
Heat-Processed Soybean Germ Extract and Lactobacillus gasseri NK109 Supplementation Reduce LPS-Induced Cognitive Impairment and Colitis in Mice


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Culture of NK109
2.3. Preparation of SE and Its Supplement
2.4. Determination of the Soayasopnin Bb Content in SE and hSE
2.5. Cultures of BV-2 Cells
2.6. Animals
2.7. Cognitive Function-Impaired Mouse Preparation
2.8. Behavioral Tasks
2.9. Immunoblotting and ELISA
2.10. Immunofluorescence Staining
2.11. Statistical Analysis
3. Results
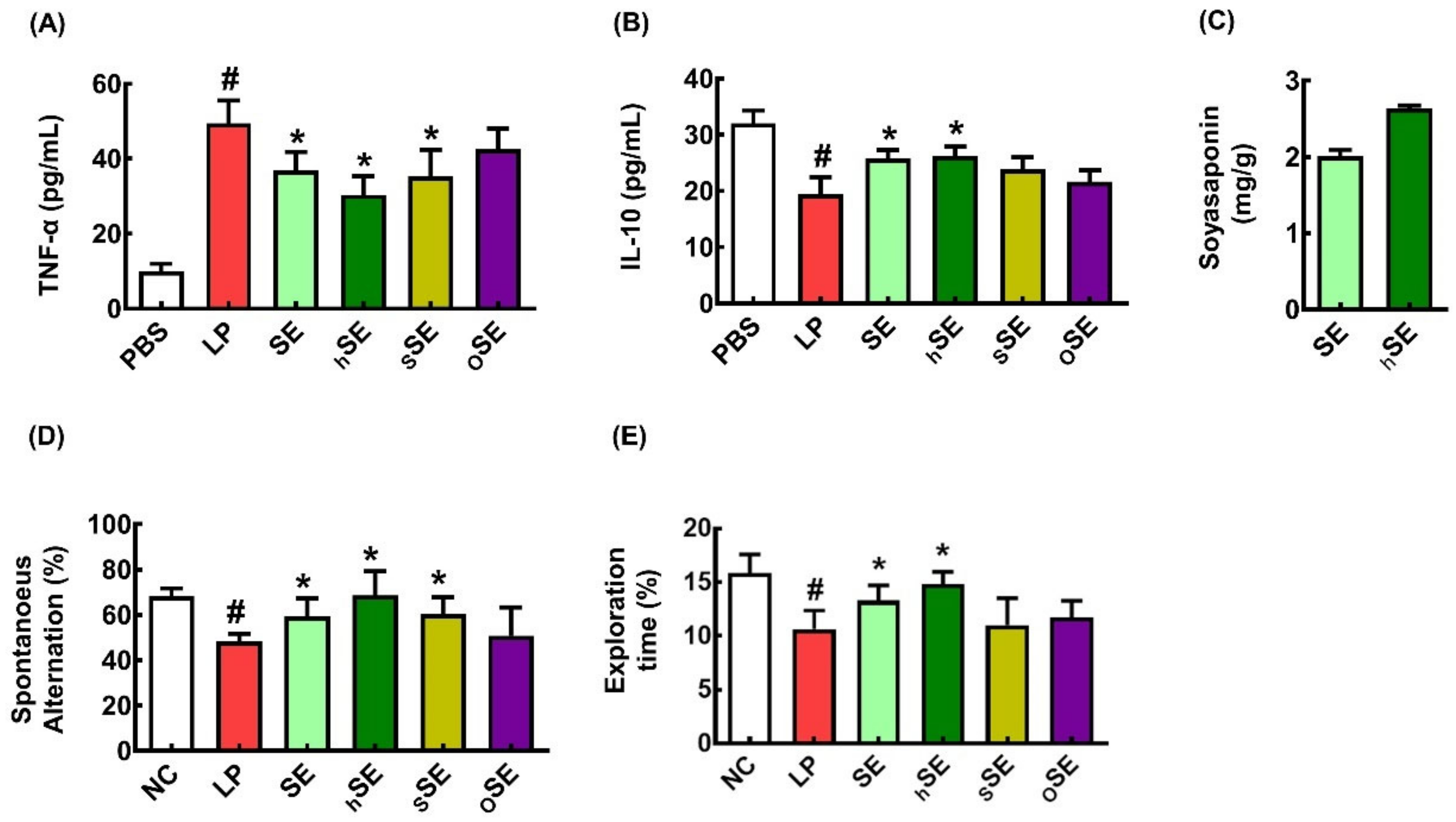
3.1. SEs Decreased LPS-Induced TNF-α Expression in BV2 Cells
3.2. SE and hSE Alleviated LCI in Mice
3.3. NK109 Decreased LPS-Induced TNF-a Expression in BV2 Cells
3.4. NK109 Alleviated LCI in Mice
3.5. The Combined Effects of NK109 with NK109 (SN) and hSE with NK109 (hSN) on LCI and Colitis in Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tarawneh, R.; Holtzman, D.M. The clinical problem of symptomatic Alzheimer disease and mild cognitive impairment. Cold Spring Harb. Perspect. Med. 2012, 2, a006148. [Google Scholar] [CrossRef]
- Cao, Q.; Tan, C.C.; Xu, W.; Hu, H.; Cao, X.P.; Dong, Q.; Tan, L.; Yu, J.T. The Prevalence of Dementia: A Systematic Review and Meta-Analysis. J. Alzheimer’s Dis. 2020, 73, 1157–1166. [Google Scholar] [CrossRef]
- Busche, M.A.; Hyman, B.T. Synergy between amyloid-β and tau in Alzheimer’s disease. Nat. Neurosci. 2020, 23, 1183–1193. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; O’Riordan, K.J.; Sandhu, K.; Peterson, V.; Dinan, T.G. The gut microbiome in neurological disorders. Lancet Neurol. 2020, 19, 179–194. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.E.; Lim, S.M.; Jeong, J.J.; Jang, H.M.; Lee, H.J.; Han, M.J.; Kim, D.H. Gastrointestinal inflammation by gut microbiota disturbance induces memory impairment in mice. Mucosal Immunol. 2018, 11, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.E.; Kim, J.K.; Kim, D.H. Orally Administered Antibiotics Vancomycin and Ampicillin Cause Cognitive Impairment With Gut Dysbiosis in Mice With Transient Global Forebrain Ischemia. Front. Microbiol. 2020, 11, 564271. [Google Scholar] [CrossRef]
- Gubert, C.; Kong, G.; Renoir, T.; Hannan, A.J. Exercise, diet and stress as modulators of gut microbiota: Implications for neurodegenerative diseases. Neurobiol. Dis. 2020, 134, 104621. [Google Scholar] [CrossRef]
- Sun, Y.; Baptista, L.C.; Roberts, L.M.; Jumbo-Lucioni, P.; McMahon, L.L.; Buford, T.W.; Carter, C.S. The Gut Microbiome as a Therapeutic Target for Cognitive Impairment. J. Gerontol. A Biol. Sci. Med. Sci. 2020, 75, 1242–1250. [Google Scholar] [CrossRef]
- Yun, S.W.; Kim, J.K.; Han, M.J.; Kim, D.H. Lacticaseibacillus paracasei NK112 mitigates Escherichia coli-induced depression and cognitive impairment in mice by regulating IL-6 expression and gut microbiota. Benef. Microbes 2021, 12, 541–551. [Google Scholar] [CrossRef]
- Yun, S.W.; Kim, J.K.; Lee, K.E.; Oh, Y.J.; Choi, H.J.; Han, M.J.; Kim, D.H. A Probiotic Lactobacillus gasseri Alleviates Escherichia coli-Induced Cognitive Impairment and Depression in Mice by Regulating IL-1β Expression and Gut Microbiota. Nutrients 2020, 12, 3441. [Google Scholar] [CrossRef]
- Yun, S.W.; Park, H.S.; Shin, Y.J.; Ma, X.; Han, M.J.; Kim, D.H. Lactobacillus gasseri NK109 and Its Supplement Alleviate Cognitive Impairment in Mice by Modulating NF-κB Activation, BDNF Expression, and Gut Microbiota Composition. Nutrients 2023, 15, 790. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Lee, K.E.; Lee, S.A.; Jang, H.M.; Kim, D.H. Interplay Between Human Gut Bacteria Escherichia coli and Lactobacillus mucosae in the Occurrence of Neuropsychiatric Disorders in Mice. Front. Immunol. 2020, 11, 273. [Google Scholar] [CrossRef]
- Lee, H.J.; Lee, K.E.; Kim, J.K.; Kim, D.H. Suppression of gut dysbiosis by Bifidobacterium longum alleviates cognitive decline in 5XFAD transgenic and aged mice. Sci. Rep. 2019, 9, 11814. [Google Scholar] [CrossRef] [PubMed]
- George, K.S.; Muñoz, J.; Akhavan, N.S.; Foley, E.M.; Siebert, S.C.; Tenenbaum, G.; Khalil, D.A.; Chai, S.C.; Arjmandi, B.H. Is soy protein effective in reducing cholesterol and improving bone health? Food Funct. 2020, 11, 544–551. [Google Scholar] [CrossRef] [PubMed]
- Lee, N. Phytoestrogens as bioactive ingredients in functional foods: Canadian regulatory update. J. AOAC Int. 2006, 89, 1135–1137. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.L.; Berhow, M.A.; Kim, J.T.; Chi, H.Y.; Lee, S.J.; Chung, I.M. Evaluation of soyasaponin, isoflavone, protein, lipid, and free sugar accumulation in developing soybean seeds. J. Agric. Food Chem. 2006, 54, 10003–10010. [Google Scholar] [CrossRef] [PubMed]
- Guang, C.; Chen, J.; Sang, S.; Cheng, S. Biological functionality of soyasaponins and soyasapogenols. J. Agric. Food Chem. 2014, 62, 8247–8255. [Google Scholar] [CrossRef]
- Kim, I.S. Current Perspectives on the Beneficial Effects of Soybean Isoflavones and Their Metabolites for Humans. Antioxidants 2021, 10, 1064. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Lim, S.M.; Ko, D.B.; Jeong, J.J.; Hwang, Y.H.; Kim, D.H. Soyasapogenol B and Genistein Attenuate Lipopolysaccharide-Induced Memory Impairment in Mice by the Modulation of NF-κB-Mediated BDNF Expression. J. Agric. Food Chem. 2017, 65, 6877–6885. [Google Scholar] [CrossRef]
- Hong, S.W.; Heo, H.; Yang, J.H.; Han, M.; Kim, D.H.; Kwon, Y.K. Soyasaponin I improved neuroprotection and regeneration in memory deficient model rats. PLoS ONE 2013, 8, e81556. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.W.; Yoo, D.H.; Woo, J.Y.; Jeong, J.J.; Yang, J.H.; Kim, D.H. Soyasaponins Ab and Bb prevent scopolamine-induced memory impairment in mice without the inhibition of acetylcholinesterase. J. Agric. Food Chem. 2014, 62, 2062–2068. [Google Scholar] [CrossRef]
- Lee, H.J.; Hwang, Y.H.; Kim, D.H. Lactobacillus plantarum C29-Fermented Soybean (DW2009) Alleviates Memory Impairment in 5XFAD Transgenic Mice by Regulating Microglia Activation and Gut Microbiota Composition. Mol. Nutr. Food Res. 2018, 62, e1800359, Erratum in Mol. Nutr. Food Res. 2021, 65, e2170024. [Google Scholar] [CrossRef]
- Jang, H.M.; Lee, K.E.; Kim, D.H. The Preventive and Curative Effects of Lactobacillus reuteri NK33 and Bifidobacterium adolescentis NK98 on Immobilization Stress-Induced Anxiety/Depression and Colitis in Mice. Nutrients 2019, 11, 819. [Google Scholar] [CrossRef]
- Percie du Sert, N.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE guidelines 2.0: Updated guidelines for reporting animal research. PLoS Biol. 2020, 18, e3000410. [Google Scholar] [CrossRef]
- Lee, K.E.; Kim, J.K.; Han, S.K.; Lee, D.Y.; Lee, H.J.; Yim, S.V.; Kim, D.H. The extracellular vesicle of gut microbial Paenalcaligenes hominis is a risk factor for vagus nerve-mediated cognitive impairment. Microbiome 2020, 8, 107. [Google Scholar] [CrossRef] [PubMed]
- Sarter, M.; Bodewitz, G.; Stephens, D.N. Attenuation of scopolamine-induced impairment of spontaneous alteration behaviour by antagonist but not inverse agonist and agonist beta-carbolines. Psychopharmacology 1988, 94, 491–495. [Google Scholar] [CrossRef]
- Morais, L.H.; Schreiber, H.L.T.; Mazmanian, S.K. The gut microbiota-brain axis in behaviour and brain disorders. Nat. Rev. Microbiol. 2021, 19, 241–255. [Google Scholar] [CrossRef]
- Fousekis, F.S.; Katsanos, A.H.; Kourtis, G.; Saridi, M.; Albani, E.; Katsanos, K.H.; Christodoulou, D.K. Inflammatory Bowel Disease and Patients With Mental Disorders: What Do We Know? J. Clin. Med. Res. 2021, 13, 466–473. [Google Scholar] [CrossRef]
- Ma, X.; Shin, Y.J.; Jang, H.M.; Joo, M.K.; Yoo, J.W.; Kim, D.H. Lactobacillus rhamnosus and Bifidobacterium longum alleviate colitis and cognitive impairment in mice by regulating IFN-γ to IL-10 and TNF-α to IL-10 expression ratios. Sci. Rep. 2021, 11, 20659. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.A.; Park, Y.J.; Yeo, H.K.; Han, M.J.; Kim, D.H. Soyasaponin I attenuates TNBS-Induced colitis in mice by inhibiting NF-κB pathway. J. Agric. Food Chem. 2010, 58, 10929–10934. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.A.; Park, Y.J.; Joh, E.H.; Kim, D.H. Soyasaponin Ab ameliorates colitis by inhibiting the binding of lipopolysaccharide (LPS) to Toll-like receptor (TLR)4 on macrophages. J. Agric. Food Chem. 2011, 59, 13165–13172. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.M.; Lee, K.E.; Lee, H.J.; Kim, D.H. Immobilization stress-induced Escherichia coli causes anxiety by inducing NF-κB activation through gut microbiota disturbance. Sci. Rep. 2018, 8, 13897. [Google Scholar] [CrossRef]
- Porter, G.A.; O’Connor, J.C. Brain-derived neurotrophic factor and inflammation in depression: Pathogenic partners in crime? World J. Psychiatry 2022, 12, 77–97. [Google Scholar] [CrossRef] [PubMed]
- Figurov, A.; Pozzo-Miller, L.D.; Olafsson, P.; Wang, T.; Lu, B. Regulation of synaptic responses to high-frequency stimulation and LTP by neurotrophins in the hippocampus. Nature 1996, 381, 706–709. [Google Scholar] [CrossRef] [PubMed]
- Akao, T.; Kida, H.; Kanaoka, M.; Hattori, M.; Kobashi, K. Intestinal bacterial hydrolysis is required for the appearance of compound K in rat plasma after oral administration of ginsenoside Rb1 from Panax ginseng. J. Pharm. Pharmacol. 1998, 50, 1155–1160. [Google Scholar] [CrossRef]
- Joh, E.H.; Kim, D.H. A sensitive liquid chromatography-electrospray tandem mass spectrometric method for lancemaside A and its metabolites in plasma and a pharmacokinetic study in mice. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2010, 878, 1875–1880. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Kim, J.K.; Yun, S.W.; Han, M.J.; Kim, D.H. DW2009 Elevates the Efficacy of Donepezil against Cognitive Impairment in Mice. Nutrients 2021, 13, 3273. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, D.; Røn, Ø.; Baardsen, G.; Smedsgaard, J.; Koppe, W.; Frøkiaer, H. Soyasaponins resist extrusion cooking and are not degraded during gut passage in Atlantic salmon (Salmo salar L.). J. Agric. Food Chem. 2006, 54, 6428–6435. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Teng, S.P.; Popovich, D.G. Generation of group B soyasaponins I and III by hydrolysis. J. Agric. Food Chem. 2009, 57, 3620–3625. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yun, S.-W.; Lee, D.-Y.; Park, H.-S.; Kim, D.-H. Heat-Processed Soybean Germ Extract and Lactobacillus gasseri NK109 Supplementation Reduce LPS-Induced Cognitive Impairment and Colitis in Mice. Nutrients 2024, 16, 2736. https://doi.org/10.3390/nu16162736
Yun S-W, Lee D-Y, Park H-S, Kim D-H. Heat-Processed Soybean Germ Extract and Lactobacillus gasseri NK109 Supplementation Reduce LPS-Induced Cognitive Impairment and Colitis in Mice. Nutrients. 2024; 16(16):2736. https://doi.org/10.3390/nu16162736
Chicago/Turabian StyleYun, Soo-Won, Dong-Yun Lee, Hee-Seo Park, and Dong-Hyun Kim. 2024. "Heat-Processed Soybean Germ Extract and Lactobacillus gasseri NK109 Supplementation Reduce LPS-Induced Cognitive Impairment and Colitis in Mice" Nutrients 16, no. 16: 2736. https://doi.org/10.3390/nu16162736
APA StyleYun, S.-W., Lee, D.-Y., Park, H.-S., & Kim, D.-H. (2024). Heat-Processed Soybean Germ Extract and Lactobacillus gasseri NK109 Supplementation Reduce LPS-Induced Cognitive Impairment and Colitis in Mice. Nutrients, 16(16), 2736. https://doi.org/10.3390/nu16162736