
Flavonoid Metabolites in Serum and Urine after the Ingestion of Selected Tropical Fruits

Abstract
1. Introduction
2. Materials and Methods
2.1. Fruit Samples
2.2. Participants
2.3. Study Design
2.4. Sample Preparation
2.5. LC/ESI-MS/MS Analysis
2.6. Calibration Curve and Method Validation
2.7. Data and Statistical Analysis
3. Results
3.1. Mass Chromatograms and Flavonoid Profiles in Serum
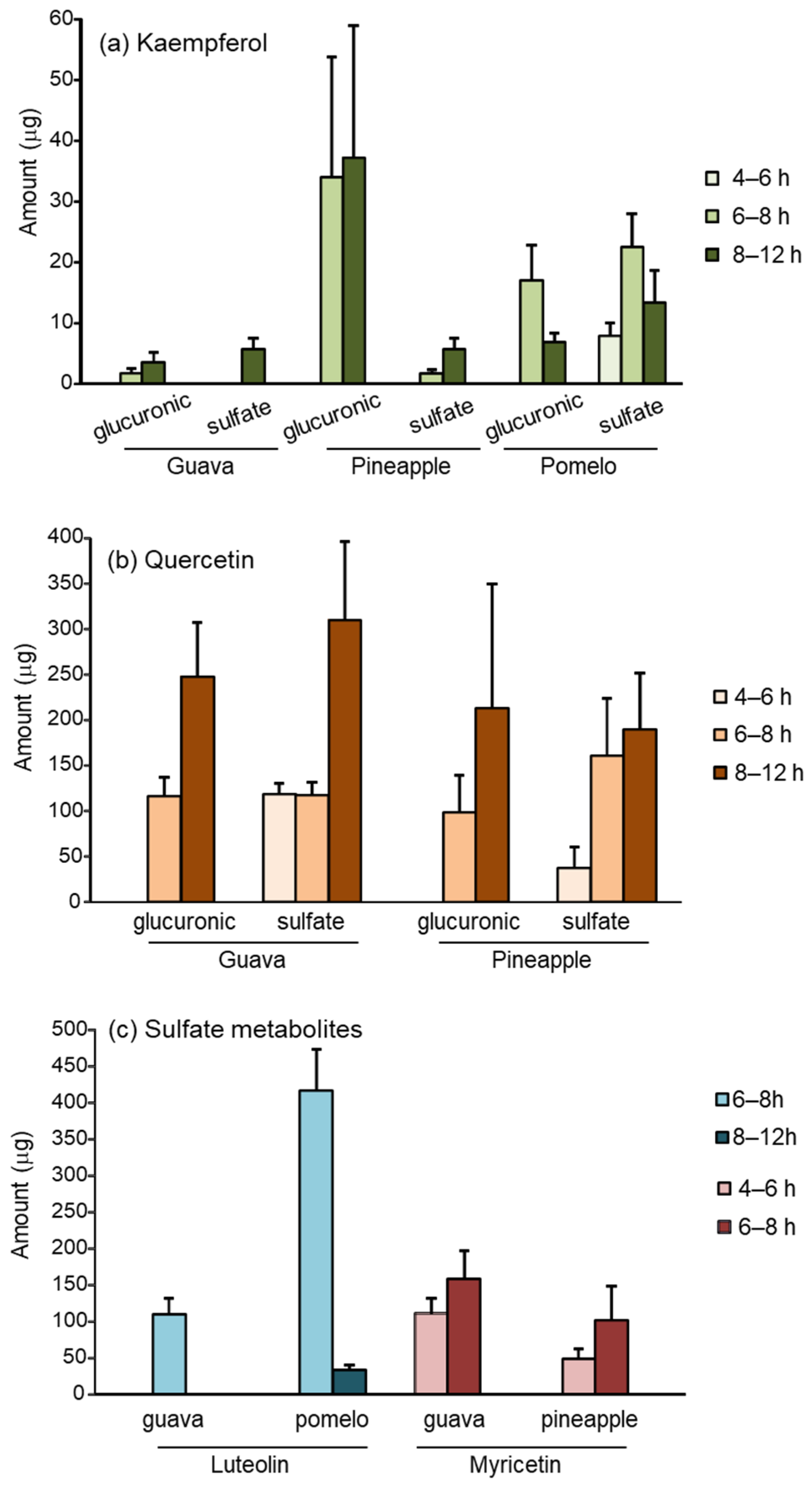
3.2. Flavonoid Excretion in Urine
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shen, N.; Wang, T.; Gan, Q.; Liu, S.; Wang, L.; Jin, B. Plant flavonoids: Classification, distribution, biosynthesis, and antioxidant activity. Food Chem. 2022, 383, 132531. [Google Scholar] [CrossRef] [PubMed]
- Maleki, S.J.; Crespo, J.F.; Cabanillas, B. Anti-inflammatory effects of flavonoids. Food Chem. 2019, 299, 125124. [Google Scholar] [CrossRef] [PubMed]
- Micek, A.; Godos, J.; Del Rio, D.; Galvano, F.; Grosso, G. Dietary flavonoids and cardiovascular Disease: A comprehensive dose-response meta-analysis. Mol. Nutr. Food Res. 2021, 65, e2001019. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.F.; Ruan, Y.; Li, Z.H.; Li, D. Flavonoid subclasses and type 2 diabetes mellitus risk: A meta-analysis of prospective cohort studies. Crit. Rev. Food Sci. Nutr. 2019, 59, 2850–2862. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Dong, J.Y.; Cui, R.; Muraki, I.; Yamagishi, K.; Sawada, N.; Iso, H.; Tsugane, S.; Japan Public Health Center-based prospective study group. Consumption of flavonoid-rich fruits, flavonoids from fruits and stroke risk: A prospective cohort study. Br. J. Nutr. 2021, 126, 1717–1724. [Google Scholar] [CrossRef] [PubMed]
- He, F.J.; Nowson, C.A.; Macgregor, G.A. Fruit and vegetables consumption and stroke: A meta-analysis of cohort studies. Lancet 2006, 367, 320–326. [Google Scholar] [CrossRef]
- Sak, K. Intake of individual flavonoids and risk of carcinogenesis: An overview of epidemiological evidence. Nutr. Cancer 2017, 69, 1119–1150. [Google Scholar] [CrossRef]
- Zhang, X.; Molsberry, S.A.; Yeh, T.S.; Cassidy, A.; Schwarzschild, M.A.; Ascherio, A.; Gao, X. Intake of flavonoids and flavonoid-rich foods and mortality risk among individuals with Parkinson disease: A prospective cohort study. Neurology 2022, 98, e1064–e1076. [Google Scholar] [CrossRef]
- Chen, L.; Cao, H.; Huang, Q.; Xiao, J.; Teng, H. Absorption, metabolism and bioavailability of flavonoids: A review. Crit Rev Food Sci. Nutr. 2022, 62, 7730–7742. [Google Scholar] [CrossRef]
- Kongkachuichai, R.; Charoensiri, R.; Sungpuag, P. Carotenoid, flavonoid profiles and dietary fiber contents of fruits commonly consumed in Thailand. Int. J. Food Sci. Nutr. 2010, 61, 536–548. [Google Scholar] [CrossRef]
- Hussain, S.; Naseer, B.; Qadri, T.; Fatima, T.; Bhat, T. Guava (Psidium Guajava)-Morphology, Taxonomy, Composition and Health Benefits. In Fruits Grown in Highland Regions of the Himalayas, Nutritional and Health Benefits; Springer Nature: Cham, Switzerland, 2021; pp. 257–267. [Google Scholar] [CrossRef]
- Makkumrai, W.; Huang, Y.; Xu, Q. Comparison of pomelo (Citrus maxima) grown in China and Thailand. Front. Agric. Sci. Eng. 2021, 8, 335–352. [Google Scholar] [CrossRef]
- Rattanathawornkiti, K.; Kanchanaketu, T.; Suwanagul, A.; Hongtrakul, V. Genetic relationship assessment of pineapple germplasm in Thailand revealed by AFLP markers. GaG 2016, 9, 56–65. [Google Scholar] [CrossRef]
- Ishii, K.; Furuta, T.; Kasuya, Y. High-performance liquid chromatographic determination of quercetin in human plasma and urine utilizing solid-phase extraction and ultraviolet detection. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2003, 794, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Biesaga, M. Influence of extraction methods on stability of flavonoids. J. Chromatogr. A 2011, 1218, 2505–2512. [Google Scholar] [CrossRef] [PubMed]
- Mullen, W.; Hartley, R.C.; Crozier, A. Flavonoids metabolites in human plasma and urine after the consumption of red onions analyses by liquid chromatography with photodiode array and tandem mass spectrometric detection. J. Chromatpgr. A 2004, 1058, 163–168. [Google Scholar] [CrossRef]
- Hollman, P.C.; van Trijp, J.M.; Buysman, M.N.; van der Gaag, M.S.; Mengelers, M.J.; de Vries, J.H.; Katan, M.B. Relative bioavailability of the antioxidant flavonoid quercetin from various foods in man. FEBS Lett. 1997, 418, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Maliakal, P.; Lu, H.; Lee, M.J.; Yang, C.S. Urinary and plasma levels of resveratrol and quercetin in humans, mice, and rats after ingestion of pure compounds and grape juice. J. Agric. Food Chem. 2004, 52, 935–942. [Google Scholar] [CrossRef] [PubMed]
- Moon, Y.J.; Wang, L.; DiCenzo, R.; Morris, M.E. Quercetin pharmacokinetics in humans. Biopharm. Drug Dispos. 2008, 29, 205–217. [Google Scholar] [CrossRef]
- Kaushik, D.; O’Fallon, K.; Clarkson, P.M.; Dunne, C.P.; Conca, K.R.; Michniak-Kohn, B. Comparison of quercetin pharmacokinetics following oral supplementation in humans. J. Food Sci. 2012, 77, H231–H238. [Google Scholar] [CrossRef]
- Graefe, E.U.; Wittig, J.; Mueller, S.; Riethling, A.K.; Uehleke, B.; Drewelow, B.; Pforte, H.; Jacobasch, G.; Derendorf, H.; Veit, M. Pharmacokinetics and bioavailability of quercetin glycosides in humans. J. Clin. Pharmacol. 2001, 41, 492–499. [Google Scholar] [CrossRef]
- Breiter, T.; Laue, C.; Kressel, G.; Gröll, S.; Engelhardt, U.H.; Hahn, A. Bioavailability and antioxidant potential of rooibos flavonoids in humans following the consumption of different rooibos formulations. Food Chem. 2011, 128, 338–347. [Google Scholar] [CrossRef] [PubMed]
- Palafox-Carlos, H.; Ayala-Zavala, J.F.; González-Aguilar, G.A. The role of dietary fiber in the bioaccessibility and bioavailability of fruit and vegetable antioxidants. J. Food Sci. 2011, 76, R6–R15. [Google Scholar] [CrossRef] [PubMed]
- Prasain, J.K.; Barnes, S. Metabolism and bioavailability of flavonoids in chemoprevention: Current analytical strategies and future prospectus. Mol. Pharm. 2007, 4, 846–864. [Google Scholar] [CrossRef]
- Scalbert, A.; Williamson, G. Dietary intake and bioavailability of polyphenols. J. Nutr. 2000, 130, 2073S–2085S. [Google Scholar] [CrossRef]
- Tang, D.; Chen, K.; Huang, L.; Li, J. Pharmacokinetic properties and drug interactions of apigenin, a natural flavone. Expert Opin. Drug Metab. Toxicol. 2017, 13, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Firrman, J.; Liu, L.; Yam, K. A review on flavonoid apigenin: Dietary intake, ADME, antimicrobial effects, and interactions with human gut microbiota. Biomed. Res. Int. 2019, 2019, 7010467. [Google Scholar] [CrossRef] [PubMed]
- Wdowiak, K.; Walkowiak, J.; Pietrzak, R.; Bazan-Woźniak, A.; Cielecka-Piontek, J. Bioavailability of hesperidin and its aglycone hesperetin-compounds found in citrus fruits as a parameter conditioning the pro-health potential (neuroprotective and antidiabetic activity)-mini-review. Nutrients 2022, 14, 2647. [Google Scholar] [CrossRef]
- Gradolatto, A.; Canivenc-Lavier, M.C.; Basly, J.P.; Siess, M.H.; Teyssier, C. Metabolism of apigenin by rat liver phase I and phase II enzymes and by isolated perfused rat liver. Drug Metab. Dispos. 2004, 32, 58–65. [Google Scholar] [CrossRef]
- Crespy, V.; Morand, C.; Besson, C.; Cotelle, N.; Vézin, H.; Demigné, C.; Rémésy, C. The splanchnic metabolism of flavonoids highly differed according to the nature of the compound. Am. J. Physiol. Gastrointest Liver Physiol. 2003, 284, G980–G988. [Google Scholar] [CrossRef]
- Koster, H.; Halsema, I.; Scholtens, E.; Knippers, M.; Mulder, G.J. Dose-dependent shifts in the sulfation and glucuronidation of phenolic compounds in the rat in vivo and in isolated hepatocytes. The role of saturation of phenol sulfotransferase. Biochem. Pharmacol. 1981, 30, 2569–2575. [Google Scholar] [CrossRef]
- Amić, D.; Davidović-Amić, D.; Beslo, D.; Rastija, V.; Lucić, B.; Trinajstić, N. SAR and QSAR of the antioxidant activity of flavonoids. Curr. Med. Chem. 2007, 14, 827–845. [Google Scholar] [CrossRef] [PubMed]
- Atmani, D.; Chaher, N.; Atmani, D.; Berboucha, M.; Debbache, N.; Boudaoud, H. Flavonoids in human health: From structure to biological activity. Curr. Nutr. Food Sci. 2009, 5, 225–237. [Google Scholar] [CrossRef]
- Morand, C.; Crespy, V.; Manach, C.; Besson, C.; Demigné, C.; Rémésy, C. Plasma metabolites of quercetin and their antioxidant properties. Am. J. Physiol. 1998, 275, R212–R219. [Google Scholar] [CrossRef] [PubMed]
- Terao, J.; Murota, K.; Kawai, Y. Conjugated quercetin glucuronides as bioactive metabolites and precursors of aglycone in vivo. Food Funct. 2011, 2, 11–17. [Google Scholar] [CrossRef]
- Luca, S.V.; Macovei, I.; Bujor, A.; Miron, A.; Skalicka-Woźniak, K.; Aprotosoaie, A.C.; Trifan, A. Bioactivity of dietary polyphenols: The role of metabolites. Crit. Rev. Food Sci. Nutr. 2020, 60, 626–659. [Google Scholar] [CrossRef]
- Williamson, G.; Barron, D.; Shimoi, K.; Terao, J. In vitro biological properties of flavonoid conjugates found in vivo. Free Radic. Res. 2005, 39, 457–469. [Google Scholar] [CrossRef]
- Day, A.J.; Bao, Y.; Morgan, M.R.; Williamson, G. Conjugation position of quercetin glucuronides and effect on biological activity. Free Radic. Biol. Med. 2000, 29, 1234–1243. [Google Scholar] [CrossRef]
- Nishikawa, M.; Kada, Y.; Kimata, M.; Sakaki, T.; Ikushiro, S. Comparison of metabolism and biological properties among positional isomers of quercetin glucuronide in LPS- and RANKL-challenged RAW264.7 cells. Biosci. Biotechnol. Biochem. 2022, 86, 1670–1679. [Google Scholar] [CrossRef]
- Yu, P.R.; Hsu, J.Y.; Tseng, C.Y.; Chen, J.H.; Lin, H.H. The inhibitory effect of quercetin-3-glucuronide on pulmonary injury in vitro and in vivo. J. Food Drug Anal. 2023, 31, 254–277. [Google Scholar] [CrossRef]
- Żyżyńska-Granica, B.; Gierlikowska, B.; Parzonko, A.; Kiss, A.K.; Granica, S. The bioactivity of flavonoid glucuronides and free aglycones in the context of their absorption, II phase metabolism and deconjugation at the inflammation site. Food Chem. Toxicol. 2020, 135, 110929. [Google Scholar] [CrossRef]
- Hong, Y.; Mitchell, A. Metabolic profiling of flavonol metabolites in human urine by liquid chromatography and tandem mass spectrometry. J. Agric. Food Chem. 2004, 52, 6794–6801. [Google Scholar] [CrossRef] [PubMed]
- Di Lorenzo, C.; Colombo, F.; Biella, S.; Stockley, C.; Restani, P. Polyphenols and human health: The role of bioavailability. Nutrients 2021, 13, 273. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Parameter | |||
|---|---|---|---|
| Fruit Ingestion | Flavonoid | Cmax (μg/mL) | AUC0–8 (μg·h/mL) |
| Guava | Luteolin | 0.8 ± 0.1 | 2.3 ± 0.9 |
| Quercetin | 1.1 ± 0.2 | 5.4 ± 1.3 | |
| Myricetin | 0.14 ± 0.03 | 0.6 ± 0.5 | |
| Pineapple | Kaempferol | 1.7 ± 0.4 | 9.9 ± 2.3 |
| Quercetin | 1.4 ± 0.4 | 6.5 ± 1.5 | |
| Myricetin | 0.2 ± 0.1 | 0.8 ± 0.3 | |
| Pomelo | Luteolin | 1.2 ± 0.3 | 6.4 ± 1.1 |
| Kaempferol | 0.8 ± 0.3 | 3.2 ± 1.0 | |
| Amount (μg) | ||||
|---|---|---|---|---|
| Fruit Ingestion | Flavonoid | Glucuronide | Sulfate | Total |
| Guava | Luteolin | - | 109.5 ± 22.0 | 109.5 ± 22.0 |
| Kaempferol | 5.2 ± 1.7 | 7.4 ± 2.0 | 12.6 ± 3.5 | |
| Quercetin | 363.8 ± 68.6 | 546.2 ± 78.4 | 910.0 ± 137.3 | |
| Myricetin | - | 269.8 ± 33.4 | 269.8 ± 33.4 | |
| Pineapple | Kaempferol | 71.2 ± 40.9 | 419.1 ± 135.4 | 490.3 ± 161.3 |
| Quercetin | 311.6 ± 171.4 | 387.8 ± 122.8 | 699.4 ± 28.4 | |
| Myricetin | - | 149.6 ± 51.8 | 149.6 ± 51.8 | |
| Pomelo | Luteolin | - | 450.3 ± 55.9 | 450.3 ± 55.9 |
| Kaempferol | 23.8 ± 4.8 | 43.6 ± 5.2 | 67.0 ± 8.9 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chomphen, L.; Yamanont, P.; Morales, N.P. Flavonoid Metabolites in Serum and Urine after the Ingestion of Selected Tropical Fruits. Nutrients 2024, 16, 161. https://doi.org/10.3390/nu16010161
Chomphen L, Yamanont P, Morales NP. Flavonoid Metabolites in Serum and Urine after the Ingestion of Selected Tropical Fruits. Nutrients. 2024; 16(1):161. https://doi.org/10.3390/nu16010161
Chicago/Turabian StyleChomphen, Lalita, Paveena Yamanont, and Noppawan Phumala Morales. 2024. "Flavonoid Metabolites in Serum and Urine after the Ingestion of Selected Tropical Fruits" Nutrients 16, no. 1: 161. https://doi.org/10.3390/nu16010161
APA StyleChomphen, L., Yamanont, P., & Morales, N. P. (2024). Flavonoid Metabolites in Serum and Urine after the Ingestion of Selected Tropical Fruits. Nutrients, 16(1), 161. https://doi.org/10.3390/nu16010161