
Chemical Composition and Neuroprotective Properties of Indonesian Stingless Bee (Geniotrigona thoracica) Propolis Extract in an In-Vivo Model of Intracerebral Hemorrhage (ICH)

 , , , ,
, , , ,  , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Propolis Extract Preparation
2.2. Total Phenolics and Flavonoids of the Propolis Extract
2.3. GC-MS Analyses of the Glyceric Extract of Propolis
2.4. In-Vivo Study
2.4.1. The ICH Model
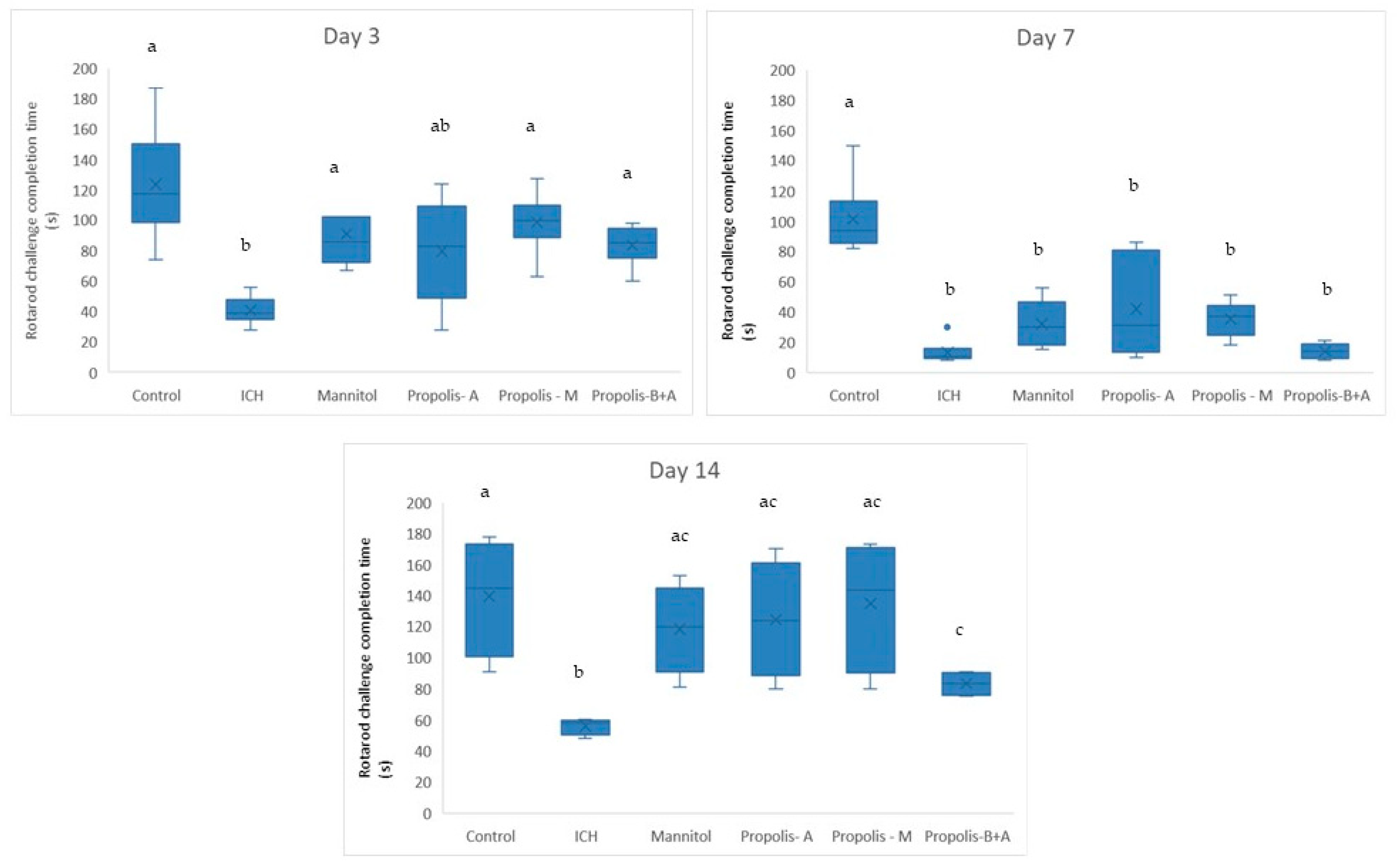
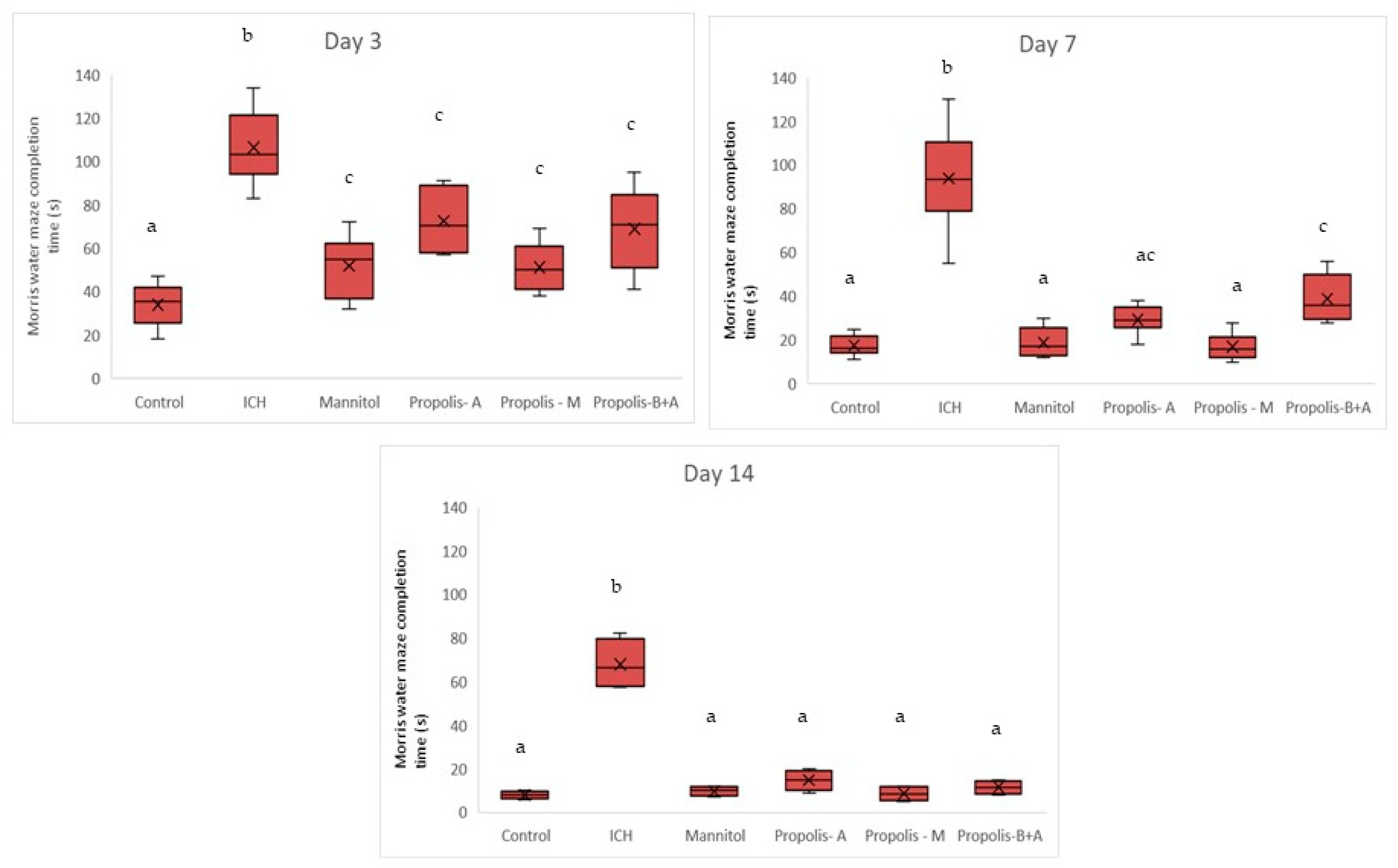
2.4.2. Assessment of Cognitive–Motor Functions
2.4.3. Cerebral Edema Analyses
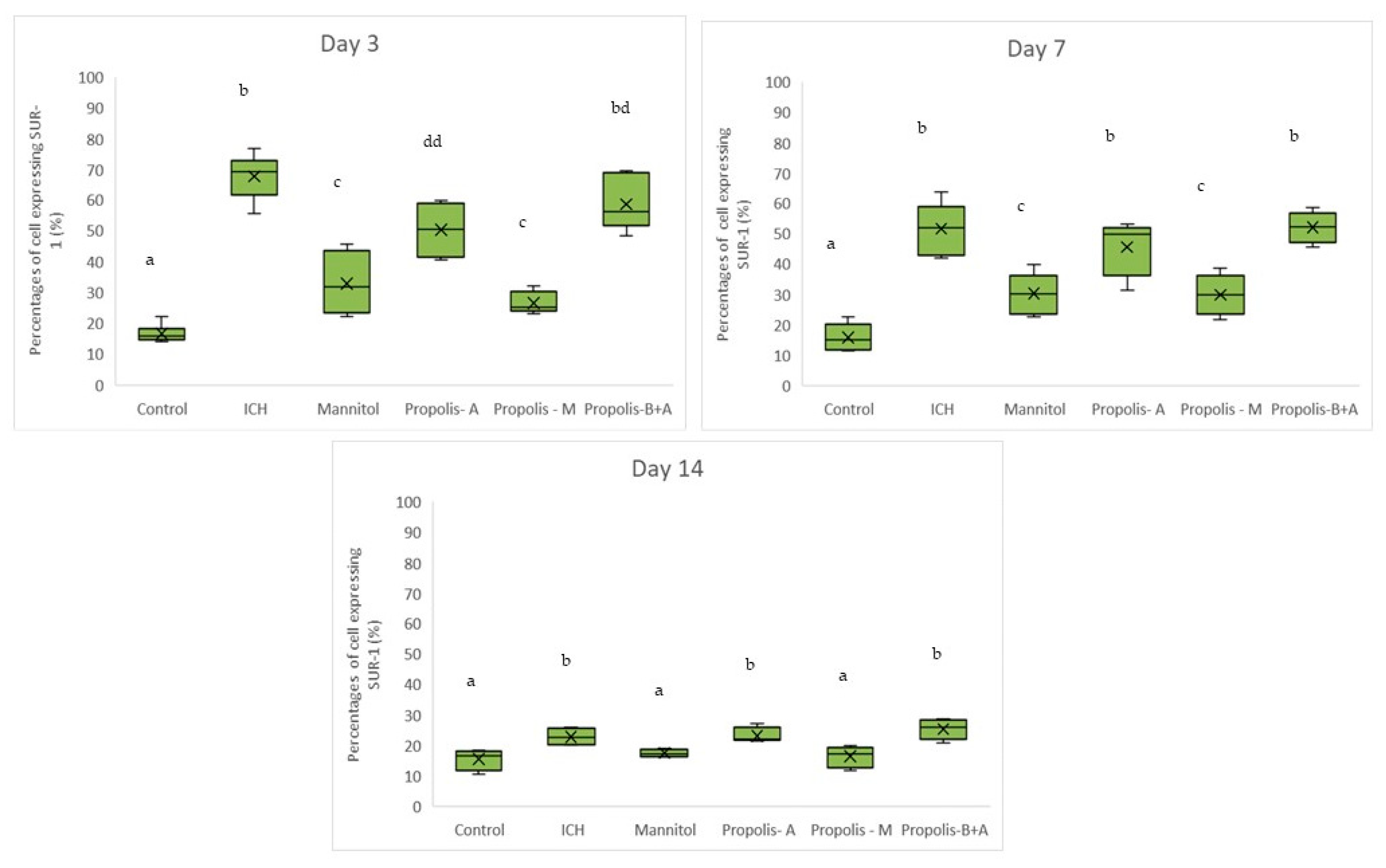
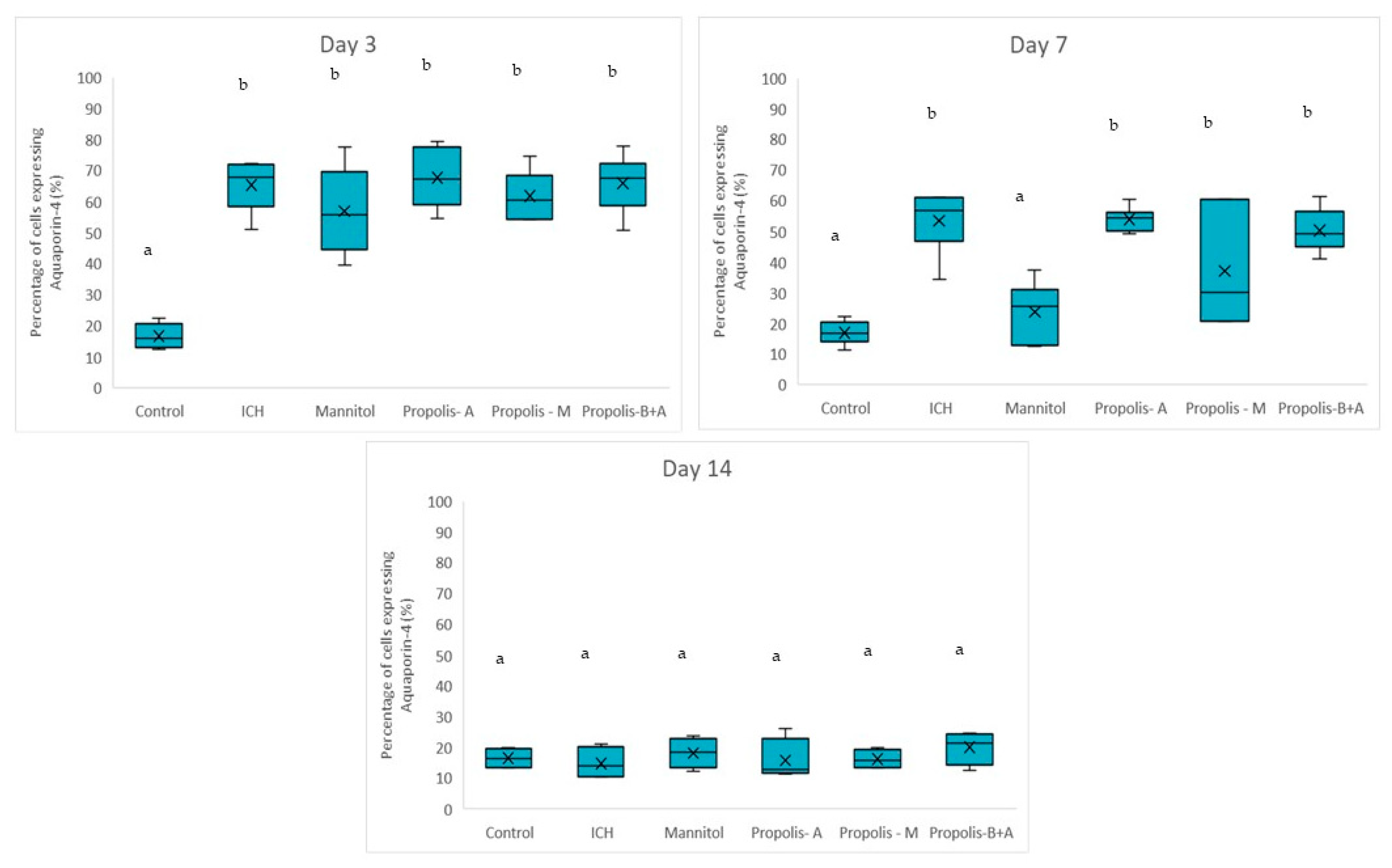
2.4.4. Immunohistochemical Analyses
2.4.5. TUNEL Assay
2.5. Statistical Analysis
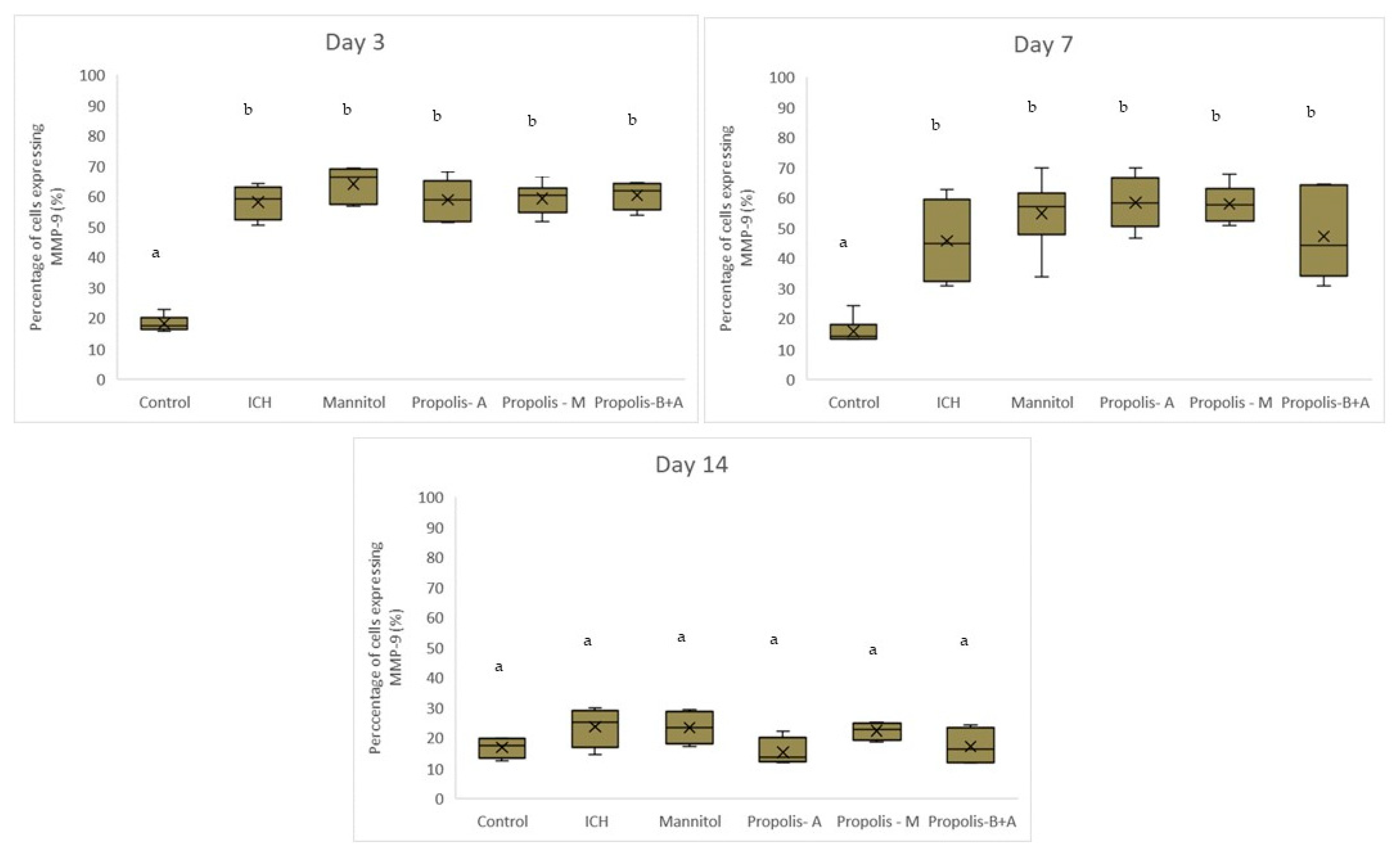
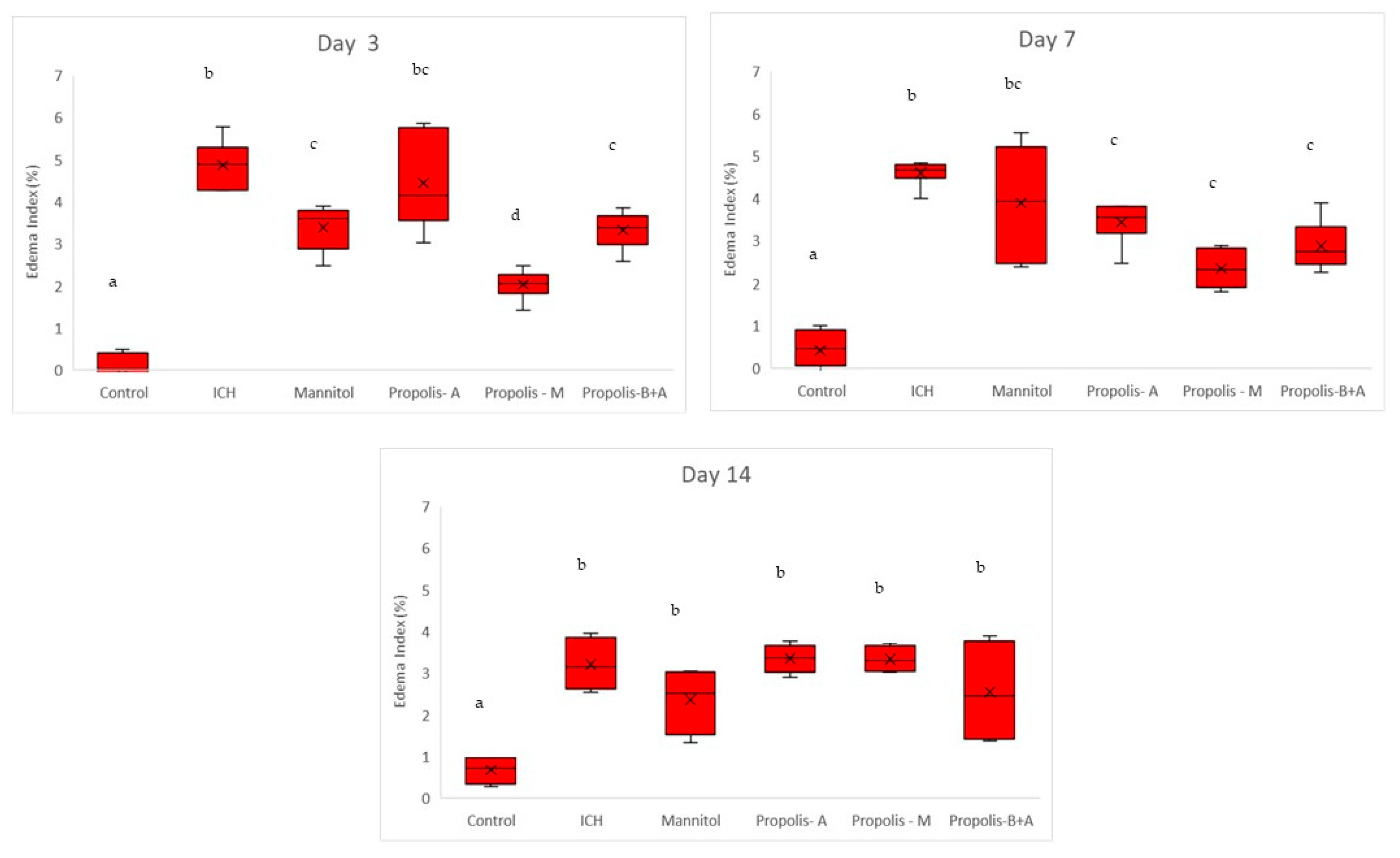
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Virani, S.S.; Alonso, A.; Aparicio, H.J.; Benjamin, E.J.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Cheng, S.; Delling, F.N.; et al. Heart Disease and Stroke Statistics—2021 Update: A Report From the American Heart Association. Circulation 2021, 143, E254–E743. [Google Scholar] [CrossRef] [PubMed]
- Montaño, A.; Hanley, D.F.; Hemphill, J.C. Hemorrhagic Stroke. Handb. Clin. Neurol. 2021, 176, 229–248. [Google Scholar] [CrossRef] [PubMed]
- Selim, M. Basic and Translational Research in Intracerebral Hemorrhage: Limitations, Priorities, and Recommendations. Stroke 2018, 49, 1308–1314. [Google Scholar] [CrossRef] [PubMed]
- Venkatasubramanian, C.; Mlynash, M.; Finley-Caulfield, A.; Eyngorn, I.; Kalimuthu, R.; Snider, R.W.; Wijman, C.A. Natural History of Perihematomal Edema after Intracerebral Hemorrhage Measured by Serial Magnetic Resonance Imaging. Stroke 2011, 42, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Rymer, M.M. Hemorrhagic Stroke: Intracerebral Hemorrhage. Mo. Med. 2011, 108, 50–54. [Google Scholar] [PubMed]
- Juvela, S.; Kase, C.S. Advances in Intracerebral Hemorrhage Management. Stroke 2006, 37, 301–304. [Google Scholar] [CrossRef] [PubMed]
- Brott, T.; Broderick, J.; Kothari, R.; Barsan, W.; Tomsick, T.; Sauerbeck, L.; Spilker, J.; Duldner, J.; Khoury, J. Early Hemorrhage Growth in Patients with Intracerebral Hemorrhage. Stroke 1997, 28, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Chen, C.; Zhang, J.; Hu, Z. Mechanism and Therapy of Brain Edema after Intracerebral Hemorrhage. Cerebrovasc. Dis. 2016, 42, 155–169. [Google Scholar] [CrossRef] [PubMed]
- Rapoport, S.I. Osmotic Opening of the Blood-Brain Barrier: Principles, Mechanism, and Therapeutic Applications. Cell. Mol. Neurobiol. 2000, 20, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Carney, N.; Totten, A.M.; O’Reilly, C.; Ullman, J.S.; Hawryluk, G.W.J.; Bell, M.J.; Bratton, S.L.; Chesnut, R.; Harris, O.A.; Kissoon, N.; et al. Guidelines for the Management of Severe TBI. Neurosurgery 2017, 80, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Lesmana, R.; Zulhendri, F.; Fearnley, J.; Irsyam, I.A.; Rasyid, R.P.H.N.; Abidin, T.; Abdulah, R.; Suwantika, A.; Paradkar, A.; Budiman, A.S.; et al. The Suitability of Propolis as a Bioactive Component of Biomaterials. Front. Pharmacol. 2022, 13, 930515. [Google Scholar] [CrossRef] [PubMed]
- Zulhendri, F.; Lesmana, R.; Tandean, S.; Christoper, A.; Chandrasekaran, K.; Irsyam, I.; Suwantika, A.A.; Abdulah, R.; Wathoni, N. Recent Update on the Anti-Inflammatory Activities of Propolis. Molecules 2022, 27, 8473. [Google Scholar] [CrossRef] [PubMed]
- Zulhendri, F.; Chandrasekaran, K.; Kowacz, M.; Ravalia, M.; Kripal, K.; Fearnley, J.; Perera, C.O. Antiviral, Antibacterial, Antifungal, and Antiparasitic Properties of Propolis: A Review. Foods 2021, 10, 1360. [Google Scholar] [CrossRef] [PubMed]
- Safari, M.; Sameni, H.R.H.R.; Badban, L.; Bandegi, A.R.A.R.; Vafaei, A.A.A.A.; Pour, A.R.A.R.; Ghahari, L. Protective Effects of Water Extract of Propolis on Dopaminergic Neurons, Brain Derived Neurotrophic Factor and Stress Oxidative Factors in the Rat Model of Parkinson’s Disease. Int. J. Pharmacol. 2015, 11, 300–308. [Google Scholar] [CrossRef]
- Barros Silva, R.; Santos, N.A.G.A.G.; Martins, N.M.M.; Ferreira, D.A.S.A.S.; Barbosa, F.; Oliveira Souza, V.C.C.; Kinoshita, Â.; Baffa, O.; Del-Bel, E.; Santos, A.C.C. Caffeic Acid Phenethyl Ester Protects against the Dopaminergic Neuronal Loss Induced by 6-Hydroxydopamine in Rats. Neuroscience 2013, 233, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Asem, N.; Gapar, N.A.A.; Abd Hapit, N.H.; Omar, E.A. Correlation between Total Phenolic and Flavonoid Contents with Antioxidant Activity of Malaysian Stingless Bee Propolis Extract. J. Apic. Res. 2020, 59, 437–442. [Google Scholar] [CrossRef]
- Bobinger, T.; Burkardt, P.; Huttner, H.B.; Manaenko, A. Programmed Cell Death after Intracerebral Hemorrhage. Curr. Neuropharmacol. 2018, 16, 1267–1281. [Google Scholar] [CrossRef] [PubMed]
- Rusche, B. The 3Rs and Animal Welfare—Conflict or the Way Forward? ALTEX Altern. Tierexp. 2003, 20, 63–76. [Google Scholar]
- Paiva, W.; Zippo, E.; Miranda, C.; Brasil, S.; Godoy, D.; De Andrade, A.; Neville, I.; Patriota, G.; Domingues, R.; Teixeira, M. Animal Models for the Study of Intracranial Hematomas (Review). Exp. Ther. Med. 2022, 25, 20. [Google Scholar] [CrossRef] [PubMed]
- Doi, T.; Fan, Y.; Gold, J.I.; Ding, L. The Caudate Nucleus Contributes Causally to Decisions That Balance Reward and Uncertain Visual Information. Elife 2020, 9, e56694. [Google Scholar] [CrossRef] [PubMed]
- Ruan, J.; Yao, Y. Behavioral Tests in Rodent Models of Stroke. Brain Hemorrhages 2020, 1, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Michinaga, S.; Koyama, Y. Pathogenesis of Brain Edema and Investigation into Anti-Edema Drugs. Int. J. Mol. Sci. 2015, 16, 9949–9975. [Google Scholar] [CrossRef] [PubMed]
- Katada, R.; Nishitani, Y.; Honmou, O.; Mizuo, K.; Okazaki, S.; Tateda, K.; Watanabe, S. Expression of Aquaporin-4 Augments Cytotoxic Brain Edema after Traumatic Brain Injury during Acute Ethanol Exposure. Am. J. Pathol. 2012, 180, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Miłek, M.; Ciszkowicz, E.; Tomczyk, M.; Sidor, E.; Zaguła, G.; Lecka-Szlachta, K.; Pasternakiewicz, A.; Dżugan, M. The Study of Chemical Profile and Antioxidant Properties of Poplar-Type Polish Propolis Considering Local Flora Diversity in Relation to Antibacterial and Anticancer Activities in Human Breast Cancer Cells. Molecules 2022, 27, 725. [Google Scholar] [CrossRef] [PubMed]
- Tumbarski, Y.; Todorova, M.; Topuzova, M.; Gineva, G.; Yanakieva, V.; Ivanov, I.; Petkova, N. Comparative Study on Physicochemical, Antioxidant and Antimicrobial Properties of Propolis Collected from Different Regions of Bulgaria. J. Apic. Sci. 2023, 67, 37–56. [Google Scholar] [CrossRef]
- Halket, J.M.; Zaikin, V.G. Derivatization in Mass Spectrometry—1. Silylation. Eur. J. Mass Spectrom. 2003, 9, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Popova, M.; Trusheva, B.; Ilieva, N.; Thanh, L.N.; Lien, N.T.P.; Bankova, V. Mangifera indica as Propolis Source: What Exactly Do Bees Collect? BMC Res. Notes 2021, 14, 448. [Google Scholar] [CrossRef] [PubMed]
- Drescher, N.; Wallace, H.M.; Katouli, M.; Massaro, C.F.; Leonhardt, S.D. Diversity Matters: How Bees Benefit from Different Resin Sources. Oecologia 2014, 176, 943–953. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, K.; Silva, A.S.; Atanassova, M.; Sharma, R.; Nepovimova, E.; Musilek, K.; Sharma, R.; Alghuthaymi, M.A.; Dhanjal, D.S.; Nicoletti, M.; et al. Conifers Phytochemicals: A Valuable Forest with Therapeutic Potential. Molecules 2021, 26, 3005. [Google Scholar] [CrossRef] [PubMed]
- Meshginfar, N.; Tavakoli, H.; Dornan, K.; Hosseinian, F. Phenolic Lipids as Unique Bioactive Compounds: A Comprehensive Review on Their Multifunctional Activity toward the Prevention of Alzheimer’s Disease. Crit. Rev. Food Sci. Nutr. 2021, 61, 1394–1403. [Google Scholar] [CrossRef] [PubMed]
- Günday, M.; Saritaş, Z.K.; Demirel, H.H.; Bülbül, A.; Saritaş, T.B.; Görücü, F.; Becit, N. Does Anzer Propolis Have a Protective Effect on Rabbit Spinal Cord Ischemia/Reperfusion Injury? Braz. J. Cardiovasc. Surg. 2021, 37, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Tandean, S.; Japardi, I.; Loe, M.L.M.L.M.L.; Riawan, W.; July, J. Protective Effects of Propolis Extract in a Rat Model of Traumatic Brain Injury via Hsp70 Induction. Open Access Maced. J. Med. Sci. 2019, 7, 2763–2766. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Z.; Du, H.; Luo, X.-L.; Liu, R.-S.; Huang, L.; Cao, C.-S. Caffeic Acid Phenethyl Ester Inhibits the Progression of Elastase Induced Aortic Aneurysm in Rats. Int. J. Pharmacol. 2019, 15, 385–393. [Google Scholar] [CrossRef]
- Bazmandegan, G.; Boroushaki, M.T.; Shamsizadeh, A.; Ayoobi, F.; Hakimizadeh, E.; Allahtavakoli, M. Brown Propolis Attenuates Cerebral Ischemia-Induced Oxidative Damage via Affecting Antioxidant Enzyme System in Mice. Biomed. Pharmacother. 2017, 85, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Durak, M.A.; Öztanir, M.N.; Başak Türkmen, N.; Çiftçi, O.; Taşlidere, A.; Tecellioğlu, M.; Önder, A. Chrysin Prevents Brain Damage Caused by Global Cerebral Ischemia/Reperfusion in a C57BL/J6 Mouse Model. Turk. J. Med. Sci. 2016, 46, 1926–1933. [Google Scholar] [CrossRef] [PubMed]
- Bazmandegan, G.; Shamsizadeh, A.; FathiNajafi, M.; Assadollahi, Z.; Allahtavakoli, M.; Kamiab, Z.; Vakilian, A.; Moghadam-Ahmadi, A.; Amirteimoury, M.; Boroushaki, M.T.M.T. Iranian Brown Propolis Possesses Neuroprotective Effect against Ischemic Neuronal Damage in Mice. J. HerbMed Pharmacol. 2020, 9, 121–129. [Google Scholar] [CrossRef]
- Zulhendri, F.; Perera, C.O.; Tandean, S. Can Propolis Be a Useful Adjuvant in Brain and Neurological Disorders and Injuries? A Systematic Scoping Review of the Latest Experimental Evidence. Biomedicines 2021, 9, 1227. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Chang, H.S.; Su, S.K.; Sun, L.P.; Wang, K. Antioxidative and Anti-Inflammatory Activities of Ethanol Extract of Geopropolis from Stingless Bees. Sci. Agric. Sin. 2019, 52, 939–948. [Google Scholar] [CrossRef]
- Zhu, A.; Wu, Z.; Zhong, X.; Ni, J.; Li, Y.; Meng, J.; Du, C.; Zhao, X.; Nakanishi, H.; Wu, S. Brazilian Green Propolis Prevents Cognitive Decline into Mild Cognitive Impairment in Elderly People Living at High Altitude. J. Alzheimer’s Dis. 2018, 63, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Asama, T.; Hiraoka, T.; Ohkuma, A.; Okumura, N.; Yamaki, A.; Urakami, K. Cognitive Improvement and Safety Assessment of a Dietary Supplement Containing Propolis Extract in Elderly Japanese: A Placebo-Controlled, Randomized, Parallel-Group, Double-Blind Human Clinical Study. Evid.-Based Complement. Altern. Med. 2021, 2021, 6664217. [Google Scholar] [CrossRef] [PubMed]
- Takashima, M.; Ichihara, K.; Hirata, Y. Neuroprotective Effects of Brazilian Green Propolis on Oxytosis/Ferroptosis in Mouse Hippocampal HT22 Cells. Food Chem. Toxicol. 2019, 132, 110669. [Google Scholar] [CrossRef] [PubMed]
- Nosratiyan, M.; Farjah, G.H.; Karimipour, M.; Pourheidar, B. The Effect of Propolis-Gum Arabic as a Novel Nerve Guidance Channel on Regeneration of Sciatic Nerve in Male Rats. Turk. Neurosurg. 2021, 31, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Shao, Q.; Zhao, M.; Pei, W.; Pu, Y.; Liu, M.; Liu, W.; Yu, Z.; Chen, K.; Liu, H.; Deng, B.; et al. Pinocembrin Promotes OPC Differentiation and Remyelination via the MTOR Signaling Pathway. Neurosci. Bull. 2021, 37, 1314–1324. [Google Scholar] [CrossRef] [PubMed]
- El-Guendouz, S.; Aazza, S.; Lyoussi, B.; Antunes, M.D.; Faleiro, M.L.; Miguel, M.G. Anti-Acetylcholinesterase, Antidiabetic, Anti-Inflammatory, Antityrosinase and Antixanthine Oxidase Activities of Moroccan Propolis. Int. J. Food Sci. Technol. 2016, 51, 1762–1773. [Google Scholar] [CrossRef]
- Do Amaral, L.; Caldas, G.R.; dos Santos, N.A.G.; Parreira, R.L.T.; Bastos, J.K.; dos Santos, A.C. Baccharin from Brazilian Green Propolis Induces Neurotrophic Signaling Pathways in PC12 Cells: Potential for Axonal and Synaptic Regeneration. Naunyn-Schmiedeb. Arch. Pharmacol. 2022, 395, 659–672. [Google Scholar] [CrossRef] [PubMed]
- Menezes da Silveira, C.C.S.; Luz, D.A.; da Silva, C.C.S.; Prediger, R.D.S.; Martins, M.D.; Martins, M.A.T.; Fontes-Júnior, E.A.; Maia, C.S.F. Propolis: A Useful Agent on Psychiatric and Neurological Disorders? A Focus on CAPE and Pinocembrin Components. Med. Res. Rev. 2021, 41, 1195–1215. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound 1 | % of TIC 2 | M+∙ | Prominent MS Peaks 3, m/z |
|---|---|---|---|
| Aromatic aldehydes, aromatic acids and their esters | 1.1 | ||
| Vanillin | 0.3 | 224 | 209, 194 (100) |
| Cinnamic acid | 0.2 | 220 | 205, 161 (100), 145, 131, 103, 75 |
| p-Hydroxybenzoic acid | 0.1 | 282 | 267 (100), 223, 193 |
| p-Methoxyphenylacetic acid | 0.1 | 238 | 223, 208, 193 (100) |
| Vanillic acid | 0.2 | 312 | 297, 267, 194, 105 (100), 91, 77 |
| Protocatechuic acid | 0.2 | 370 | 355 (100), 311 |
| Diterpenes | 2.3 | ||
| Pimaric acid | 0.4 | 374 | 359, 257, 241, 121 (100) |
| Isopimaric acid | 0.8 | 374 | 359, 256, 241 (100) |
| Palustric acid | 0.1 | 374 | 359, 241 (100) |
| Dehydroabietic acid | 0.4 | 372 | 357, 239 (100) |
| Abietic acid | 0.4 | 374 | 256 (100), 241, 185 |
| Mercusic (junicedric) acid | 0.2 | 480 | 465, 362 (100), 121 |
| Cardanols | 1.0 | ||
| Cardanol C17H31 | 0.1 | 400 | 180 (100) |
| Cardanol C17H35 | 0.7 | 404 | 180 (100) |
| Cardanol C19H35 | 0.2 | 430 | 180 (100) |
| Cardols | 3.7 | ||
| Cardol C15H31 | 0.4 | 464 | 268 (100) |
| Cardol C17H31 | 1.2 | 488 | 268 (100) |
| Cardol C17H33 | 0.1 | 490 | 268 (100) |
| Cardol C17H33 (isomer) | 1.1 | 490 | 268 (100) |
| Cardol C17H35 | 0.3 | 492 | 268 (100) |
| Cardol C19H35 | 0.6 | 518 | 268 (100) |
| Anacardic acids | 2.0 | ||
| Anacardic acid C15H31 | 0.3 | 492 | 477 (100), 219 |
| Anacardic acid C17H31 | 0.5 | 516 | 501, 219 (100) |
| Anacardic acid C17H35 | 0.7 | 520 | 505 (100), 219 |
| Anacardic acid C17H33 | 0.5 | 546 | 531 (100), 219 |
| Lignans | 3.9 | ||
| Pinoresinol | 3.9 | 502 | 487, 235, 223 (100), 209 |
| Triterpenes | 22.4 | ||
| Lanosterol | 0.6 | 498 | 483, 393 (100), 109 |
| β-Amyrin | 2.1 | 498 | 218 (100), 203, 189 |
| α-Amyrenone | 3.0 | 424 | 218 (100), 203, 189 |
| α-Amyrin | 2.1 | 498 | 218 (100), 203, 189 |
| Cycloartenol | 7.9 | 498 | 408 (100), 393, 365, 339 |
| Ursolic acid | 1.7 | 600 | 482, 320, 203 (100), 189, 133 |
| Oleanolic acid | 2.6 | 600 | 482, 320, 203 (100), 189 |
| Mangiferolic acid | 2.4 | 600 | 585, 510, 495, 467 (100), 441, 388 |
| Others | |||
| Palmitic acid | 0.2 | 328 | 313, 129, 117 (100) |
| Oleic acid | 0.1 | 354 | 339, 129, 117 (100) |
| Stearic acid | 0.1 | 356 | 341, 129, 117 (100) |
| Arachidic acid | 0.1 | 384 | 369, 129, 117 (100) |
| 3-Hydroxyheneicosenoic acid | 0.6 | 484 | 469, 427, 233 (100) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tandean, S.; Japardi, I.; Rusda, M.; Indharty, R.S.; Lelo, A.; Aman, R.A.; Amin, M.M.; Siahaan, A.M.P.; Eyanoer, P.C.; D’Prinzessin, C.A.; et al. Chemical Composition and Neuroprotective Properties of Indonesian Stingless Bee (Geniotrigona thoracica) Propolis Extract in an In-Vivo Model of Intracerebral Hemorrhage (ICH). Nutrients 2024, 16, 1880. https://doi.org/10.3390/nu16121880
Tandean S, Japardi I, Rusda M, Indharty RS, Lelo A, Aman RA, Amin MM, Siahaan AMP, Eyanoer PC, D’Prinzessin CA, et al. Chemical Composition and Neuroprotective Properties of Indonesian Stingless Bee (Geniotrigona thoracica) Propolis Extract in an In-Vivo Model of Intracerebral Hemorrhage (ICH). Nutrients. 2024; 16(12):1880. https://doi.org/10.3390/nu16121880
Chicago/Turabian StyleTandean, Steven, Iskandar Japardi, Muhammad Rusda, Rr Suzy Indharty, Aznan Lelo, Renindra Ananda Aman, Mustafa Mahmud Amin, Andre Marolop Pangihutan Siahaan, Putri Chairani Eyanoer, Celine Augla D’Prinzessin, and et al. 2024. "Chemical Composition and Neuroprotective Properties of Indonesian Stingless Bee (Geniotrigona thoracica) Propolis Extract in an In-Vivo Model of Intracerebral Hemorrhage (ICH)" Nutrients 16, no. 12: 1880. https://doi.org/10.3390/nu16121880
APA StyleTandean, S., Japardi, I., Rusda, M., Indharty, R. S., Lelo, A., Aman, R. A., Amin, M. M., Siahaan, A. M. P., Eyanoer, P. C., D’Prinzessin, C. A., Lesmana, R., Popova, M., Trusheva, B., Bankova, V., & Zulhendri, F. (2024). Chemical Composition and Neuroprotective Properties of Indonesian Stingless Bee (Geniotrigona thoracica) Propolis Extract in an In-Vivo Model of Intracerebral Hemorrhage (ICH). Nutrients, 16(12), 1880. https://doi.org/10.3390/nu16121880