
The PPIase Activity of CypB Is Essential for the Activation of Both AKT/mTOR and XBP1s Signaling Pathways during the Differentiation of 3T3-L1 Preadipocytes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Differentiation and Culture of Cells
2.2. Plasmid and Transfection
2.3. Western Blot Analysis
2.4. Oil-Red-O Staining
2.5. FACS Analysis
2.6. Statistical Analyses
3. Results
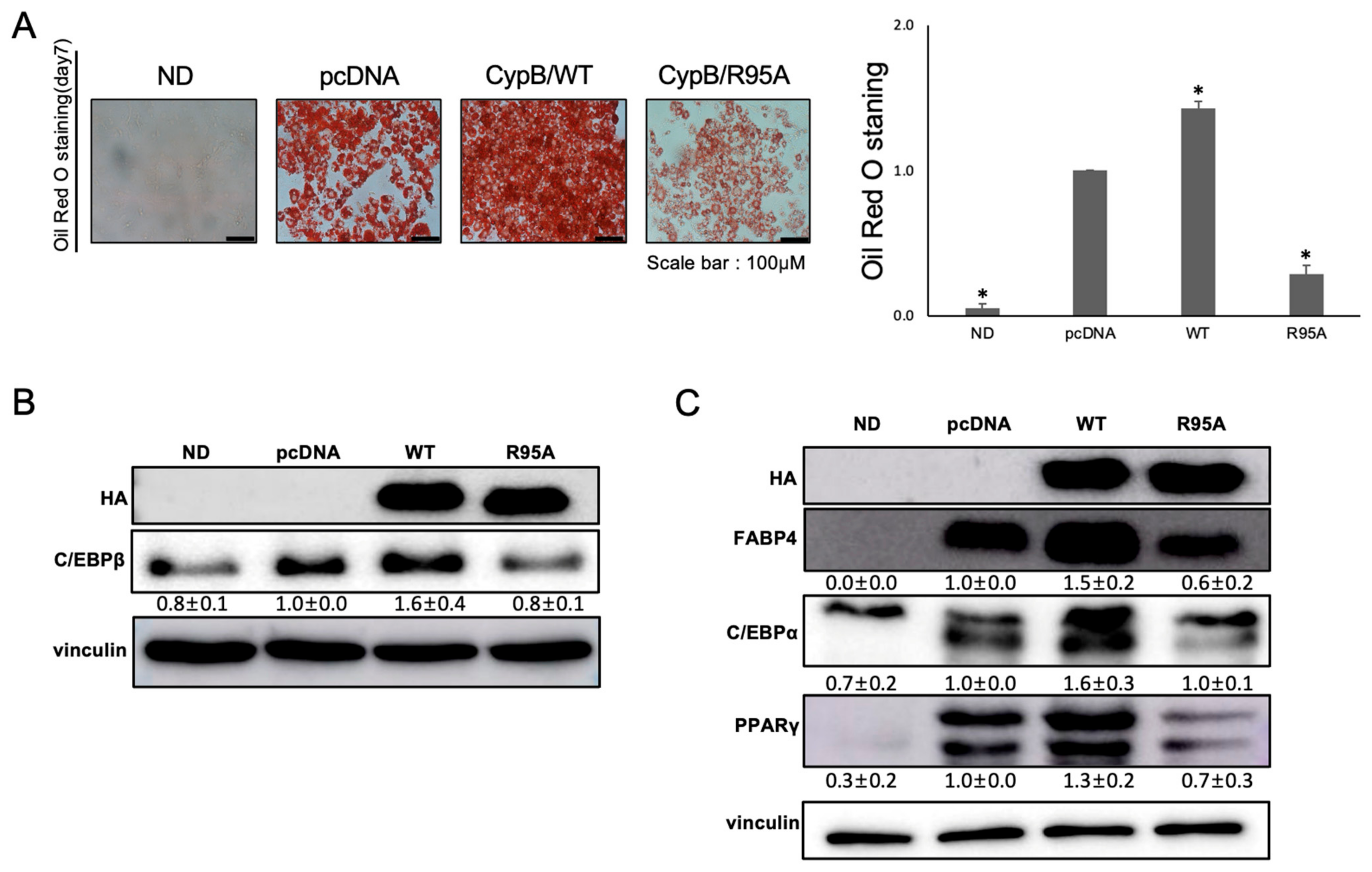
3.1. Impact of CypB PPIase on 3T3-L1 Cell Differentiation
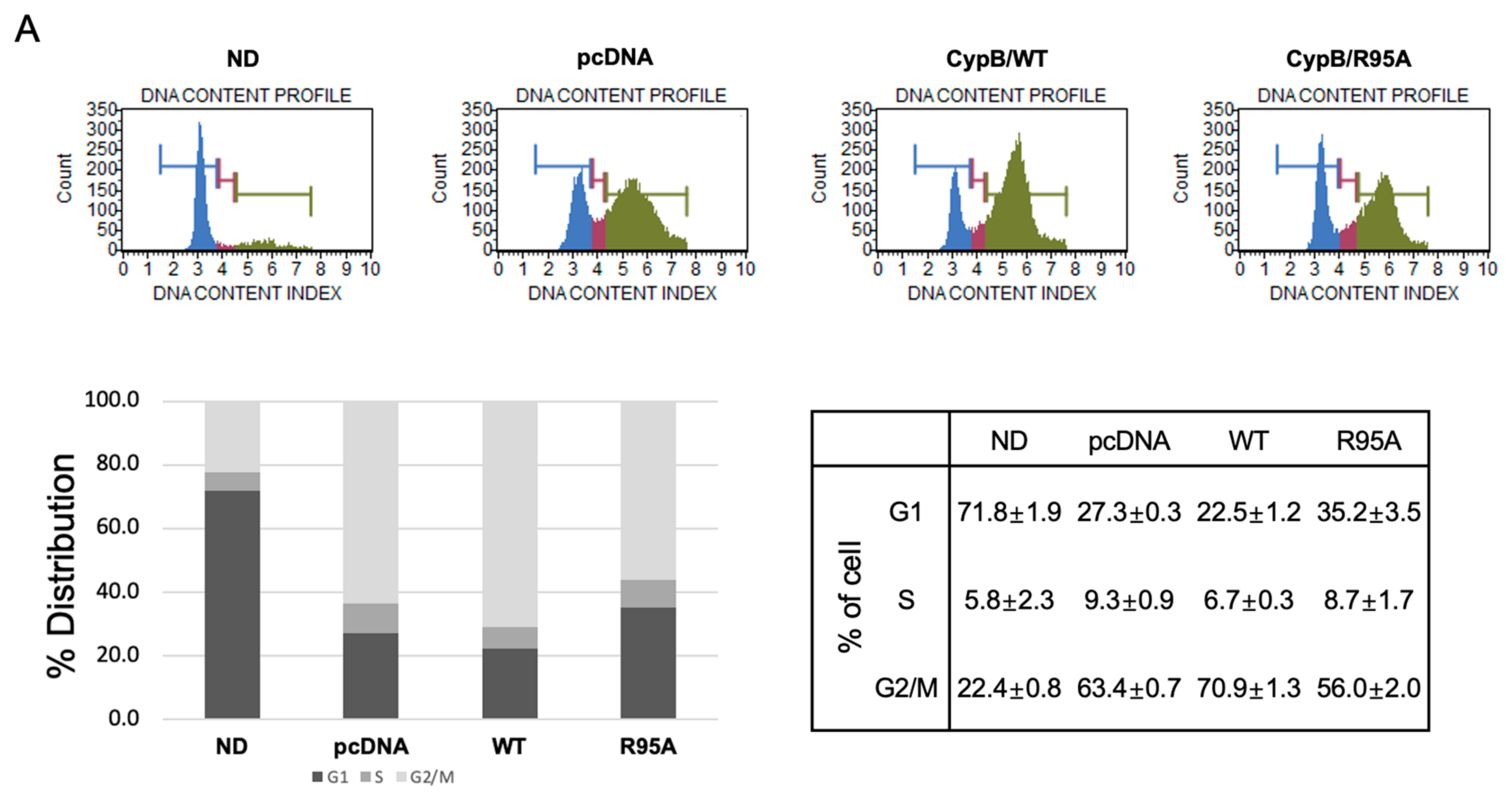
3.2. Inhibition of CypB’s PPIase Activity Suppresses Mitotic Clonal Expansion
3.3. Inhibition of CypB’s PPIase Activity Suppresses of AKT/mTOR Signaling Pathway in 3T3-L1 Cells
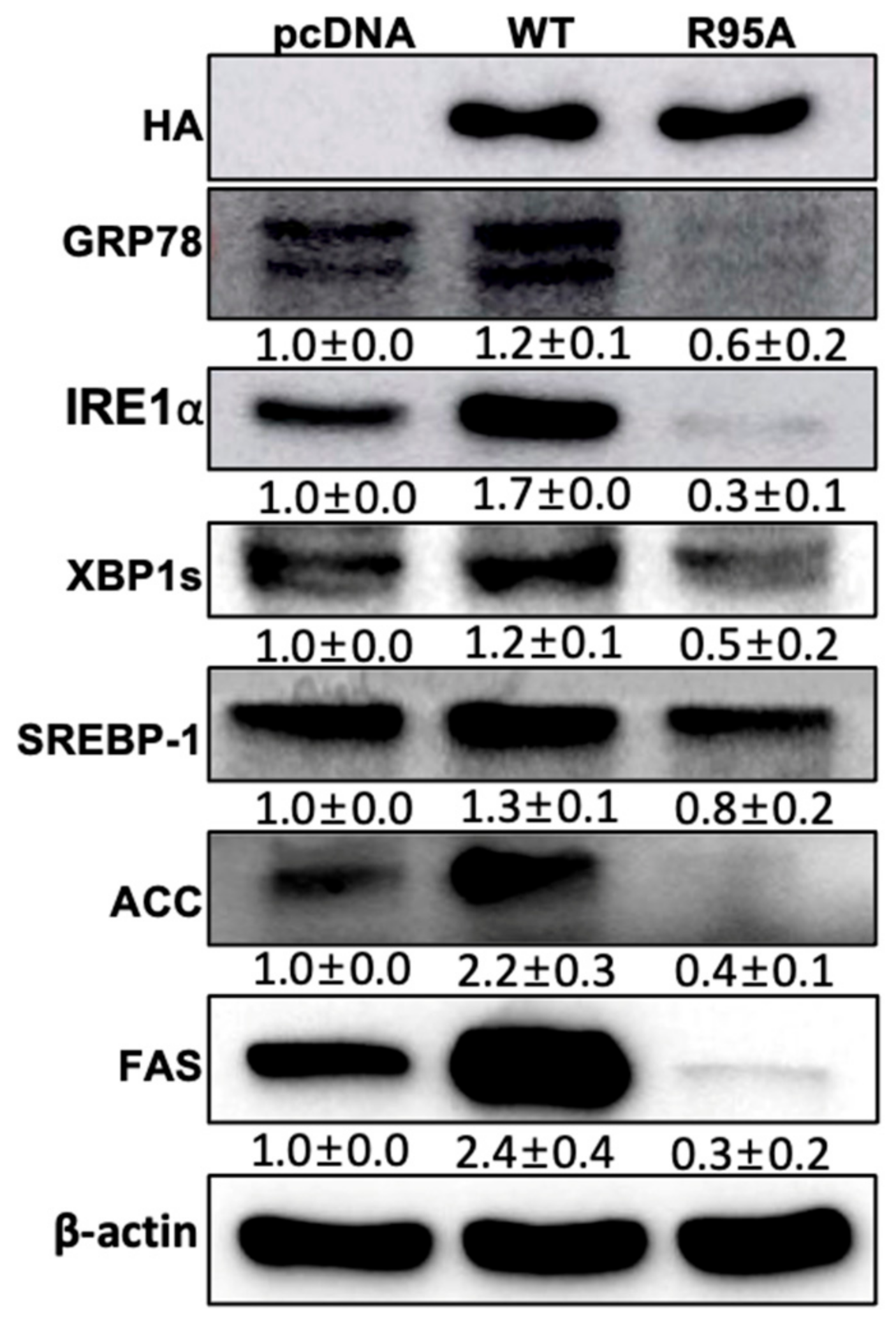
3.4. The PPIase Activity of CypB Affects Lipid Metabolism via the XBP1s Pathway
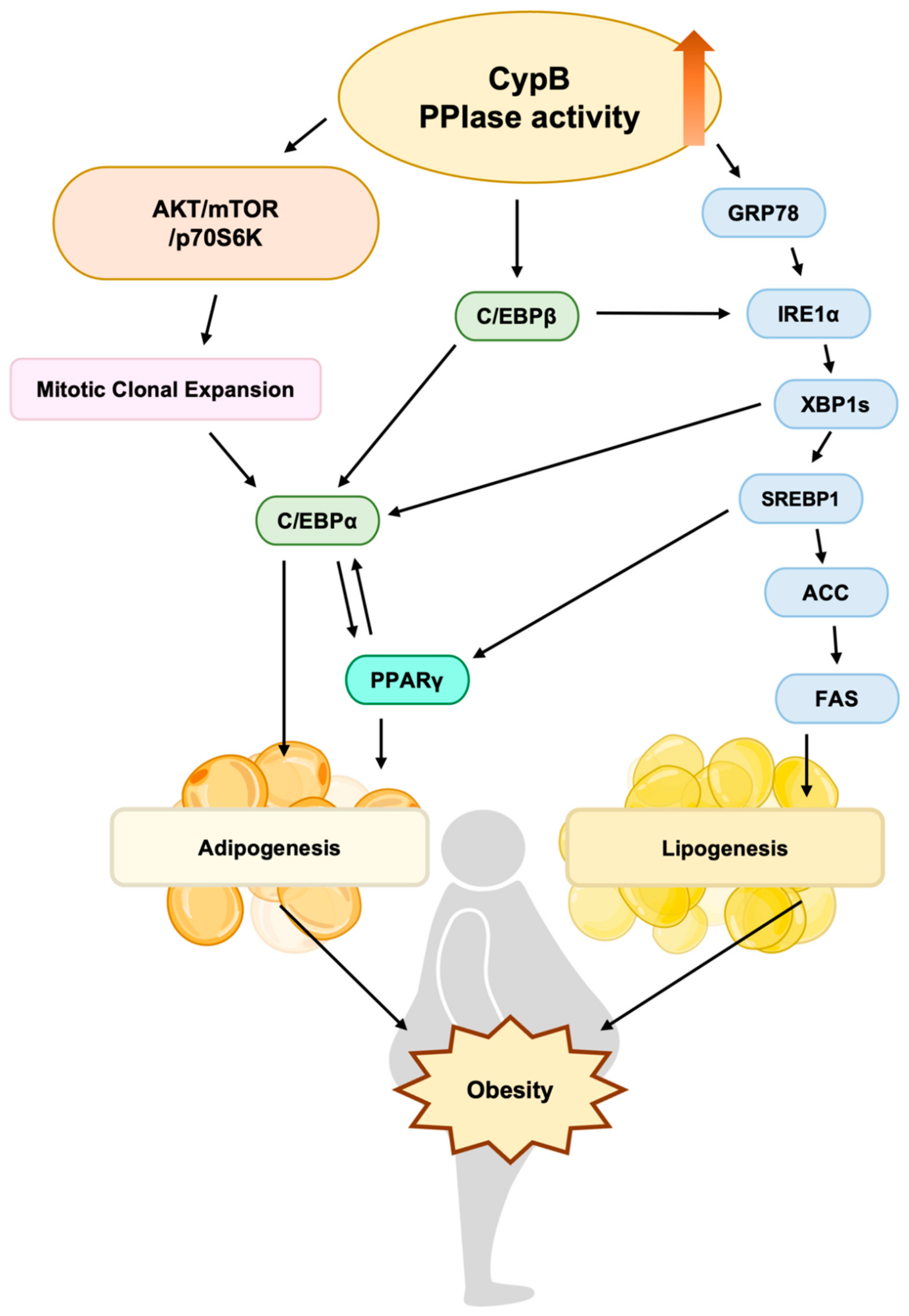
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- One in eight people are now living with obesity. Saudi Med. J. 2024, 45, 448–449.
- Chen, C.; Cui, Q.; Zhang, X.; Luo, X.; Liu, Y.; Zuo, J.; Peng, Y. Long non-coding RNAs regulation in adipogenesis and lipid metabolism: Emerging insights in obesity. Cell. Signal. 2018, 51, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Hachula, M.; Kosowski, M.; Zielanska, K.; Basiak, M.; Okopien, B. The Impact of Various Methods of Obesity Treatment on the Quality of Life and Mental Health—A Narrative Review. Int. J. Environ. Res. Public Health 2023, 20, 2122. [Google Scholar] [CrossRef] [PubMed]
- Heeren, J.; Scheja, L. Metabolic-associated fatty liver disease and lipoprotein metabolism. Mol. Metab. 2021, 50, 101238. [Google Scholar] [CrossRef] [PubMed]
- Cave, E.; Crowther, N.J. The Use of 3T3-L1 Murine Preadipocytes as a Model of Adipogenesis. Methods Mol. Biol. 2019, 1916, 263–272. [Google Scholar] [PubMed]
- Ruiz-Ojeda, F.J.; Ruperez, A.I.; Gomez-Llorente, C.; Gil, A.; Aguilera, C.M. Cell Models and Their Application for Studying Adipogenic Differentiation in Relation to Obesity: A Review. Int. J. Mol. Sci. 2016, 17, 1040. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Jeong, J.H.; Ryu, J.H. Lignan from Alnus japonica Inhibits Adipocyte Differentiation via Cell Cycle and FOXO1 Regulation. Molecules 2020, 25, 3346. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.J.; Seo, D.W.; Ju, J.Y.; Cha, Y.Y.; An, H.J. The Antiobesity Effects of Buginawa in 3T3-L1 Preadipocytes and in a Mouse Model of High-Fat Diet-Induced Obesity. Biomed. Res. Int. 2019, 2019, 3101987. [Google Scholar] [CrossRef] [PubMed]
- Madsen, M.S.; Siersbaek, R.; Boergesen, M.; Nielsen, R.; Mandrup, S. Peroxisome proliferator-activated receptor gamma and C/EBPalpha synergistically activate key metabolic adipocyte genes by assisted loading. Mol. Cell. Biol. 2014, 34, 939–954. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.S.; Kim, S.S.; Ha, J.; Kang, I.; Choe, W. Cyclophilin B, a molecule chaperone, promotes adipogenesis in 3T3-L1 preadipocytes via AKT/mTOR pathway. Int. J. Mol. Med. 2023, 51, 6. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.; Lee, J.; Ha, J.; Kang, I.; Choe, W. Endoplasmic Reticulum Stress and Its Impact on Adipogenesis: Molecular Mechanisms Implicated. Nutrients 2023, 15, 5082. [Google Scholar] [CrossRef] [PubMed]
- Barneda, D.; Christian, M. Lipid droplet growth: Regulation of a dynamic organelle. Curr. Opin. Cell Biol. 2017, 47, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Scott, C.C.; Vossio, S.; Rougemont, J.; Gruenberg, J. TFAP2 transcription factors are regulators of lipid droplet biogenesis. eLife 2018, 7, e36330. [Google Scholar] [CrossRef] [PubMed]
- Bravo, R.; Parra, V.; Gatica, D.; Rodriguez, A.E.; Torrealba, N.; Paredes, F.; Wang, Z.V.; Zorzano, A.; Hill, J.A.; Jaimovich, E.; et al. Endoplasmic reticulum and the unfolded protein response: Dynamics and metabolic integration. Int. Rev. Cell Mol. Biol. 2013, 301, 215–290. [Google Scholar] [PubMed]
- Cao, H.; Zhou, X.; Xu, B.; Hu, H.; Guo, J.; Ma, Y.; Wang, M.; Li, N.; Jun, Z. Advances in the study of protein folding and endoplasmic reticulum-associated degradation in mammal cells. J. Zhejiang Univ. Sci. B 2024, 25, 212–232. [Google Scholar] [CrossRef] [PubMed]
- Boden, G.; Duan, X.; Homko, C.; Molina, E.J.; Song, W.; Perez, O.; Cheung, P.; Merali, S. Increase in endoplasmic reticulum stress-related proteins and genes in adipose tissue of obese, insulin-resistant individuals. Diabetes 2008, 57, 2438–2444. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-S.; Jeong, S.; Kim, K.-Y.; Yoon, J.-S.; Kim, S.; Yoon, K.-S.; Ha, J.; Kang, I.; Choe, W. Honokiol inhibits hepatoma carcinoma cell migration through downregulated Cyclophilin B expression. Biochem. Biophys. Res. Commun. 2021, 552, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Dunyak, B.M.; Gestwicki, J.E. Peptidyl-Proline Isomerases (PPIases): Targets for Natural Products and Natural Product-Inspired Compounds. J. Med. Chem. 2016, 59, 9622–9644. [Google Scholar] [CrossRef] [PubMed]
- Galigniana, N.M.; Ruiz, M.C.; Piwien-Pilipuk, G. FK506 binding protein 51: Its role in the adipose organ and beyond. J. Cell. Biochem. 2022, early view, 1–14. [Google Scholar] [CrossRef]
- Inoue, M.-K.; Nakatsu, Y.; Yamamotoya, T.; Hasei, S.; Kanamoto, M.; Naitou, M.; Matsunaga, Y.; Sakoda, H.; Fujishiro, M.; Ono, H.; et al. Pin1 Plays Essential Roles in NASH Development by Modulating Multiple Target Proteins. Cells 2019, 8, 1545. [Google Scholar] [CrossRef] [PubMed]
- Stechschulte, L.A.; Hinds, T.D., Jr.; Khuder, S.S.; Shou, W.; Najjar, S.M.; Sanchez, E.R. FKBP51 controls cellular adipogenesis through p38 kinase-mediated phosphorylation of GRalpha and PPARgamma. Mol. Endocrinol. 2014, 28, 1265–1275. [Google Scholar] [CrossRef] [PubMed]
- Nakatsu, Y.; Sakoda, H.; Kushiyama, A.; Zhang, J.; Ono, H.; Fujishiro, M.; Kikuchi, T.; Fukushima, T.; Yoneda, M.; Ohno, H.; et al. Peptidyl-prolyl cis/trans isomerase NIMA-interacting 1 associates with insulin receptor substrate-1 and enhances insulin actions and adipogenesis. J. Biol. Chem. 2011, 286, 20812–20822. [Google Scholar] [CrossRef] [PubMed]
- Schiene-Fischer, C.; Fischer, G.; Braun, M. Non-Immunosuppressive Cyclophilin Inhibitors. Angew. Chem. Int. Ed. 2022, 61, e202201597. [Google Scholar] [CrossRef] [PubMed]
- Stauffer, W.T.; Goodman, A.Z.; Bobardt, M.; Ure, D.R.; Foster, R.T.; Gallay, P. Mice lacking cyclophilin B, but not cyclophilin A, are protected from the development of NASH in a diet and chemical-induced model. PLoS ONE 2024, 19, e0298211. [Google Scholar] [CrossRef] [PubMed]
- Kanna, M.; Nakatsu, Y.; Yamamotoya, T.; Encinas, J.; Ito, H.; Okabe, T.; Asano, T.; Sakaguchi, T. Roles of peptidyl prolyl isomerase Pin1 in viral propagation. Front. Cell Dev. Biol. 2022, 10, 1005325. [Google Scholar] [CrossRef]
- Liu, J.; Zuo, Y.; Qu, G.M.; Song, X.; Liu, Z.H.; Zhang, T.G.; Zheng, Z.H.; Wang, H.K. CypB promotes cell proliferation and metastasis in endometrial carcinoma. BMC Cancer 2021, 21, 747. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Ma, Z.; Zhang, Y.; Zhang, M.; Shi, X.; Zhang, M.; Zhang, W.; Liu, W. The role of cyclophilins in viral infection and the immune response. J. Infect. 2022, 85, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Teng, M.R.; Huang, J.A.; Zhu, Z.T.; Li, H.; Shen, J.F.; Chen, Q. Cyclophilin B promotes cell proliferation, migration, invasion and angiogenesis via regulating the STAT3 pathway in non-small cell lung cancer. Pathol. Res. Pract. 2019, 215, 152417. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, Z.; Zhang, B.; He, H.; Bai, Y. PPIA is a novel adipogenic factor implicated in obesity. Obesity 2015, 23, 2093–2100. [Google Scholar] [CrossRef]
- Torpey, J.; Madine, J.; Wood, A.; Lian, L.Y. Cyclophilin D binds to the acidic C-terminus region of alpha-Synuclein and affects its aggregation characteristics. Sci. Rep. 2020, 10, 10159. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-C.; Wang, W.-D.; Wang, J.-S.; Pan, J.-C. PPIase independent chaperone-like function of recombinant human Cyclophilin A during arginine kinase refolding. FEBS Lett. 2013, 587, 666–672. [Google Scholar] [CrossRef] [PubMed]
- Rostam, M.A.; Piva, T.J.; Rezaei, H.B.; Kamato, D.; Little, P.J.; Zheng, W.; Osman, N. Peptidyl-prolyl isomerases: Functionality and potential therapeutic targets in cardiovascular disease. Clin. Exp. Pharmacol. Physiol. 2015, 42, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Battineni, G.; Sagaro, G.G.; Chintalapudi, N.; Amenta, F.; Tomassoni, D.; Tayebati, S.K. Impact of Obesity-Induced Inflammation on Cardiovascular Diseases (CVD). Int. J. Mol. Sci. 2021, 22, 4798. [Google Scholar] [CrossRef] [PubMed]
- Jeong, K.; Kim, H.; Kim, K.; Kim, S.-J.; Hahn, B.-S.; Jahng, G.-H.; Yoon, K.-S.; Kim, S.-S.; Ha, J.; Kang, I.; et al. Cyclophilin B is involved in p300-mediated degradation of CHOP in tumor cell adaptation to hypoxia. Cell Death Differ. 2014, 21, 438–450. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.-H.; Jang, S.-H.; Won, C.; Kim, C.-H.; Kim, H.-D.; Kim, T.-H.; Cho, J.-H. Derhamnosylmaysin Inhibits Adipogenesis via Inhibiting Expression of PPARgamma and C/EBPalpha in 3T3-L1 Cells. Molecules 2022, 27, 4232. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Yang, X.; Zhao, Q.; Li, Z.; Fu, F.; Zhang, H.; Zheng, M.; Zhang, S. Molecular Mechanism of Stem Cell Differentiation into Adipocytes and Adipocyte Differentiation of Malignant Tumor. Stem Cells Int. 2020, 2020, 8892300. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Wang, S.; Malhotra, J.; Hassler, J.R.; Back, S.H.; Wang, G.; Chang, L.; Xu, W.; Miao, H.; Leonardi, R.; et al. The unfolded protein response transducer IRE1alpha prevents ER stress-induced hepatic steatosis. EMBO J. 2011, 30, 1357–1375. [Google Scholar] [CrossRef] [PubMed]
- Sha, H.; He, Y.; Chen, H.; Wang, C.; Zenno, A.; Shi, H.; Yang, X.; Zhang, X.; Qi, L. The IRE1alpha-XBP1 pathway of the unfolded protein response is required for adipogenesis. Cell Metab. 2009, 9, 556–564. [Google Scholar] [CrossRef] [PubMed]
- Ning, J.; Hong, T.; Ward, A.; Pi, J.; Liu, Z.; Liu, H.Y.; Cao, W. Constitutive role for IRE1alpha-XBP1 signaling pathway in the insulin-mediated hepatic lipogenic program. Endocrinology 2011, 152, 2247–2255. [Google Scholar] [CrossRef]
- Lee, Y.-J.; Choi, H.-S.; Seo, M.-J.; Jeon, H.-J.; Kim, K.-J.; Lee, B.-Y. Kaempferol suppresses lipid accumulation by inhibiting early adipogenesis in 3T3-L1 cells and zebrafish. Food Funct. 2015, 6, 2824–2833. [Google Scholar] [CrossRef]
- Han, J.; Kaufman, R.J. The role of ER stress in lipid metabolism and lipotoxicity. J. Lipid Res. 2016, 57, 1329–1338. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.H.; Scapa, E.F.; Cohen, D.E.; Glimcher, L.H. Regulation of hepatic lipogenesis by the transcription factor XBP1. Science 2008, 320, 1492–1496. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, G.; Yoon, K.-S.; Ha, J.; Kang, I.; Choe, W. The PPIase Activity of CypB Is Essential for the Activation of Both AKT/mTOR and XBP1s Signaling Pathways during the Differentiation of 3T3-L1 Preadipocytes. Nutrients 2024, 16, 2465. https://doi.org/10.3390/nu16152465
Kim G, Yoon K-S, Ha J, Kang I, Choe W. The PPIase Activity of CypB Is Essential for the Activation of Both AKT/mTOR and XBP1s Signaling Pathways during the Differentiation of 3T3-L1 Preadipocytes. Nutrients. 2024; 16(15):2465. https://doi.org/10.3390/nu16152465
Chicago/Turabian StyleKim, Gyuhui, Kyung-Sik Yoon, Joohun Ha, Insug Kang, and Wonchae Choe. 2024. "The PPIase Activity of CypB Is Essential for the Activation of Both AKT/mTOR and XBP1s Signaling Pathways during the Differentiation of 3T3-L1 Preadipocytes" Nutrients 16, no. 15: 2465. https://doi.org/10.3390/nu16152465
APA StyleKim, G., Yoon, K.-S., Ha, J., Kang, I., & Choe, W. (2024). The PPIase Activity of CypB Is Essential for the Activation of Both AKT/mTOR and XBP1s Signaling Pathways during the Differentiation of 3T3-L1 Preadipocytes. Nutrients, 16(15), 2465. https://doi.org/10.3390/nu16152465