
The Effects of Polyphenol Supplementation on BDNF, Cytokines and Cognition in Trained Male Cyclists following Acute Ozone Exposure during High-Intensity Cycling

Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
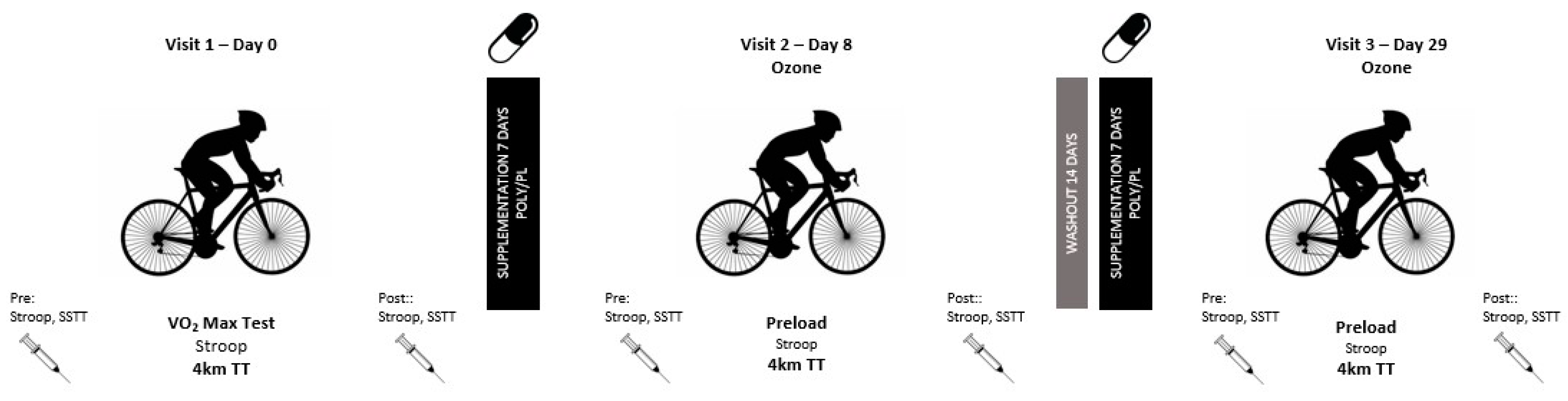
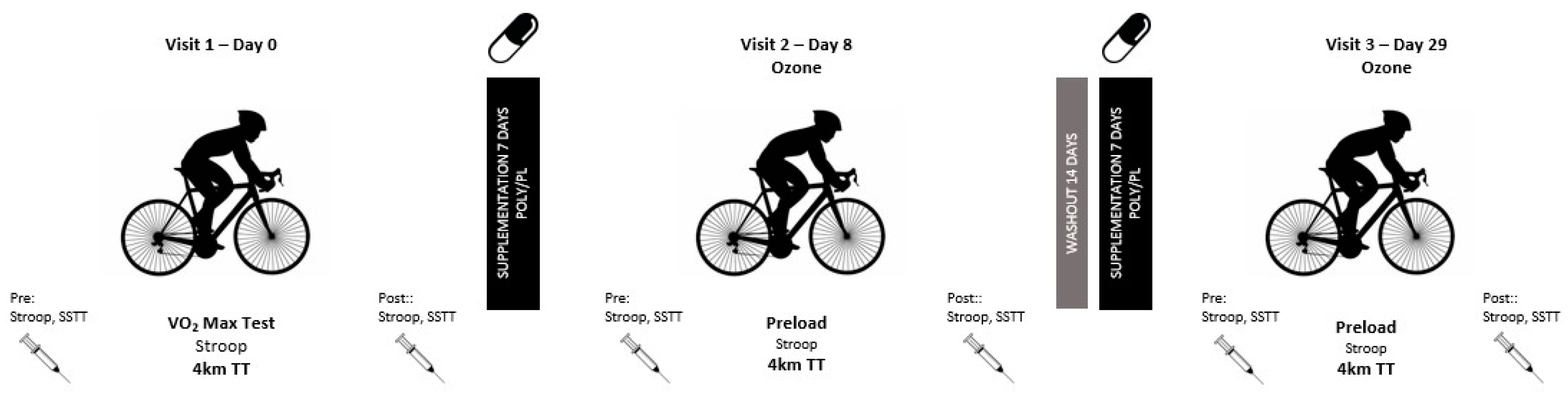
2.2. Experimental Design
2.3. Ozone Exposure
2.4. Experimental Trial
2.5. Blood Sampling
2.6. Sample Analysis
2.6.1. BDNF
2.6.2. IL-6, IL-10 and tnf-α
2.7. Cognitive Tests
2.7.1. Stroop Test
2.7.2. Serial Subtraction Task (3’s and 7’s)
“I want you to perform a mental, cognitive task that requires subtracting. I am going to say a number and I would like you to subtract 3/7 from that number and continue to subtract 3/7 from your answer until I tell you to stop. Work as quickly as you can and try not to make any mis-takes. Any questions? Ready? Here is the number XX”.
2.8. Data Analysis Procedures and Statistical Analysis
3. Results
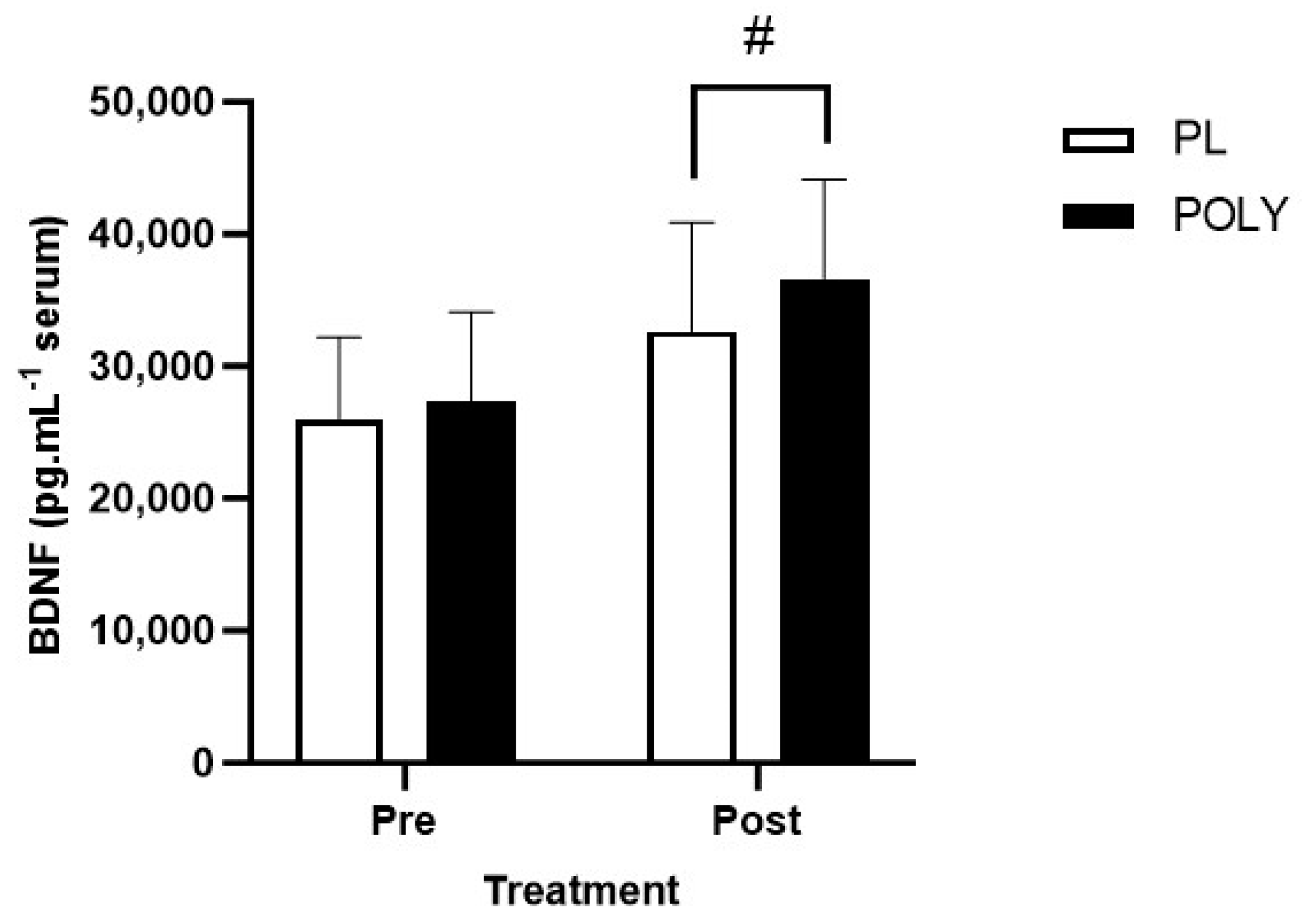
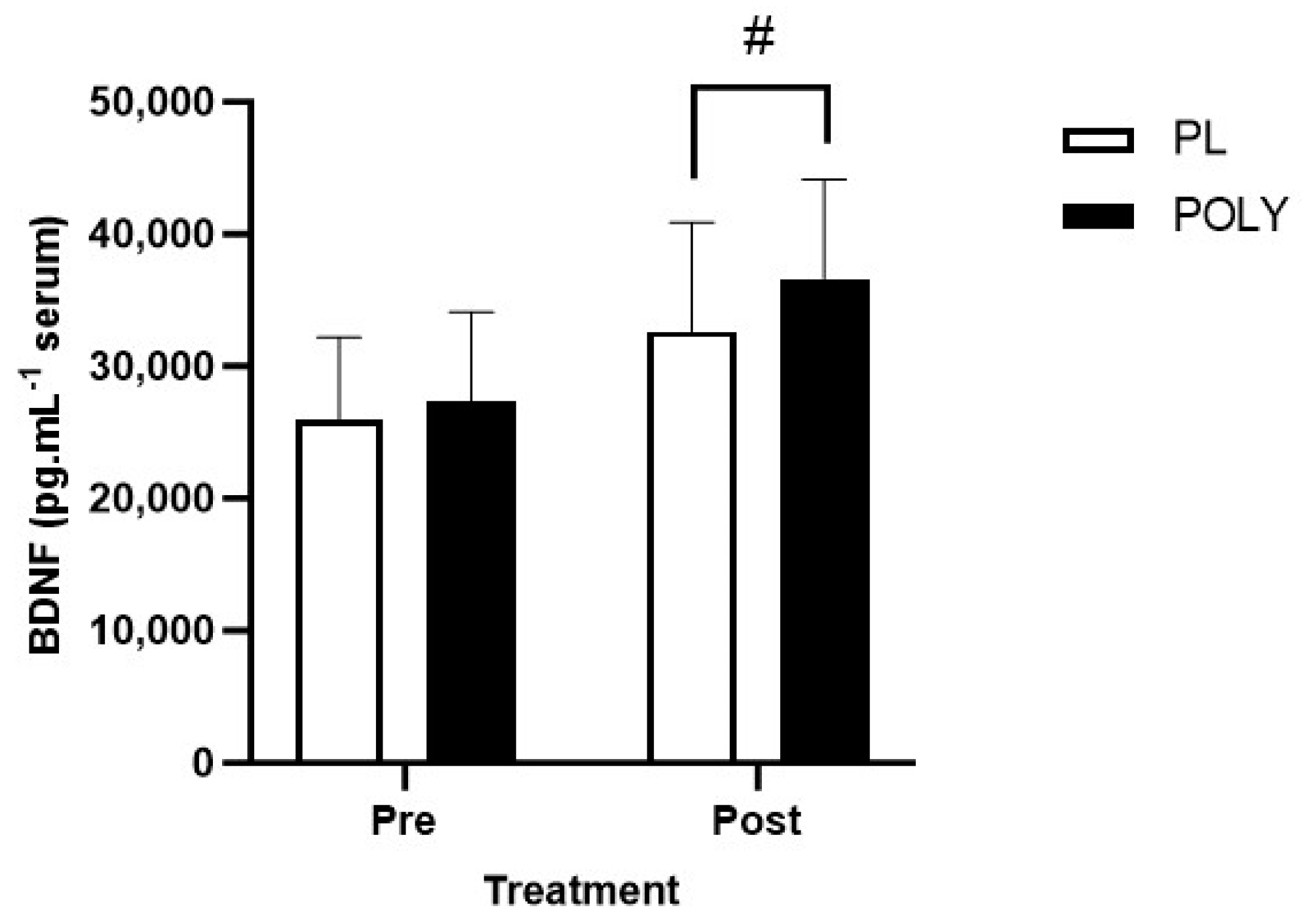
3.1. BDNF
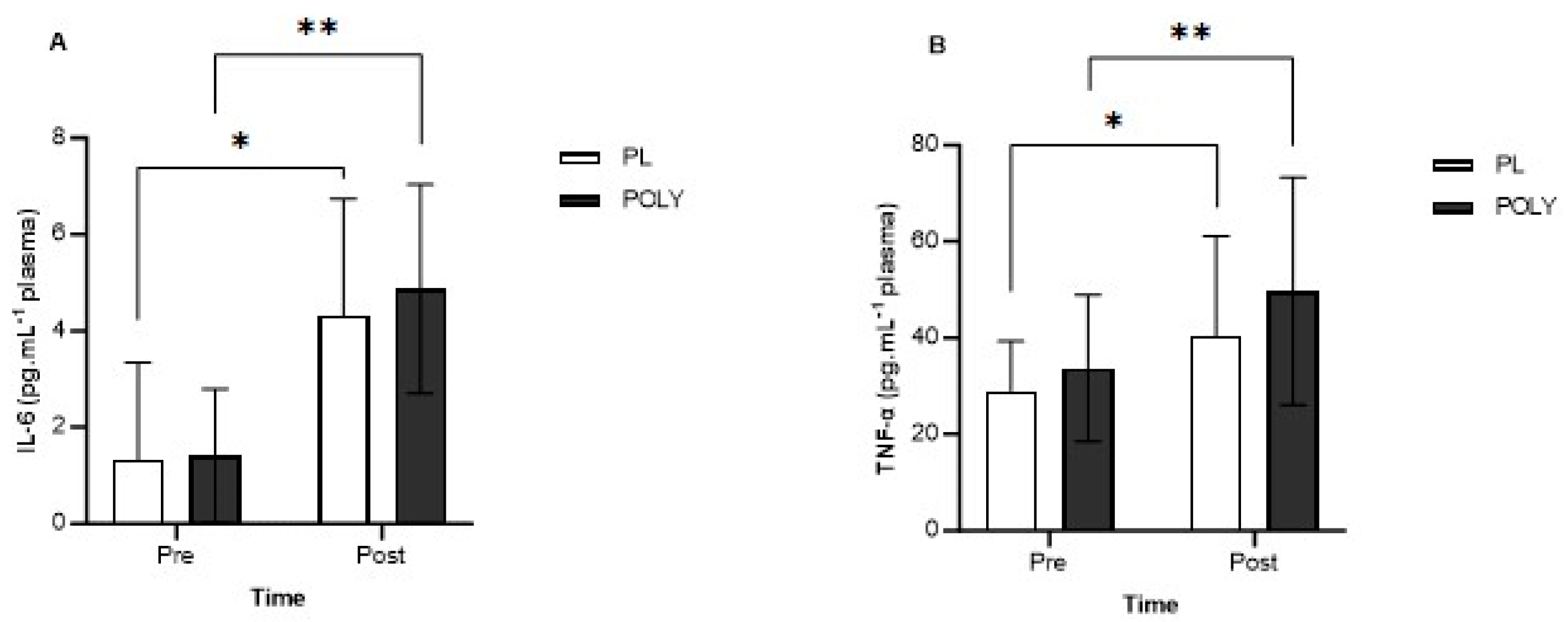
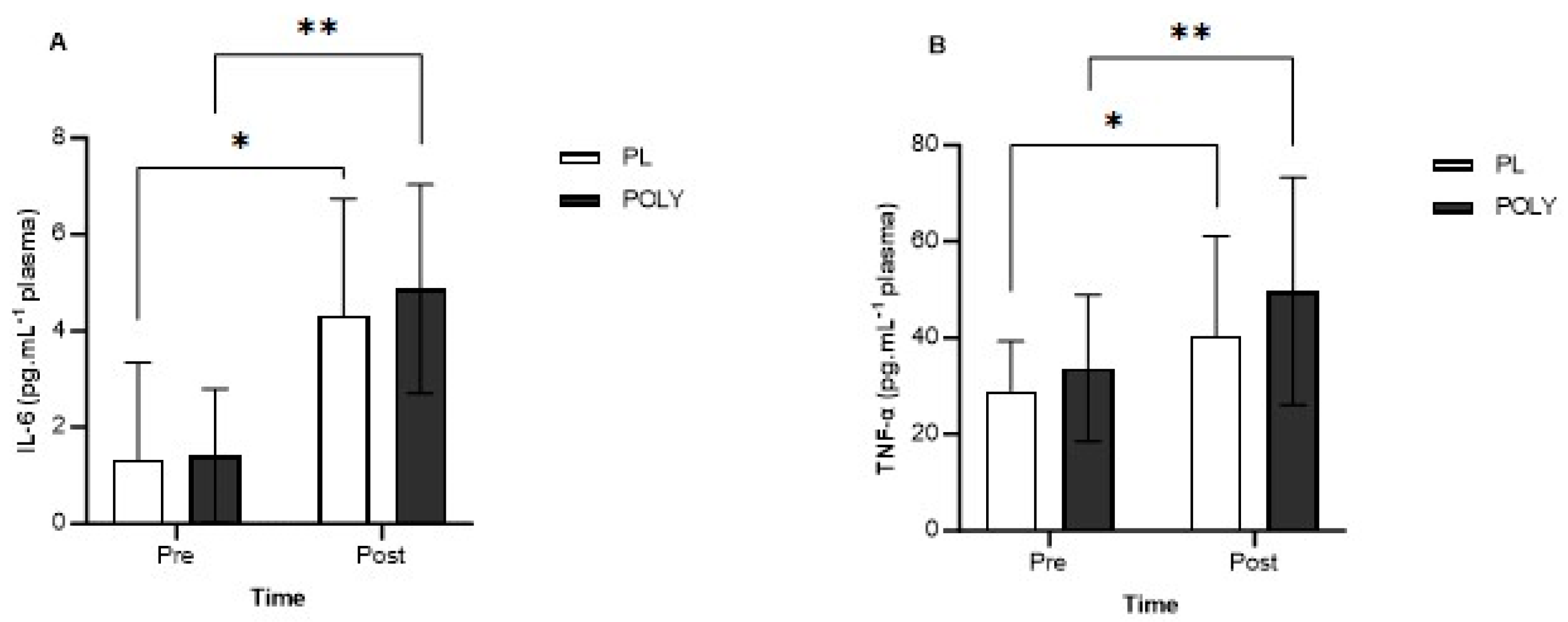
3.2. IL-6, IL-10 and TNF-α
3.3. Stroop Test
3.4. Serial Subtraction Task
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bromberg, P.A. Mechanisms of the acute effects of inhaled ozone in humans. Biochim. Biophys. Acta BBA-Gen. Subj. 2016, 1860, 2771–2781. [Google Scholar] [CrossRef] [PubMed]
- Bello-Medina, P.C.; Rodríguez-Martínez, E.; Prado-Alcalá, R.A.; Rivas-Arancibia, S. Ozone pollution, oxidative stress, synaptic plasticity, and neurodegeneration. Neurología 2022, 37, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Bosson, J.; Barath, S.; Pourazar, J.; Behndig, A.F.; Sandström, T.; Blomberg, A.; Ädelroth, E. Diesel exhaust exposure enhances the ozone-induced airway inflammation in healthy humans. Eur. Respir. J. 2008, 31, 1234–1240. [Google Scholar] [CrossRef]
- Gackière, F.; Saliba, L.; Baude, A.; Bosler, O.; Strube, C. Ozone inhalation activates stress-responsive regions of the CNS. J. Neurochem. 2011, 117, 961–972. [Google Scholar] [CrossRef] [PubMed]
- Galiè, M.; Covi, V.; Tabaracci, G.; Malatesta, M. The role of Nrf2 in the antioxidant cellular response to medical ozone exposure. Int. J. Mol. Sci. 2019, 20, 4009. [Google Scholar] [CrossRef]
- Nery-Flores, S.D.; Ramírez-Herrera, M.A.; Mendoza-Magaña, M.L.; Romero-Prado, M.M.; Ramírez-Vázquez, J.D.; Bañuelos-Pineda, J.; Espinoza-Gutiérrez, H.A.; Ramírez-Mendoza, A.A.; Tostado, M.C. Dietary curcumin prevented astrocytosis, microgliosis, and apoptosis caused by acute and chronic exposure to ozone. Molecules 2019, 24, 2839. [Google Scholar] [CrossRef] [PubMed]
- Croze, M.L.; Zimmer, L. Ozone atmospheric pollution and Alzheimer’s disease: From epidemiological facts to molecular mechanisms. J. Alzheimer’s Dis. 2018, 62, 503–522. [Google Scholar] [CrossRef]
- Chen, M.C.; Wang, C.F.; Lai, B.C.; Hsieh, S.W.; Chen, S.C.; Hung, C.H.; Kuo, C.H. Air pollution is associated with poor cognitive function in Taiwanese adults. Int. J. Environ. Res. Public Health 2021, 18, 316. [Google Scholar] [CrossRef]
- Gao, Q.; Zang, E.; Bi, J.; Dubrow, R.; Lowe, S.R.; Chen, H.; Zeng, Y.; Shi, L.; Chen, K. Long-term ozone exposure and cognitive impairment among Chinese older adults: A cohort study. Environ. Int. 2022, 160, 107072. [Google Scholar] [CrossRef]
- Lo, Y.T.; Lu, Y.C.; Chang, Y.H.; Kao, S.; Huang, H.B. Air pollution exposure and cognitive function in Taiwanese older adults: A repeated measurement study. Int. J. Environ. Res. Public Health 2019, 16, 2976. [Google Scholar] [CrossRef]
- Power, M.C.; Adar, S.D.; Yanosky, J.D.; Weuve, J. Exposure to air pollution as a potential contributor to cognitive function, cognitive decline, brain imaging, and dementia: A systematic review of epidemiologic research. Neurotoxicology 2016, 56, 235–253. [Google Scholar] [CrossRef] [PubMed]
- Erickson, M.A.; Jude, J.; Zhao, H.; Rhea, E.M.; Salameh, T.S.; Jester, W.; Pu, S.; Harrowitz, J.; Nguyen, N.; Banks, W.A.; et al. Serum amyloid A: An ozone-induced circulating factor with potentially important functions in the lung-brain axis. FASEB J. 2017, 31, 3950. [Google Scholar] [CrossRef] [PubMed]
- Gaetani, L.; Paoletti, F.P.; Bellomo, G.; Mancini, A.; Simoni, S.; Di Filippo, M.; Parnetti, L. CSF and blood biomarkers in neuroinflammatory and neurodegenerative diseases: Implications for treatment. Trends Pharmacol. Sci. 2020, 41, 1023–1037. [Google Scholar] [CrossRef]
- Terrone, G.; Frigerio, F.; Balosso, S.; Ravizza, T.; Vezzani, A. Inflammation and reactive oxygen species in status epilepticus: Biomarkers and implications for therapy. Epilepsy Behav. 2019, 101, 106275. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Markevych, I.; Romanos, M.; Nowak, D.; Heinrich, J. Ambient ozone exposure and mental health: A systematic review of epidemiological studies. Environ. Res. 2018, 165, 459–472. [Google Scholar] [CrossRef] [PubMed]
- Schuch, F.B.; da Silveira, L.E.; de Zeni, T.C.; da Silva, D.P.; Wollenhaupt-Aguiar, B.; Ferrari, P.; Reischak-Oliveira, Á.; Kapczinski, F. Effects of a single bout of maximal aerobic exercise on BDNF in bipolar disorder: A gender-based response. Psychiatry Res. 2015, 229, 57–62. [Google Scholar] [CrossRef]
- Jiménez-Maldonado, A.; Rentería, I.; García-Suárez, P.C.; Moncada-Jiménez, J.; Freire-Royes, L.F. The impact of high-intensity interval training on brain derived neurotrophic factor in brain: A mini-review. Front. Neurosci. 2018, 12, 839. [Google Scholar] [CrossRef]
- Bos, I.; Jacobs, L.; Nawrot, T.S.; De Geus, B.; Torfs, R.; Panis, L.I.; Degraeuwe, B.; Meeusen, R. No exercise-induced increase in serum BDNF after cycling near a major traffic road. Neurosci. Lett. 2011, 500, 129–132. [Google Scholar] [CrossRef]
- Bos, I.; De Boever, P.; Int Panis, L.; Meeusen, R. Physical activity, air pollution and the brain. Sports Med. 2014, 44, 1505–1518. [Google Scholar] [CrossRef]
- Bos, I.; De Boever, P.; Vanparijs, J.; Pattyn, N.; Panis, L.I.; Meeusen, R. Subclinical effects of aerobic training in urban environment. Med. Sci. Sports Exerc. 2013, 45, 439–447. [Google Scholar] [CrossRef]
- Griffin, É.W.; Mullally, S.; Foley, C.; Warmington, S.A.; O’Mara, S.M.; Kelly, Á.M. Aerobic exercise improves hippocampal function and increases BDNF in the serum of young adult males. Physiol. Behav. 2011, 104, 934–941. [Google Scholar] [CrossRef] [PubMed]
- Miodownik, C.; Maayan, R.; Ratner, Y.; Lerner, V.; Pintov, L.; Mar, M.; Weizman, A.; Ritsner, M.S. Serum levels of brain-derived neurotrophic factor and cortisol to sulfate of dehydroepiandrosterone molar ratio associated with clinical response to L-theanine as augmentation of antipsychotic therapy in schizophrenia and schizoaffective disorder patients. Clin. Neuropharmacol. 2011, 34, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Revi, N.; Rengan, A.K. Impact of dietary polyphenols on neuroinflammation-associated disorders. Neurol. Sci. 2021, 42, 3101–3119. [Google Scholar] [CrossRef]
- Choi, D.Y.; Lee, Y.J.; Hong, J.T.; Lee, H.J. Antioxidant properties of natural polyphenols and their therapeutic potentials for Alzheimer’s disease. Brain Res. Bull. 2012, 87, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Gravesteijn, E.; Mensink, R.P.; Plat, J. Effects of nutritional interventions on BDNF concentrations in humans: A systematic review. Nutr. Neurosci. 2022, 25, 1425–1436. [Google Scholar] [CrossRef]
- Hidese, S.; Ogawa, S.; Ota, M.; Ishida, I.; Yasukawa, Z.; Ozeki, M.; Kunugi, H. Effects of L-theanine administration on stress-related symptoms and cognitive functions in healthy adults: A randomized controlled trial. Nutrients 2019, 11, 2362. [Google Scholar] [CrossRef]
- Jiang, T.A. Health benefits of culinary herbs and spices. J. AOAC Int. 2019, 102, 395–411. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, K.; Matsumoto, T.; Kumazawa, Y. Effects of antioxidant polyphenols on TNF-alpha-related diseases. Curr. Top. Med. Chem. 2011, 11, 1767–1779. [Google Scholar] [CrossRef]
- Lee, W.H.; Loo, C.Y.; Bebawy, M.; Luk, F.; Mason, R.S.; Rohanizadeh, R. Curcumin and its derivatives: Their application in neuropharmacology and neuroscience in the 21st century. Curr. Neuropharmacol. 2013, 11, 338–378. [Google Scholar] [CrossRef]
- Sangiovanni, E.; Brivio, P.; Dell’Agli, M.; Calabrese, F. Botanicals as modulators of neuroplasticity: Focus on BDNF. Neural Plast. 2017, 2017, 5965371. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, K.; Zhai, M.; Lin, X.; Hu, Y.; Feng, L.; Yang, J.; Yu, H.; Wu, C. Protective effect and mechanism of Lycium barbarum L. polyphenol on cognitive impairment induced by ethanol in mice. Phytomedicine 2022, 100, 154033. [Google Scholar] [CrossRef] [PubMed]
- Zhu, A.; Yuan, C.; Pretty, J.; Ji, J.S. Plant-based dietary patterns and cognitive function: A prospective cohort analysis of elderly individuals in China (2008–2018). Brain Behav. 2022, 12, e2670. [Google Scholar] [CrossRef] [PubMed]
- Ammar, A.; Trabelsi, K.; Boukhris, O.; Bouaziz, B.; Mueller, P.; Glenn, J.M.; Bott, N.T.; Mueller, N.; Chtourou, H.; Driss, T.; et al. Effects of polyphenol-rich interventions on cognition and brain health in healthy young and middle-aged adults: Systematic review and meta-analysis. J. Clin. Med. 2020, 9, 1598. [Google Scholar] [CrossRef]
- Bao, M.J.; Shen, J.; Jia, Y.L.; Li, F.F.; Ma, W.J.; Shen, H.J.; Shen, L.L.; Lin, X.X.; Zhang, L.H.; Dong, X.W.; et al. Apple polyphenol protects against cigarette smoke-induced acute lung injury. Nutrition 2013, 29, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Larsen, V.; Thawer, N.; Charles, D.; Cassidy, A.; Van Zele, T.; Thilsing, T.; Ahlström, M.; Haahtela, T.; Keil, T.; Matricardi, P.M.; et al. Dietary intake of flavonoids and ventilatory function in European adults: A GA2LEN study. Nutrients 2018, 10, 95. [Google Scholar] [CrossRef]
- Nyanhanda, T.; Gould, E.M.; McGhie, T.; Shaw, O.M.; Harper, J.L.; Hurst, R.D. Blackcurrant cultivar polyphenolic extracts suppress CCL26 secretion from alveolar epithelial cells. Food Funct. 2014, 5, 671–677. [Google Scholar] [CrossRef]
- Qi, G.; Mi, Y.; Fan, R.; Zhao, B.; Ren, B.; Liu, X. Tea polyphenols ameliorates neural redox imbalance and mitochondrial dysfunction via mechanisms linking the key circadian regular Bmal1. Food Chem. Toxicol. 2017, 110, 189–199. [Google Scholar] [CrossRef]
- Shaw, O.M.; Nyanhanda, T.; McGhie, T.K.; Harper, J.L.; Hurst, R.D. Blackcurrant anthocyanins modulate CCL11 secretion and suppress allergic airway inflammation. Mol. Nutr. Food Res. 2017, 61, 1600868. [Google Scholar] [CrossRef]
- Tabak, C.; Arts, I.C.; Smit, H.A.; Heederik, D.; Kromhout, D. Chronic obstructive pulmonary disease and intake of catechins, flavonols, and flavones: The MORGEN Study. Am. J. Respir. Crit. Care Med. 2001, 164, 61–64. [Google Scholar] [CrossRef]
- Vergara, D.; Ávila, D.; Escobar, E.; Carrasco-Pozo, C.; Sánchez, A.; Gotteland, M. The intake of maqui (Aristotelia chilensis) berry extract normalizes H2O2 and IL-6 concentrations in exhaled breath condensate from healthy smokers-an explorative study. Nutr. J. 2015, 14, 1–5. [Google Scholar] [CrossRef]
- Preedy, V.R.; Watson, R.R.; Zibadi, S. (Eds.) Polyphenols in Human Health and Disease; Academic Press: Cambridge, MA, USA, 2014. [Google Scholar]
- Pounis, G.; Arcari, A.; Costanzo, S.; Di Castelnuovo, A.; Bonaccio, M.; Persichillo, M.; Donati, M.B.; de Gaetano, G.; Iacoviello, L. Favorable association of polyphenol-rich diets with lung function: Cross-sectional findings from the Moli-sani study. Respir. Med. 2018, 136, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Morton, L.C.; Paton, C.D.; Merry, T.; Braakhuis, A.J. Effects of 7-day polyphenol powder supplementation on cycling performance and lung function in an ozone-polluted environment. Eur. J. Appl. Physiol. 2023, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Marquez, C.M.; Vanaudenaerde, B.; Troosters, T.; Wenderoth, N. High-intensity interval training evokes larger serum BDNF levels compared with intense continuous exercise. J. Appl. Physiol. 2015, 119, 1363–1373. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, J.S.; Heuman, D.M.; Sterling, R.K.; Sanyal, A.J.; Siddiqui, M.; Matherly, S.; Luketic, V.; Stravitz, R.T.; Fuchs, M.; Thacker, L.R.; et al. Validation of EncephalApp, smartphone-based Stroop test, for the diagnosis of covert hepatic encephalopathy. Clin. Gastroenterol. Hepatol. 2015, 13, 1828–1835. [Google Scholar] [CrossRef] [PubMed]
- Bristow, T.; Jih, C.S.; Slabich, A.; Gunn, J. Standardization and adult norms for the sequential subtracting tasks of serial 3’s and 7’s. Appl. Neuropsychol. Adult 2016, 23, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences; Academic Press: Cambridge, MA, USA, 2013. [Google Scholar]
- Chen, J.C.; Schwartz, J. Neurobehavioral effects of ambient air pollution on cognitive performance in US adults. Neurotoxicology 2009, 30, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Knaepen, K.; Goekint, M.; Heyman, E.M.; Meeusen, R. Neuroplasticity—Exercise-induced response of peripheral brain-derived neurotrophic factor: A systematic review of experimental studies in human subjects. Sports Med. 2010, 40, 765–801. [Google Scholar] [CrossRef]
- Silveira, A.C.; Hasegawa, J.S.; Cruz, R.; Matsuda, M.; Marquezini, M.V.; Lima-Silva, A.E.; Giles, L.V.; Saldiva, P.; Koehle, M.S.; Bertuzzi, R. Effects of air pollution exposure on inflammatory and endurance performance in recreationally trained cyclists adapted to traffic-related air pollution. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2022, 322, R562–R570. [Google Scholar] [CrossRef]
- Pedersen, B.K.; Febbraio, M.A. Muscle as an endocrine organ: Focus on muscle-derived interleukin-6. Physiol. Rev. 2008, 88, 1379–1406. [Google Scholar] [CrossRef]
- Steensberg, A.; Fischer, C.P.; Keller, C.; Møller, K.; Pedersen, B.K. IL-6 enhances plasma IL-1ra, IL-10, and cortisol in humans. Am. J. Physiol.-Endocrinol. Metab. 2003, 285, E433–E437. [Google Scholar] [CrossRef]
- Ullum, H.E.; Haahr, P.M.; Diamant, M.A.; Palmo, J.; Halkjaer-Kristensen, J.; Pedersen, B.K. Bicycle exercise enhances plasma IL-6 but does not change IL-1 alpha, IL-1 beta, IL-6, or TNF-alpha pre-mRNA in BMNC. J. Appl. Physiol. 1994, 77, 93–97. [Google Scholar] [CrossRef] [PubMed]
- González-Guevara, E.; Martínez-Lazcano, J.C.; Custodio, V.; Hernández-Cerón, M.; Rubio, C.; Paz, C. Exposure to ozone induces a systemic inflammatory response: Possible source of the neurological alterations induced by this gas. Inhal. Toxicol. 2014, 26, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A. Overview of the IL-1 family in innate inflammation and acquired immunity. Immunol. Rev. 2018, 281, 8–27. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Liu, Y.M.; Wu, W.D.; Jiang, Y.; Zhuo, L.B. Combined exposure of heat stress and ozone enhanced cognitive impairment via neuroinflammation and blood brain barrier disruption in male rats. Sci. Total Environ. 2023, 857, 159599. [Google Scholar] [CrossRef] [PubMed]
- Rivas-Arancibia, S.; Guevara-Guzmán, R.; López-Vidal, Y.; Rodríguez-Martínez, E.; Zanardo-Gomes, M.; Angoa-Pérez, M.; Raisman-Vozari, R. Oxidative stress caused by ozone exposure induces loss of brain repair in the hippocampus of adult rats. Toxicol. Sci. 2010, 113, 187–197. [Google Scholar] [CrossRef]
- Bowtell, J.L.; Aboo-Bakkar, Z.; Conway, M.E.; Adlam, A.L.; Fulford, J. Enhanced task-related brain activation and resting perfusion in healthy older adults after chronic blueberry supplementation. Appl. Physiol. Nutr. Metab. 2017, 42, 773–779. [Google Scholar] [CrossRef]
- Fang, J.L.; Luo, Y.; Jin, S.H.; Yuan, K.; Guo, Y. Ameliorative effect of anthocyanin on depression mice by increasing monoamine neurotransmitter and up-regulating BDNF expression. J. Funct. Foods 2020, 66, 103757. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| PL | POLY | |||||
|---|---|---|---|---|---|---|
| Pre | Post 1 | Post 2 | Pre | Post 1 | Post 2 | |
| Stroop speed (s) | 13.63 ± 3.07 | 13.18 ± 3.27 | 13.48 ± 3.26 | 14.26 ± 3.65 | 13.10 ± 3.02 | 13.58 ± 3.17 |
| Stroop time (s) | 27.26 ± 6.14 | 26.37 ± 6.53 | 26.96 ± 6.52 | 28.55 ± 7.33 | 26.77 ± 7.13 | 27.66 ± 7.02 |
| Stroop errors | 0.20 ± 0.63 | 0.10 ± 0.32 | 0.30 ± 0.48 | 0 ± 0 | 0.20 ± 6.32 | 0.20 ± 0.42 |
| PL | POLY | |||
|---|---|---|---|---|
| Pre | Post | Pre | Post | |
| Serial 3 s | 13.80 ± 3.43 | 14.20 ± 2.20 | 14.20 ± 2.62 | 15.00 ± 2.31 |
| Serial 7 s | 6.40 ± 3.78 | 7.70 ± 4.73 | 8.10 ± 3.84 | 9.10 ± 3.70 |
| Serial 3 errors | 0.40 ± 0.84 | 0.20 ± 0.42 | 0.40 ± 0.84 | 0.0 ± 0.0 * |
| Serial 7 errors | 0.80 ± 0.79 | 0.90 ± 1.01 | 0.50 ± 0.71 | 1.00 ± 1.25 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morton, L.; Paton, C.; Braakhuis, A. The Effects of Polyphenol Supplementation on BDNF, Cytokines and Cognition in Trained Male Cyclists following Acute Ozone Exposure during High-Intensity Cycling. Nutrients 2024, 16, 233. https://doi.org/10.3390/nu16020233
Morton L, Paton C, Braakhuis A. The Effects of Polyphenol Supplementation on BDNF, Cytokines and Cognition in Trained Male Cyclists following Acute Ozone Exposure during High-Intensity Cycling. Nutrients. 2024; 16(2):233. https://doi.org/10.3390/nu16020233
Chicago/Turabian StyleMorton, Lillian, Carl Paton, and Andrea Braakhuis. 2024. "The Effects of Polyphenol Supplementation on BDNF, Cytokines and Cognition in Trained Male Cyclists following Acute Ozone Exposure during High-Intensity Cycling" Nutrients 16, no. 2: 233. https://doi.org/10.3390/nu16020233
APA StyleMorton, L., Paton, C., & Braakhuis, A. (2024). The Effects of Polyphenol Supplementation on BDNF, Cytokines and Cognition in Trained Male Cyclists following Acute Ozone Exposure during High-Intensity Cycling. Nutrients, 16(2), 233. https://doi.org/10.3390/nu16020233