
The Effects of Different Dietary Patterns on Bone Health


Abstract
1. Introduction
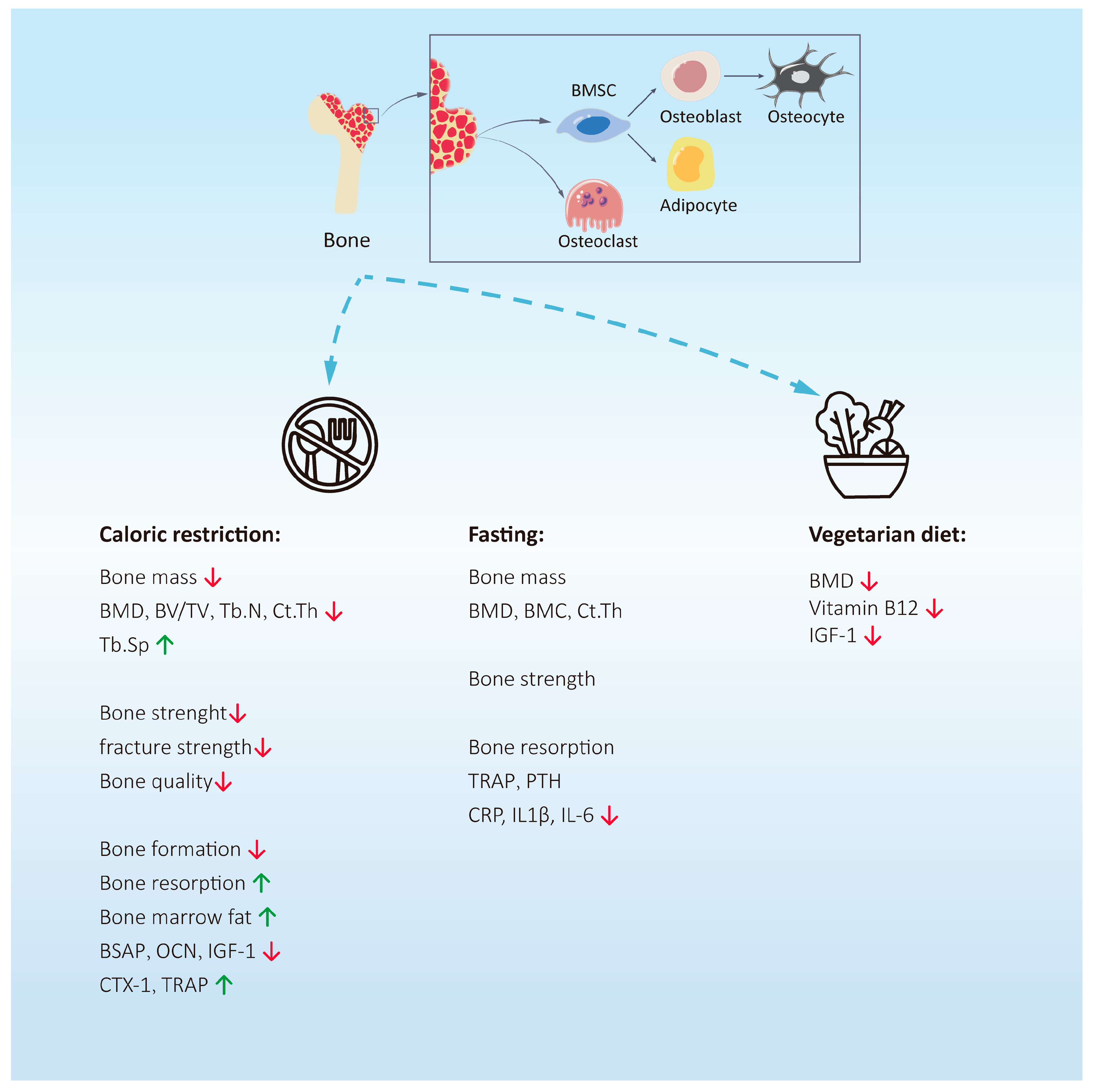
2. Intermittent Fasting and Bone Health
2.1. Classification of Intermittent Fasting
2.2. Basic Research on Intermittent Fasting and Bone Metabolism
2.3. Clinical Research on Intermittent Fasting and Bone Metabolism
3. Caloric Restriction and Bone Health
4. Vegetarian Diet and Bone Health
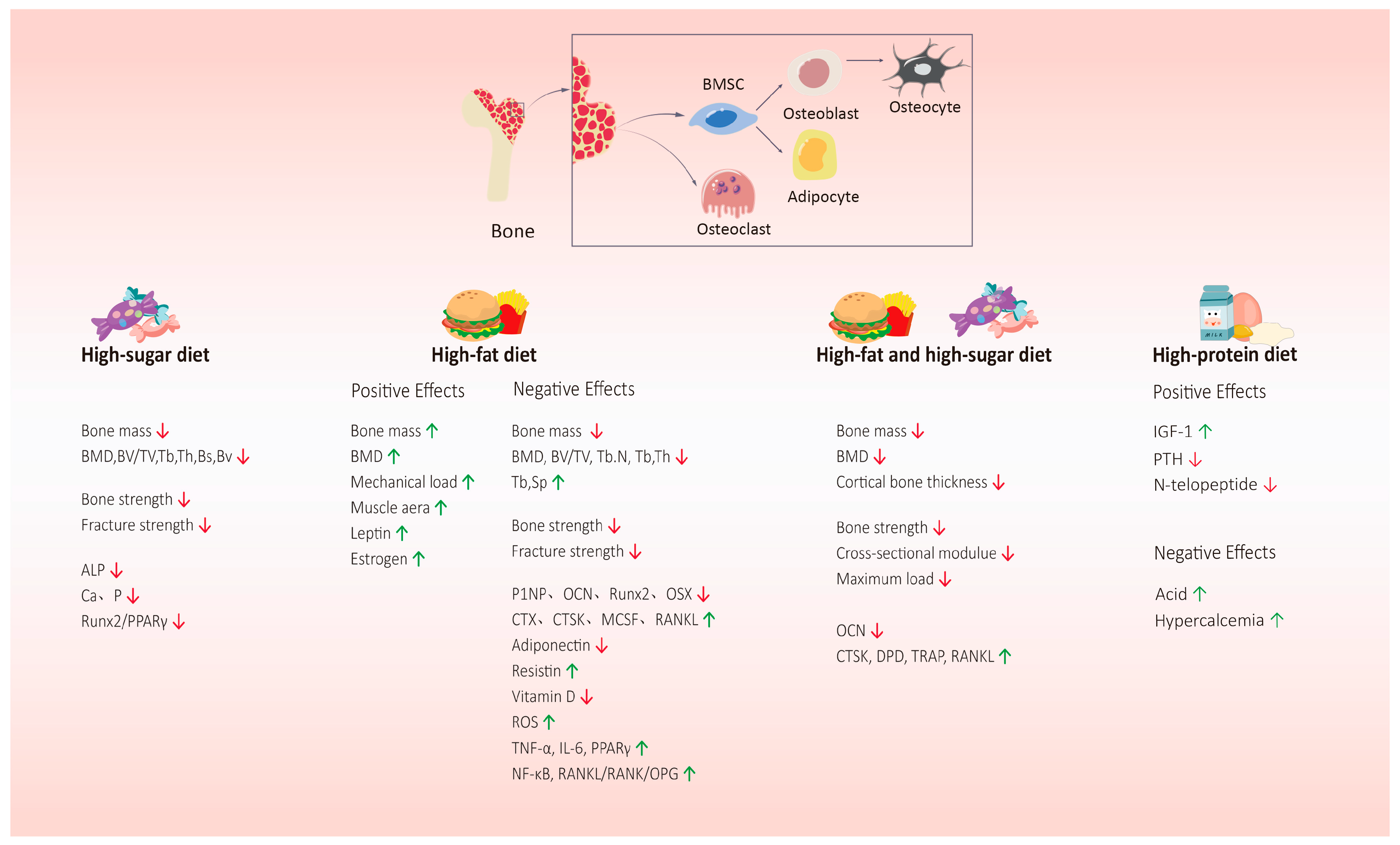
5. High-Sugar and/or High-Fat Dietary Patterns and Bone Health
5.1. HSD and Bone Health
5.1.1. Effect of HSD on Bone Mass and Bone Strength
5.1.2. Effect of HSD on Bone Metabolism
5.2. HFD and Bone Health
5.2.1. Effect of HFD on Bone Mass and Bone Strength
5.2.2. Effect of HFD on Bone Metabolism
5.3. HSFD and Bone Health
5.3.1. Effect of HSFD on Bone Mass and Bone Strength
5.3.2. Effect of HSFD on Bone Metabolism
6. High-Protein Dietary Patterns and Bone Health
7. Intake of Calcium, Vitamin D, and Dairy Products and Bone Health
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Raggatt, L.J.; Partridge, N.C. Cellular and molecular mechanisms of bone remodeling. J. Biol. Chem. 2010, 285, 25103–25108. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Garach, A.; García-Fontana, B.; Muñoz-Torres, M. Nutrients and Dietary Patterns Related to Osteoporosis. Nutrients 2020, 12, 1986. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.H.; Chen, L.R.; Chen, K.H. Osteoporosis Due to Hormone Imbalance: An Overview of the Effects of Estrogen Deficiency and Glucocorticoid Overuse on Bone Turnover. Int. J. Mol. Sci. 2022, 23, 1376. [Google Scholar] [CrossRef] [PubMed]
- Tsugawa, N.; Shiraki, M. Vitamin K Nutrition and Bone Health. Nutrients 2020, 12, 1909. [Google Scholar] [CrossRef] [PubMed]
- Rizzoli, R.; Biver, E.; Brennan-Speranza, T.C. Nutritional intake and bone health. Lancet Diabetes Endocrinol. 2021, 9, 606–621. [Google Scholar] [CrossRef] [PubMed]
- Ji, T.; Fang, B.; Wu, F.; Liu, Y.; Cheng, L.; Li, Y.; Wang, R.; Zhu, L. Diet Change Improves Obesity and Lipid Deposition in High-Fat Diet-Induced Mice. Nutrients 2023, 15, 4978. [Google Scholar] [CrossRef] [PubMed]
- Waddington, G.S. COVID-19, mental health and physical activity. J. Sci. Med. Sport 2021, 24, 319. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Zhong, Z.; Huang, C.; Wang, W. The effects of popular diets on bone health in the past decade: A narrative review. Front. Endocrinol. 2023, 14, 1287140. [Google Scholar] [CrossRef] [PubMed]
- Ogilvie, A.R.; McGuire, B.D.; Meng, L.; Shapses, S.A. Fracture Risk in Vegetarians and Vegans: The Role of Diet and Metabolic Factors. Curr. Osteoporos. Rep. 2022, 20, 442–452. [Google Scholar] [CrossRef]
- Grajower, M.M.; Horne, B.D. Clinical Management of Intermittent Fasting in Patients with Diabetes Mellitus. Nutrients 2019, 11, 873. [Google Scholar] [CrossRef] [PubMed]
- Klempel, M.C.; Kroeger, C.M.; Bhutani, S.; Trepanowski, J.F.; Varady, K.A. Intermittent fasting combined with calorie restriction is effective for weight loss and cardio-protection in obese women. Nutr. J. 2012, 11, 98. [Google Scholar] [CrossRef] [PubMed]
- Patterson, R.E.; Laughlin, G.A.; LaCroix, A.Z.; Hartman, S.J.; Natarajan, L.; Senger, C.M.; Martinez, M.E.; Villasenor, A.; Sears, D.D.; Marinac, C.R.; et al. Intermittent Fasting and Human Metabolic Health. J. Acad. Nutr. Diet. 2015, 115, 1203–1212. [Google Scholar] [CrossRef] [PubMed]
- Santos, H.O.; Macedo, R.C.O. Impact of intermittent fasting on the lipid profile: Assessment associated with diet and weight loss. Clin. Nutr. ESPEN 2018, 24, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Mattson, M.P.; Longo, V.D.; Harvie, M. Impact of intermittent fasting on health and disease processes. Ageing Res. Rev. 2017, 39, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Donahue, S.W.; Vaughan, M.R.; Demers, L.M.; Donahue, H.J. Serum markers of bone metabolism show bone loss in hibernating bears. Clin. Orthop. Relat. Res. 2003, 408, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Hisatomi, Y.; Kugino, K. Changes in bone density and bone quality caused by single fasting for 96 hours in rats. PeerJ 2019, 6, e6161. [Google Scholar] [CrossRef] [PubMed]
- Alrowaili, M.G.; Hussein, A.M.; Eid, E.A.; Serria, M.S.; Abdellatif, H.; Sakr, H.F. Effect of Intermittent Fasting on Glucose Homeostasis and Bone Remodeling in Glucocorticoid-Induced Osteoporosis Rat Model. J. Bone Metab. 2021, 28, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Ding, J.; Wu, X.; Huang, Z.; Kong, G.; Liu, Q.; Yang, Z.; Huang, Z.; Zhu, Q. Bone microstructure and metabolism changes under the combined intervention of ketogenic diet with intermittent fasting: An in vivo study of rats. Exp. Anim. 2019, 68, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Bahijri, S.M.; Ajabnoor, G.M.; Borai, A.; Al-Aama, J.Y.; Chrousos, G.P. Effect of Ramadan fasting in Saudi Arabia on serum bone profile and immunoglobulins. Ther. Adv. Endocrinol. Metab. 2015, 6, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Kroll, M.H. Parathyroid hormone temporal effects on bone formation and resorption. Bull. Math. Biol. 2000, 62, 163–188. [Google Scholar] [CrossRef] [PubMed]
- Ben Nessib, D.; Maatallah, K.; Ferjani, H.; Kaffel, D.; Hamdi, W. The potential effect of Ramadan fasting on musculoskeletal diseases: New perspectives. Clin. Rheumatol. 2021, 40, 833–839. [Google Scholar] [CrossRef]
- Rodopaios, N.E.; Petridou, A.; Mougios, V.; Koulouri, A.A.; Vasara, E.; Papadopoulou, S.K.; Skepastianos, P.; Hassapidou, M.; Kafatos, A.G. Vitamin D status, vitamin D intake, and sunlight exposure in adults adhering or not to periodic religious fasting for decades. Int. J. Food Sci. Nutr. 2021, 72, 989–996. [Google Scholar] [CrossRef] [PubMed]
- Barnosky, A.; Kroeger, C.M.; Trepanowski, J.F.; Klempel, M.C.; Bhutani, S.; Hoddy, K.K.; Gabel, K.; Shapses, S.A.; Varady, K.A. Effect of alternate day fasting on markers of bone metabolism: An exploratory analysis of a 6-month randomized controlled trial. Nutr. Healthy Aging 2017, 4, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Clayton, D.J.; James, L.J.; Sale, C.; Templeman, I.; Betts, J.A.; Varley, I. Severely restricting energy intake for 24 h does not affect markers of bone metabolism at rest or in response to re-feeding. Eur. J. Nutr. 2020, 59, 3527–3535. [Google Scholar] [CrossRef] [PubMed]
- Martens, C.R.; Rossman, M.J.; Mazzo, M.R.; Jankowski, L.R.; Nagy, E.E.; Denman, B.A.; Richey, J.J.; Johnson, S.A.; Ziemba, B.P.; Wang, Y.; et al. Short-term time-restricted feeding is safe and feasible in non-obese healthy midlife and older adults. Geroscience 2020, 42, 667–686. [Google Scholar] [CrossRef] [PubMed]
- Most, J.; Tosti, V.; Redman, L.M.; Fontana, L. Calorie restriction in humans: An update. Ageing Res. Rev. 2017, 39, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Welton, S.; Minty, R.; O’Driscoll, T.; Willms, H.; Poirier, D.; Madden, S.; Kelly, L. Intermittent fasting and weight loss: Systematic review. Can. Fam. Physician 2020, 66, 117–125. [Google Scholar] [PubMed]
- Veronese, N.; Stubbs, B.; Koyanagi, A.; Hebert, J.R.; Cooper, C.; Caruso, M.G.; Guglielmi, G.; Reginster, J.Y.; Rizzoli, R.; Maggi, S.; et al. Pro-inflammatory dietary pattern is associated with fractures in women: An eight-year longitudinal cohort study. Osteoporos. Int. 2018, 29, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Gao, Y.; Hua, R.; Lu, M.; Chen, H.; Wang, Z.; Li, L.; Hu, K.; Yin, Y.; Xu, K.; et al. Calorie intake rather than food quantity consumed is the key factor for the anti-aging effect of calorie restriction. Aging 2021, 13, 21526–21546. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.J. Caloric restriction combined with exercise is effective in reducing adiposity and mitigating bone structural deterioration in obese rats. Ann. N. Y. Acad. Sci. 2018, 1433, 41–52. [Google Scholar] [CrossRef] [PubMed]
- McGrath, C.; Sankaran, J.S.; Misaghian-Xanthos, N.; Sen, B.; Xie, Z.; Styner, M.A.; Zong, X.; Rubin, J.; Styner, M. Exercise Degrades Bone in Caloric Restriction, Despite Suppression of Marrow Adipose Tissue (MAT). J. Bone Miner Res. 2020, 35, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Devlin, M.J.; Cloutier, A.M.; Thomas, N.A.; Panus, D.A.; Lotinun, S.; Pinz, I.; Baron, R.; Rosen, C.J.; Bouxsein, M.L. Caloric restriction leads to high marrow adiposity and low bone mass in growing mice. J. Bone Miner Res. 2010, 25, 2078–2088. [Google Scholar] [CrossRef] [PubMed]
- Singh, L.; Tyagi, S.; Myers, D.; Duque, G. Good, Bad, or Ugly: The Biological Roles of Bone Marrow Fat. Curr. Osteoporos. Rep. 2018, 16, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Duque, G.; Al Saedi, A.; Rivas, D.; Miard, S.; Ferland, G.; Picard, F.; Gaudreau, P. Differential Effects of Long-Term Caloric Restriction and Dietary Protein Source on Bone and Marrow Fat of the Aging Rat. J. Gerontol. A Biol. Sci. Med. Sci. 2020, 75, 2031–2036. [Google Scholar] [CrossRef] [PubMed]
- Devlin, M.J.; Brooks, D.J.; Conlon, C.; Vliet, M.; Louis, L.; Rosen, C.J.; Bouxsein, M.L. Daily leptin blunts marrow fat but does not impact bone mass in calorie-restricted mice. J. Endocrinol. 2016, 229, 295–306. [Google Scholar] [CrossRef]
- Talbott, S.M.; Cifuentes, M.; Dunn, M.G.; Shapses, S.A. Energy restriction reduces bone density and biomechanical properties in aged female rats. J. Nutr. 2001, 131, 2382–2387. [Google Scholar] [CrossRef] [PubMed]
- Bodnar, M.; Skalicky, M.; Viidik, A.; Erben, R.G. Interaction between exercise, dietary restriction and age-related bone loss in a rodent model of male senile osteoporosis. Gerontology 2012, 58, 139–149. [Google Scholar] [CrossRef] [PubMed]
- LaMothe, J.M.; Hepple, R.T.; Zernicke, R.F. Selected contribution: Bone adaptation with aging and long-term caloric restriction in Fischer 344 x Brown-Norway F1-hybrid rats. J. Appl. Physiol. 2003, 95, 1739–1745. [Google Scholar] [CrossRef] [PubMed]
- Behrendt, A.K.; Kuhla, A.; Osterberg, A.; Polley, C.; Herlyn, P.; Fischer, D.C.; Scotland, M.; Wree, A.; Histing, T.; Menger, M.D.; et al. Dietary Restriction-Induced Alterations in Bone Phenotype: Effects of Lifelong Versus Short-Term Caloric Restriction on Femoral and Vertebral Bone in C57BL/6 Mice. J. Bone Miner Res. 2016, 31, 852–863. [Google Scholar] [CrossRef] [PubMed]
- Ahn, H.; Seo, D.H.; Kim, H.S.; Choue, R. Calorie restriction aggravated cortical and trabecular bone architecture in ovariectomy-induced estrogen-deficient rats. Nutr. Res. 2014, 34, 707–713. [Google Scholar] [CrossRef] [PubMed]
- Hamrick, M.W.; Ding, K.H.; Ponnala, S.; Ferrari, S.L.; Isales, C.M. Caloric restriction decreases cortical bone mass but spares trabecular bone in the mouse skeleton: Implications for the regulation of bone mass by body weight. J. Bone Miner Res. 2008, 23, 870–878. [Google Scholar] [CrossRef] [PubMed]
- Aikawa, Y.; Agata, U.; Kakutani, Y.; Kato, S.; Noma, Y.; Hattori, S.; Ogata, H.; Ezawa, I.; Omi, N. The Preventive Effect of Calcium Supplementation on Weak Bones Caused by the Interaction of Exercise and Food Restriction in Young Female Rats During the Period from Acquiring Bone Mass to Maintaining Bone Mass. Calcif. Tissue Int. 2016, 98, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Jeon, Y.K.; Kim, W.J.; Shin, M.J.; Chung, H.Y.; Kim, S.S.; Kim, B.H.; Kim, S.J.; Kim, Y.K.; Kim, I.J. Short-term caloric restriction does not reduce bone mineral density in rats with early type 2 diabetes. Endocrinol. Metab. 2014, 29, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Fontinele, R.G.; Krause Neto, W.; Gama, E.F.; Brito Mari, R.; de Souza, R.R.; Conrado, A.; Mochizuki, L.; Kfoury Junior, J.R. Caloric restriction minimizes aging effects on the femoral medial condyle. Aging Male 2017, 20, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Villareal, D.T.; Kotyk, J.J.; Armamento-Villareal, R.C.; Kenguva, V.; Seaman, P.; Shahar, A.; Wald, M.J.; Kleerekoper, M.; Fontana, L. Reduced bone mineral density is not associated with significantly reduced bone quality in men and women practicing long-term calorie restriction with adequate nutrition. Aging Cell 2011, 10, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Villareal, D.T.; Fontana, L.; Weiss, E.P.; Racette, S.B.; Steger-May, K.; Schechtman, K.B.; Klein, S.; Holloszy, J.O. Bone mineral density response to caloric restriction-induced weight loss or exercise-induced weight loss: A randomized controlled trial. Arch. Intern. Med. 2006, 166, 2502–2510. [Google Scholar] [CrossRef] [PubMed]
- Villareal, D.T.; Fontana, L.; Das, S.K.; Redman, L.; Smith, S.R.; Saltzman, E.; Bales, C.; Rochon, J.; Pieper, C.; Huang, M.; et al. Effect of Two-Year Caloric Restriction on Bone Metabolism and Bone Mineral Density in Non-Obese Younger Adults: A Randomized Clinical Trial. J. Bone Miner Res. 2016, 31, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Razny, U.; Goralska, J.; Calder, P.C.; Gruca, A.; Childs, C.E.; Kapusta, M.; Slowinska-Solnica, K.; Dembinska-Kiec, A.; Solnica, B.; Malczewska-Malec, M. The Effect of Caloric Restriction with and without n-3 PUFA Supplementation on Bone Turnover Markers in Blood of Subjects with Abdominal Obesity: A Randomized Placebo-Controlled Trial. Nutrients 2021, 13, 3096. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, N.; Yano, T. Concomitant assessment of DNA oxidation and bone resorption over a rapid body mass reduction period in female judokas. J. Biol. Regul. Homeost. Agents 2018, 32, 781–790. [Google Scholar] [PubMed]
- Villareal, D.T.; Chode, S.; Parimi, N.; Sinacore, D.R.; Hilton, T.; Armamento-Villareal, R.; Napoli, N.; Qualls, C.; Shah, K. Weight loss, exercise, or both and physical function in obese older adults. N. Engl. J. Med. 2011, 364, 1218–1229. [Google Scholar] [CrossRef]
- Flanagan, E.W.; Most, J.; Mey, J.T.; Redman, L.M. Calorie Restriction and Aging in Humans. Annu. Rev. Nutr. 2020, 40, 105–133. [Google Scholar] [CrossRef] [PubMed]
- Locher, J.L.; Goldsby, T.U.; Goss, A.M.; Kilgore, M.L.; Gower, B.; Ard, J.D. Calorie restriction in overweight older adults: Do benefits exceed potential risks? Exp. Gerontol. 2016, 86, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, S.M.; Raposo, A.; Saraiva, A.; Zandonadi, R.P. Vegetarian Diet: An Overview through the Perspective of Quality of Life Domains. Int. J. Environ. Res. Public Health 2021, 18, 4067. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, N.S.; Sabate, J.; Jaceldo-Siegl, K.; Fraser, G.E. Vegetarian dietary patterns are associated with a lower risk of metabolic syndrome: The adventist health study 2. Diabetes Care 2011, 34, 1225–1227. [Google Scholar] [CrossRef] [PubMed]
- Lau, E.M.; Kwok, T.; Woo, J.; Ho, S.C. Bone mineral density in Chinese elderly female vegetarians, vegans, lacto-vegetarians and omnivores. Eur. J. Clin. Nutr. 1998, 52, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Ho-Pham, L.T.; Nguyen, N.D.; Nguyen, T.V. Effect of vegetarian diets on bone mineral density: A Bayesian meta-analysis. Am. J. Clin. Nutr. 2009, 90, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Iguacel, I.; Miguel-Berges, M.L.; Gomez-Bruton, A.; Moreno, L.A.; Julian, C. Veganism, vegetarianism, bone mineral density, and fracture risk: A systematic review and meta-analysis. Nutr. Rev. 2019, 77, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Pawlak, R. Vitamin B12 status is a risk factor for bone fractures among vegans. Med. Hypotheses 2021, 153, 110625. [Google Scholar] [CrossRef] [PubMed]
- Chuang, T.L.; Koo, M.; Chuang, M.H.; Lin, C.H.; Huang, C.H.; Wang, Y.F. Changes in Bone Mineral Density and Trabecular Bone Score over Time between Vegetarian and Non-Vegetarian Middle-Aged and Older Women: A Three-Year Retrospective Medical Record Review. Int. J. Environ. Res. Public Health 2022, 19, 2445. [Google Scholar] [CrossRef] [PubMed]
- Knurick, J.R.; Johnston, C.S.; Wherry, S.J.; Aguayo, I. Comparison of correlates of bone mineral density in individuals adhering to lacto-ovo, vegan, or omnivore diets: A cross-sectional investigation. Nutrients 2015, 7, 3416–3426. [Google Scholar] [CrossRef] [PubMed]
- Tesar, R.; Notelovitz, M.; Shim, E.; Kauwell, G.; Brown, J. Axial and peripheral bone density and nutrient intakes of postmenopausal vegetarian and omnivorous women. Am. J. Clin. Nutr. 1992, 56, 699–704. [Google Scholar] [CrossRef]
- Chuang, T.L.; Lin, C.H.; Wang, Y.F. Effects of vegetarian diet on bone mineral density. Tzu Chi Med. J. 2021, 33, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Brants, H.A.; Löwik, M.R.; Westenbrink, S.; Hulshof, K.F.; Kistemaker, C. Adequacy of a vegetarian diet at old age (Dutch Nutrition Surveillance System). J. Am. Coll. Nutr. 1990, 9, 292–302. [Google Scholar] [CrossRef] [PubMed]
- Berg, J.; Seyedsadjadi, N.; Grant, R. Increased Consumption of Plant Foods Is Associated with Increased Bone Mineral Density. J. Nutr. Health Aging 2020, 24, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Aleksandrova, K.; Koelman, L.; Rodrigues, C.E. Dietary patterns and biomarkers of oxidative stress and inflammation: A systematic review of observational and intervention studies. Redox Biol. 2021, 42, 101869. [Google Scholar] [CrossRef] [PubMed]
- Sahni, S.; Hannan, M.T.; Blumberg, J.; Cupples, L.A.; Kiel, D.P.; Tucker, K.L. Inverse association of carotenoid intakes with 4-y change in bone mineral density in elderly men and women: The Framingham Osteoporosis Study. Am. J. Clin. Nutr. 2009, 89, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Burckhardt, P. The role of low acid load in vegetarian diet on bone health: A narrative review. Swiss Med. Wkly. 2016, 146, w14277. [Google Scholar] [CrossRef]
- Baker, K.D.; Loughman, A.; Spencer, S.J.; Reichelt, A.C. The impact of obesity and hypercaloric diet consumption on anxiety and emotional behavior across the lifespan. Neurosci. Biobehav. Rev. 2017, 83, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.J.; Nakagawa, T.; Sanchez-Lozada, L.G.; Shafiu, M.; Sundaram, S.; Le, M.; Ishimoto, T.; Sautin, Y.Y.; Lanaspa, M.A. Sugar, uric acid, and the etiology of diabetes and obesity. Diabetes 2013, 62, 3307–3315. [Google Scholar] [CrossRef] [PubMed]
- Kawano, Y.; Edwards, M.; Huang, Y.; Bilate, A.M.; Araujo, L.P.; Tanoue, T.; Atarashi, K.; Ladinsky, M.S.; Reiner, S.L.; Wang, H.H.; et al. Microbiota imbalance induced by dietary sugar disrupts immune-mediated protection from metabolic syndrome. Cell 2022, 185, 3501–3519.e3520. [Google Scholar] [CrossRef] [PubMed]
- Frieler, R.A.; Vigil, T.M.; Song, J.; Leung, C.; Lumeng, C.N.; Mortensen, R.M. High-fat and high-sodium diet induces metabolic dysfunction in the absence of obesity. Obesity 2021, 29, 1868–1881. [Google Scholar] [CrossRef] [PubMed]
- Atak, S.; Boye, A.; Peciña, S.; Liu, Z.X. High-fat-sugar diet is associated with impaired hippocampus-dependent memory in humans. Physiol. Behav. 2023, 268, 114225. [Google Scholar] [CrossRef] [PubMed]
- Tsanzi, E.; Fitch, C.W.; Tou, J.C. Effect of consuming different caloric sweeteners on bone health and possible mechanisms. Nutr. Rev. 2008, 66, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Ahn, H.; Park, Y.K. Sugar-sweetened beverage consumption and bone health: A systematic review and meta-analysis. Nutr. J. 2021, 20, 41. [Google Scholar] [CrossRef] [PubMed]
- Bragança, M.; Bogea, E.G.; de Almeida Fonseca Viola, P.C.; Dos Santos Vaz, J.; Confortin, S.C.; Menezes, A.M.B.; Gonçalves, H.; Bettiol, H.; Barbieri, M.A.; Cardoso, V.C.; et al. High Consumption of Sugar-Sweetened Beverages Is Associated with Low Bone Mineral Density in Young People: The Brazilian Birth Cohort Consortium. Nutrients 2023, 15, 324. [Google Scholar] [CrossRef] [PubMed]
- Bennett, A.M.; Murray, K.; Ambrosini, G.L.; Oddy, W.H.; Walsh, J.P.; Zhu, K. Prospective Associations of Sugar-Sweetened Beverage Consumption During Adolescence with Body Composition and Bone Mass at Early Adulthood. J. Nutr. 2022, 152, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Tjäderhane, L.; Larmas, M. A high sucrose diet decreases the mechanical strength of bones in growing rats. J. Nutr. 1998, 128, 1807–1810. [Google Scholar] [CrossRef] [PubMed]
- Bass, E.F.; Baile, C.A.; Lewis, R.D.; Giraudo, S.Q. Bone quality and strength are greater in growing male rats fed fructose compared with glucose. Nutr. Res. 2013, 33, 1063–1071. [Google Scholar] [CrossRef] [PubMed]
- Tsanzi, E.; Light, H.R.; Tou, J.C. The effect of feeding different sugar-sweetened beverages to growing female Sprague-Dawley rats on bone mass and strength. Bone 2008, 42, 960–968. [Google Scholar] [CrossRef]
- Felice, J.I.; Gangoiti, M.V.; Molinuevo, M.S.; McCarthy, A.D.; Cortizo, A.M. Effects of a metabolic syndrome induced by a fructose-rich diet on bone metabolism in rats. Metabolism 2014, 63, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Minematsu, A.; Nishii, Y.; Sakata, S. High-fat/high-sucrose diet results in higher bone mass in aged rats. Bone Rep. 2018, 8, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Vandewalle, S.; Taes, Y.; Van Helvoirt, M.; Debode, P.; Herregods, N.; Ernst, C.; Roef, G.; Van Caenegem, E.; Roggen, I.; Verhelle, F.; et al. Bone size and bone strength are increased in obese male adolescents. J. Clin. Endocrinol. Metab. 2013, 98, 3019–3028. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Jung, H.W.; Hong, H.; Kim, J.H.; Shin, C.H.; Yang, S.W.; Lee, Y.A. The Role of Overweight and Obesity on Bone Health in Korean Adolescents with a Focus on Lean and Fat Mass. J. Korean Med. Sci. 2017, 32, 1633–1641. [Google Scholar] [CrossRef] [PubMed]
- Leonard, M.B.; Zemel, B.S.; Wrotniak, B.H.; Klieger, S.B.; Shults, J.; Stallings, V.A.; Stettler, N. Tibia and radius bone geometry and volumetric density in obese compared to non-obese adolescents. Bone 2015, 73, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Woo, D.G.; Lee, B.Y.; Lim, D.; Kim, H.S. Relationship between nutrition factors and osteopenia: Effects of experimental diets on immature bone quality. J. Biomech. 2009, 42, 1102–1107. [Google Scholar] [CrossRef] [PubMed]
- Inzana, J.A.; Kung, M.; Shu, L.; Hamada, D.; Xing, L.P.; Zuscik, M.J.; Awad, H.A.; Mooney, R.A. Immature mice are more susceptible to the detrimental effects of high fat diet on cancellous bone in the distal femur. Bone 2013, 57, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Wee, N.K.Y.; Enriquez, R.F.; Nguyen, A.D.; Horsnell, H.; Kulkarni, R.; Khor, E.C.; Herzog, H.; Baldock, P.A. Diet-induced obesity suppresses cortical bone accrual by a neuropeptide Y-dependent mechanism. Int. J. Obes. 2018, 42, 1925–1938. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Yang, X.; Gao, X.; Du, H.; Han, Y.; Zhang, D.; Wang, Z.; Sun, L. Inhibiting myostatin signaling prevents femoral trabecular bone loss and microarchitecture deterioration in diet-induced obese rats. Exp. Biol. Med. 2016, 241, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, Z.; Pei, L.; Zhang, X.; Li, B.; Meng, Y.; Zhou, X. How high-fat diet affects bone in mice: A systematic review and meta-analysis. Obes. Rev. 2022, 23, e13493. [Google Scholar] [CrossRef] [PubMed]
- Doucette, C.R.; Horowitz, M.C.; Berry, R.; MacDougald, O.A.; Anunciado-Koza, R.; Koza, R.A.; Rosen, C.J. A High Fat Diet Increases Bone Marrow Adipose Tissue (MAT) But Does Not Alter Trabecular or Cortical Bone Mass in C57BL/6J Mice. J. Cell Physiol. 2015, 230, 2032–2037. [Google Scholar] [CrossRef]
- Liu, S.Q.; Wu, J.; Mo, J.; Sun, Z.X.; Yang, H.B.; Huang, C.W.; Lei, M.X.; Peng, L.W.; Xu, L. Serum leptin level and its association with bone mineral density in obese children. Zhongguo Dang Dai Er Ke Za Zhi 2009, 11, 745–748. [Google Scholar] [PubMed]
- Głogowska-Szeląg, J.; Kos-Kudła, B.; Marek, B.; Nowak, M.; Siemińska, L. Assessment of selected adipocytokines in obese women with postmenopausal osteoporosis. Endokrynol. Pol. 2019, 70, 478–483. [Google Scholar] [CrossRef] [PubMed]
- Ducy, P.; Amling, M.; Takeda, S.; Priemel, M.; Schilling, A.F.; Beil, F.T.; Shen, J.; Vinson, C.; Rueger, J.M.; Karsenty, G. Leptin inhibits bone formation through a hypothalamic relay: A central control of bone mass. Cell 2000, 100, 197–207. [Google Scholar] [CrossRef] [PubMed]
- López-Gómez, J.J.; Pérez-Castrillón, J.L.; García de Santos, I.; Pérez-Alonso, M.; Izaola-Jauregui, O.; Primo-Martín, D.; De Luis-Román, D.A. Influence of Obesity on Bone Turnover Markers and Fracture Risk in Postmenopausal Women. Nutrients 2022, 14, 1617. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Lee, J.W.; Im, J.A.; Hwang, H.J. Serum osteocalcin is related to abdominal obesity in Korean obese and overweight men. Clin. Chim. Acta 2010, 411, 2054–2057. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; He, C. Pro-inflammatory cytokines: The link between obesity and osteoarthritis. Cytokine Growth Factor Rev. 2018, 44, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Abuna, R.P.; De Oliveira, F.S.; Santos Tde, S.; Guerra, T.R.; Rosa, A.L.; Beloti, M.M. Participation of TNF-α in Inhibitory Effects of Adipocytes on Osteoblast Differentiation. J. Cell Physiol. 2016, 231, 204–214. [Google Scholar] [CrossRef] [PubMed]
- Vranić, L.; Mikolašević, I.; Milić, S. Vitamin D Deficiency: Consequence or Cause of Obesity? Medicina 2019, 55, 541. [Google Scholar] [CrossRef] [PubMed]
- Patsch, J.M.; Kiefer, F.W.; Varga, P.; Pail, P.; Rauner, M.; Stupphann, D.; Resch, H.; Moser, D.; Zysset, P.K.; Stulnig, T.M.; et al. Increased bone resorption and impaired bone microarchitecture in short-term and extended high-fat diet-induced obesity. Metabolism 2011, 60, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Ootsuka, T.; Nakanishi, A.; Tsukamoto, I. Increase in osteoclastogenesis in an obese Otsuka Long-Evans Tokushima fatty rat model. Mol Med Rep. 2015, 12, 3874–3880. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Wei, L.; Xu, W.; Lu, J.; Wang, C.; Bao, Y.; Jia, W. CTSK inhibitor exert its anti-obesity effects through regulating adipocyte differentiation in high-fat diet induced obese mice. Endocr. J. 2015, 62, 309–317. [Google Scholar] [CrossRef]
- Shu, L.; Beier, E.; Sheu, T.; Zhang, H.; Zuscik, M.J.; Puzas, E.J.; Boyce, B.F.; Mooney, R.A.; Xing, L. High-fat diet causes bone loss in young mice by promoting osteoclastogenesis through alteration of the bone marrow environment. Calcif. Tissue Int. 2015, 96, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Oshima, K.; Nampei, A.; Matsuda, M.; Iwaki, M.; Fukuhara, A.; Hashimoto, J.; Yoshikawa, H.; Shimomura, I. Adiponectin increases bone mass by suppressing osteoclast and activating osteoblast. Biochem. Biophys. Res. Commun. 2005, 331, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.P.; Li, X.P.; Wang, M.; Zhao, L.L.; Li, H.; Xie, H.; Lu, Z.Y. Adiponectin exerts its negative effect on bone metabolism via OPG/RANKL pathway: An in vivo study. Endocrine 2014, 47, 845–853. [Google Scholar] [CrossRef] [PubMed]
- Montalvany-Antonucci, C.C.; Zicker, M.C.; Ferreira, A.V.M.; Macari, S.; Ramos-Junior, E.S.; Gomez, R.S.; Pereira, T.S.F.; Madeira, M.F.M.; Fukada, S.Y.; Andrade, I., Jr.; et al. High-fat diet disrupts bone remodeling by inducing local and systemic alterations. J. Nutr. Biochem. 2018, 59, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Xu, P.; Wang, C.; Ha, X.; Gu, Y.; Wang, Y.; Zhang, J.; Xie, J. The effects of fat-induced obesity on bone metabolism in rats. Obes. Res. Clin. Pract. 2017, 11, 454–463. [Google Scholar] [CrossRef] [PubMed]
- Cai, F.; Yusufu, A.; Liu, K.; Chen, W.; Zhao, R.; Liu, Y.; Liu, Y. High-fat diet causes undesirable bone regeneration by altering the bone marrow environment in rats. Front. Endocrinol. 2023, 14, 1088508. [Google Scholar] [CrossRef] [PubMed]
- Cortez, M.; Carmo, L.S.; Rogero, M.M.; Borelli, P.; Fock, R.A. A high-fat diet increases IL-1, IL-6, and TNF-α production by increasing NF-κB and attenuating PPAR-γ expression in bone marrow mesenchymal stem cells. Inflammation 2013, 36, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Steppan, C.M.; Lazar, M.A. Resistin and obesity-associated insulin resistance. Trends Endocrinol. Metab. 2002, 13, 18–23. [Google Scholar] [CrossRef] [PubMed]
- De Benedetti, F.; Rucci, N.; Del Fattore, A.; Peruzzi, B.; Paro, R.; Longo, M.; Vivarelli, M.; Muratori, F.; Berni, S.; Ballanti, P.; et al. Impaired skeletal development in interleukin-6-transgenic mice: A model for the impact of chronic inflammation on the growing skeletal system. Arthritis Rheum. 2006, 54, 3551–3563. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Cui, J.; Li, Y.X.; Shi, Y.H.; Wang, B.; Le, G.W.; Wang, Z.P. Dyslipidemic high-fat diet affects adversely bone metabolism in mice associated with impaired antioxidant capacity. Nutrition 2011, 27, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Zengin, A.; Kropp, B.; Chevalier, Y.; Junnila, R.; Sustarsic, E.; Herbach, N.; Fanelli, F.; Mezzullo, M.; Milz, S.; Bidlingmaier, M.; et al. Low-carbohydrate, high-fat diets have sex-specific effects on bone health in rats. Eur. J. Nutr. 2016, 55, 2307–2320. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.L.; Li, C.M.; Cao, S.S.; Zhou, L.P.; Wong, M.S. A High-Saturated-Fat, High-Sucrose Diet Aggravates Bone Loss in Ovariectomized Female Rats. J. Nutr. 2016, 146, 1172–1179. [Google Scholar] [CrossRef] [PubMed]
- Zernicke, R.F.; Salem, G.J.; Barnard, R.J.; Schramm, E. Long-term, high-fat-sucrose diet alters rat femoral neck and vertebral morphology, bone mineral content, and mechanical properties. Bone 1995, 16, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Wang, C.; Xie, Y.; Wan, S.; Zhang, K.; Yu, X. High Fructose and High Fat Exert Different Effects on Changes in Trabecular Bone Micro-structure. J. Nutr. Health Aging 2018, 22, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Jatkar, A.; Kurland, I.J.; Judex, S. Diets High in Fat or Fructose Differentially Modulate Bone Health and Lipid Metabolism. Calcif. Tissue Int. 2017, 100, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Lorincz, C.; Reimer, R.A.; Boyd, S.K.; Zernicke, R.F. High-fat, sucrose diet impairs geometrical and mechanical properties of cortical bone in mice. Br. J. Nutr. 2010, 103, 1302–1308. [Google Scholar] [CrossRef] [PubMed]
- Li, K.C.; Zernicke, R.F.; Barnard, R.J.; Li, A.F. Effects of a high fat-sucrose diet on cortical bone morphology and biomechanics. Calcif. Tissue Int. 1990, 47, 308–313. [Google Scholar] [CrossRef]
- World Health Organization. Protein and Amino Acid Requirements in Human Nutrition; World Health Organization: Geneva, Switzerland, 2007; pp. 1–265. [Google Scholar]
- Cao, J.J. High Dietary Protein Intake and Protein-Related Acid Load on Bone Health. Curr. Osteoporos. Rep. 2017, 15, 571–576. [Google Scholar] [CrossRef]
- Shams-White, M.M.; Chung, M.; Du, M.; Fu, Z.; Insogna, K.L.; Karlsen, M.C.; LeBoff, M.S.; Shapses, S.A.; Sackey, J.; Wallace, T.C.; et al. Dietary protein and bone health: A systematic review and meta-analysis from the National Osteoporosis Foundation. Am. J. Clin. Nutr. 2017, 105, 1528–1543. [Google Scholar] [CrossRef]
- Antonio, J.; Ellerbroek, A.; Evans, C.; Silver, T.; Peacock, C.A. High protein consumption in trained women: Bad to the bone? J. Int. Soc. Sports Nutr. 2018, 15, 6. [Google Scholar] [CrossRef] [PubMed]
- Calvez, J.; Poupin, N.; Chesneau, C.; Lassale, C.; Tomé, D. Protein intake, calcium balance and health consequences. Eur. J. Clin. Nutr. 2012, 66, 281–295. [Google Scholar] [CrossRef] [PubMed]
- Kerstetter, J.E.; Caseria, D.M.; Mitnick, M.E.; Ellison, A.F.; Gay, L.F.; Liskov, T.A.; Carpenter, T.O.; Insogna, K.L. Increased circulating concentrations of parathyroid hormone in healthy, young women consuming a protein-restricted diet. Am. J. Clin. Nutr. 1997, 66, 1188–1196. [Google Scholar] [CrossRef] [PubMed]
- Langdahl, B.L.; Kassem, M.; Møller, M.K.; Eriksen, E.F. The effects of IGF-I and IGF-II on proliferation and differentiation of human osteoblasts and interactions with growth hormone. Eur. J. Clin. Investig. 1998, 28, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Dawson-Hughes, B.; Harris, S.S.; Rasmussen, H.; Song, L.; Dallal, G.E. Effect of dietary protein supplements on calcium excretion in healthy older men and women. J. Clin. Endocrinol. Metab. 2004, 89, 1169–1173. [Google Scholar] [CrossRef] [PubMed]
- Tirapegui, J.; Ribeiro, S.M.; Pires, I.S.; Rogero, M.M. Effects of two different levels of dietary protein on body composition and protein nutritional status of growing rats. Nutrients 2012, 4, 1328–1337. [Google Scholar] [CrossRef]
- Nebot, E.; Aparicio, V.A.; Coll-Risco, I.; Camiletti-Moirón, D.; Schneider, J.; Kapravelou, G.; Heimel, P.; Martínez, R.; Andrade, A.; Slezak, P.; et al. Effects of a moderately high-protein diet and interval aerobic training combined with strength-endurance exercise on markers of bone metabolism, microarchitecture and turnover in obese Zucker rats. Bone 2016, 92, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Kerstetter, J.E.; O’Brien, K.O.; Insogna, K.L. Dietary protein, calcium metabolism, and skeletal homeostasis revisited. Am. J. Clin. Nutr. 2003, 78, 584s–592s. [Google Scholar] [CrossRef] [PubMed]
- Matikainen, N.; Pekkarinen, T.; Ryhänen, E.M.; Schalin-Jäntti, C. Physiology of Calcium Homeostasis: An Overview. Endocrinol. Metab. Clin. N. Am. 2021, 50, 575–590. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Hu, J.; Kong, X.; Zhu, Z. Association between Dietary Calcium Intake and Bone Mineral Density in Older Adults. Ecol. Food Nutr. 2021, 60, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Closa-Monasterolo, R.; Zaragoza-Jordana, M.; Ferré, N.; Luque, V.; Grote, V.; Koletzko, B.; Verduci, E.; Vecchi, F.; Escribano, J. Adequate calcium intake during long periods improves bone mineral density in healthy children. Data from the Childhood Obesity Project. Clin. Nutr. 2018, 37, 890–896. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.M.; Huang, Z.W.; Yang, X.G.; Su, Y.X. Calcium supplementation and bone mineral accretion in Chinese adolescents aged 12–14 years: A 12-month, dose-response, randomised intervention trial. Br. J. Nutr. 2014, 112, 1510–1520. [Google Scholar] [CrossRef] [PubMed]
- Vannucci, L.; Fossi, C.; Quattrini, S.; Guasti, L.; Pampaloni, B.; Gronchi, G.; Giusti, F.; Romagnoli, C.; Cianferotti, L.; Marcucci, G.; et al. Calcium Intake in Bone Health: A Focus on Calcium-Rich Mineral Waters. Nutrients 2018, 10, 1930. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.M.; Sawyer, R.K.; Moore, A.J.; Morris, H.A.; O’Loughlin, P.D.; Anderson, P.H. Adequate dietary vitamin D and calcium are both required to reduce bone turnover and increased bone mineral volume. J. Steroid Biochem. Mol. Biol. 2014, 144 Pt A, 159–162. [Google Scholar] [CrossRef]
- Carmeliet, G.; Dermauw, V.; Bouillon, R. Vitamin D signaling in calcium and bone homeostasis: A delicate balance. Best Pract. Res. Clin. Endocrinol. Metab. 2015, 29, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Kuang, X.; Li, K.; Guo, X.; Deng, Q.; Li, D. Effects of combined calcium and vitamin D supplementation on osteoporosis in postmenopausal women: A systematic review and meta-analysis of randomized controlled trials. Food Funct. 2020, 11, 10817–10827. [Google Scholar] [CrossRef] [PubMed]
- Bolland, M.J.; Leung, W.; Tai, V.; Bastin, S.; Gamble, G.D.; Grey, A.; Reid, I.R. Calcium intake and risk of fracture: Systematic review. BMJ 2015, 351, h4580. [Google Scholar] [CrossRef] [PubMed]
- Rizzoli, R. Dairy products and bone health. Aging Clin. Exp. Res. 2022, 34, 9–24. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, A.E.; Zawada, A.; Rychter, A.M.; Dobrowolska, A.; Krela-Kaźmierczak, I. Milk and Dairy Products: Good or Bad for Human Bone? Practical Dietary Recommendations for the Prevention and Management of Osteoporosis. Nutrients 2021, 13, 1329. [Google Scholar] [CrossRef]
- van den Heuvel, E.; Steijns, J. Dairy products and bone health: How strong is the scientific evidence? Nutr. Res. Rev. 2018, 31, 164–178. [Google Scholar] [CrossRef] [PubMed]
- Li, B.Y.; Mahe, J.L.; Hao, J.Y.; Ye, W.H.; Bai, X.F.; Feng, H.T.; Szeto, I.M.; Jing, L.P.; Zhao, Z.F.; Chen, Y.M. Formula Milk Supplementation and Bone Acquisition in 4-6 Years Chinese Children: A 12-Month Cluster-Randomized Controlled Trial. Nutrients 2023, 15, 2012. [Google Scholar] [CrossRef] [PubMed]
- Gui, J.C.; Brašić, J.R.; Liu, X.D.; Gong, G.Y.; Zhang, G.M.; Liu, C.J.; Gao, G.Q. Bone mineral density in postmenopausal Chinese women treated with calcium fortification in soymilk and cow’s milk. Osteoporos. Int. 2012, 23, 1563–1570. [Google Scholar] [CrossRef] [PubMed]
- Michaëlsson, K.; Wolk, A.; Langenskiöld, S.; Basu, S.; Warensjö Lemming, E.; Melhus, H.; Byberg, L. Milk intake and risk of mortality and fractures in women and men: Cohort studies. BMJ 2014, 349, g6015. [Google Scholar] [CrossRef] [PubMed]
- Salari, N.; Darvishi, N.; Bartina, Y.; Larti, M.; Kiaei, A.; Hemmati, M.; Shohaimi, S.; Mohammadi, M. Global prevalence of osteoporosis among the world older adults: A comprehensive systematic review and meta-analysis. J. Orthop. Surg. Res. 2021, 16, 669. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Butler, L.M.; van Dam, R.M.; Ang, L.W.; Yuan, J.M.; Koh, W.P. Adherence to a vegetable-fruit-soy dietary pattern or the Alternative Healthy Eating Index is associated with lower hip fracture risk among Singapore Chinese. J. Nutr. 2014, 144, 511–518. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Protocol | |
|---|---|
| Alternate-Day Fasting | Alternate fasting days and eating days. |
| Modified Fasting Regimens | Fasting days allow for 20–25% of daily energy requirements, alternating with feeding days. |
| Time-Restricted Feeding | Consume random energy intake at specific times of the day and prohibit the intake of calorie-containing foods and beverages at specific times. |
| Ramadan Fasting | Followers of some religions must fast from sunrise to sunset during Ramadan. |
| Types of Intermittent Fasting | Subjects | Effects on Bone | Reference |
|---|---|---|---|
| 96 h of single fasting | 6-week-old male Wister rats | Lumbar vertebra BMD ↓ Bone strength index minimum cross-sectional moment, polar moment ↓ | [16] |
| 16–18 h of fasting per day | Rats with glucocorticoid-induced osteoporosis | Serum OPG, ALP, OCN ↑ TRAP-5b, NTX-1, DPD ↓ | [17] |
| Alternate-day fasting | 6-week-old male Sprague Dawley rats + ketogenic diet | BMD ↑ Serum TRAP ↓ | [18] |
| Types of Intermittent Fasting | Gender (M/F) | Age (Year) | Subjects | Effects on Bone | Reference |
|---|---|---|---|---|---|
| Ramadan fasting | 18/5 | 18–42 | Saudi Arabian Muslims | Serum PTH during Ramadan fasting ↓. | [19] |
| Ramadan fasting | 143/257 | 24–58 | People with and without Ramadan fasting habits | 25-hydroxyvitamin D concentrations were higher in winter and spring fasters than in non-fasters ↓. | [22] |
| Alternate-day fasting | 8/54 | 18–65 | Overweight and obese people | BMC, BMD, serum CTX-1, OPG showed no significant difference. | [23] |
| 24 h of single fasting | 8/8 | 19–25 | Healthy subjects | Serum CTX-1, P1NP, PTH showed no significant difference. | [24] |
| 8 h food restriction | 10/12 | 55–79 | Healthy elderly subjects | No significant effect on BMD. | [25] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Wu, Y.; Bennett, S.; Zou, J.; Xu, J.; Zhang, L. The Effects of Different Dietary Patterns on Bone Health. Nutrients 2024, 16, 2289. https://doi.org/10.3390/nu16142289
Liu X, Wu Y, Bennett S, Zou J, Xu J, Zhang L. The Effects of Different Dietary Patterns on Bone Health. Nutrients. 2024; 16(14):2289. https://doi.org/10.3390/nu16142289
Chicago/Turabian StyleLiu, Xiaohua, Yangming Wu, Samuel Bennett, Jun Zou, Jiake Xu, and Lingli Zhang. 2024. "The Effects of Different Dietary Patterns on Bone Health" Nutrients 16, no. 14: 2289. https://doi.org/10.3390/nu16142289
APA StyleLiu, X., Wu, Y., Bennett, S., Zou, J., Xu, J., & Zhang, L. (2024). The Effects of Different Dietary Patterns on Bone Health. Nutrients, 16(14), 2289. https://doi.org/10.3390/nu16142289