
Maternal Body Mass Index Associates with Prenatal Characteristics and Fecal Microbial Communities

 , , ,
, , , 
Abstract

1. Introduction
2. Materials and Methods
2.1. Study Participants
2.2. Data Collection
2.3. Maternal Pre-Pregnancy Body Mass Index Classification
2.4. DNA Extraction and Amplification
2.5. Statistical Analysis
3. Results
3.1. Participant Characteristics
3.2. Alpha Diversity
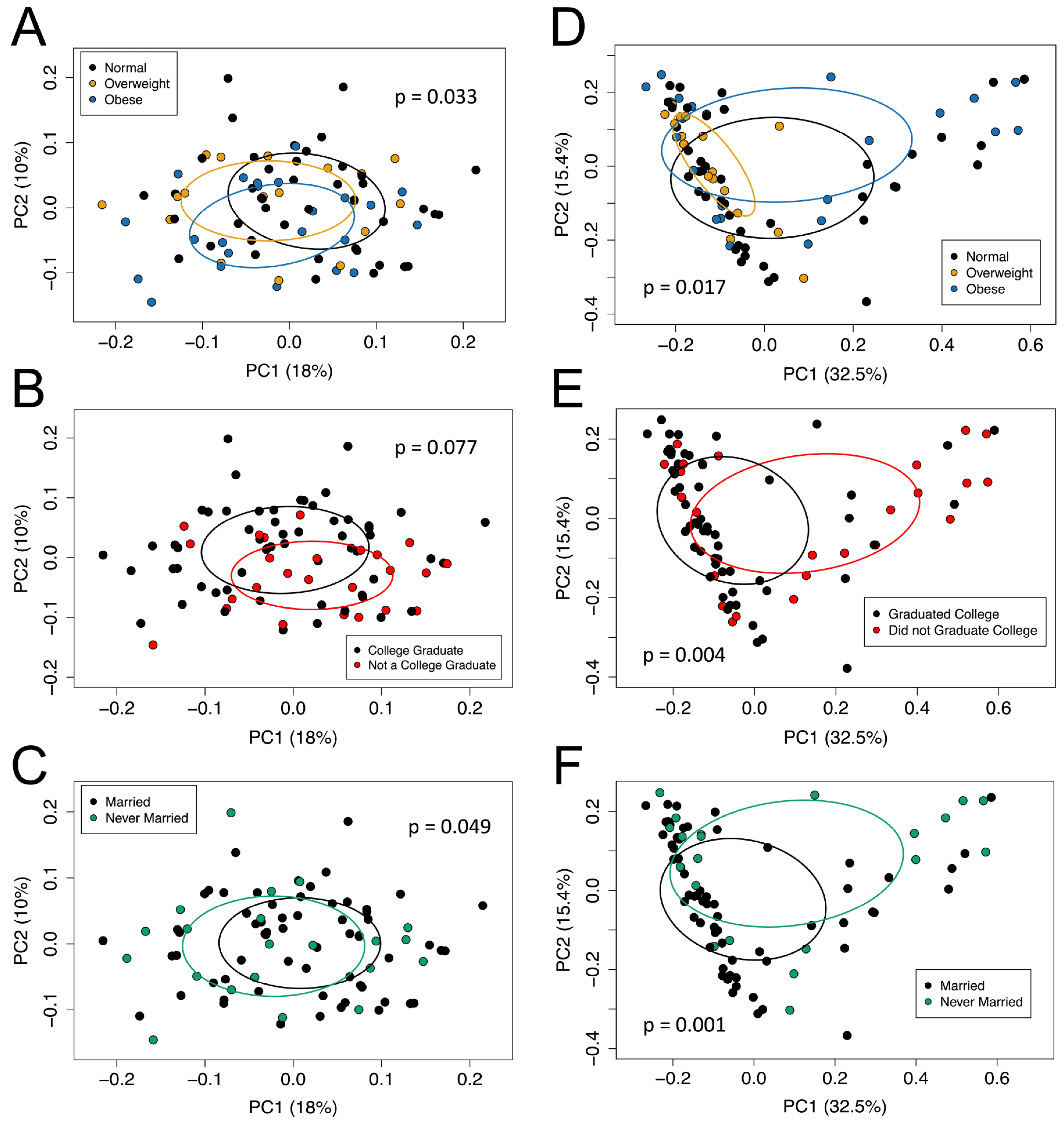
3.3. Beta Diversity
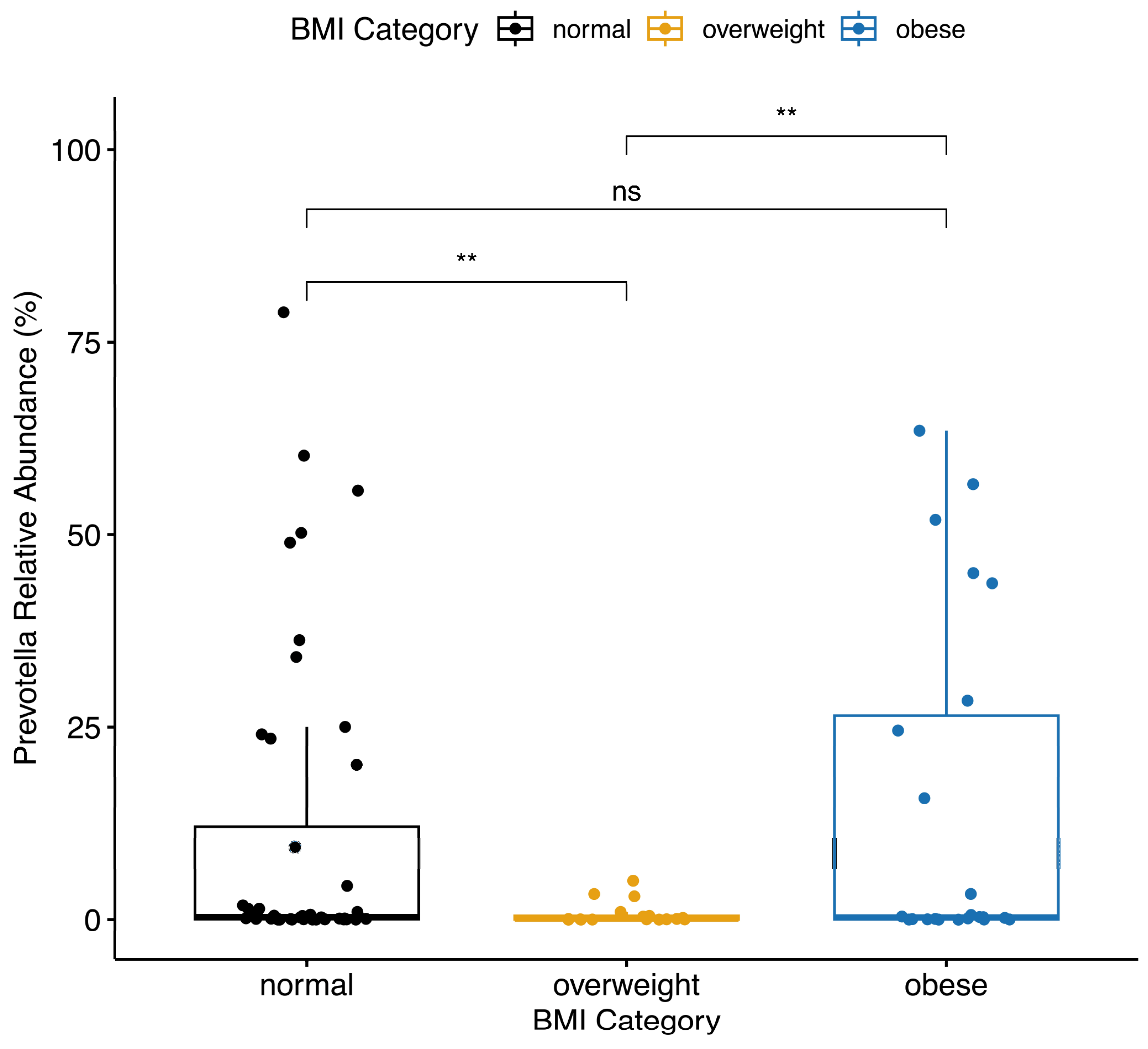
3.4. Differences in Bacterial Taxa
3.5. Multivariable Analysis
3.5.1. Alpha Diversity
3.5.2. Beta Diversity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Driscoll, A.K.; Gregory, E.C.W. Increases in Prepregnancy Obesity: United States, 2016–2019. NCHS Data Brief 2020, 392, 1–8. [Google Scholar]
- Yao, Y.; Cai, X.; Ye, Y.; Wang, F.; Chen, F.; Zheng, C. The Role of Microbiota in Infant Health: From Early Life to Adulthood. Front. Immunol. 2021, 12, 708472. [Google Scholar] [CrossRef] [PubMed]
- Sajdel-Sulkowska, E.M. The Impact of Maternal Gut Microbiota during Pregnancy on Fetal Gut–Brain Axis Development and Life-Long Health Outcomes. Microorganisms 2023, 11, 2199. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Liang, X.; Bao, H.; Ma, G.; Tang, X.; Luo, H.; Xiao, X. Multi-Omics Analysis Reveals the Associations between Altered Gut Microbiota, Metabolites, and Cytokines during Pregnancy. mSystems 2024, 9, e01252-23. [Google Scholar] [CrossRef]
- Turroni, F.; Milani, C.; Duranti, S.; Lugli, G.A.; Bernasconi, S.; Margolles, A.; di Pierro, F.; van Sinderen, D.; Ventura, M. The Infant Gut Microbiome as a Microbial Organ Influencing Host Well-Being. Ital. J. Pediatr. 2020, 46, 16. [Google Scholar] [CrossRef]
- Kalbermatter, C.; Fernandez Trigo, N.; Christensen, S.; Ganal-Vonarburg, S.C. Maternal Microbiota, Early Life Colonization and Breast Milk Drive Immune Development in the Newborn. Front. Immunol. 2021, 12, 683022. [Google Scholar] [CrossRef]
- Portela, A.; Esteller, M. Epigenetic Modifications and Human Disease. Nat. Biotechnol. 2010, 28, 1057–1068. [Google Scholar] [CrossRef]
- Li, Y. Epigenetic Mechanisms Link Maternal Diets and Gut Microbiome to Obesity in the Offspring. Front. Genet. 2018, 9, 390209. [Google Scholar] [CrossRef] [PubMed]
- Sharp, G.C.; Salas, L.A.; Monnereau, C.; Allard, C.; Yousefi, P.; Everson, T.M.; Bohlin, J.; Xu, Z.; Huang, R.-C.; Reese, S.E.; et al. Maternal BMI at the Start of Pregnancy and Offspring Epigenome-Wide DNA Methylation: Findings from the Pregnancy and Childhood Epigenetics (PACE) Consortium. Hum. Mol. Genet. 2017, 26, 4067–4085. [Google Scholar] [CrossRef]
- Koemel, N.A.; Skilton, M.R. Epigenetic Aging in Early Life: Role of Maternal and Early Childhood Nutrition. Curr. Nutr. Rep. 2022, 11, 318–328. [Google Scholar] [CrossRef]
- Phang, M.; Ross, J.; Raythatha, J.H.; Dissanayake, H.U.; McMullan, R.L.; Kong, Y.; Hyett, J.; Gordon, A.; Molloy, P.; Skilton, M.R. Epigenetic Aging in Newborns: Role of Maternal Diet. Am. J. Clin. Nutr. 2020, 111, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Monasso, G.S.; Küpers, L.K.; Jaddoe, V.W.V.; Heil, S.G.; Felix, J.F. Associations of Circulating Folate, Vitamin B12 and Homocysteine Concentrations in Early Pregnancy and Cord Blood with Epigenetic Gestational Age: The Generation R Study. Clin. Epigenet. 2021, 13, 95. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.; Quan, S.; Yang, G.; Ye, Q.; Chen, M.; Yu, H.; Wang, G.; Wang, Y.; Zeng, X.; Qiao, S. One Carbon Metabolism and Mammalian Pregnancy Outcomes. Mol. Nutr. Food Res. 2021, 65, 2000734. [Google Scholar] [CrossRef] [PubMed]
- Sugino, K.Y.; Paneth, N.; Comstock, S.S. Michigan Cohorts to Determine Associations of Maternal Pre-Pregnancy Body Mass Index with Pregnancy and Infant Gastrointestinal Microbial Communities: Late Pregnancy and Early Infancy. PLoS ONE 2019, 14, e0213733. [Google Scholar] [CrossRef] [PubMed]
- Ruebel, M.L.; Gilley, S.P.; Sims, C.R.; Zhong, Y.; Turner, D.; Chintapalli, S.V.; Piccolo, B.D.; Andres, A.; Shankar, K. Associations between Maternal Diet, Body Composition and Gut Microbial Ecology in Pregnancy. Nutrients 2021, 13, 3295. [Google Scholar] [CrossRef] [PubMed]
- Strobel, K.M.; Juul, S.E.; Hendrixson, D.T. Maternal Nutritional Status and the Microbiome across the Pregnancy and the Post-Partum Period. Microorganisms 2023, 11, 1569. [Google Scholar] [CrossRef] [PubMed]
- Pinart, M.; Dötsch, A.; Schlicht, K.; Laudes, M.; Bouwman, J.; Forslund, S.K.; Pischon, T.; Nimptsch, K. Gut Microbiome Composition in Obese and Non-Obese Persons: A Systematic Review and Meta-Analysis. Nutrients 2021, 14, 12. [Google Scholar] [CrossRef] [PubMed]
- Koliada, A.; Syzenko, G.; Moseiko, V.; Budovska, L.; Puchkov, K.; Perederiy, V.; Gavalko, Y.; Dorofeyev, A.; Romanenko, M.; Tkach, S.; et al. Association between Body Mass Index and Firmicutes/Bacteroidetes Ratio in an Adult Ukrainian Population. BMC Microbiol. 2017, 17, 120. [Google Scholar] [CrossRef]
- Zacarías, M.F.; Collado, M.C.; Gómez-Gallego, C.; Flinck, H.; Aittoniemi, J.; Isolauri, E.; Salminen, S. Pregestational Overweight and Obesity Are Associated with Differences in Gut Microbiota Composition and Systemic Inflammation in the Third Trimester. PLoS ONE 2018, 13, e0200305. [Google Scholar] [CrossRef]
- Aatsinki, A.-K.; Uusitupa, H.-M.; Munukka, E.; Pesonen, H.; Rintala, A.; Pietilä, S.; Lahti, L.; Eerola, E.; Karlsson, L.; Karlsson, H. Gut Microbiota Composition in Mid-Pregnancy Is Associated with Gestational Weight Gain but Not Prepregnancy Body Mass Index. J. Womens Health 2018, 27, 1293–1301. [Google Scholar] [CrossRef]
- Daliry, A.; Pereira, E.N.G. da S. Role of Maternal Microbiota and Nutrition in Early-Life Neurodevelopmental Disorders. Nutrients 2021, 13, 3533. [Google Scholar] [CrossRef] [PubMed]
- Agustí, A.; García-Pardo, M.P.; López-Almela, I.; Campillo, I.; Maes, M.; Romaní-Pérez, M.; Sanz, Y. Interplay between the Gut-Brain Axis, Obesity and Cognitive Function. Front. Neurosci. 2018, 12, 155. [Google Scholar] [CrossRef] [PubMed]
- Asadi, A.; Shadab Mehr, N.; Mohamadi, M.H.; Shokri, F.; Heidary, M.; Sadeghifard, N.; Khoshnood, S. Obesity and Gut–Microbiota–Brain Axis: A Narrative Review. J. Clin. Lab. Anal. 2022, 36, e24420. [Google Scholar] [CrossRef] [PubMed]
- Mayer, E.A.; Nance, K.; Chen, S. The Gut–Brain Axis. Annu. Rev. Med. 2022, 73, 439–453. [Google Scholar] [CrossRef] [PubMed]
- Edlow, A.G. Maternal Obesity and Neurodevelopmental and Psychiatric Disorders in Offspring. Prenat. Diagn. 2017, 37, 95–110. [Google Scholar] [CrossRef] [PubMed]
- Buss, C.; Entringer, S.; Davis, E.P.; Hobel, C.J.; Swanson, J.M.; Wadhwa, P.D.; Sandman, C.A. Impaired Executive Function Mediates the Association between Maternal Pre-Pregnancy Body Mass Index and Child ADHD Symptoms. PLoS ONE 2012, 7, e37758. [Google Scholar] [CrossRef] [PubMed]
- Howell, K.R.; Powell, T.L. Effects of Maternal Obesity on Placental Function and Fetal Development. Reproduction 2017, 153, R97–R108. [Google Scholar] [CrossRef]
- Hu, C.; Yan, Y.; Ji, F.; Zhou, H. Maternal Obesity Increases Oxidative Stress in Placenta and It Is Associated with Intestinal Microbiota. Front. Cell. Infect. Microbiol. 2021, 11, 671347. [Google Scholar] [CrossRef]
- CHARM: Child Health Advances from Research with Mothers. Available online: https://charmstudy.epibio.msu.edu (accessed on 15 October 2023).
- Ma, T.; Bu, S.; Paneth, N.; Kerver, J.M.; Comstock, S.S. Vitamin D Supplementation in Exclusively Breastfed Infants Is Associated with Alterations in the Fecal Microbiome. Nutrients 2022, 14, 202. [Google Scholar] [CrossRef]
- Paneth, N.; Monk, C. The Importance of Cohort Research Starting Early in Life to Understanding Child Health. Curr. Opin. Pediatr. 2018, 30, 292–296. [Google Scholar] [CrossRef]
- Haddad, E.N.; Comstock, S.S. Archive for Research in Child Health (ARCH) and Baby Gut: Study Protocol for a Remote, Prospective, Longitudinal Pregnancy and Birth Cohort to Address Microbiota Development and Child Health. Methods Protoc. 2021, 4, 52. [Google Scholar] [CrossRef] [PubMed]
- Defining Adult Overweight and Obesity. Available online: https://www.cdc.gov/obesity (accessed on 17 August 2022).
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a Dual-Index Sequencing Strategy and Curation Pipeline for Analyzing Amplicon Sequence Data on the MiSeq Illumina Sequencing Platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Simpson, G.L.; Blanchet, G.; Kindt, R. Vegan: Community Ecology Package. R Version 2.6-2. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 20 January 2021).
- R Core Team. R: A Language and Environment for Statistical Computing 2020; R Core Team: Vienna, Austria, 2020. [Google Scholar]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S; Springer: New York, NY, USA, 2002; ISBN 978-1-4419-3008-8. [Google Scholar]
- Scott, F.I.; Horton, D.B.; Mamtani, R.; Haynes, K.; Goldberg, D.S.; Lee, D.Y.; Lewis, J.D. Administration of Antibiotics to Children Before Age 2 Years Increases Risk for Childhood Obesity. Gastroenterology 2016, 151, 120–129.e5. [Google Scholar] [CrossRef] [PubMed]
- Furlong, M.; Deming-Halverson, S.; Sandler, D.P. Chronic Antibiotic Use during Adulthood and Weight Change in the Sister Study. PLoS ONE 2019, 14, e0216959. [Google Scholar] [CrossRef] [PubMed]
- Bongers, K.S.; McDonald, R.A.; Winner, K.M.; Falkowski, N.R.; Brown, C.A.; Baker, J.M.; Hinkle, K.J.; Fergle, D.J.; Dickson, R.P. Antibiotics Cause Metabolic Changes in Mice Primarily through Microbiome Modulation Rather than Behavioral Changes. PLoS ONE 2022, 17, e0265023. [Google Scholar] [CrossRef] [PubMed]
- Vallianou, N.; Dalamaga, M.; Stratigou, T.; Karampela, I.; Tsigalou, C. Do Antibiotics Cause Obesity Through Long-Term Alterations in the Gut Microbiome? A Review of Current Evidence. Curr. Obes. Rep. 2021, 10, 244–262. [Google Scholar] [CrossRef]
- Leong, K.S.W.; Derraik, J.G.B.; Hofman, P.L.; Cutfield, W.S. Antibiotics, Gut Microbiome and Obesity. Clin. Endocrinol. 2018, 88, 185–200. [Google Scholar] [CrossRef] [PubMed]
- del Fiol, F.S.; Balcão, V.M.; Barberato-Fillho, S.; Lopes, L.C.; Bergamaschi, C.C. Obesity: A New Adverse Effect of Antibiotics? Front. Pharmacol. 2018, 9, 1408. [Google Scholar] [CrossRef]
- Dare, S.; Mackay, D.F.; Pell, J.P. Relationship between Smoking and Obesity: A Cross-Sectional Study of 499,504 Middle-Aged Adults in the UK General Population. PLoS ONE 2015, 10, e0123579. [Google Scholar] [CrossRef]
- Klesges, R.C.; Meyers, A.W.; Klesges, L.M.; LaVasque, M.E. Smoking, Body Weight, and Their Effects on Smoking Behavior: A Comprehensive Review of the Literature. Psychol. Bull. 1989, 106, 204–230. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, S.L.; Hagedorn, A.; Yeom, J.; Saito, Y.; Yokoyama, E.; Crimmins, E.M. A Tale of Two Countries—The United States and Japan: Are Differences in Health Due to Differences in Overweight? J. Epidemiol. 2008, 18, 280–290. [Google Scholar] [CrossRef] [PubMed]
- Tzotzas, T.; Vlahavas, G.; Papadopoulou, S.K.; Kapantais, E.; Kaklamanou, D.; Hassapidou, M. Marital Status and Educational Level Associated to Obesity in Greek Adults: Data from the National Epidemiological Survey. BMC Public Health 2010, 10, 732. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, Y. Socioeconomic Inequality of Obesity in the United States: Do Gender, Age, and Ethnicity Matter? Soc. Sci. Med. 2004, 58, 1171–1180. [Google Scholar] [CrossRef]
- Addoum, J.M.; Korniotis, G.; Kumar, A. Stature, Obesity, and Portfolio Choice. Manag. Sci. 2017, 63, 3393–3413. [Google Scholar] [CrossRef]
- Gallè, F.; Valeriani, F.; Cattaruzza, M.S.; Gianfranceschi, G.; Liguori, R.; Antinozzi, M.; Mederer, B.; Liguori, G.; Romano Spica, V. Mediterranean Diet, Physical Activity and Gut Microbiome Composition: A Cross-Sectional Study among Healthy Young Italian Adults. Nutrients 2020, 12, 2164. [Google Scholar] [CrossRef]
- Gao, X.; Zhang, M.; Xue, J.; Huang, J.; Zhuang, R.; Zhou, X.; Zhang, H.; Fu, Q.; Hao, Y. Body Mass Index Differences in the Gut Microbiota Are Gender Specific. Front. Microbiol. 2018, 9, 1250. [Google Scholar] [CrossRef]
- Peters, B.A.; Shapiro, J.A.; Church, T.R.; Miller, G.; Trinh-Shevrin, C.; Yuen, E.; Friedlander, C.; Hayes, R.B.; Ahn, J. A Taxonomic Signature of Obesity in a Large Study of American Adults. Sci. Rep. 2018, 8, 9749. [Google Scholar] [CrossRef]
- Kasai, C.; Sugimoto, K.; Moritani, I.; Tanaka, J.; Oya, Y.; Inoue, H.; Tameda, M.; Shiraki, K.; Ito, M.; Takei, Y.; et al. Comparison of the Gut Microbiota Composition between Obese and Non-Obese Individuals in a Japanese Population, as Analyzed by Terminal Restriction Fragment Length Polymorphism and next-Generation Sequencing. BMC Gastroenterol. 2015, 15, 100. [Google Scholar] [CrossRef]
- Stanislawski, M.A.; Dabelea, D.; Lange, L.A.; Wagner, B.D.; Lozupone, C.A. Gut Microbiota Phenotypes of Obesity. NPJ Biofilms Microbiomes 2019, 5, 18. [Google Scholar] [CrossRef]
- de la Cuesta-Zuluaga, J.; Kelley, S.T.; Chen, Y.; Escobar, J.S.; Mueller, N.T.; Ley, R.E.; McDonald, D.; Huang, S.; Swafford, A.D.; Knight, R.; et al. Age- and Sex-Dependent Patterns of Gut Microbial Diversity in Human Adults. mSystems 2019, 4, e00261-19. [Google Scholar] [CrossRef] [PubMed]
- Kovatcheva-Datchary, P.; Nilsson, A.; Akrami, R.; Lee, Y.S.; De Vadder, F.; Arora, T.; Hallen, A.; Martens, E.; Björck, I.; Bäckhed, F. Dietary Fiber-Induced Improvement in Glucose Metabolism Is Associated with Increased Abundance of Prevotella. Cell Metab. 2015, 22, 971–982. [Google Scholar] [CrossRef]
- Savin, Z.; Kivity, S.; Yonath, H.; Yehuda, S. Smoking and the Intestinal Microbiome. Arch. Microbiol. 2018, 200, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Human Gut Microbes Associated with Obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef] [PubMed]
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The Firmicutes/Bacteroidetes Ratio: A Relevant Marker of Gut Dysbiosis in Obese Patients? Nutrients 2020, 12, 1474. [Google Scholar] [CrossRef]
- Chen, J.; Ryu, E.; Hathcock, M.; Ballman, K.; Chia, N.; Olson, J.E.; Nelson, H. Impact of Demographics on Human Gut Microbial Diversity in a US Midwest Population. PeerJ 2016, 4, e1514. [Google Scholar] [CrossRef]
- Ozato, N.; Saito, S.; Yamaguchi, T.; Katashima, M.; Tokuda, I.; Sawada, K.; Katsuragi, Y.; Kakuta, M.; Imoto, S.; Ihara, K.; et al. Blautia Genus Associated with Visceral Fat Accumulation in Adults 20–76 Years of Age. NPJ Biofilms Microbiomes 2019, 5, 28. [Google Scholar] [CrossRef]
- Zhang, W.; Li, J.; Lu, S.; Han, N.; Miao, J.; Zhang, T.; Qiang, Y.; Kong, Y.; Wang, H.; Gao, T.; et al. Gut Microbiota Community Characteristics and Disease-Related Microorganism Pattern in a Population of Healthy Chinese People. Sci. Rep. 2019, 9, 1594. [Google Scholar] [CrossRef]
- Russell, W.R.; Gratz, S.W.; Duncan, S.H.; Holtrop, G.; Ince, J.; Scobbie, L.; Duncan, G.; Johnstone, A.M.; Lobley, G.E.; Wallace, R.J.; et al. High-Protein, Reduced-Carbohydrate Weight-Loss Diets Promote Metabolite Profiles Likely to Be Detrimental to Colonic Health. Am. J. Clin. Nutr. 2011, 93, 1062–1072. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet Rapidly and Reproducibly Alters the Human Gut Microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef]
- Meehan, C.J.; Beiko, R.G. A Phylogenomic View of Ecological Specialization in the Lachnospiraceae, a Family of Digestive Tract-Associated Bacteria. Genome Biol. Evol. 2014, 6, 703–713. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.; Chung, H.; Weatherspoon, L.; Song, W.O. Validity of Prepregnancy Weight Status Estimated from Self-Reported Height and Weight. Matern. Child Health J. 2014, 18, 1667–1674. [Google Scholar] [CrossRef] [PubMed]
- Hodge, J.M.; Shah, R.; McCullough, M.L.; Gapstur, S.M.; Patel, A.v. Validation of Self-Reported Height and Weight in a Large, Nationwide Cohort of U.S. Adults. PLoS ONE 2020, 15, e0231229. [Google Scholar] [CrossRef] [PubMed]
- Yehya, N. Gut Bacteria Differences between Black and White Women Linked to Insulin Sensitivity. Available online: https://health.ucdavis.edu/news/headlines/gut-bacteria-differences-between-black-and-white-women-linked-to-insulin-sensitivity/2022/01 (accessed on 22 February 2023).
- Brooks, A.W.; Priya, S.; Blekhman, R.; Bordenstein, S.R. Gut Microbiota Diversity across Ethnicities in the United States. PLoS Biol. 2018, 16, e2006842. [Google Scholar] [CrossRef] [PubMed]
- Knapp, E.A.; Kress, A.M.; Parker, C.B.; Page, G.P.; McArthur, K.; Gachigi, K.K.; Alshawabkeh, A.N.; Aschner, J.L.; Bastain, T.M.; Breton, C.V.; et al. The Environmental Influences on Child Health Outcomes (ECHO)-Wide Cohort. Am. J. Epidemiol. 2023, 192, 1249–1263. [Google Scholar] [CrossRef] [PubMed]
- Bragg, M.G.; Westlake, M.; Alshawabkeh, A.N.; Bekelman, T.A.; Camargo, C.A.; Catellier, D.J.; Comstock, S.S.; Dabelea, D.; Dunlop, A.L.; Hedderson, M.M.; et al. Opportunities for Examining Child Health Impacts of Early-Life Nutrition in the ECHO Program: Maternal and Child Dietary Intake Data from Pregnancy to Adolescence. Curr. Dev. Nutr. 2023, 7, 102019. [Google Scholar] [CrossRef] [PubMed]
- Adithya, K.K.; Rajeev, R.; Selvin, J.; Seghal Kiran, G. Dietary Influence on the Dynamics of the Human Gut Microbiome: Prospective Implications in Interventional Therapies. ACS Food Sci. Technol. 2021, 1, 717–736. [Google Scholar] [CrossRef]
- Durack, J.; Lynch, S.v. The Gut Microbiome: Relationships with Disease and Opportunities for Therapy. J. Exp. Med. 2019, 216, 20–40. [Google Scholar] [CrossRef] [PubMed]
- Santarossa, S.; Sitarik, A.R.; Cassidy-Bushrow, A.E.; Comstock, S.S. Prenatal Physical Activity and the Gut Microbiota of Pregnant Women: Results from a Preliminary Investigation. Phys. Act. Nutr. 2023, 27, 001–007. [Google Scholar] [CrossRef]
- Maugeri, A.; Barchitta, M.; Favara, G.; La Rosa, M.C.; La Mastra, C.; Magnano San Lio, R.; Agodi, A. Maternal Dietary Patterns Are Associated with Pre-Pregnancy Body Mass Index and Gestational Weight Gain: Results from the “Mamma & Bambino” Cohort. Nutrients 2019, 11, 1308. [Google Scholar] [CrossRef]
- Ferreira, L.B.; Lobo, C.V.; Miranda, A.E.d.S.; Carvalho, B.d.C.; dos Santos, L.C. Dietary Patterns during Pregnancy and Gestational Weight Gain: A Systematic Review. Rev. Bras. de Ginecol. e Obs./RBGO Gynecol. Obstet. 2022, 44, 540–547. [Google Scholar] [CrossRef] [PubMed]
- Wesolowski, S.R.; El Kasmi, K.C.; Jonscher, K.R.; Friedman, J.E. Developmental Origins of NAFLD: A Womb with a Clue. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 81–96. [Google Scholar] [CrossRef] [PubMed]
- Vähämiko, S.; Laiho, A.; Lund, R.; Isolauri, E.; Salminen, S.; Laitinen, K. The Impact of Probiotic Supplementation during Pregnancy on DNA Methylation of Obesity-Related Genes in Mothers and Their Children. Eur. J. Nutr. 2019, 58, 367–377. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Pre-Pregnancy Characteristic | Normal BMI (n = 44) n, (%) | Overweight BMI (n = 17) n, (%) | Obese BMI (n = 23) n, (%) | p-Value |
| Race | 0.371 | |||
| White | 40 (90.9) | 16 (94.1) | 18 (78.3) | |
| Black | 2 (4.5) | 1 (5.9) | 4 (17.4) | |
| Other | 2 (4.5) | 0 (0) | 1 (4.3) | |
| Antibiotic use * | 13 (29.5) | 12 (70.6) | 16 (69.6) | 0.001 |
| Ever smoked cigarettes | 2 (4.5) | 1 (5.9) | 7 (30.4) | 0.006 |
| Ever smoked marijuana | 3 (7.0) | 1 (6.3) | 3 (13.0) | 0.657 |
| Missing, n (%) | 1 (2.3) | 1 (5.9) | 0 (0) | |
| Asthma diagnosis | 5 (11.4) | 5 (29.4) | 6 (26.1) | 0.165 |
| Allergy diagnosis | 4 (9.3) | 2 (11.8) | 2 (8.7) | 0.943 |
| Missing, n (%) | 1 (2.3) | 0 (0) | 0 (0) | |
| College graduate | 34 (77.3) | 13 (81.3) | 11 (47.8) | 0.024 |
| Missing, n (%) | 0 (0) | 1 (5.9) | 0 (0) | |
| Married | 39 (88.6) | 12 (70.6) | 12 (52.2) | 0.004 |
| Owns a car | 43 (97.7) | 14 (82.4) | 19 (82.6) | 0.060 |
| Owns a home | 33 (76.7) | 11 (64.7) | 16 (69.6) | 0.607 |
| Missing, n (%) | 1 (2.3) | 0 (0) | 0 (0) | |
| Owns stocks or bonds | 24 (54.5) | 7 (41.2) | 5 (22.7) | 0.048 |
| Missing, n (%) | 0 (0) | 0 (0) | 1 (4.3) | |
| Pre-Pregnancy Characteristic | Mean (± SD) | Mean (± SD) | Mean (± SD) | p-Value |
| Maternal age | 31.5 (5.1) | 32.8 (7.0) | 30.7 (4.1) | 0.293 |
| Missing, n (%) | 2 (4.5) | 0 (0) | 1 (4.3) | |
| Parity | 1.38 (1.7) | 1.06 (1.1) | 2.05 (1.6) | 0.129 |
| Missing, n (%) | 2 (4.5) | 0 (0) | 1 (4.3) |
| Pre-Pregnancy Characteristics | Chao Mean (SD) | Shannon Mean (SD) | Inverse Simpson Mean (SD) |
|---|---|---|---|
| Normal BMI | 116.7 (30.6) | 2.7 (0.4) | 9.7 (3.6) |
| Overweight BMI | 110.1 (21.4) | 2.7 (0.3) | 9.8 (3.5) |
| Obese BMI | 102.0 (19.5) | 2.5 (0.4) | 7.5 (3.3) |
| p-value | 0.156 | 0.032 | 0.035 |
| ≤31 years of age | 106.4 (21.7) | 2.5 (0.4) | 7.7 (3.6) |
| >31 years of age | 114.0 (26.3) | 2.8 (0.4) | 10.4 (3.1) |
| p-value | 0.152 | <0.001 | <0.001 |
| College graduate | 113.4 (27.8) | 2.7 (0.3) | 9.7 (3.1) |
| Did not graduate college | 105.3 (23.3) | 2.5 (0.4) | 7.5 (3.8) |
| p-value | 0.176 | 0.020 | 0.014 |
| Married | 111.5 (25.0) | 2.7 (0.3) | 9.7 (3.3) |
| Unmarried | 111.0 (32.0) | 2.5 (0.4) | 7.4 (4.1) |
| p-value | 0.635 | 0.007 | 0.025 |
| Owned bonds | 111.1 (23.2) | 2.8 (0.2) | 10.6 (2.6) |
| Did not own bonds | 111.5 (29.5) | 2.5 (0.4) | 8.0 (3.8) |
| p-value | 0.696 | <0.001 | <0.001 |
| Model (Overall p-Value) | Variable | Regression Coefficient | Standard Error | p-Value | |
|---|---|---|---|---|---|
| Chao1 Index | Model 1 (p = 0.098) | Intercept | 114.098 | 3.708 | <0.0001 |
| Normal BMI | - | - | - | ||
| Overweight BMI | −3.987 | 6.907 | 0.565 | ||
| Obese BMI | −11.536 | 6.324 | 0.072 | ||
| Model 2 (p = 0.444) | Intercept | 109.330 | 6.831 | <0.0001 | |
| Normal BMI | - | - | - | ||
| Overweight BMI | −5.625 | 7.693 | 0.467 | ||
| Obese BMI | −14.752 | 7.705 | 0.059 | ||
| Antibiotic usage | 2.084 | 6.420 | 0.746 | ||
| Smoking status | 11.445 | 9.360 | 0.225 | ||
| College grad | −0.243 | 7.107 | 0.973 | ||
| Marital status | −1.831 | 7.914 | 0.818 | ||
| Maternal age | 8.477 | 6.050 | 0.166 | ||
| Shannon Index | Model 1 (p = 0.076) | Intercept | 2.677 | 0.058 | <0.0001 |
| Normal BMI | - | - | - | ||
| Overweight BMI | 0.063 | 0.108 | 0.561 | ||
| Obese BMI | −0.192 | 0.099 | 0.057 | ||
| Model 2 (p = 0.006) | Intercept | 2.596 | 0.099 | <0.0001 | |
| Normal BMI | - | - | - | ||
| Overweight BMI | 0.116 | 0.112 | 0.305 | ||
| Obese BMI | −0.122 | 0.112 | 0.279 | ||
| Antibiotic usage | −0.085 | 0.093 | 0.367 | ||
| Smoking status | 0.227 | 0.136 | 0.100 | ||
| College grad | 0.021 | 0.103 | 0.837 | ||
| Marital status | −0.198 | 0.115 | 0.089 | ||
| Maternal age | 0.197 | 0.088 | 0.028 | ||
| Inverse Simpson Index | Model 1 (p = 0.047) | Intercept | 9.553 | 0.543 | <0.0001 |
| Normal BMI | - | - | - | ||
| Overweight BMI | 0.279 | 1.012 | 0.783 | ||
| Obese BMI | −2.122 | 0.543 | <0.0001 | ||
| Model 2 (p = 0.006) | Intercept | 8.722 | 0.935 | <0.0001 | |
| Normal BMI | - | - | - | ||
| Overweight BMI | 0.440 | 1.053 | 0.677 | ||
| Obese BMI | −1.651 | 1.055 | 0.122 | ||
| Antibiotic usage | −0.245 | 0.879 | 0.781 | ||
| Smoking status | 1.133 | 1.281 | 0.379 | ||
| College grad | −0.116 | 0.973 | 0.905 | ||
| Marriage status | −1.608 | 1.083 | 0.142 | ||
| Maternal age | 2.274 | 0.828 | 0.008 |
| Beta Diversity Model (Overall p-Value) | Variable | p-Value | R2 | Overall R2 | F-Statistic |
|---|---|---|---|---|---|
| Sorensen (p = 0.024) | BMI | 0.113 | 0.031 | 0.080 | 1.348 |
| Antibiotic usage | 0.480 | 0.011 | |||
| Smoking status | 0.353 | 0.013 | |||
| College graduation status | 0.082 | 0.018 | |||
| Bray–Curtis (p = 0.007) | BMI | 0.109 | 0.034 | 0.103 | 1.792 |
| Antibiotic usage | 0.490 | 0.010 | |||
| Smoking status | 0.489 | 0.010 | |||
| College graduation status | 0.003 | 0.041 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nel, N.H.; Haddad, E.N.; Kerver, J.M.; Cassidy-Bushrow, A.E.; Comstock, S.S. Maternal Body Mass Index Associates with Prenatal Characteristics and Fecal Microbial Communities. Nutrients 2024, 16, 1881. https://doi.org/10.3390/nu16121881
Nel NH, Haddad EN, Kerver JM, Cassidy-Bushrow AE, Comstock SS. Maternal Body Mass Index Associates with Prenatal Characteristics and Fecal Microbial Communities. Nutrients. 2024; 16(12):1881. https://doi.org/10.3390/nu16121881
Chicago/Turabian StyleNel, Nikita H., Eliot N. Haddad, Jean M. Kerver, Andrea E. Cassidy-Bushrow, and Sarah S. Comstock. 2024. "Maternal Body Mass Index Associates with Prenatal Characteristics and Fecal Microbial Communities" Nutrients 16, no. 12: 1881. https://doi.org/10.3390/nu16121881
APA StyleNel, N. H., Haddad, E. N., Kerver, J. M., Cassidy-Bushrow, A. E., & Comstock, S. S. (2024). Maternal Body Mass Index Associates with Prenatal Characteristics and Fecal Microbial Communities. Nutrients, 16(12), 1881. https://doi.org/10.3390/nu16121881