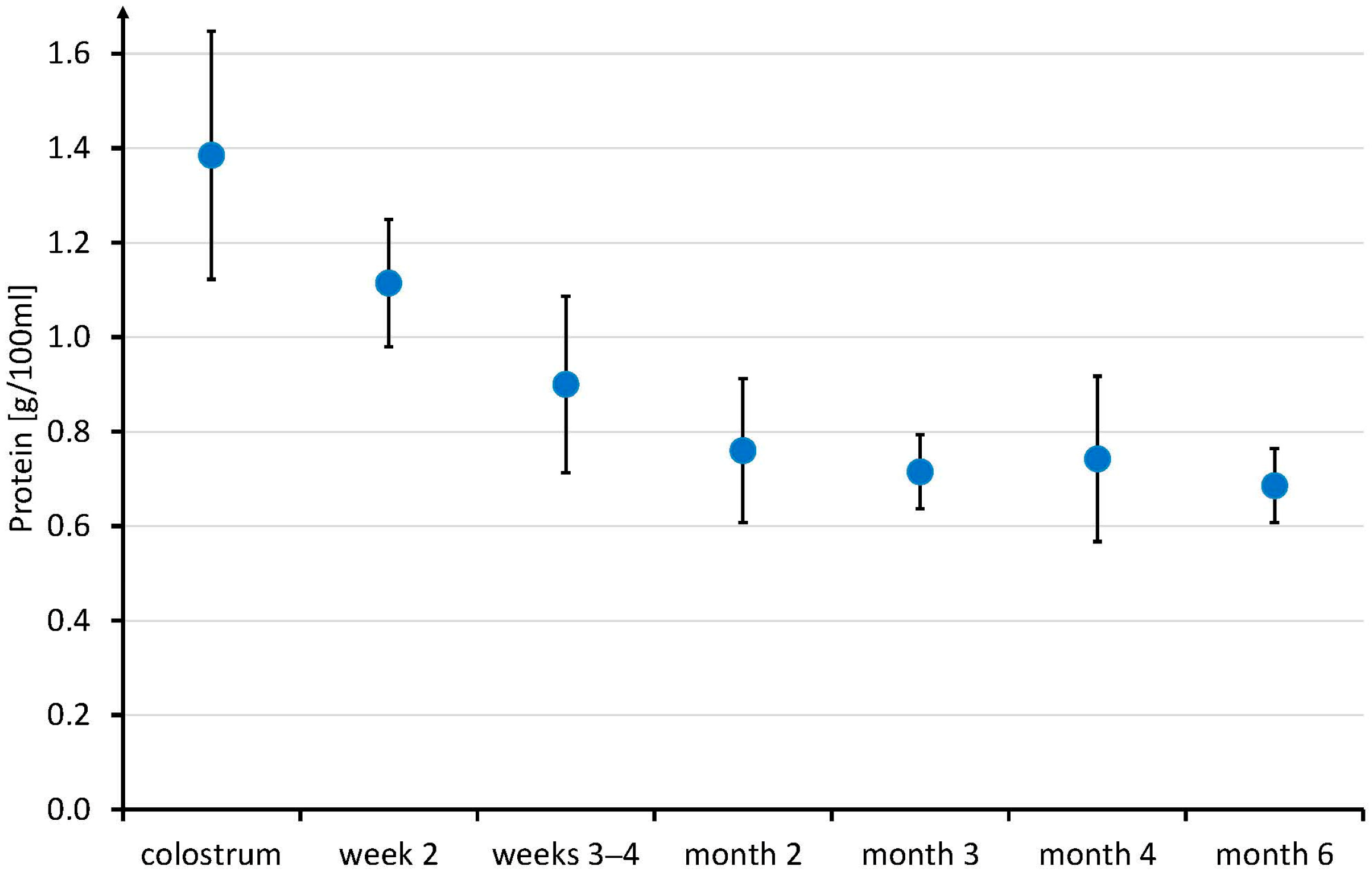
3.3. Longitudinal Study on Breast Milk Creatine in Omnivore Mothers
4. Discussion
4.1. Umbilical Creatine Flux
4.2. Intra- and Interindividual Breast Milk Creatine Variability
4.3. Longitudinal Study on Breast Milk Creatine
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- National Library of Medicine (US). Creatine; National Library of Medicine (US): Bethesda, MD, USA, 2006. [Google Scholar]
- Garwolinska, D.; Namiesnik, J.; Kot-Wasik, A.; Hewelt-Belka, W. Chemistry of Human Breast Milk-A Comprehensive Review of the Composition and Role of Milk Metabolites in Child Development. J. Agric. Food Chem. 2018, 66, 11881–11896. [Google Scholar] [CrossRef]
- Wyss, M.; Kaddurah-Daouk, R. Creatine and creatinine metabolism. Physiol. Rev. 2000, 80, 1107–1213. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.B. Creatine: Biosynthesis, regulation, and function. Adv. Enzymol. Relat. Areas Mol. Biol. 1979, 50, 177–242. [Google Scholar] [CrossRef] [PubMed]
- Brosnan, M.E.; Edison, E.E.; da Silva, R.; Brosnan, J.T. New insights into creatine function and synthesis. Adv. Enzym. Regul. 2007, 47, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Battini, R.; Alessandri, M.G.; Leuzzi, V.; Moro, F.; Tosetti, M.; Bianchi, M.C.; Cioni, G. Arginine:glycine amidinotransferase (AGAT) deficiency in a newborn: Early treatment can prevent phenotypic expression of the disease. J. Pediatr. 2006, 148, 828–830. [Google Scholar] [CrossRef] [PubMed]
- Braissant, O.; Henry, H.; Beard, E.; Uldry, J. Creatine deficiency syndromes and the importance of creatine synthesis in the brain. Amino Acids 2011, 40, 1315–1324. [Google Scholar] [CrossRef] [PubMed]
- Nasrallah, F.; Feki, M.; Kaabachi, N. Creatine and creatine deficiency syndromes: Biochemical and clinical aspects. Pediatr. Neurol. 2010, 42, 163–171. [Google Scholar] [CrossRef]
- Edison, E.E.; Brosnan, M.E.; Aziz, K.; Brosnan, J.T. Creatine and guanidinoacetate content of human milk and infant formulas: Implications for creatine deficiency syndromes and amino acid metabolism. Br. J. Nutr. 2013, 110, 1075–1078. [Google Scholar] [CrossRef]
- Brosnan, M.E.; Brosnan, J.T. The role of dietary creatine. Amino Acids 2016, 48, 1785–1791. [Google Scholar] [CrossRef]
- Ipsiroglu, O.S.; Stromberger, C.; Ilas, J.; Hoger, H.; Muhl, A.; Stockler-Ipsiroglu, S. Changes of tissue creatine concentrations upon oral supplementation of creatine-monohydrate in various animal species. Life Sci. 2001, 69, 1805–1815. [Google Scholar] [CrossRef]
- Kaviani, M.; Shaw, K.; Chilibeck, P.D. Benefits of Creatine Supplementation for Vegetarians Compared to Omnivorous Athletes: A Systematic Review. Int. J. Environ. Res. Public Health 2020, 17, 3041. [Google Scholar] [CrossRef] [PubMed]
- Balsom, P.D.; Soderlund, K.; Ekblom, B. Creatine in humans with special reference to creatine supplementation. Sport. Med. 1994, 18, 268–280. [Google Scholar] [CrossRef] [PubMed]
- Kan, H.E.; van der Graaf, M.; Klomp, D.W.; Vlak, M.H.; Padberg, G.W.; Heerschap, A. Intake of 13C-4 creatine enables simultaneous assessment of creatine and phosphocreatine pools in human skeletal muscle by 13C MR spectroscopy. Magn. Reason. Med. 2006, 56, 953–957. [Google Scholar] [CrossRef] [PubMed]
- Stockler, S.; Holzbach, U.; Hanefeld, F.; Marquardt, I.; Helms, G.; Requart, M.; Hanicke, W.; Frahm, J. Creatine deficiency in the brain: A new, treatable inborn error of metabolism. Pediatr. Res. 1994, 36, 409–413. [Google Scholar] [CrossRef] [PubMed]
- Salomons, G.S.; van Dooren, S.J.; Verhoeven, N.M.; Cecil, K.M.; Ball, W.S.; Degrauw, T.J.; Jakobs, C. X-linked creatine-transporter gene (SLC6A8) defect: A new creatine-deficiency syndrome. Am. J. Hum. Genet. 2001, 68, 1497–1500. [Google Scholar] [CrossRef] [PubMed]
- Schulze, A.; Ebinger, F.; Rating, D.; Mayatepek, E. Improving treatment of guanidinoacetate methyltransferase deficiency: Reduction of guanidinoacetic acid in body fluids by arginine restriction and ornithine supplementation. Mol. Genet. Metab. 2001, 74, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.C.; Nevill, M.; Harris, D.B.; Fallowfield, J.L.; Bogdanis, G.C.; Wise, J.A. Absorption of creatine supplied as a drink, in meat or in solid form. J. Sport. Sci. 2002, 20, 147–151. [Google Scholar] [CrossRef]
- Hulsemann, J.; Manz, F.; Wember, T.; Schoch, G. Administration of creatine and creatinine with breast milk and infant milk preparations. Klin. Padiatr. 1987, 199, 292–295. [Google Scholar] [CrossRef]
- Spevacek, A.R.; Smilowitz, J.T.; Chin, E.L.; Underwood, M.A.; German, J.B.; Slupsky, C.M. Infant Maturity at Birth Reveals Minor Differences in the Maternal Milk Metabolome in the First Month of Lactation. J. Nutr. 2015, 145, 1698–1708. [Google Scholar] [CrossRef]
- Wu, J.; Domellof, M.; Zivkovic, A.M.; Larsson, G.; Ohman, A.; Nording, M.L. NMR-based metabolite profiling of human milk: A pilot study of methods for investigating compositional changes during lactation. Biochem. Biophys. Res. Commun. 2016, 469, 626–632. [Google Scholar] [CrossRef]
- Creatine Disorders Panel CRDPP. Available online: https://www.mayocliniclabs.com/api/sitecore/TestCatalog/DownloadTestCatalog?testId=606130 (accessed on 5 May 2023).
- Lamarre, S.G.; Edison, E.E.; Wijekoon, E.P.; Brosnan, M.E.; Brosnan, J.T. Suckling rat pups accumulate creatine primarily via de novo synthesis rather than from dam milk. J. Nutr. 2010, 140, 1570–1573. [Google Scholar] [CrossRef]
- Ireland, Z.; Russell, A.P.; Wallimann, T.; Walker, D.W.; Snow, R. Developmental changes in the expression of creatine synthesizing enzymes and creatine transporter in a precocial rodent, the spiny mouse. BMC Dev. Biol. 2009, 9, 39. [Google Scholar] [CrossRef] [PubMed]
- Lage, S.; Andrade, F.; Prieto, J.A.; Asla, I.; Rodriguez, A.; Ruiz, N.; Echeverria, J.; Luz Couce, M.; Sanjurjo, P.; Aldamiz-Echevarria, L. Arginine-guanidinoacetate-creatine pathway in preterm newborns: Creatine biosynthesis in newborns. J. Pediatr. Endocrinol. Metab. 2013, 26, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Muccini, A.M.; Tran, N.T.; de Guingand, D.L.; Philip, M.; Della Gatta, P.A.; Galinsky, R.; Sherman, L.S.; Kelleher, M.A.; Palmer, K.R.; Berry, M.J.; et al. Creatine Metabolism in Female Reproduction, Pregnancy and Newborn Health. Nutrients 2021, 13, 490. [Google Scholar] [CrossRef] [PubMed]
- Tran, N.T.; Kelly, S.B.; Snow, R.J.; Walker, D.W.; Ellery, S.J.; Galinsky, R. Assessing Creatine Supplementation for Neuroprotection against Perinatal Hypoxic-Ischaemic Encephalopathy: A Systematic Review of Perinatal and Adult Pre-Clinical Studies. Cells 2021, 10, 2902. [Google Scholar] [CrossRef] [PubMed]
- Tran, N.T.; Kowalski, G.M.; Muccini, A.M.; Nitsos, I.; Hale, N.; Snow, R.J.; Walker, D.W.; Ellery, S.J. Creatine supplementation reduces the cerebral oxidative and metabolic stress responses to acute in utero hypoxia in the late-gestation fetal sheep. J. Physiol. 2022, 600, 3193–3210. [Google Scholar] [CrossRef] [PubMed]
- Davidson, J.O.; Gunn, A.J.; Dean, J.M. Back to the beginning: Can we stop brain injury before it starts? J. Physiol. 2022, 600, 3013–3014. [Google Scholar] [CrossRef]
- Heland, S.; Fields, N.; Ellery, S.J.; Fahey, M.; Palmer, K.R. The role of nutrients in human neurodevelopment and their potential to prevent neurodevelopmental adversity. Front. Nutr. 2022, 9, 992120. [Google Scholar] [CrossRef]
- Bodamer, O.A.; Bloesch, S.M.; Gregg, A.R.; Stockler-Ipsiroglu, S.; O’Brien, W.E. Analysis of guanidinoacetate and creatine by isotope dilution electrospray tandem mass spectrometry. Clin. Chim. Acta 2001, 308, 173–178. [Google Scholar] [CrossRef]
- Bernt, E.; Bergmeyer, H.U.; Moellering, H. Creatine. In Methoden der Enzymatischen Analysen, 7th ed.; Bergmeyer, H.U., Ed.; Verlag Chemie: Weinheim, Germany, 1985; Volume II, p. 505ff. [Google Scholar]
- Jaffe, M. Ueber den Niederschlag, welchen Pikrinsaeure in normalem Harn erzeugt und über eine neue Reaction des Kreatinins. Z. Für Physiol. Chem. 1886, 10, 391–400. [Google Scholar]
- Kiserud, T.; Ebbing, C.; Kessler, J.; Rasmussen, S. Fetal cardiac output, distribution to the placenta and impact of placental compromise. Ultrasound Obstet. Gynecol. 2006, 28, 126–136. [Google Scholar] [CrossRef]
- Menjo, A.; Mizuno, K.; Murase, M.; Nishida, Y.; Taki, M.; Itabashi, K.; Shimono, T.; Namba, K. Bedside analysis of human milk for adjustable nutrition strategy. Acta Paediatr. 2009, 98, 380–384. [Google Scholar] [CrossRef]
- Fusch, G.; Kwan, C.; Kotrri, G.; Fusch, C. “Bed Side” Human Milk Analysis in the Neonatal Intensive Care Unit: A Systematic Review. Clin. Perinatol. 2017, 44, 209–267. [Google Scholar] [CrossRef]
- Bakdash, J.; Marusich, L. Repeated Measures Correlation, R Package Version 0.5.4, R_Foundation; 2022. Available online: https://CRAN.R-project.org/package=rmcorr (accessed on 5 May 2023).
- Lentner, C. Geigy Scientific Tables, 8th ed.; Medical Education Division, Ciba-Geigy Corp.: Basel, Switzerland, 1992. [Google Scholar]
- Andresen, J.; Zegarra, J.A.; Estrada, C.A.; Dumler, F. Serum creatinine concentrations in healthy newborns: Reference ranges during the first five days of life. Henry Ford. Hosp. Med. J. 1988, 36, 193–194. [Google Scholar]
- Gidrewicz, D.A.; Fenton, T.R. A systematic review and meta-analysis of the nutrient content of preterm and term breast milk. BMC Pediatr. 2014, 14, 216. [Google Scholar] [CrossRef] [PubMed]
- Ng, W.W.; Miller, R.K. Transport of Nutrients in the Early Human Placenta: Amino Acid, Creatine, Vitamin B12. In Fetal Nutrition, Metabolism, and Immunology; Trophoblast, Research; Miller, R.K., Thiede, H.A., Eds.; Springer: Boston, MA, USA, 1984; Volume 1, pp. 121–134. [Google Scholar]
- Ellery, S.J.; Della Gatta, P.A.; Bruce, C.R.; Kowalski, G.M.; Davies-Tuck, M.; Mockler, J.C.; Murthi, P.; Walker, D.W.; Snow, R.J.; Dickinson, H. Creatine biosynthesis and transport by the term human placenta. Placenta 2017, 52, 86–93. [Google Scholar] [CrossRef]
- Miller, R.K.; Berndt, W.O. Mechanisms of transport across the placenta: An in vitro approach. Life Sci. 1975, 16, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Baharom, S.; De Matteo, R.; Ellery, S.; Della Gatta, P.; Bruce, C.R.; Kowalski, G.M.; Hale, N.; Dickinson, H.; Harding, R.; Walker, D.; et al. Does maternal-fetal transfer of creatine occur in pregnant sheep? Am. J. Physiol. Endocrinol. Metab. 2017, 313, E75–E83. [Google Scholar] [CrossRef] [PubMed]
- Davis, B.M.; Miller, R.K.; Brent, R.L.; Koszalka, T.R. Materno-fetal transport of creatine in the rat. Biol. Neonate 1978, 33, 43–54. [Google Scholar] [CrossRef]
- Ellery, S.J.; Murthi, P.; Davies-Tuck, M.L.; Della Gatta, P.A.; May, A.K.; Kowalski, G.M.; Callahan, D.L.; Bruce, C.R.; Alers, N.O.; Miller, S.L.; et al. Placental creatine metabolism in cases of placental insufficiency and reduced fetal growth. Mol. Hum. Reprod. 2019, 25, 495–505. [Google Scholar] [CrossRef]
- Braissant, O.; Henry, H.; Villard, A.M.; Speer, O.; Wallimann, T.; Bachmann, C. Creatine synthesis and transport during rat embryogenesis: Spatiotemporal expression of AGAT, GAMT and CT1. BMC Dev. Biol. 2005, 5, 9. [Google Scholar] [CrossRef] [PubMed]
- Haughton, C.L.; Gawriluk, T.R.; Seifert, A.W. The Biology and Husbandry of the African Spiny Mouse (Acomys cahirinus) and the Research Uses of a Laboratory Colony. J. Am. Assoc. Lab. Anim. Sci. 2016, 55, 9–17. [Google Scholar] [PubMed]
- Neville, M.C.; Keller, R.P.; Seacat, J.; Casey, C.E.; Allen, J.C.; Archer, P. Studies on human lactation. I. Within-feed and between-breast variation in selected components of human milk. Am. J. Clin. Nutr. 1984, 40, 635–646. [Google Scholar] [CrossRef]
- Rodriguez, G.; Ventura, P.; Samper, M.P.; Moreno, L.; Sarria, A.; Perez-Gonzalez, J.M. Changes in body composition during the initial hours of life in breast-fed healthy term newborns. Biol. Neonate 2000, 77, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Roggero, P.; Gianni, M.L.; Orsi, A.; Piemontese, P.; Amato, O.; Moioli, C.; Mosca, F. Neonatal period: Body composition changes in breast-fed full-term newborns. Neonatology 2010, 97, 139–143. [Google Scholar] [CrossRef]
- Santacruz, L.; Arciniegas, A.J.L.; Darrabie, M.; Mantilla, J.G.; Baron, R.M.; Bowles, D.E.; Mishra, R.; Jacobs, D.O. Hypoxia decreases creatine uptake in cardiomyocytes, while creatine supplementation enhances HIF activation. Physiol. Rep. 2017, 5, e13382. [Google Scholar] [CrossRef]
- Lawn, J.; Shibuya, K.; Stein, C. No cry at birth: Global estimates of intrapartum stillbirths and intrapartum-related neonatal deaths. Bull. World Health Organ. 2005, 83, 409–417. [Google Scholar]
- Berry, M.J.; Schlegel, M.; Kowalski, G.M.; Bruce, C.R.; Callahan, D.L.; Davies-Tuck, M.L.; Dickinson, H.; Goodson, A.; Slocombe, A.; Snow, R.J.; et al. UNICORN Babies: Understanding Circulating and Cerebral Creatine Levels of the Preterm Infant. An Observational Study Protocol. Front. Physiol. 2019, 10, 142. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mihatsch, W.A.; Stahl, B.; Braun, U. The Umbilical Cord Creatine Flux and Time Course of Human Milk Creatine across Lactation. Nutrients 2024, 16, 345. https://doi.org/10.3390/nu16030345
Mihatsch WA, Stahl B, Braun U. The Umbilical Cord Creatine Flux and Time Course of Human Milk Creatine across Lactation. Nutrients. 2024; 16(3):345. https://doi.org/10.3390/nu16030345
Chicago/Turabian StyleMihatsch, Walter Alexander, Bernd Stahl, and Ulrike Braun. 2024. "The Umbilical Cord Creatine Flux and Time Course of Human Milk Creatine across Lactation" Nutrients 16, no. 3: 345. https://doi.org/10.3390/nu16030345
APA StyleMihatsch, W. A., Stahl, B., & Braun, U. (2024). The Umbilical Cord Creatine Flux and Time Course of Human Milk Creatine across Lactation. Nutrients, 16(3), 345. https://doi.org/10.3390/nu16030345