
Activation of the Gut–Brain Interaction by Urolithin A and Its Molecular Basis


Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Reagent
2.2. Mitochondria
2.3. Quantitative Reverse Transcriptase–Polymerase Chain Reaction (RT-qPCR)
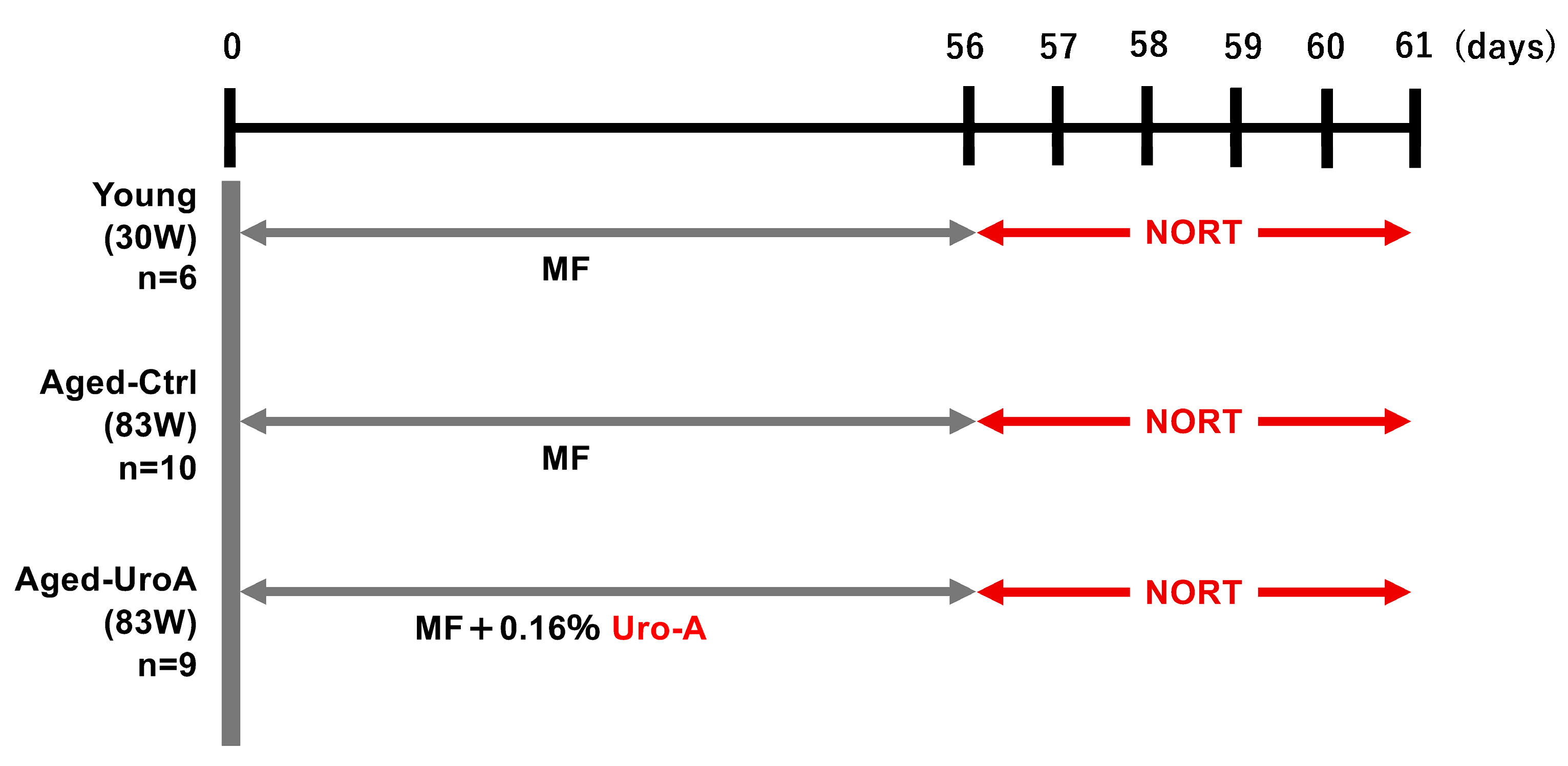
2.4. Animals
2.5. Novel Object Recognition Test (NORT)
2.6. Fluorescent Immunocytochemistry
2.7. RNA Sequencing (RNAseq)
2.8. Exosome Isolation
2.9. miRNA Microarray Assay
2.10. Statistical Analysis
3. Results
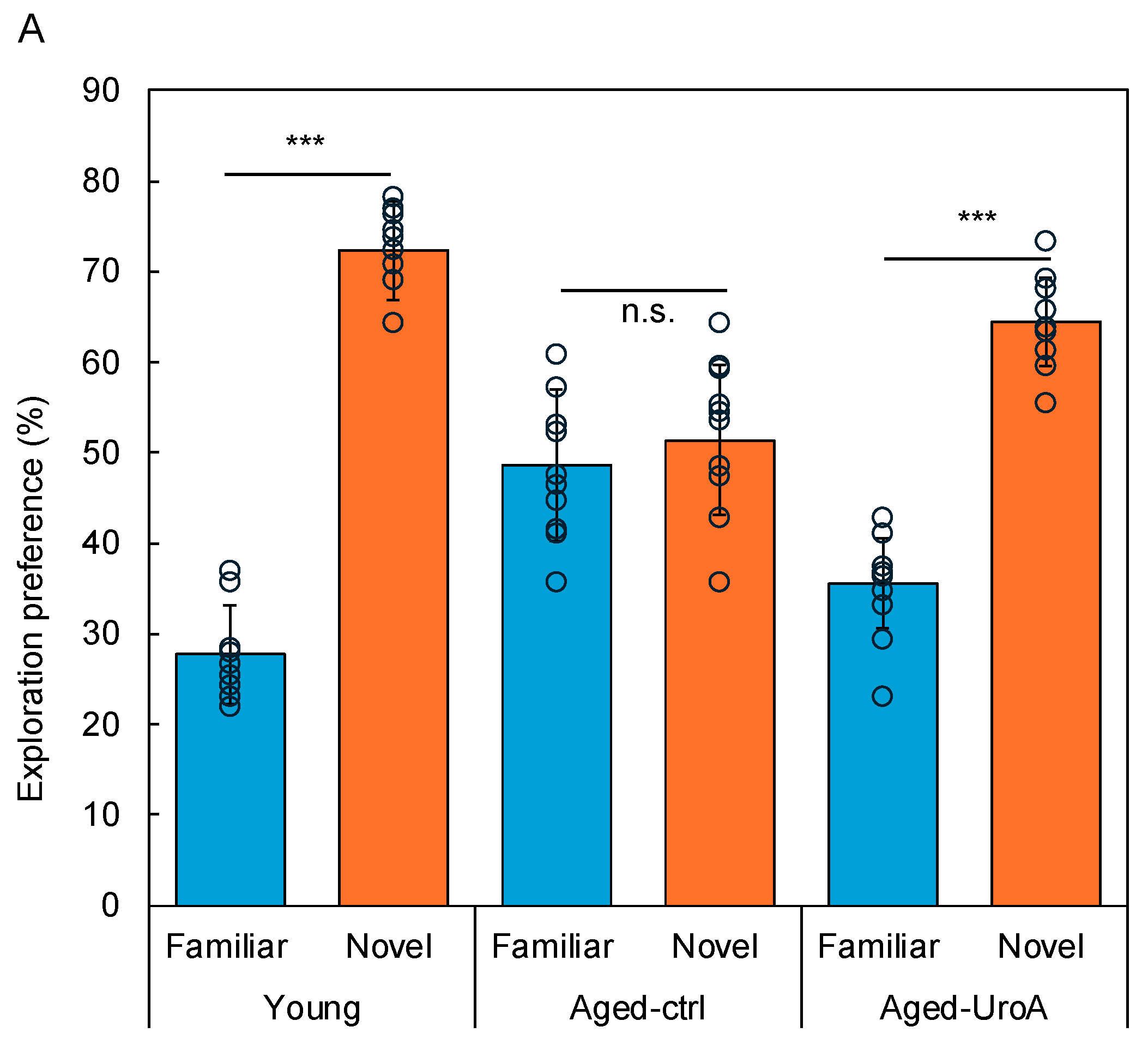
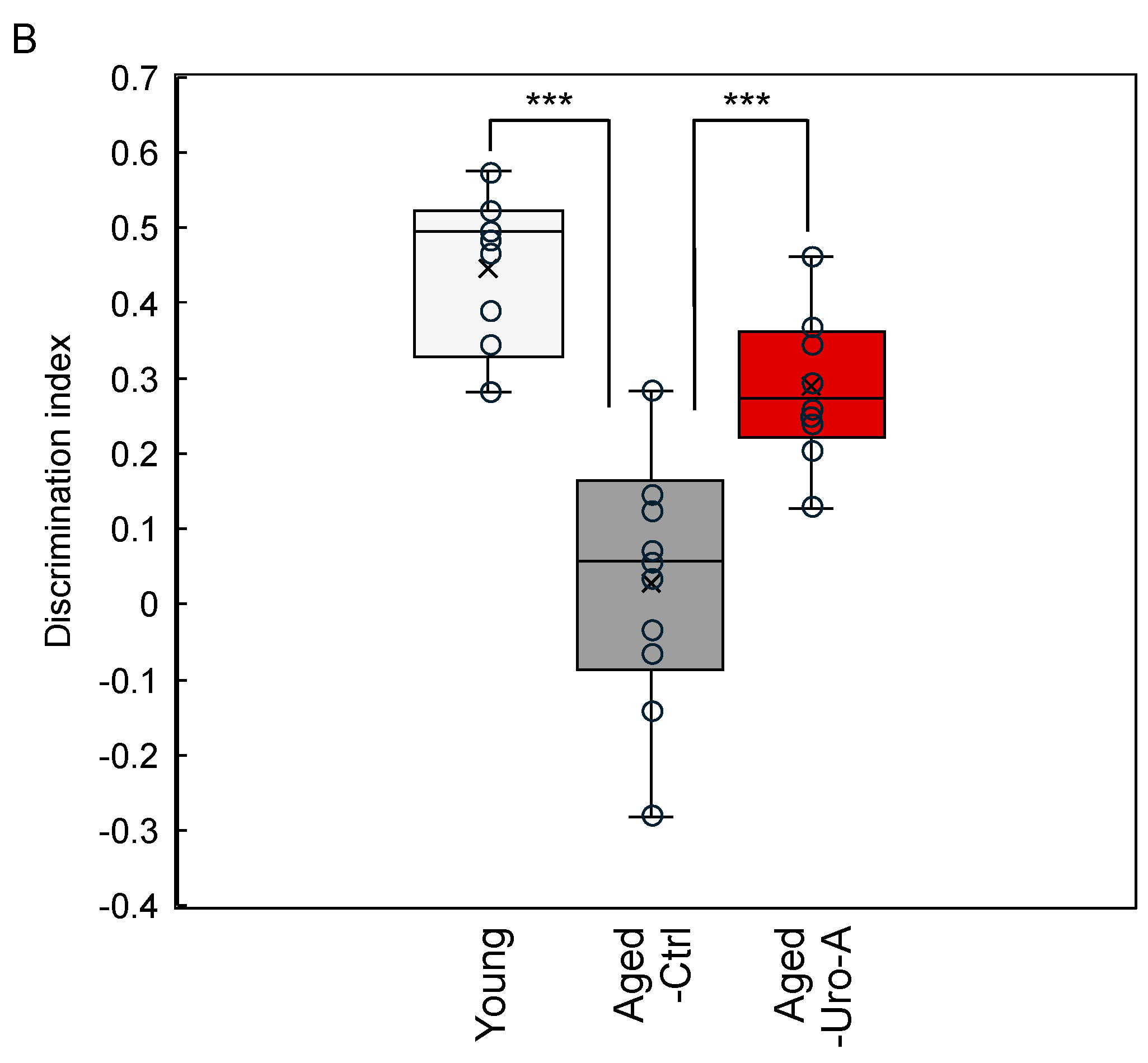
3.1. Effects of Uro-A on Age-Related Memory Impairment in Aged Mice
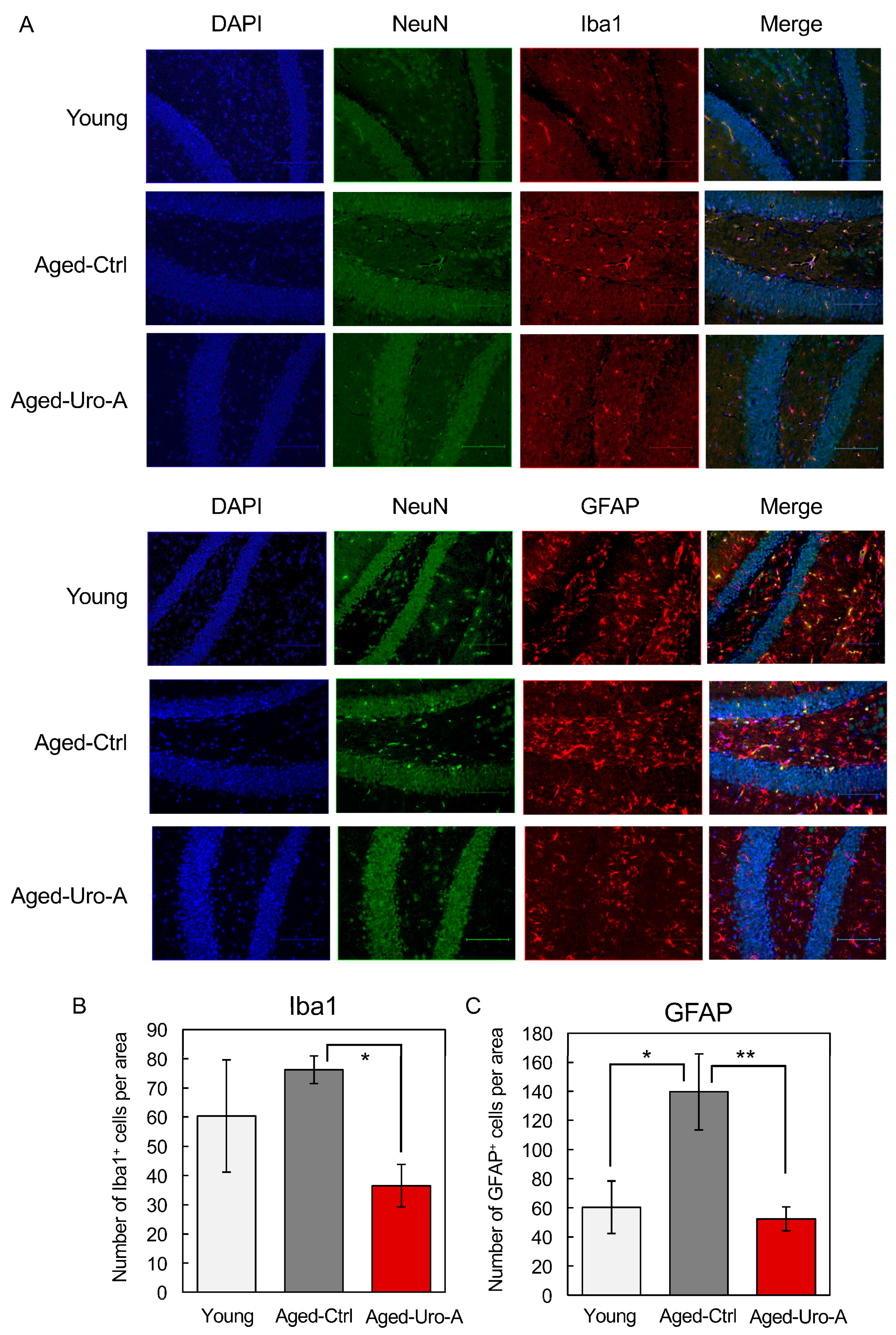
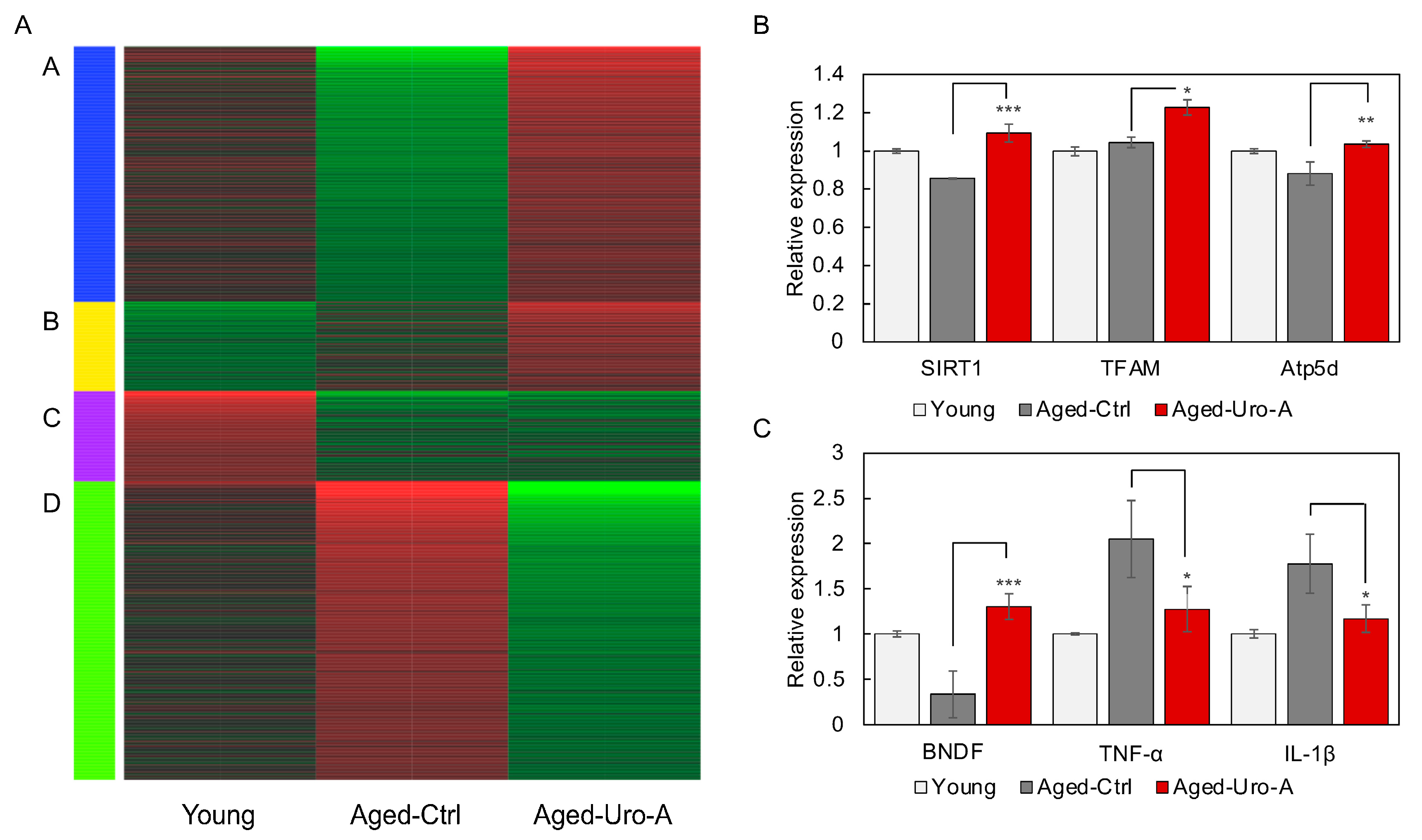
3.2. Effects of Uro-A on Age-Related Inflammation in Aged Mice
3.3. Effects of Uro-A on Gene Expression in the Hippocampus of Aged Mice
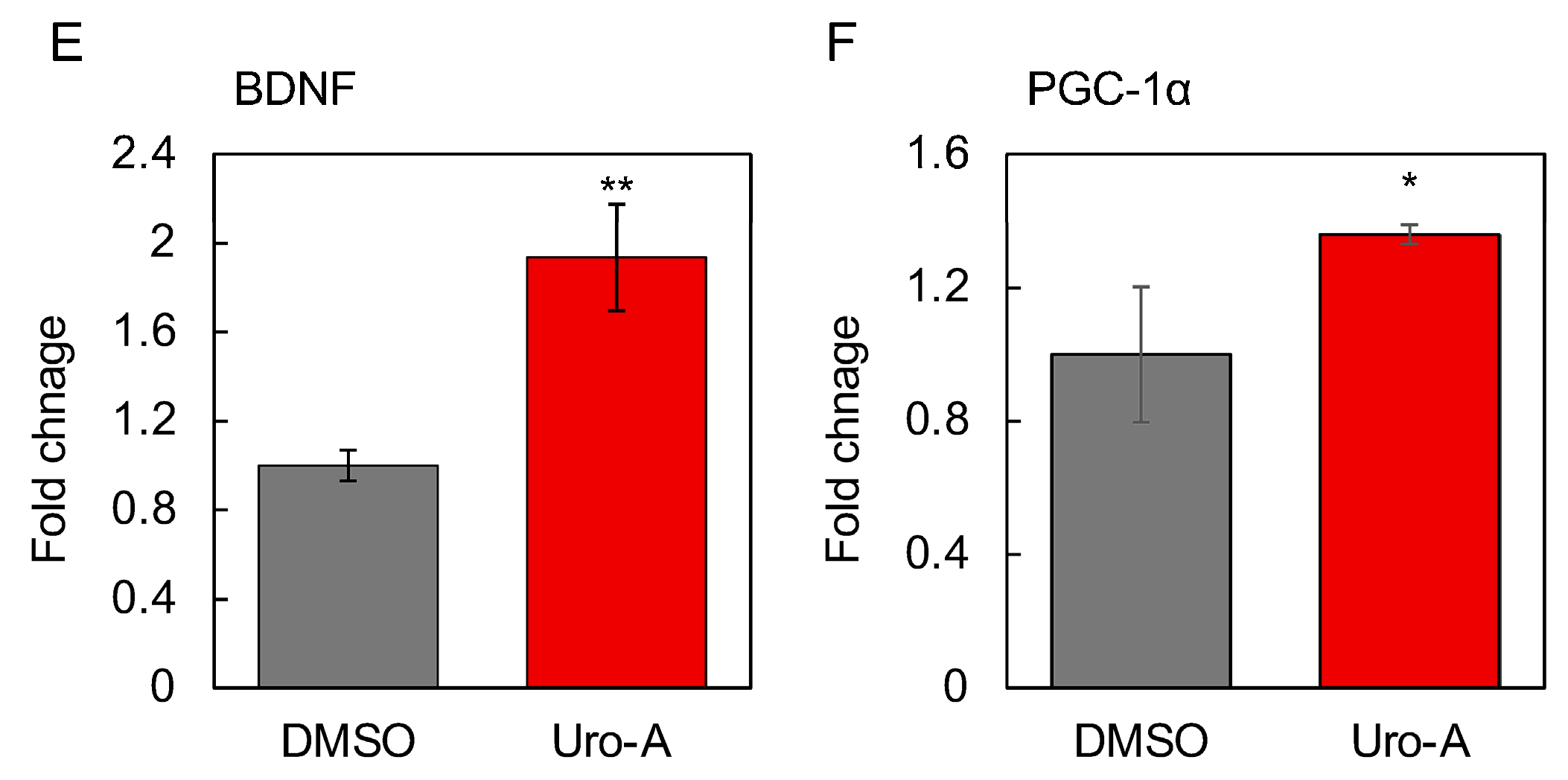
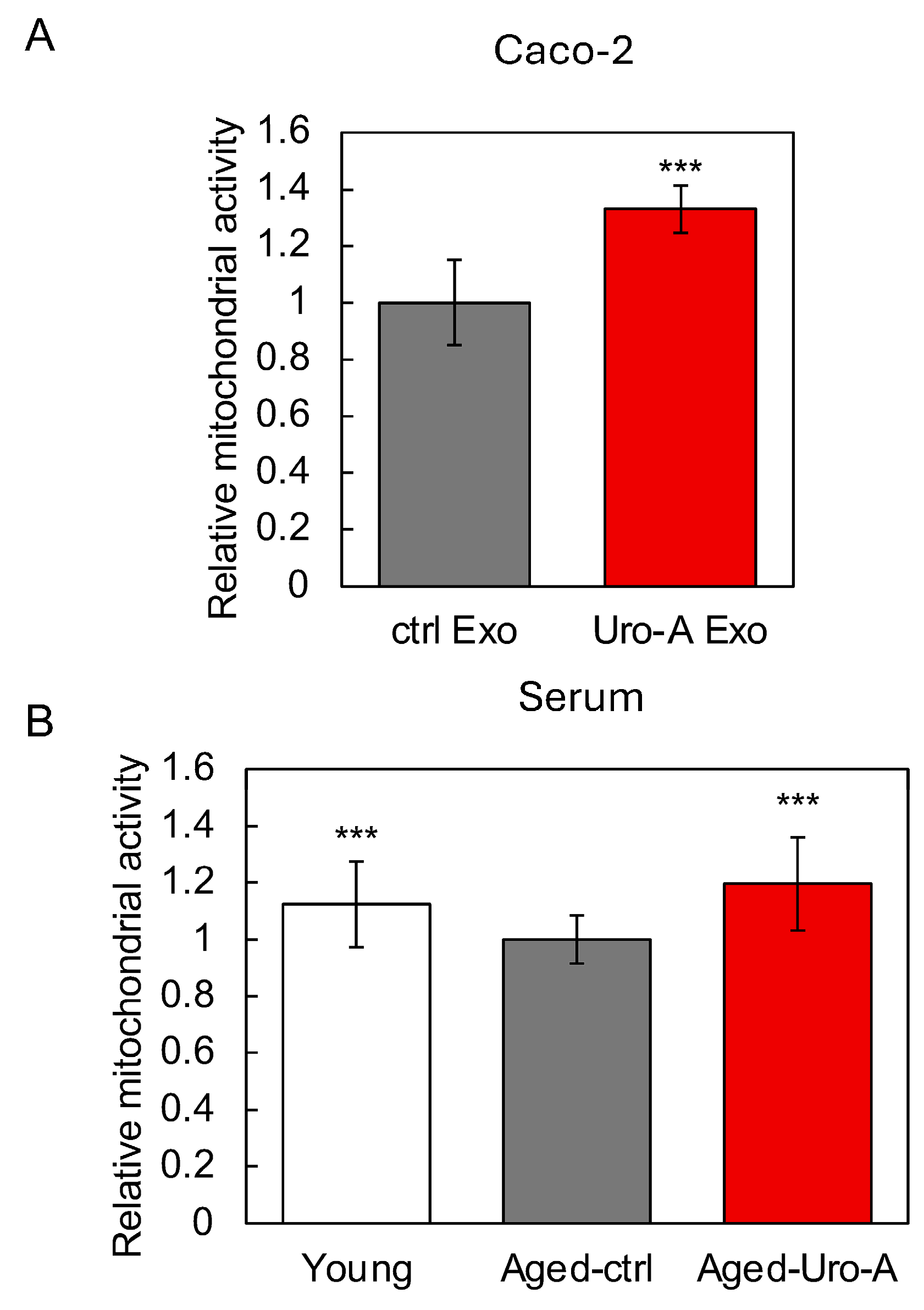
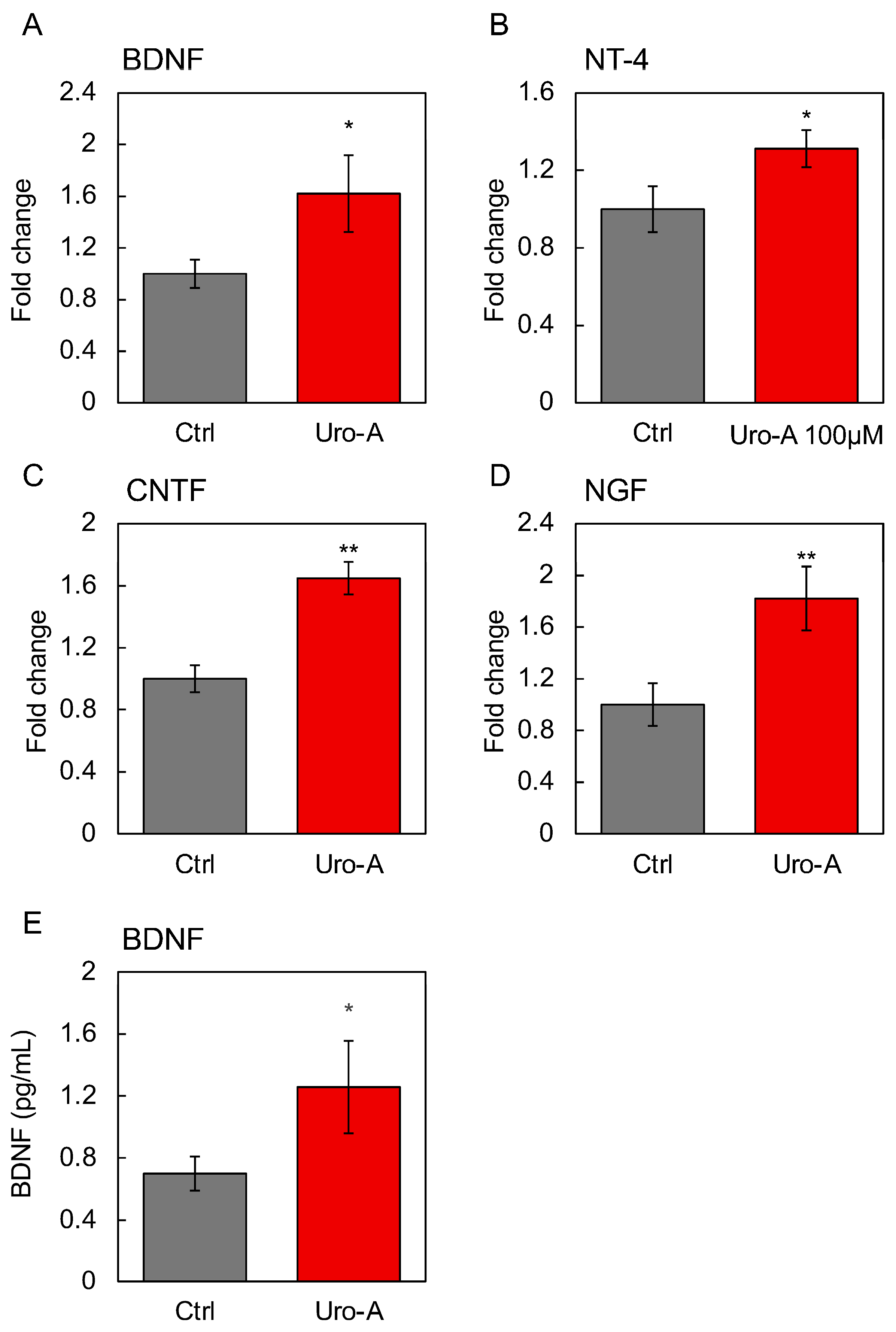
3.4. Supernatants from Caco-2 Cells Treated with Uro-A-Activated SH-SY5Y Cells
3.5. Functional Evaluation of Uro-A-Induced Exosomes
3.6. Involvement of Secreted Factors Other than Exosomes in the Functionality of Uro-A
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- D’Amico, D.; Andreux, P.A.; Valdes, P.; Singh, A.; Rinsch, C.; Auwerx, J. Impact of the Natural Compound Urolithin A on Health, Disease, and Aging. Trends Mol. Med. 2021, 27, 687–699. [Google Scholar] [CrossRef]
- Gong, Z.; Huang, J.; Xu, B.; Ou, Z.; Zhang, L.; Lin, X.; Ye, X.; Kong, X.; Long, D.; Sun, X.; et al. Urolithin A attenuates memory impairment and neuroinflammation in APP/PS1 mice. J. Neuroinflamm. 2019, 16, 62. [Google Scholar] [CrossRef]
- Fang, E.F.; Hou, Y.; Palikaras, K.; Adriaanse, B.A.; Kerr, J.S.; Yang, B.; Lautrup, S.; Hasan-Olive, M.M.; Caponio, D.; Dan, X.; et al. Mitophagy inhibits amyloid-beta and tau pathology and reverses cognitive deficits in models of Alzheimer’s disease. Nat. Neurosci. 2019, 22, 401–412. [Google Scholar] [CrossRef]
- Chong, Z.; Matsuo, H.; Onoue, S.; Yamamoto, H.; Ito, H.; Katakura, Y. Identification of polyphenols that repair the ultraviolet-B-induced DNA damage via SIRT1-dependent XPC/XPA activation. J. Funct. Foods 2019, 54, 119–127. [Google Scholar] [CrossRef]
- Ishibashi, A.; Udono, M.; Sato, M.; Katakura, Y. Molecular Mechanisms for the Carnosine-Induced Activation of Muscle-Brain Interaction. Nutrients 2023, 15, 1479. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Shoji, H.; Miyakawa, T. Age-related behavioral changes from young to old age in male mice of a C57BL/6J strain maintained under a genetic stability program. Neuropsychopharmacol. Rep. 2019, 39, 100–118. [Google Scholar] [CrossRef]
- Shoji, H.; Takao, K.; Hattori, S.; Miyakawa, T. Age-related changes in behavior in C57BL/6J mice from young adulthood to middle age. Mol. Brain 2016, 9, 11. [Google Scholar] [CrossRef]
- Tanimizu, T.; Kono, K.; Kida, S. Brain networks activated to form object recognition memory. Brain Res. Bull. 2018, 141, 27–34. [Google Scholar] [CrossRef]
- Kubo, C.; Ogawa, M.; Uehara, N.; Katakura, Y. Fisetin Promotes Hair Growth by Augmenting TERT Expression. Front. Cell Dev. Biol. 2020, 8, 566617. [Google Scholar] [CrossRef]
- Ogawa, M.; Udono, M.; Teruya, K.; Uehara, N.; Katakura, Y. Exosomes Derived from Fisetin-Treated Keratinocytes Mediate Hair Growth Promotion. Nutrients 2021, 13, 2087. [Google Scholar] [CrossRef]
- Ge, S.X.; Son, E.W.; Yao, R. iDEP: An integrated web application for differential expression and pathway analysis of RNA-Seq data. BMC Bioinform. 2018, 19, 534. [Google Scholar] [CrossRef]
- Sugihara, Y.; Onoue, S.; Tashiro, K.; Sato, M.; Hasegawa, T.; Katakura, Y. Carnosine induces intestinal cells to secrete exosomes that activate neuronal cells. PLoS ONE 2019, 14, e0217394. [Google Scholar] [CrossRef]
- Bolstad, B.M.; Irizarry, R.A.; Astrand, M.; Speed, T.P. A comparison of normalization methods for high density oligonucleotide array data based on variance and bias. Bioinformatics 2003, 19, 185–193. [Google Scholar] [CrossRef]
- Inotsuka, R.; Udono, M.; Yamatsu, A.; Kim, M.; Katakura, Y. Exosome-Mediated Activation of Neuronal Cells Triggered by gamma-Aminobutyric Acid (GABA). Nutrients 2021, 13, 2544. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef]
- Zhu, H.; Liu, J.; Zhou, J.; Jin, Y.; Zhao, Q.; Jiang, X.; Gao, H. Notopterygium incisum roots extract (NRE) alleviates neuroinflammation pathology in Alzheimer’s disease through TLR4-NF-κB pathway. J. Ethnopharmacol. 2024, 335, 118651. [Google Scholar] [CrossRef]
- Sales, I.S.L.; de Souza, A.G.; Chaves Filho, A.J.M.; Sampaio, T.L.; da Silva, D.M.A.; Valentim, J.T.; Chaves, R.C.; Soares, M.V.R.; Costa Junior, D.C.; Barbosa Filho, J.M.; et al. Antidepressant-like effect of riparin I and riparin II against CUMS-induced neuroinflammation via astrocytes and microglia modulation in mice. Behav. Pharmacol. 2024, 35, 314–326. [Google Scholar] [CrossRef]
- Izadi, S.; Abdolrezaei, M.; Soukhaklari, R.; Moosavi, M. Memory impairment induced by aluminum nanoparticles is associated with hippocampal IL-1 and IBA-1 upregulation in mice. Neurol. Res. 2024, 46, 284–290. [Google Scholar] [CrossRef]
- Lituma, P.J.; Woo, E.; O’Hara, B.F.; Castillo, P.E.; Sibinga, N.E.S.; Nandi, S. Altered synaptic connectivity and brain function in mice lacking microglial adapter protein Iba1. Proc. Natl. Acad. Sci. USA 2021, 118, e2115539118. [Google Scholar] [CrossRef]
- Wang, L.; Lu, Y.; Liu, J.M.; Wang, S.Y.; Fei, Z.P.; Zhang, K.W.; Zhang, D.F.; Jin, X. Gegen Qinlian tablets delay Alzheimer ‘s disease progression via inhibiting glial neuroinflammation and remodeling gut microbiota homeostasis. Phytomedicine 2024, 128, 155394. [Google Scholar] [CrossRef]
- Shen, Y.; Ye, B.; Chen, P.; Wang, Q.; Fan, C.; Shu, Y.; Xiang, M. Cognitive Decline, Dementia, Alzheimer’s Disease and Presbycusis: Examination of the Possible Molecular Mechanism. Front. Neurosci. 2018, 12, 394. [Google Scholar] [CrossRef]
- Chaturvedi, R.K.; Flint Beal, M. Mitochondrial diseases of the brain. Free Radic. Biol. Med. 2013, 63, 1–29. [Google Scholar] [CrossRef]
- Patro, S.; Ratna, S.; Yamamoto, H.A.; Ebenezer, A.T.; Ferguson, D.S.; Kaur, A.; McIntyre, B.C.; Snow, R.; Solesio, M.E. ATP Synthase and Mitochondrial Bioenergetics Dysfunction in Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 22, 11185. [Google Scholar] [CrossRef]
- Wang, Y.H.; Mondal, G.; Khan, W.; Gurley, B.J.; Yates, C.R. Development of a liquid chromatography-tandem mass spectrometry (LC-MS/MS) method for characterizing pomegranate extract pharmacokinetics in humans. J. Pharm. Biomed. Anal. 2023, 233, 115477. [Google Scholar] [CrossRef]
- Tang, B.L. Sirt1 and the Mitochondria. Mol. Cells 2016, 39, 87–95. [Google Scholar] [CrossRef]
- Zhou, L.; Pinho, R.; Gu, Y.; Radak, Z. The Role of SIRT3 in Exercise and Aging. Cells 2022, 11, 2596. [Google Scholar] [CrossRef]
- Imai, S.; Guarente, L. NAD+ and sirtuins in aging and disease. Trends Cell Biol. 2014, 24, 464–471. [Google Scholar] [CrossRef]
- Gao, L.; Zhang, Y.; Sterling, K.; Song, W. Brain-derived neurotrophic factor in Alzheimer’s disease and its pharmaceutical potential. Transl. Neurodegener. 2022, 11, 4. [Google Scholar] [CrossRef]
- Shaito, A.; Al-Mansoob, M.; Ahmad, S.M.S.; Haider, M.Z.; Eid, A.H.; Posadino, A.M.; Pintus, G.; Giordo, R. Resveratrol-Mediated Regulation of Mitochondria Biogenesis-associated Pathways in Neurodegenerative Diseases: Molecular Insights and Potential Therapeutic Applications. Curr. Neuropharmacol. 2023, 21, 1184–1201. [Google Scholar] [CrossRef] [PubMed]
- Inotsuka, R.; Uchimura, K.; Yamatsu, A.; Kim, M.; Katakura, Y. gamma-Aminobutyric acid (GABA) activates neuronal cells by inducing the secretion of exosomes from intestinal cells. Food Funct. 2020, 11, 9285–9290. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Song, G.; Zhu, H.; Qian, H.; Pan, X.; Song, X.; Xie, Y.; Liu, C. Pharmacological Effects of Urolithin A and Its Role in Muscle Health and Performance: Current Knowledge and Prospects. Nutrients 2023, 15, 4441. [Google Scholar] [CrossRef]
- Wang, I.F.; Ho, P.C.; Tsai, K.J. MicroRNAs in Learning and Memory and Their Impact on Alzheimer’s Disease. Biomedicines 2022, 10, 1856. [Google Scholar] [CrossRef]
- Su, L.; Li, R.; Zhang, Z.; Liu, J.; Du, J.; Wei, H. Identification of altered exosomal microRNAs and mRNAs in Alzheimer’s disease. Ageing Res. Rev. 2022, 73, 101497. [Google Scholar] [CrossRef] [PubMed]
- Swerdlow, R.H. Mitochondria and Mitochondrial Cascades in Alzheimer’s Disease. J. Alzheimer’s Dis. 2018, 62, 1403–1416. [Google Scholar] [CrossRef]
- Kadooka, K.; Fujii, K.; Matsumoto, T.; Sato, M.; Morimatsu, F.; Tashiro, K.; Kuhara, S.; Katakura, Y. Mechanisms and consequences of carnosine-induced activation of intestinal epithelial cells. J. Funct. Foods 2015, 13, 32–37. [Google Scholar] [CrossRef]
- Matin, S.; Dadkhah, M. BDNF/CREB signaling pathway contribution in depression pathogenesis: A survey on the non-pharmacological therapeutic opportunities for gut microbiota dysbiosis. Brain Res. Bull. 2024, 207, 110882. [Google Scholar] [CrossRef]
- Lin, P.H.; Kuo, L.T.; Luh, H.T. The Roles of Neurotrophins in Traumatic Brain Injury. Life 2021, 12, 26. [Google Scholar] [CrossRef]
- Bolognin, S.; Buffelli, M.; Puolivali, J.; Iqbal, K. Rescue of cognitive-aging by administration of a neurogenic and/or neurotrophic compound. Neurobiol. Aging 2014, 35, 2134–2146. [Google Scholar] [CrossRef]
- Amadoro, G.; Latina, V.; Balzamino, B.O.; Squitti, R.; Varano, M.; Calissano, P.; Micera, A. Nerve Growth Factor-Based Therapy in Alzheimer’s Disease and Age-Related Macular Degeneration. Front. Neurosci. 2021, 15, 735928. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Origin | Caco-2 | |||||
|---|---|---|---|---|---|---|---|
| miR | 4730 | 6126 | 663a | 4497 | 4745-5p | 3663-3p | |
| Memory | Axon guidance | * | *** | ** | * | ** | *** |
| Long-term potentiation | * | ||||||
| Neurotrophin signaling pathway | * | *** | |||||
| Neurological Function | GABAergic synapse | * | * | ||||
| Long-term depression | |||||||
| Dopaminergic synapse | ** | ||||||
| Oxytocin signaling pathway | *** | * | |||||
| Longevity Signal | Longevity regulating pathway | *** | * | * | |||
| FoxO signaling pathway | *** | ||||||
| Calcium signaling pathway | ** | * | * | ** | |||
| Category | Origin | Mouse Serum | |||||
|---|---|---|---|---|---|---|---|
| miR | 29a-5p | 449c-3p | 6240 | 5100 | 3547-5p | 2861 | |
| Memory | Axon guidance | *** | *** | *** | *** | *** | |
| Long-term potentiation | ** | ** | *** | *** | * | ||
| Neurotrophin signaling pathway | ** | ** | *** | *** | |||
| Neurological Function | GABAergic synapse | ** | *** | *** | * | ||
| Long-term depression | * | ** | ** | ** | ** | ||
| Dopaminergic synapse | *** | *** | *** | *** | |||
| Oxytocin signaling pathway | ** | * | ** | *** | *** | ||
| Longevity Signal | Longevity regulating pathway | ** | *** | *** | * | ||
| FoxO signaling pathway | *** | ** | ** | *** | |||
| Calcium signaling pathway | *** | ** | ** | *** | *** | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kubota, D.; Sato, M.; Udono, M.; Kohara, A.; Kudoh, M.; Ukawa, Y.; Teruya, K.; Katakura, Y. Activation of the Gut–Brain Interaction by Urolithin A and Its Molecular Basis. Nutrients 2024, 16, 3369. https://doi.org/10.3390/nu16193369
Kubota D, Sato M, Udono M, Kohara A, Kudoh M, Ukawa Y, Teruya K, Katakura Y. Activation of the Gut–Brain Interaction by Urolithin A and Its Molecular Basis. Nutrients. 2024; 16(19):3369. https://doi.org/10.3390/nu16193369
Chicago/Turabian StyleKubota, Daiki, Momoka Sato, Miyako Udono, Akiko Kohara, Masatake Kudoh, Yuichi Ukawa, Kiichiro Teruya, and Yoshinori Katakura. 2024. "Activation of the Gut–Brain Interaction by Urolithin A and Its Molecular Basis" Nutrients 16, no. 19: 3369. https://doi.org/10.3390/nu16193369
APA StyleKubota, D., Sato, M., Udono, M., Kohara, A., Kudoh, M., Ukawa, Y., Teruya, K., & Katakura, Y. (2024). Activation of the Gut–Brain Interaction by Urolithin A and Its Molecular Basis. Nutrients, 16(19), 3369. https://doi.org/10.3390/nu16193369