
Effects of Saponins on Lipid Metabolism: The Gut–Liver Axis Plays a Key Role

Abstract
1. Introduction
2. Saponin
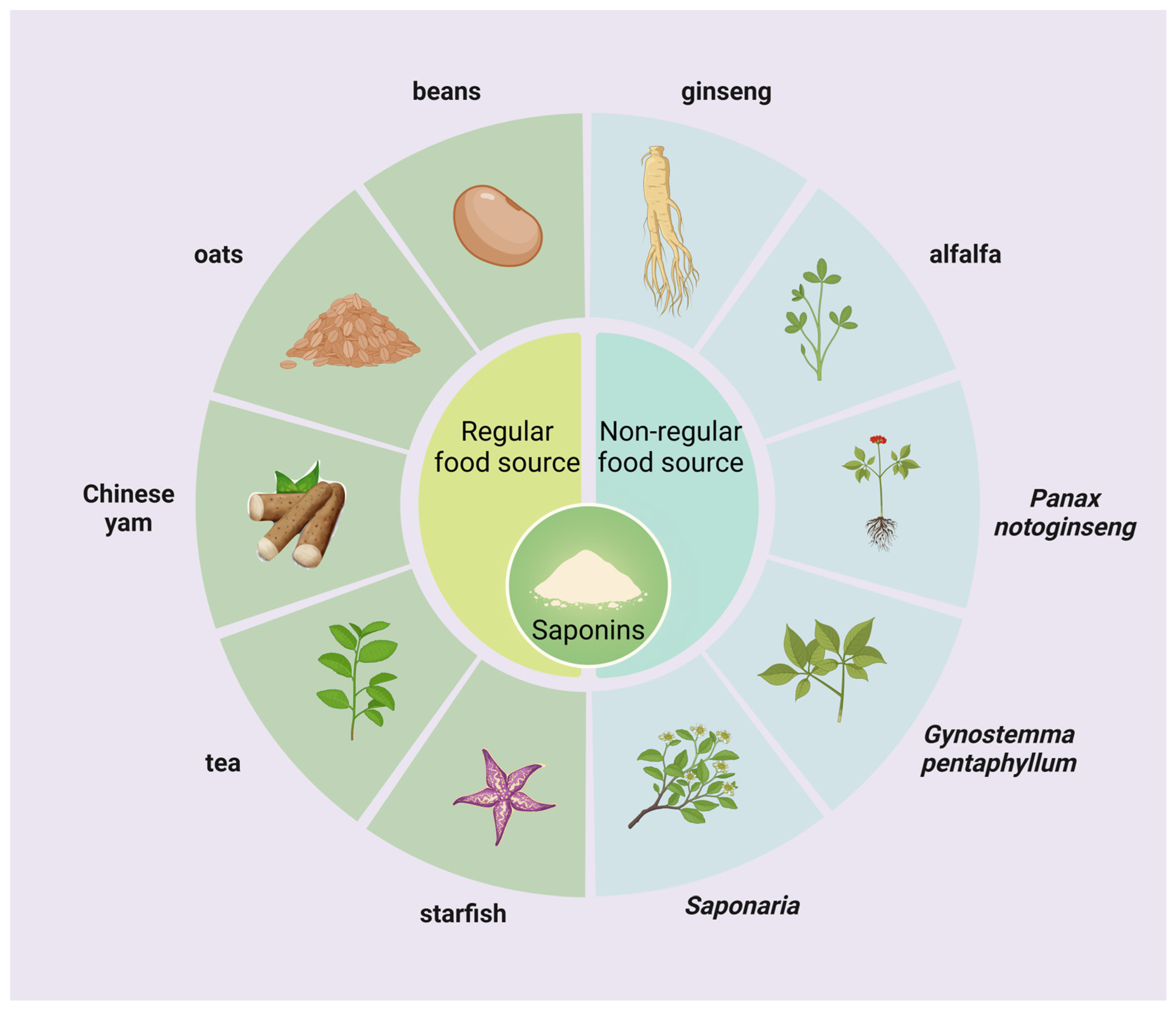
2.1. Sources of Saponins
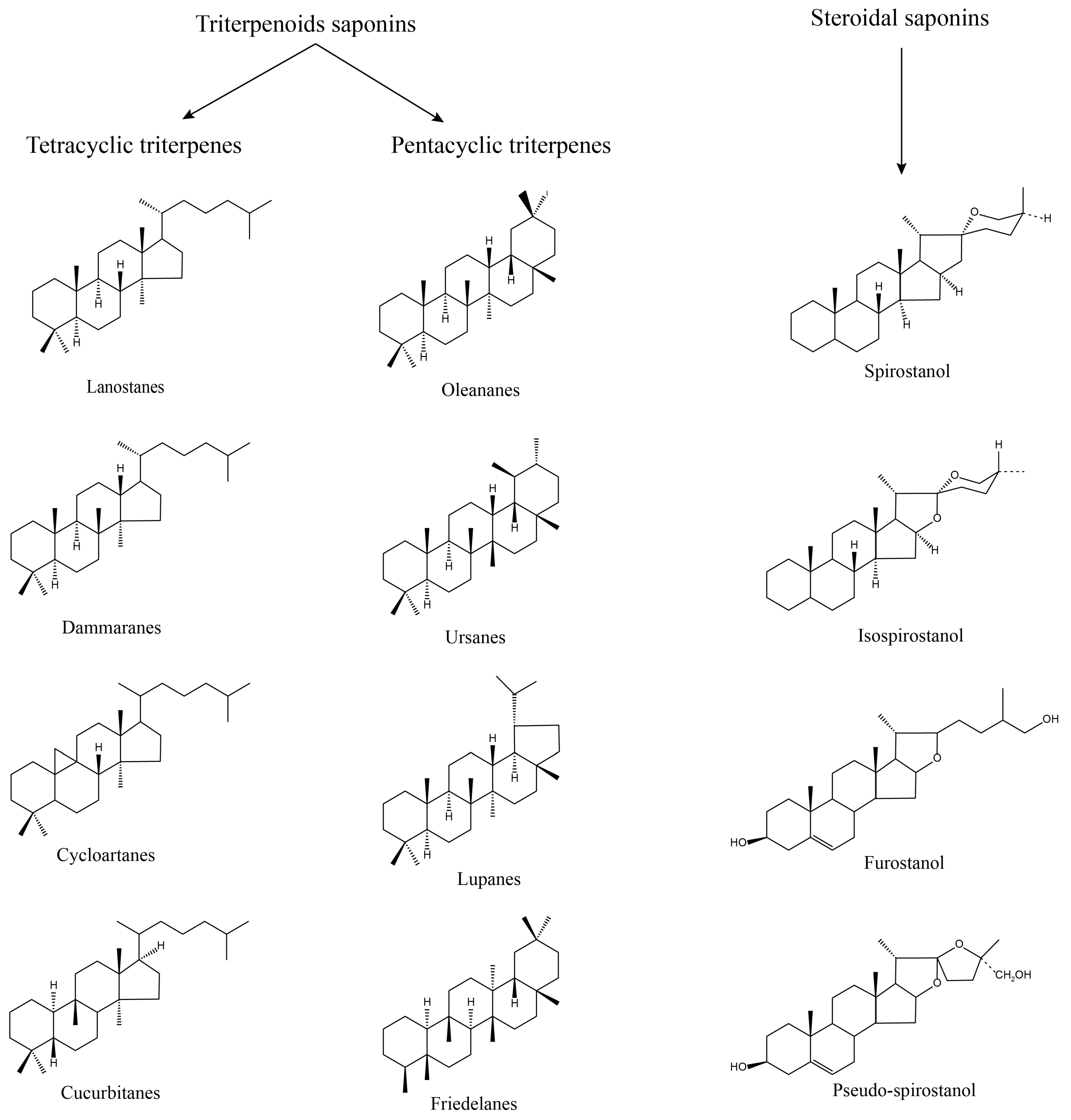
2.2. Structure and Physicochemical Properties of Saponins
2.3. Toxicity and Safety of Saponins
3. Saponins Inhibit Fat Accumulation
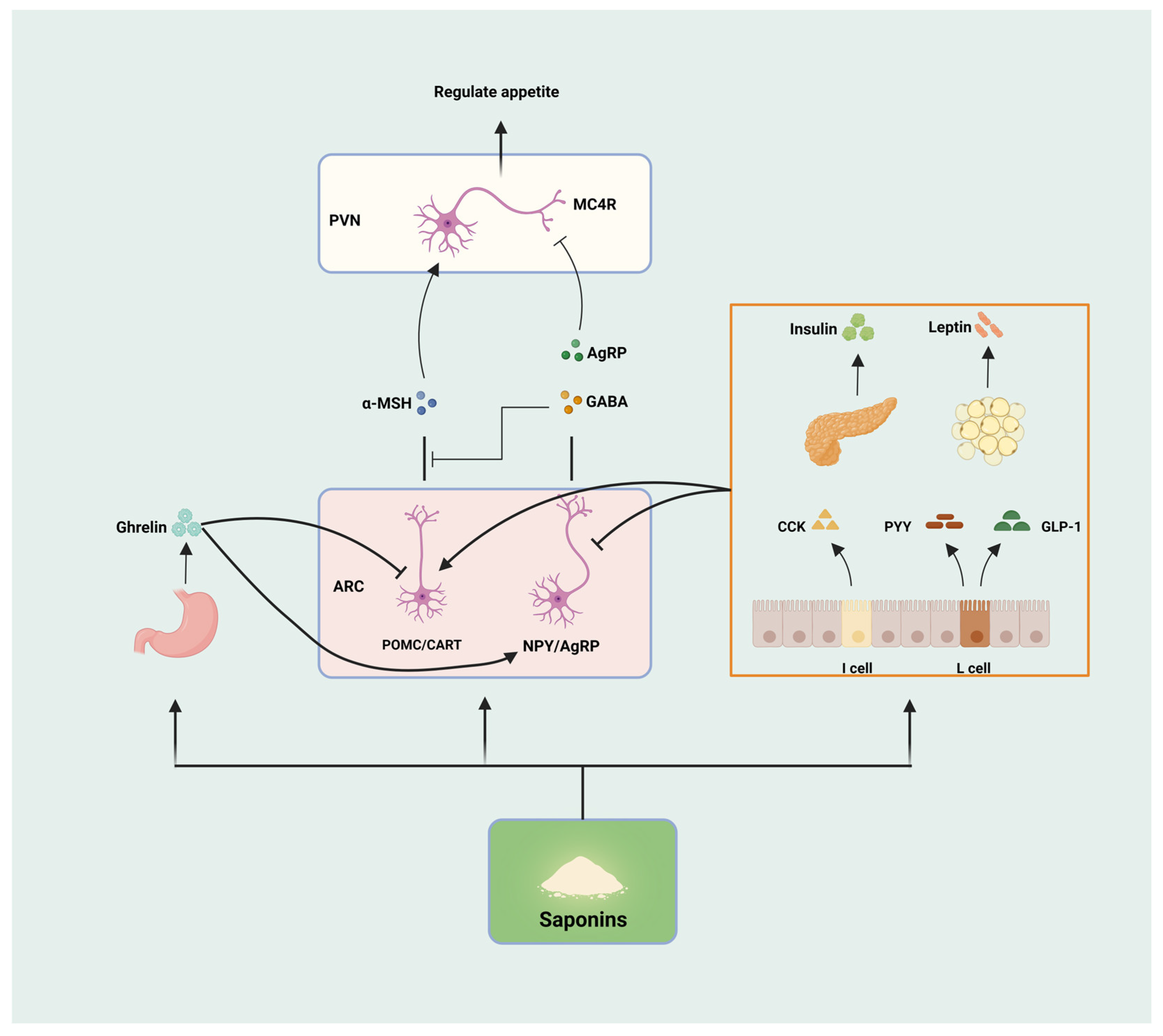
3.1. Saponins Regulate Appetite
3.2. Saponins Inhibit Pancreatic Lipase Activity
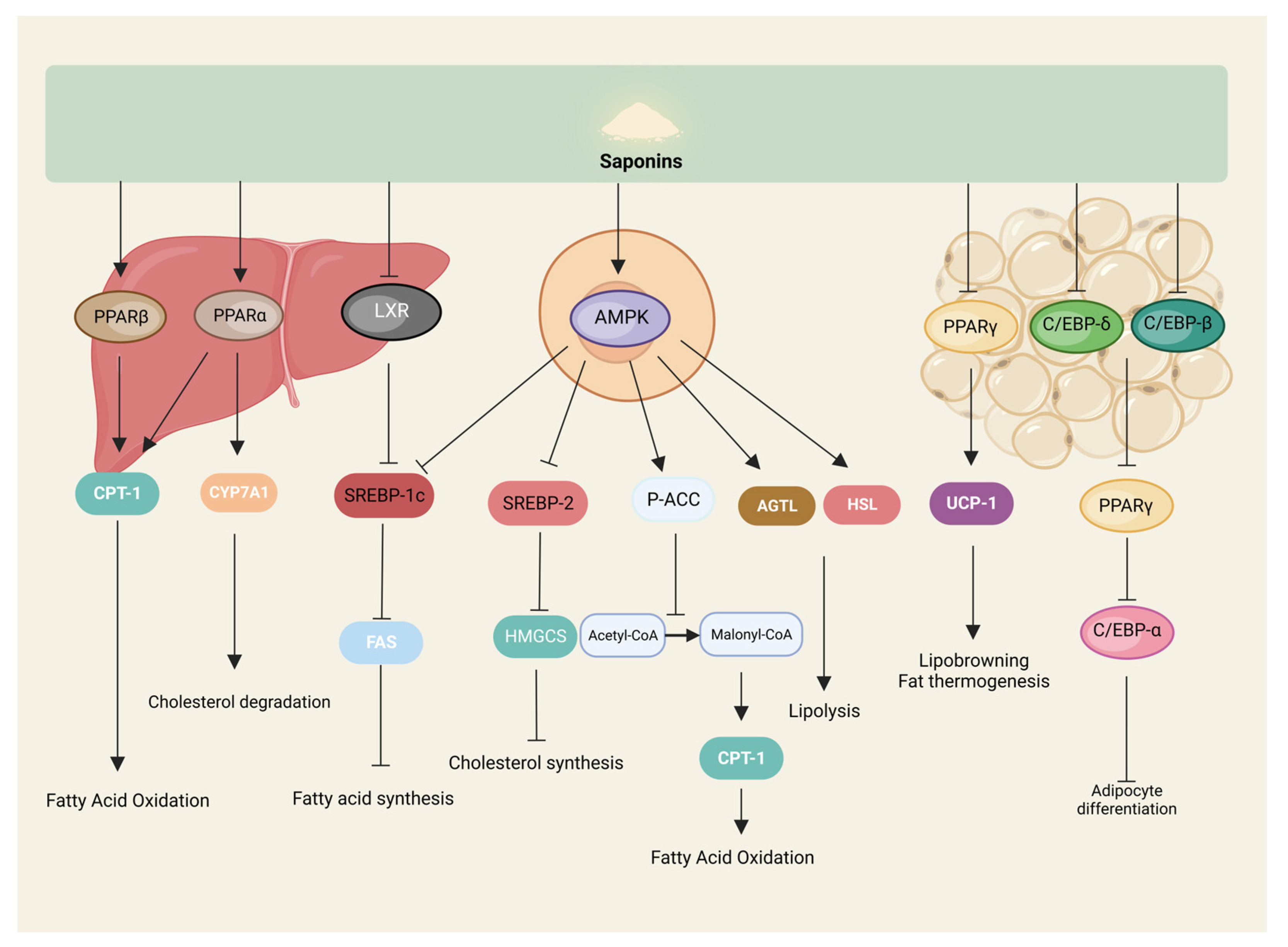
3.3. Saponins Regulate the Synthesis and Breakdown of Fats
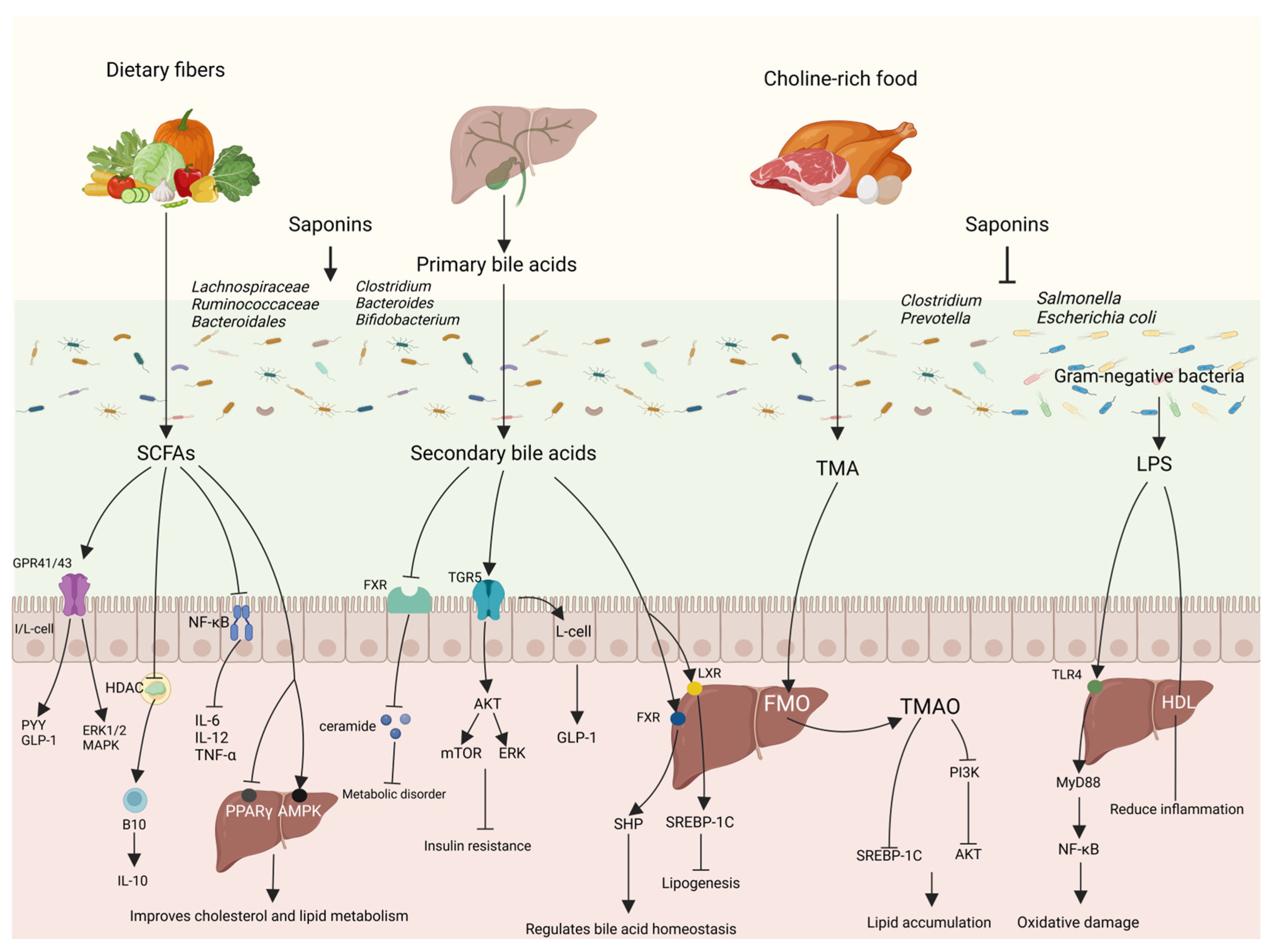
4. Saponins Regulate Lipid Metabolism via the Gut–Liver Axis
4.1. Protective Effects of Saponins on the Gut–Liver Axis
4.1.1. Protective Effects of Saponins on the Intestinal Barrier
4.1.2. Protective Effects of Saponins on the Liver
4.2. Saponins Regulate Lipid Metabolism via the Gut–Liver Axis
4.2.1. Short-Chain Fatty Acids
4.2.2. Bile Acids
4.2.3. Trimethylamine
4.2.4. Lipopolysaccharide
5. Conclusions and Outlook
Author Contributions
Funding
Conflicts of Interest
References
- Hu, P.; Li, K.; Peng, X.; Kan, Y.; Li, H.; Zhu, Y.; Wang, Z.; Li, Z.; Liu, H.-Y.; Cai, D. Nuclear Receptor PPARα as a Therapeutic Target in Diseases Associated with Lipid Metabolism Disorders. Nutrients 2023, 15, 4772. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Guo, W.-L.; Zhang, W.; Xu, J.-X.; Qian, M.; Bai, W.-D.; Zhang, Y.-Y.; Rao, P.-F.; Ni, L.; Lv, X.-C. Grifola Frondosa Polysaccharides Ameliorate Lipid Metabolic Disorders and Gut Microbiota Dysbiosis in High-Fat Diet Fed Rats. Food Funct. 2019, 10, 2560–2572. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Zhang, H.; Lu, Y.; Jiang, X.; Liu, Q.; Hu, Y.; Sun, H.; Ma, A. Effects of the Polypeptide from Peanut Meal Mixed Fermentation on Lipid Metabolism and Intestinal Flora of Hyperlipidemic Mice. J. Sci. Food Agric. 2023, 103, 4351–4359. [Google Scholar] [CrossRef] [PubMed]
- Murphy, A.J.; Febbraio, M.A. Immune-Based Therapies in Cardiovascular and Metabolic Diseases: Past, Present and Future. Nat. Rev. Immunol. 2021, 21, 669–679. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Luo, M.; Chen, Q.; Zhang, Q.; Qin, L.; Wang, Y.; Zhao, Y.; He, Y. Hypolipidemic Effect and Gut Microbiota Regulation of Gypenoside Aglycones in Rats Fed a High-Fat Diet. J. Ethnopharmacol. 2024, 328, 118066. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Zheng, Y.; Tang, W.; Yan, W.; Nie, H.; Fang, J.; Liu, G. Dietary Polyphenols in Lipid Metabolism: A Role of Gut Microbiome. Anim. Nutr. 2020, 6, 404–409. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-H.; Jeong, H.Y.; Choi, P.G.; Kim, M.J.; Seo, H.-D.; Hahm, J.-H.; Ahn, J.; Jung, C.H. C-Glycosidic Flavone-Rich Passiflora incarnata L. Leaf Extracts Decrease Body Weight and Fatty Liver in Obese Mice. Food Biosci. 2023, 55, 103028. [Google Scholar] [CrossRef]
- Yang, X.; Xiong, X.; Wang, H.; Wang, J. Protective Effects of Panax Notoginseng Saponins on Cardiovascular Diseases: A Comprehensive Overview of Experimental Studies. Evid. Based Complement. Alternat. Med. 2014, 2014, 204840. [Google Scholar] [CrossRef]
- Sparg, S.G.; Light, M.E.; van Staden, J. Biological Activities and Distribution of Plant Saponins. J. Ethnopharmacol. 2004, 94, 219–243. [Google Scholar] [CrossRef]
- Güçlü-Ustündağ, O.; Mazza, G. Saponins: Properties, Applications and Processing. Crit. Rev. Food Sci. Nutr. 2007, 47, 231–258. [Google Scholar] [CrossRef]
- Yoshiki, Y.; Kudou, S.; Okubo, K. Relationship between Chemical Structures and Biological Activities of Triterpenoid Saponins from Soybean. Biosci. Biotechnol. Biochem. 1998, 62, 2291–2299. [Google Scholar] [CrossRef] [PubMed]
- Bialy, Z.; Jurzysta, M.; Oleszek, W.; Piacente, S.; Pizza, C. Saponins in Alfalfa (Medicago sativa L.) Root and Their Structural Elucidation. J. Agric. Food Chem. 1999, 47, 3185–3192. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Laval, S.; Yu, B. Chemical Synthesis of Saponins. Adv. Carbohydr. Chem. Biochem. 2014, 71, 137–226. [Google Scholar] [CrossRef] [PubMed]
- Reichert, C.L.; Salminen, H.; Weiss, J. Quillaja Saponin Characteristics and Functional Properties. Annu. Rev. Food Sci. Technol. 2019, 10, 43–73. [Google Scholar] [CrossRef] [PubMed]
- Fink, R.; Filip, S. Surface-Active Natural Saponins. Properties, Safety, and Efficacy. Int. J. Environ. Health Res. 2023, 33, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Nakaziba, R.; Lubega, A.; Ogwal-Okeng, J.; Alele, P.E. Phytochemical Analysis, Acute Toxicity, as Well as Antihyperglycemic and Antidiabetic Activities of Corchorus olitorius L. Leaf Extracts. Sci. World J. 2022, 2022, 1376817. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhai, B.; Sun, J.; Fan, Y.; Zou, J.; Cheng, J.; Zhang, X.; Shi, Y.; Guo, D. Antioxidant, Anti-Aging and Organ Protective Effects of Total Saponins from Aralia taibaiensis. Drug Des. Dev. Ther. 2021, 15, 4025–4042. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.-Z.; Yi, H.-W.; Huang, W.; Pang, T.; Zhou, H.-P.; Wu, X.-D. Fatty Liver Diseases, Mechanisms, and Potential Therapeutic Plant Medicines. Chin. J. Nat. Med. 2020, 18, 161–168. [Google Scholar] [CrossRef]
- Ren, Z.; Chen, X.; Hong, L.; Zhao, X.; Cui, G.; Li, A.; Liu, Y.; Zhou, L.; Sun, R.; Shen, S.; et al. Nanoparticle Conjugation of Ginsenoside Rg3 Inhibits Hepatocellular Carcinoma Development and Metastasis. Small 2020, 16, e1905233. [Google Scholar] [CrossRef]
- Liu, Y.; Deng, J.; Fan, D. Ginsenoside Rk3 Ameliorates High-Fat-Diet/Streptozocin Induced Type 2 Diabetes Mellitus in Mice via the AMPK/Akt Signaling Pathway. Food Funct. 2019, 10, 2538–2551. [Google Scholar] [CrossRef]
- Dai, Y.; Liu, P.; Wen, W.; Li, P.; Yang, C.; Wang, P.; Xu, S. Sarsasapogenin, a Principal Active Component Absorbed into Blood of Total Saponins of Anemarrhena, Attenuates Proliferation and Invasion in Rheumatoid Arthritis Fibroblast-like Syno-viocytes through Downregulating PKM2 Inhibited Pathological Glycolysis. Phytother. Res. 2023, 37, 1951–1967. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Jin, S.; Xiang, X.; Tian, J.; Huang, R.; Li, S.; Chen, C.; Xu, H.; Song, C. The Metabolic Change in Serum Lysoglycerophospholipids Intervened by Triterpenoid Saponins from Kuding Tea on Hyperlipidemic Mice. Food Funct. 2019, 10, 7782–7792. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-B.; Zhao, M.; Wang, J.; Shi, J.-T.; Wang, W.-F.; Zhang, Y.; Meng, X.-H.; Sang, C.-Y.; Zhu, L.-L.; Yang, J.-L. Gypenosides (GPs) Alleviates Hypoxia-Induced Injury in PC12 Cells and Enhances Tolerance to Anoxia in C57BL/6 Mice. J. Food Biochem. 2022, 46, e14448. [Google Scholar] [CrossRef]
- Kala, K.; Sivalingam, S.; Gayathri, J.; Devi, Y. Encapsulation of Diosgenin in Chitosan Nanoparticles with Enhanced In Vitro and In Vivo Anticancer Activity in Female Sprague Dawley Rats. Surf. Interfaces 2023, 41, 103158. [Google Scholar] [CrossRef]
- Paarvanova, B.; Tacheva, B.; Savova, G.; Karabaliev, M.; Georgieva, R. Hemolysis by Saponin Is Accelerated at Hypertonic Conditions. Molecules 2023, 28, 7096. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, F.; Kong, D.; Zhu, X.; Chen, W.; Wang, A.; Zheng, S. Saikosaponin D Disrupts Platelet-Derived Growth Factor-β Receptor/P38 Pathway Leading to Mitochondrial Apoptosis in Human LO2 Hepatocyte Cells: A Potential Mechanism of Hepatotoxicity. Chem.-Biol. Interact. 2013, 206, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-H.; Chen, M.-F.; Huang, S.J.; Huang, C.-Y.; Wang, H.-K.; Hsieh, W.-C.; Huang, C.-H.; Liu, L.-F.; Shiu, L.-Y. Saikosaponin a Induces Apoptosis through Mitochondria-Dependent Pathway in Hepatic Stellate Cells. Am. J. Chin. Med. 2017, 45, 351–368. [Google Scholar] [CrossRef]
- Sun, X.-C.; Song, X.; Guo, F.; Yuan, Y.-H.; Wang, S.-Y.; Wang, S.; Liu, K.-L.; Lv, X.-Y.; Han, B.; Zhang, C.; et al. Terrestrosin D, a Spirostanol Saponin from Tribulus terrestris L. with Potential Hepatorenal Toxicity. J. Ethnopharmacol. 2022, 283, 114716. [Google Scholar] [CrossRef] [PubMed]
- Qin, T.; Fu, X.; Yu, J.; Zhang, R.; Deng, X.; Fu, Q.; Ma, Z.; Ma, S. Modification of GSK3β/β-Catenin Signaling on Saikosaponins-d-Induced Inhibition of Neural Progenitor Cell Proliferation and Adult Neurogenesis. Toxicology 2019, 424, 152233. [Google Scholar] [CrossRef] [PubMed]
- Chan, L.Y.; Chiu, P.Y.; Lau, T.K. Embryotoxicity Study of Ginsenoside Rc and Re in In Vitro Rat Whole Embryo Culture. Reprod. Toxicol. 2004, 19, 131–134. [Google Scholar] [CrossRef]
- Hundahl, C.; Kotzbeck, P.; Burm, H.B.; Christiansen, S.H.; Torz, L.; Helge, A.W.; Madsen, M.P.; Ratner, C.; Serup, A.K.; Thompson, J.J.; et al. Hypothalamic Hormone-Sensitive Lipase Regulates Appetite and Energy Homeostasis. Mol. Metab. 2021, 47, 101174. [Google Scholar] [CrossRef]
- Peris-Sampedro, F.; Stoltenborg, I.; Le May, M.V.; Zigman, J.M.; Adan, R.A.H.; Dickson, S.L. Genetic Deletion of the Ghrelin Receptor (GHSR) Impairs Growth and Blunts Endocrine Response to Fasting in Ghsr-IRES-Cre Mice. Mol. Metab. 2021, 51, 101223. [Google Scholar] [CrossRef]
- Jeong, B.; Kim, K.-K.; Lee, T.-H.; Kim, H.-R.; Park, B.-S.; Park, J.-W.; Jeong, J.-K.; Seong, J.-Y.; Lee, B.-J. Spexin Regulates Hypothalamic Leptin Action on Feeding Behavior. Biomolecules 2022, 12, 236. [Google Scholar] [CrossRef]
- Jia, W.; Peng, J.; Zhang, Y.; Zhu, J.; Qiang, X.; Zhang, R.; Shi, L. Amelioration Impact of Gut-Brain Communication on Obesity Control by Regulating Gut Microbiota Composition through the Ingestion of Animal-Plant-Derived Peptides and Dietary Fiber: Can Food Reward Effect as a Hidden Regulator? Crit. Rev. Food Sci. Nutr. 2023, 1–15, Online ahead of print. [Google Scholar] [CrossRef]
- Camilleri, M.; Lupianez-Merly, C. Effects of GLP-1 and Other Gut Hormone Receptors on the Gastrointestinal Tract and Implications in Clinical Practice. Am. J. Gastroenterol. 2023. Online ahead of print. [Google Scholar] [CrossRef]
- Shin, N.R.; Bose, S.; Choi, Y.; Kim, Y.-M.; Chin, Y.-W.; Song, E.-J.; Nam, Y.-D.; Kim, H. Anti-Obesity Effect of Fermented Panax Notoginseng Is Mediated Via Modulation of Appetite and Gut Microbial Population. Front. Pharmacol. 2021, 12, 665881. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Hahm, D.H.; Yang, D.C.; Kim, J.H.; Lee, H.J.; Shim, I. Effect of Crude Saponin of Korean Red Ginseng on High-Fat Diet-Induced Obesity in the Rat. J. Pharmacol. Sci. 2005, 97, 124–131. [Google Scholar] [CrossRef]
- Kim, J.H.; Kang, S.A.; Han, S.-M.; Shim, I. Comparison of the Antiobesity Effects of the Protopanaxadiol- and Protopanaxatriol-Type Saponins of Red Ginseng. Phytother. Res. 2009, 23, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Gao, Y.; Shi, H.-L.; Qin, L.-Y.; Huang, F.; Lan, Y.-Y.; Zhang, B.-B.; Hu, Z.-B.; Wu, X.-J. Astragaloside IV Improves Lipid Metabolism in Obese Mice by Alleviation of Leptin Resistance and Regulation of Thermogenic Network. Sci. Rep. 2016, 6, 30190. [Google Scholar] [CrossRef] [PubMed]
- Hamao, M.; Matsuda, H.; Nakamura, S.; Nakashima, S.; Semura, S.; Maekubo, S.; Wakasugi, S.; Yoshikawa, M. Anti-Obesity Effects of the Methanolic Extract and Chakasaponins from the Flower Buds of Camellia Sinensis in Mice. Bioorg. Med. Chem. 2011, 19, 6033–6041. [Google Scholar] [CrossRef]
- Elmaleh-Sachs, A.; Schwartz, J.L.; Bramante, C.T.; Nicklas, J.M.; Gudzune, K.A.; Jay, M. Obesity Management in Adults: A Review. JAMA 2023, 330, 2000–2015. [Google Scholar] [CrossRef]
- Sanneur, K.; Leksawasdi, N.; Sumonsiri, N.; Techapun, C.; Taesuwan, S.; Nunta, R.; Khemacheewakul, J. Inhibitory Effects of Saponin-Rich Extracts from Pouteria Cambodiana against Digestive Enzymes α-Glucosidase and Pancreatic Lipase. Foods 2023, 12, 3738. [Google Scholar] [CrossRef] [PubMed]
- Navarro Del Hierro, J.; Casado-Hidalgo, G.; Reglero, G.; Martin, D. The Hydrolysis of Saponin-Rich Extracts from Fenugreek and Quinoa Improves Their Pancreatic Lipase Inhibitory Activity and Hypocholesterolemic Effect. Food Chem. 2021, 338, 128113. [Google Scholar] [CrossRef] [PubMed]
- Yoshizumi, K.; Hirano, K.; Ando, H.; Hirai, Y.; Ida, Y.; Tsuji, T.; Tanaka, T.; Satouchi, K.; Terao, J. Lupane-Type Saponins from Leaves of Acanthopanax Sessiliflorus and Their Inhibitory Activity on Pancreatic Lipase. J. Agric. Food Chem. 2006, 54, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Angin, Y.; Beauloye, C.; Horman, S.; Bertrand, L. Regulation of Carbohydrate Metabolism, Lipid Metabolism, and Protein Metabolism by AMPK. Exp. Suppl. 2016, 107, 23–43. [Google Scholar] [CrossRef] [PubMed]
- Sarjeant, K.; Stephens, J.M. Adipogenesis. Cold Spring Harb. Perspect. Biol. 2012, 4, a008417. [Google Scholar] [CrossRef] [PubMed]
- Straub, L.G.; Scherer, P.E. Metabolic Messengers: Adiponectin. Nat. Metab. 2019, 1, 334–339. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Sheng, W.; Gao, R.; Feng, J.; Huang, W.; Cui, L.; Liu, J.; Li, Y. Ethanolic Extract of Root from Arctium lappa L Ameliorates Obesity and Hepatic Steatosis in Rats by Regulating the AMPK/ACC/CPT-1 Pathway. J. Food Biochem. 2022, 46, e14455. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ai, Z.; Xing, X.; Fan, Y.; Zhang, Y.; Nan, B.; Li, X.; Wang, Y.; Liu, J. The Ameliorative Effect of Probiotics on Diet-Induced Lipid Metabolism Disorders: A Review. Crit. Rev. Food Sci. Nutr. 2024, 64, 3556–3572. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.-T.; Wang, Y.-S.; Chou, H.-C.; Chang, C.-C.; Lee, C.-K.; Juan, S.-H. Kinsenoside-Mediated Lipolysis through an AMPK-Dependent Pathway in C3H10T1/2 Adipocytes: Roles of AMPK and PPARα in the Lipolytic Effect of Kinsenoside. Phytomedicine 2015, 22, 641–647. [Google Scholar] [CrossRef]
- Sun, C.; Mao, S.; Chen, S.; Zhang, W.; Liu, C. PPARs-Orchestrated Metabolic Homeostasis in the Adipose Tissue. Int. J. Mol. Sci. 2021, 22, 8974. [Google Scholar] [CrossRef]
- Zhang, T.; Zhong, S.; Li, T.; Zhang, J. Saponins as Modulators of Nuclear Receptors. Crit. Rev. Food Sci. Nutr. 2020, 60, 94–107. [Google Scholar] [CrossRef]
- Qin, R.; Zhang, J.; Li, C.; Zhang, X.; Xiong, A.; Huang, F.; Yin, Z.; Li, K.; Qin, W.; Chen, M.; et al. Protective Effects of Gypenosides against Fatty Liver Disease Induced by High Fat and Cholesterol Diet and Alcohol in Rats. Arch. Pharm. Res. 2012, 35, 1241–1250. [Google Scholar] [CrossRef] [PubMed]
- Malliou, F.; Andreadou, I.; Gonzalez, F.J.; Lazou, A.; Xepapadaki, E.; Vallianou, I.; Lambrinidis, G.; Mikros, E.; Marselos, M.; Skaltsounis, A.-L.; et al. The Olive Constituent Oleuropein, as a PPARα Agonist, Markedly Reduces Serum Triglycerides. J. Nutr. Biochem. 2018, 59, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xiao, Y.; Gao, W.; Pan, J.; Zhao, Q.; Zhang, Z. Gymnemic Acid Alleviates Inflammation and Insulin Resistance via PPARδ- and NFκB-Mediated Pathways in Db/Db Mice. Food Funct. 2019, 10, 5853–5862. [Google Scholar] [CrossRef]
- Liu, H.; Liu, M.; Jin, Z.; Yaqoob, S.; Zheng, M.; Cai, D.; Liu, J.; Guo, S. Ginsenoside Rg2 Inhibits Adipogenesis in 3T3-L1 Preadipocytes and Suppresses Obesity in High-Fat-Diet-Induced Obese Mice through the AMPK Pathway. Food Funct. 2019, 10, 3603–3614. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.H.; Ahn, E.-K.; Lee, J.A.; Shin, T.-S.; Tsukamoto, C.; Suh, J.; Mei, I.; Chung, G. Soyasaponins Aa and Ab Exert an Anti-Obesity Effect in 3T3-L1 Adipocytes through Downregulation of PPARγ. Phytother. Res. 2015, 29, 281–287. [Google Scholar] [CrossRef]
- Kim, K.; Nam, K.H.; Yi, S.A.; Park, J.W.; Han, J.-W.; Lee, J. Ginsenoside Rg3 Induces Browning of 3T3-L1 Adipocytes by Activating AMPK Signaling. Nutrients 2020, 12, 427. [Google Scholar] [CrossRef]
- Mu, Q.; Fang, X.; Li, X.; Zhao, D.; Mo, F.; Jiang, G.; Yu, N.; Zhang, Y.; Guo, Y.; Fu, M.; et al. Ginsenoside Rb1 Promotes Browning through Regulation of PPARγ in 3T3-L1 Adipocytes. Biochem. Biophys. Res. Commun. 2015, 466, 530–535. [Google Scholar] [CrossRef]
- Ramji, D.P.; Foka, P. CCAAT/Enhancer-Binding Proteins: Structure, Function and Regulation. Biochem. J. 2002, 365, 561–575. [Google Scholar] [CrossRef]
- Lee, S.-G.; Lee, Y.-J.; Jang, M.-H.; Kwon, T.-R.; Nam, J.-O. Panax Ginseng Leaf Extracts Exert Anti-Obesity Effects in High-Fat Diet-Induced Obese Rats. Nutrients 2017, 9, 999. [Google Scholar] [CrossRef]
- Gu, W.; Kim, K.-A.; Kim, D.-H. Ginsenoside Rh1 Ameliorates High Fat Diet-Induced Obesity in Mice by Inhibiting Adipocyte Differentiation. Biol. Pharm. Bull. 2013, 36, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Manzoor, M.; Muroi, M.; Ogawa, N.; Kobayashi, H.; Nishimura, H.; Chen, D.; Fasina, O.B.; Wang, J.; Osada, H.; Yoshida, M.; et al. Isoquercitrin from Apocynum venetum L. Produces an Anti-Obesity Effect on Obese Mice by Targeting C-1-Tetrahydrofolate Synthase, Carbonyl Reductase, and Glutathione S-Transferase P and Modification of the AMPK/SREBP-1c/FAS/CD36 Signaling Pathway in Mice In Vivo. Food Funct. 2022, 13, 10923–10936. [Google Scholar] [CrossRef] [PubMed]
- Fang, K.; Wu, F.; Chen, G.; Dong, H.; Li, J.; Zhao, Y.; Xu, L.; Zou, X.; Lu, F. Diosgenin Ameliorates Palmitic Acid-Induced Lipid Accumulation via AMPK/ACC/CPT-1A and SREBP-1c/FAS Signaling Pathways in LO2 Cells. BMC Complement. Altern. Med. 2019, 19, 255. [Google Scholar] [CrossRef]
- Gu, Y.; Duan, S.; Ding, M.; Zheng, Q.; Fan, G.; Li, X.; Li, Y.; Liu, C.; Sun, R.; Liu, R. Saikosaponin D Attenuates Metabolic Associated Fatty Liver Disease by Coordinately Tuning PPARα and INSIG/SREBP1c Pathway. Phytomedicine 2022, 103, 154219. [Google Scholar] [CrossRef] [PubMed]
- Tsai, W.-T.; Nakamura, Y.; Akasaka, T.; Katakura, Y.; Tanaka, Y.; Shirouchi, B.; Jiang, Z.; Yuan, X.; Sato, M. Soyasaponin Ameliorates Obesity and Reduces Hepatic Triacylglycerol Accumulation by Suppressing Lipogenesis in High-Fat Diet-Fed Mice. J. Food Sci. 2021, 86, 2103–2117. [Google Scholar] [CrossRef] [PubMed]
- Renga, B.; Festa, C.; De Marino, S.; Di Micco, S.; D’Auria, M.V.; Bifulco, G.; Fiorucci, S.; Zampella, A. Molecular Decodification of Gymnemic Acids from Gymnema Sylvestre. Discovery of a New Class of Liver X Receptor Antagonists. Steroids 2015, 96, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Albillos, A.; de Gottardi, A.; Rescigno, M. The Gut-Liver Axis in Liver Disease: Pathophysiological Basis for Therapy. J. Hepatol. 2020, 72, 558–577. [Google Scholar] [CrossRef] [PubMed]
- Schoeler, M.; Caesar, R. Dietary Lipids, Gut Microbiota and Lipid Metabolism. Rev. Endocr. Metab. Disord. 2019, 20, 461–472. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Huang, J.; Sun, M.; Jiang, Y.; Wang, S.; Wang, L.; Yu, N.; Peng, D.; Wang, Y.; Chen, W.; et al. Poria Cocos Polysaccharide Improves Intestinal Barrier Function and Maintains Intestinal Homeostasis in Mice. Int. J. Biol. Macromol. 2023, 249, 125953. [Google Scholar] [CrossRef]
- Chopyk, D.M.; Grakoui, A. Contribution of the Intestinal Microbiome and Gut Barrier to Hepatic Disorders. Gastroenterology 2020, 159, 849–863. [Google Scholar] [CrossRef]
- Dong, J.; Liang, W.; Wang, T.; Sui, J.; Wang, J.; Deng, Z.; Chen, D. Saponins Regulate Intestinal Inflammation in Colon Cancer and IBD. Pharmacol. Res. 2019, 144, 66–72. [Google Scholar] [CrossRef]
- Cai, J.; Liu, J.; Fan, P.; Dong, X.; Zhu, K.; Liu, X.; Zhang, N.; Cao, Y. Dioscin Prevents DSS-Induced Colitis in Mice with Enhancing Intestinal Barrier Function and Reducing Colon Inflammation. Int. Immunopharmacol. 2021, 99, 108015. [Google Scholar] [CrossRef]
- Zhang, M.; Zhao, J.; Deng, J.; Duan, Z.; Zhu, C.; Fan, D. The Protective Effect of Protopanaxatriol-Type Saponin on Intestinal Health in Antibiotic-Treated Mice. Food Funct. 2019, 10, 4124–4133. [Google Scholar] [CrossRef]
- Zhao, W.; Xiao, M.; Yang, J.; Zhang, L.; Ba, Y.; Xu, R.; Liu, Z.; Zou, H.; Yu, P.; Wu, X.; et al. The Combination of Ilexhainanoside D and Ilexsaponin A1 Reduces Liver Inflammation and Improves Intestinal Barrier Function in Mice with High-Fat Diet-Induced Non-Alcoholic Fatty Liver Disease. Phytomedicine 2019, 63, 153039. [Google Scholar] [CrossRef]
- Zhang, J.; Ma, X.; Fan, D. Ginsenoside CK Ameliorates Hepatic Lipid Accumulation via Activating the LKB1/AMPK Pathway In Vitro and In Vivo. Food Funct. 2022, 13, 1153–1167. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, L.; Liu, J.; Chan, A.S.C.; Li, D. Saponins of Tomato Extract Improve Non-Alcoholic Fatty Liver Disease by Regulating Oxidative Stress and Lipid Homeostasis. Antioxidants 2023, 12, 1848. [Google Scholar] [CrossRef]
- Ding, R.-B.; Tian, K.; Cao, Y.-W.; Bao, J.-L.; Wang, M.; He, C.; Hu, Y.; Su, H.; Wan, J.-B. Protective Effect of Panax Notoginseng Saponins on Acute Ethanol-Induced Liver Injury Is Associated with Ameliorating Hepatic Lipid Accumulation and Reducing Ethanol-Mediated Oxidative Stress. J. Agric. Food Chem. 2015, 63, 2413–2422. [Google Scholar] [CrossRef]
- Xu, K.; Hu, B.; Ding, X.; Zhan, Z. Alleviation of D-Gal-Induced Senile Liver Injury by Rg3, a Signature Component of Red Ginseng. Aging 2023, 15, 6749–6756. [Google Scholar] [CrossRef]
- O’Riordan, K.J.; Collins, M.K.; Moloney, G.M.; Knox, E.G.; Aburto, M.R.; Fülling, C.; Morley, S.J.; Clarke, G.; Schellekens, H.; Cryan, J.F. Short Chain Fatty Acids: Microbial Metabolites for Gut-Brain Axis Signalling. Mol. Cell Endocrinol. 2022, 546, 111572. [Google Scholar] [CrossRef]
- van der Hee, B.; Wells, J.M. Microbial Regulation of Host Physiology by Short-Chain Fatty Acids. Trends Microbiol. 2021, 29, 700–712. [Google Scholar] [CrossRef]
- Tan, J.; McKenzie, C.; Potamitis, M.; Thorburn, A.N.; Mackay, C.R.; Macia, L. The Role of Short-Chain Fatty Acids in Health and Disease. Adv. Immunol. 2014, 121, 91–119. [Google Scholar] [CrossRef]
- He, J.; Zhang, P.; Shen, L.; Niu, L.; Tan, Y.; Chen, L.; Zhao, Y.; Bai, L.; Hao, X.; Li, X.; et al. Short-Chain Fatty Acids and Their Association with Signalling Pathways in Inflammation, Glucose and Lipid Metabolism. Int. J. Mol. Sci. 2020, 21, 6356. [Google Scholar] [CrossRef]
- Kim, M.H.; Kang, S.G.; Park, J.H.; Yanagisawa, M.; Kim, C.H. Short-Chain Fatty Acids Activate GPR41 and GPR43 on Intestinal Epithelial Cells to Promote Inflammatory Responses in Mice. Gastroenterology 2013, 145, 396–406.e10. [Google Scholar] [CrossRef]
- Zou, F.; Qiu, Y.; Huang, Y.; Zou, H.; Cheng, X.; Niu, Q.; Luo, A.; Sun, J. Effects of Short-Chain Fatty Acids in Inhibiting HDAC and Activating P38 MAPK Are Critical for Promoting B10 Cell Generation and Function. Cell Death Dis. 2021, 12, 582. [Google Scholar] [CrossRef]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef]
- Lan, Y.; Sun, Q.; Ma, Z.; Peng, J.; Zhang, M.; Wang, C.; Zhang, X.; Yan, X.; Chang, L.; Hou, X.; et al. Seabuckthorn Polysaccharide Ameliorates High-Fat Diet-Induced Obesity by Gut Microbiota-SCFAs-Liver Axis. Food Funct. 2022, 13, 2925–2937. [Google Scholar] [CrossRef]
- den Besten, G.; Bleeker, A.; Gerding, A.; van Eunen, K.; Havinga, R.; van Dijk, T.H.; Oosterveer, M.H.; Jonker, J.W.; Groen, A.K.; Reijngoud, D.-J.; et al. Short-Chain Fatty Acids Protect Against High-Fat Diet-Induced Obesity via a PPARγ-Dependent Switch from Lipogenesis to Fat Oxidation. Diabetes 2015, 64, 2398–2408. [Google Scholar] [CrossRef]
- Yin, Y.; Sichler, A.; Ecker, J.; Laschinger, M.; Liebisch, G.; Höring, M.; Basic, M.; Bleich, A.; Zhang, X.-J.; Kübelsbeck, L.; et al. Gut Microbiota Promote Liver Regeneration through Hepatic Membrane Phospholipid Biosynthesis. J. Hepatol. 2023, 78, 820–835. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, T.; Yan, C.; Xie, R.; Guo, Z.; Wang, S.; Zhang, Y.; Li, Z.; Wang, B.; Cao, H. Diammonium Glycyrrhizinate Protects against Nonalcoholic Fatty Liver Disease in Mice through Modulation of Gut Microbiota and Restoration of Intestinal Barrier. Mol. Pharm. 2018, 15, 3860–3870. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Peng, Y.; Zhao, L.; Zhou, G.; Li, X. Regulating the Gut Microbiota and SCFAs in the Faeces of T2DM Rats Should Be One of Antidiabetic Mechanisms of Mogrosides in the Fruits of Siraitia grosvenorii. J. Ethnopharmacol. 2021, 274, 114033. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Yang, X.; Yang, Z.; Huang, W.; Kou, J.; Li, F. Akebia Saponin D Regulates the Metabolome and Intestinal Microbiota in High Fat Diet-Induced Hyperlipidemic Rats. Molecules 2019, 24, 1268. [Google Scholar] [CrossRef] [PubMed]
- Fu, K.; Ma, C.; Wang, C.; Zhou, H.; Gong, L.; Zhang, Y.; Li, Y. Forsythiaside A Alleviated Carbon Tetrachloride-Induced Liver Fibrosis by Modulating Gut Microbiota Composition to Increase Short-Chain Fatty Acids and Restoring Bile Acids Metabolism Disorder. Biomed. Pharmacother. 2022, 151, 113185. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Song, Y.; Xu, W.; Chen, J.; Zhou, R.; Yang, M.; Zhu, G.; Luo, X.; Ai, Z.; Liu, Y.; et al. Pulsatilla Chinensis Saponins Improve SCFAs Regulating GPR43-NLRP3 Signaling Pathway in the Treatment of Ulcerative Colitis. J. Ethnopharmacol. 2023, 308, 116215. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Zhao, J.; Liu, C.; Zhang, Z.; Zhang, Y.; Meng, D. Xanthoceraside Exerts Anti-Alzheimer’s Disease Effect by Remodeling Gut Microbiota and Modulating Microbial-Derived Metabolites Level in Rats. Phytomedicine 2022, 98, 153937. [Google Scholar] [CrossRef]
- Cai, J.; Sun, L.; Gonzalez, F.J. Gut Microbiota-Derived Bile Acids in Intestinal Immunity, Inflammation, and Tumorigenesis. Cell Host Microbe 2022, 30, 289–300. [Google Scholar] [CrossRef]
- Lin, S.; Wang, S.; Wang, P.; Tang, C.; Wang, Z.; Chen, L.; Luo, G.; Chen, H.; Liu, Y.; Feng, B.; et al. Bile Acids and Their Receptors in Regulation of Gut Health and Diseases. Prog. Lipid Res. 2023, 89, 101210. [Google Scholar] [CrossRef] [PubMed]
- Vaz, F.M.; Ferdinandusse, S. Bile Acid Analysis in Human Disorders of Bile Acid Biosynthesis. Mol. Asp. Med. 2017, 56, 10–24. [Google Scholar] [CrossRef]
- Xiang, J.; Zhang, Z.; Xie, H.; Zhang, C.; Bai, Y.; Cao, H.; Che, Q.; Guo, J.; Su, Z. Effect of Different Bile Acids on the Intestine through Enterohepatic Circulation Based on FXR. Gut Microbes 2021, 13, 1949095. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Zhang, H.; Xiao, D.; Wei, H.; Chen, Y. Farnesoid X Receptor (FXR): Structures and Ligands. Comput. Struct. Biotechnol. J. 2021, 19, 2148–2159. [Google Scholar] [CrossRef]
- Tuominen, I.; Beaven, S.W. Intestinal Farnesoid X Receptor Puts a Fresh Coat of Wax on Fatty Liver. Hepatology 2015, 62, 646–648. [Google Scholar] [CrossRef]
- Xie, C.; Jiang, C.; Shi, J.; Gao, X.; Sun, D.; Sun, L.; Wang, T.; Takahashi, S.; Anitha, M.; Krausz, K.W.; et al. An Intestinal Farnesoid X Receptor–Ceramide Signaling Axis Modulates Hepatic Gluconeogenesis in Mice. Diabetes 2016, 66, 613–626. [Google Scholar] [CrossRef] [PubMed]
- Simbrunner, B.; Trauner, M.; Reiberger, T. Review Article: Therapeutic Aspects of Bile Acid Signalling in the Gut-Liver Axis. Aliment. Pharmacol. Ther. 2021, 54, 1243–1262. [Google Scholar] [CrossRef] [PubMed]
- Kwong, E.; Li, Y.; Hylemon, P.B.; Zhou, H. Bile Acids and Sphingosine-1-Phosphate Receptor 2 in Hepatic Lipid Metabolism. Acta Pharm. Sin. B 2015, 5, 151–157. [Google Scholar] [CrossRef]
- Perino, A.; Pols, T.W.H.; Nomura, M.; Stein, S.; Pellicciari, R.; Schoonjans, K. TGR5 Reduces Macrophage Migration through mTOR-Induced C/EBPβ Differential Translation. J. Clin. Investig. 2014, 124, 5424–5436. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhuang, T.; Wang, Z.; Wang, X.; Liu, L.; Luo, Y.; Wang, R.; Li, L.; Huang, W.; Wang, Z.; et al. Red Ginseng Extracts Ameliorate High-Fat Diet-Induced Obesity and Insulin Resistance by Activating the Intestinal TGR5-Mediated Bile Acids Signaling Pathway. Phytomedicine 2023, 119, 154982. [Google Scholar] [CrossRef] [PubMed]
- Jadhav, K.; Xu, Y.; Xu, Y.; Li, Y.; Xu, J.; Zhu, Y.; Adorini, L.; Lee, Y.K.; Kasumov, T.; Yin, L.; et al. Reversal of Metabolic Disorders by Pharmacological Activation of Bile Acid Receptors TGR5 and FXR. Mol. Metab. 2018, 9, 131–140. [Google Scholar] [CrossRef]
- Zhao, X.-Y.; Xiong, X.; Liu, T.; Mi, L.; Peng, X.; Rui, C.; Guo, L.; Li, S.; Li, X.; Lin, J.D. Long Noncoding RNA Licensing of Obesity-Linked Hepatic Lipogenesis and NAFLD Pathogenesis. Nat. Commun. 2018, 9, 2986. [Google Scholar] [CrossRef]
- Liu, Y.; Jin, Z.-Y.; Wang, J.-X.; Wang, D.; Liu, H.; Li, D.; Zhu, J.; Luo, Z.-B.; Han, S.-Z.; Chang, S.-Y.; et al. Ginsenoside Rg1 Activates Brown Adipose Tissue to Counteract Obesity in High-Fat Diet-Fed Mice by Regulating Gut Microbes and Bile Acid Composition. Food Funct. 2023, 14, 4696–4705. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Man, S.; Liang, Y.; Ma, L.; Guo, L.; Huang, L.; Gao, W. Diosgenin Alleviates Nonalcoholic Steatohepatitis through Affecting Liver-Gut Circulation. Pharmacol. Res. 2023, 187, 106621. [Google Scholar] [CrossRef]
- Feng, J.; Ma, H.; Yue, Y.; Wang, L.; Hao, K.; Zhang, Y.; Li, J.; Xiang, Y.; Min, Y. Saikosaponin a Ameliorates Diet-Induced Fatty Liver via Regulating Intestinal Microbiota and Bile Acid Profile in Laying Hens. Poult. Sci. 2023, 102, 103155. [Google Scholar] [CrossRef]
- Malinow, M.R.; Connor, W.E.; McLaughlin, P.; Stafford, C.; Lin, D.S.; Livingston, A.L.; Kohler, G.O.; McNulty, W.P. Cholesterol and Bile Acid Balance in Macaca Fascicularis. Effects of Alfalfa Saponins. J. Clin. Investig. 1981, 67, 156–162. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Tan, C.-P.; Xu, Y.-J.; Liu, Y. Gut Microbiota-Derived Trimethylamine-N-Oxide: A Bridge between Dietary Fatty Acid and Cardiovascular Disease? Food Res. Int. 2020, 138, 109812. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Chang, M.; Guo, Y.; Zhang, L.; Xue, C.; Yanagita, T.; Zhang, T.; Wang, Y. Trimethylamine-N-Oxide (TMAO)-Induced Atherosclerosis Is Associated with Bile Acid Metabolism. Lipids Health Dis. 2018, 17, 286. [Google Scholar] [CrossRef]
- Gao, X.; Liu, X.; Xu, J.; Xue, C.; Xue, Y.; Wang, Y. Dietary Trimethylamine N-Oxide Exacerbates Impaired Glucose Tolerance in Mice Fed a High Fat Diet. J. Biosci. Bioeng. 2014, 118, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Guo, X.; Xie, C.; Cao, Z.; Wang, X.; Liu, L.; Yang, P. Unraveling the Metabolic Pathway of Choline-TMA-TMAO: Effects of Gypenosides and Implications for the Therapy of TMAO Related Diseases. Pharmacol. Res. 2021, 173, 105884. [Google Scholar] [CrossRef] [PubMed]
- Xue, P.; Yang, X.; Zhao, L.; Hou, Z.; Zhang, R.; Zhang, F.; Ren, G. Relationship between Antimicrobial Activity and Amphipathic Structure of Ginsenosides. Ind. Crops Prod. 2020, 143, 111929. [Google Scholar] [CrossRef]
- Yu, H.; Chai, X.; Geng, W.-C.; Zhang, L.; Ding, F.; Guo, D.-S.; Wang, Y. Facile and Label-Free Fluorescence Strategy for Evaluating the Influence of Bioactive Ingredients on FMO3 Activity via Supramolecular Host-Guest Reporter Pair. Biosens. Bioelectron. 2021, 192, 113488. [Google Scholar] [CrossRef]
- Yu, T.; Wang, Y.; Chen, X.; Xiong, W.; Tang, Y.; Lin, L. Spirulina Platensis Alleviates Chronic Inflammation with Modulation of Gut Microbiota and Intestinal Permeability in Rats Fed a High-Fat Diet. J. Cell. Mol. Med. 2020, 24, 8603–8613. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Wang, L.; Wang, S.; Wang, J.; Su, C.; Zhang, L.; Li, C.; Liu, S. Phenolics from Noni (Morinda citrifolia L.) Fruit Alleviate Obesity in High Fat Diet-Fed Mice via Modulating the Gut Microbiota and Mitigating Intestinal Damage. Food Chem. 2023, 402, 134232. [Google Scholar] [CrossRef]
- Shen, S.; Wang, K.; Zhi, Y.; Shen, W.; Huang, L. Gypenosides Improves Nonalcoholic Fatty Liver Disease Induced by High-Fat Diet Induced through Regulating LPS/TLR4 Signaling Pathway. Cell Cycle 2020, 19, 3042–3053. [Google Scholar] [CrossRef]
- Wang, P.-F.; Xie, K.; Cao, Y.-X.; Zhang, A. Hepatoprotective Effect of Mitochondria-Targeted Antioxidant Mito-TEMPO against Lipopolysaccharide-Induced Liver Injury in Mouse. Mediat. Inflamm. 2022, 2022, e6394199. [Google Scholar] [CrossRef] [PubMed]
- Lebrun, L.J.; Pallot, G.; Nguyen, M.; Tavernier, A.; Dusuel, A.; Pilot, T.; Deckert, V.; Dugail, I.; Le Guern, N.; Pais De Barros, J.-P.; et al. Increased Weight Gain and Insulin Resistance in HF-Fed PLTP Deficient Mice Is Related to Altered Inflammatory Response and Plasma Transport of Gut-Derived LPS. Int. J. Mol. Sci. 2022, 23, 13226. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Seki, E. An Intestine-Derived HDL as a Novel Regulator of the Activity of Gut-Derived LPS: Ushering in a New Era of Research on the Gut-Liver Axis. Gastroenterology 2022, 162, 651–652. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Liu, S.; Kong, L.; Wu, S.; Zhong, Z.; Yu, L.; Yang, Q.; Zhang, J.; Li, J.; Zheng, G. Lipopolysaccharide-Induced Persistent Inflammation Ameliorates Fat Accumulation by Promoting Adipose Browning In Vitro and In Vivo. Int. J. Biol. Macromol. 2023, 252, 126511. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wang, N.; Tan, H.-Y.; Li, S.; Zhang, C.; Feng, Y. Gut-Liver Axis Modulation of Panax Notoginseng Saponins in Nonalcoholic Fatty Liver Disease. Hepatol. Int. 2021, 15, 350–365. [Google Scholar] [CrossRef]
- Yu, Y.; Cui, S.; Zheng, T.; Ma, H.; Xie, Z.; Jiang, H.; Li, Y.; Zhu, K.; Huang, C.; Li, J.; et al. Sarsasapogenin Improves Adipose Tissue Inflammation and Ameliorates Insulin Resistance in High-Fat Diet-Fed C57BL/6J Mice. Acta Pharmacol. Sin. 2021, 42, 272–281. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Saponin | Pharmacological Properties | Model | References |
|---|---|---|---|
| Garlic saponins | Lipid lowering, lowering of cholesterol | mice | [10] |
| Ginsenosides | Anticancer activity | mice | [10] |
| Aralia taibaiensis saponins | Antioxidant, anti-aging, organ protection | rat | [17] |
| Gnsenoside Rb2 | Improvement of fatty liver disease | mice | [18] |
| Gnsenoside Rg3 | Anticancer activity | mice | [19] |
| Gnsenoside Rk3 | Lowering blood glucose, improving insulin resistance | mice | [20] |
| Sarsasapogenin | Improvement of rheumatoid arthritis | rat | [21] |
| Kuding tea saponins | Decrease serum lipids, improve lipid metabolism | mice | [22] |
| Gypenosides | Enhancement of hypoxia tolerance | mice | [23] |
| Diosgenin | Anticancer activity | rat | [24] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, S.; Liu, M.; Han, Y.; Li, S.; Zhu, X.; Li, D.; Shi, Y.; Liu, B. Effects of Saponins on Lipid Metabolism: The Gut–Liver Axis Plays a Key Role. Nutrients 2024, 16, 1514. https://doi.org/10.3390/nu16101514
Cao S, Liu M, Han Y, Li S, Zhu X, Li D, Shi Y, Liu B. Effects of Saponins on Lipid Metabolism: The Gut–Liver Axis Plays a Key Role. Nutrients. 2024; 16(10):1514. https://doi.org/10.3390/nu16101514
Chicago/Turabian StyleCao, Shixi, Mengqi Liu, Yao Han, Shouren Li, Xiaoyan Zhu, Defeng Li, Yinghua Shi, and Boshuai Liu. 2024. "Effects of Saponins on Lipid Metabolism: The Gut–Liver Axis Plays a Key Role" Nutrients 16, no. 10: 1514. https://doi.org/10.3390/nu16101514
APA StyleCao, S., Liu, M., Han, Y., Li, S., Zhu, X., Li, D., Shi, Y., & Liu, B. (2024). Effects of Saponins on Lipid Metabolism: The Gut–Liver Axis Plays a Key Role. Nutrients, 16(10), 1514. https://doi.org/10.3390/nu16101514