
Abalone Viscera Fermented with Aspergillus oryzae 001 Prevents Pressure Elevation by Inhibiting Angiotensin Converting Enzyme

Abstract
:1. Introduction
2. Materials and Methods
2.1. Material and Reagents
2.2. Fermentation by A. oryzae 001
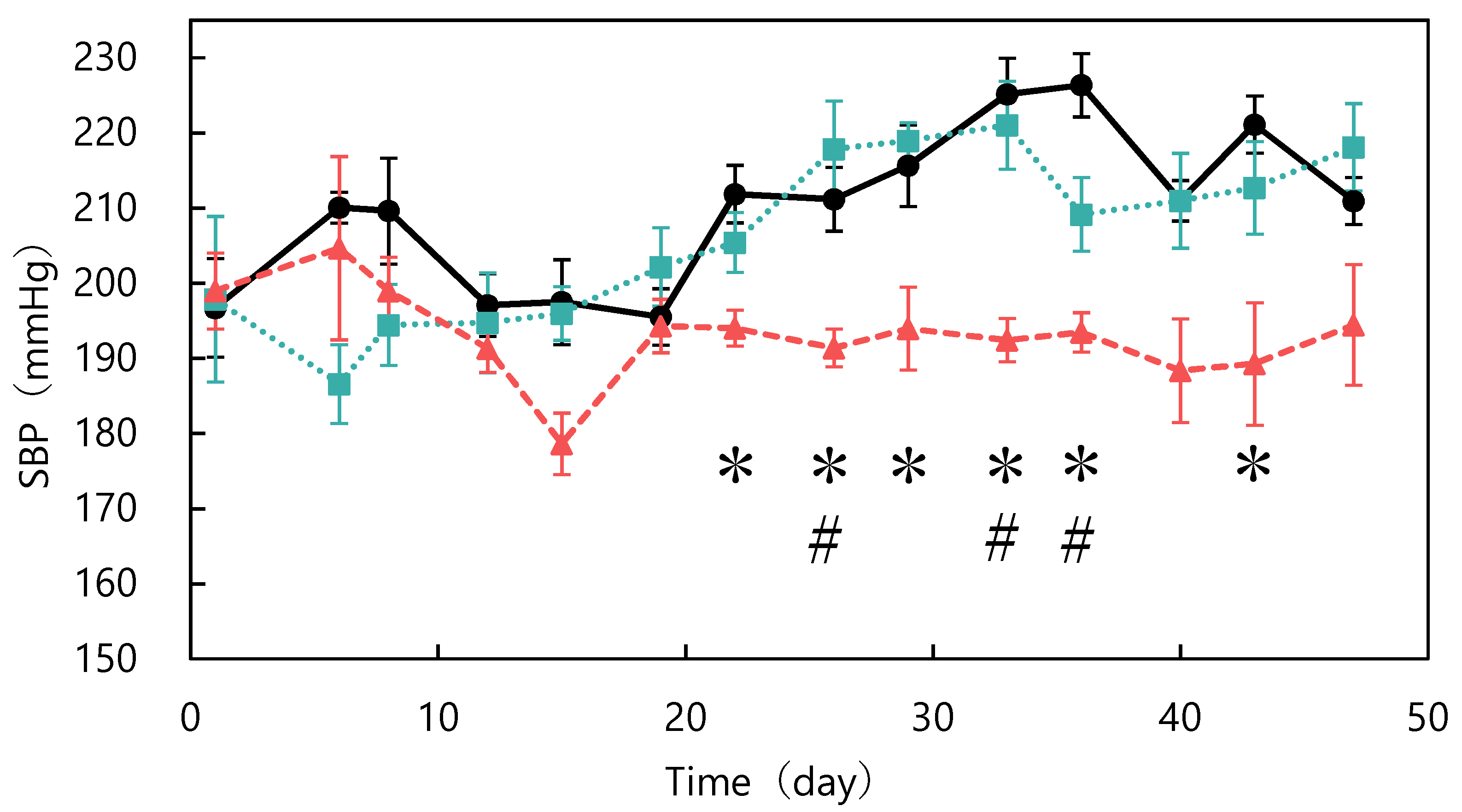
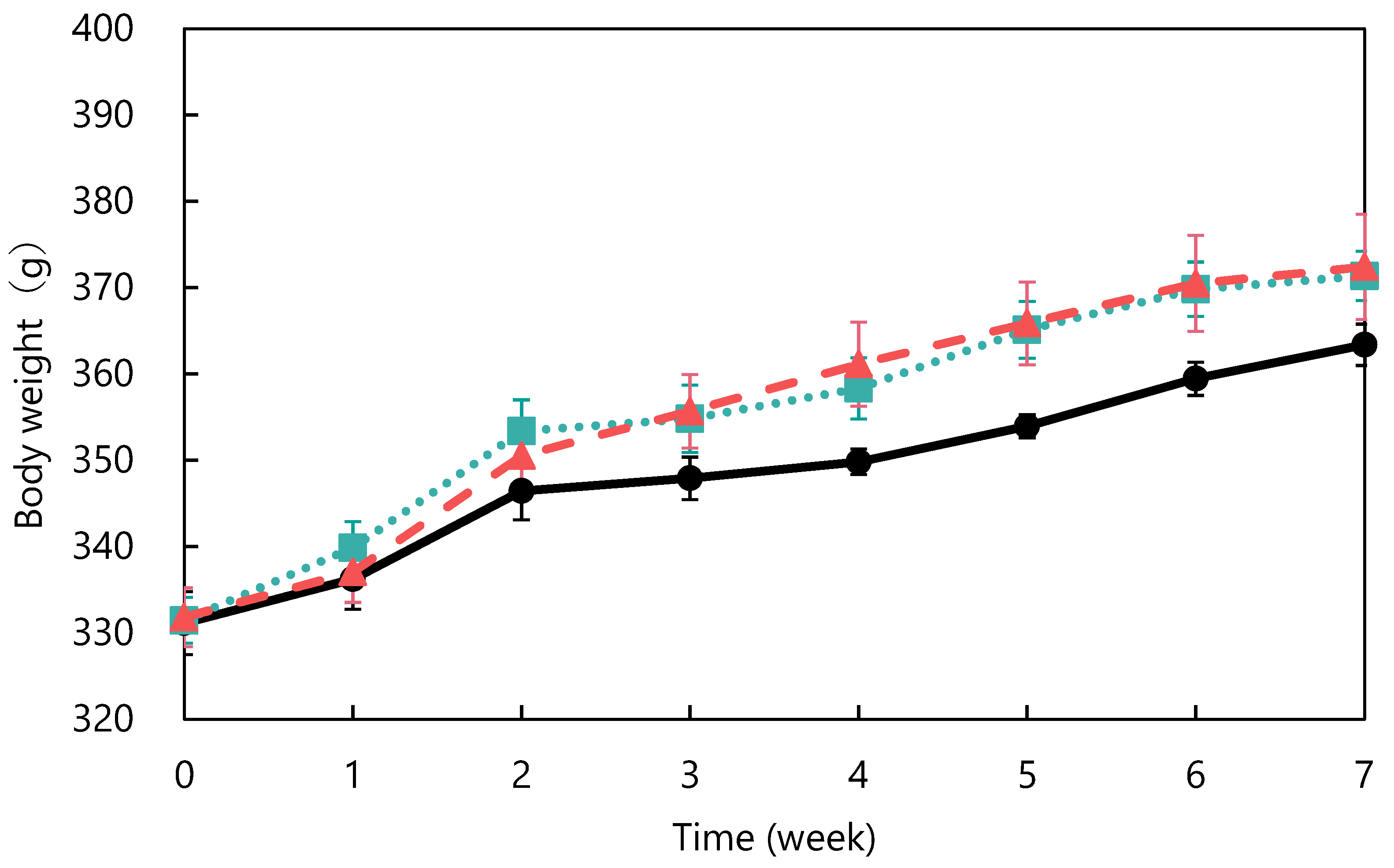
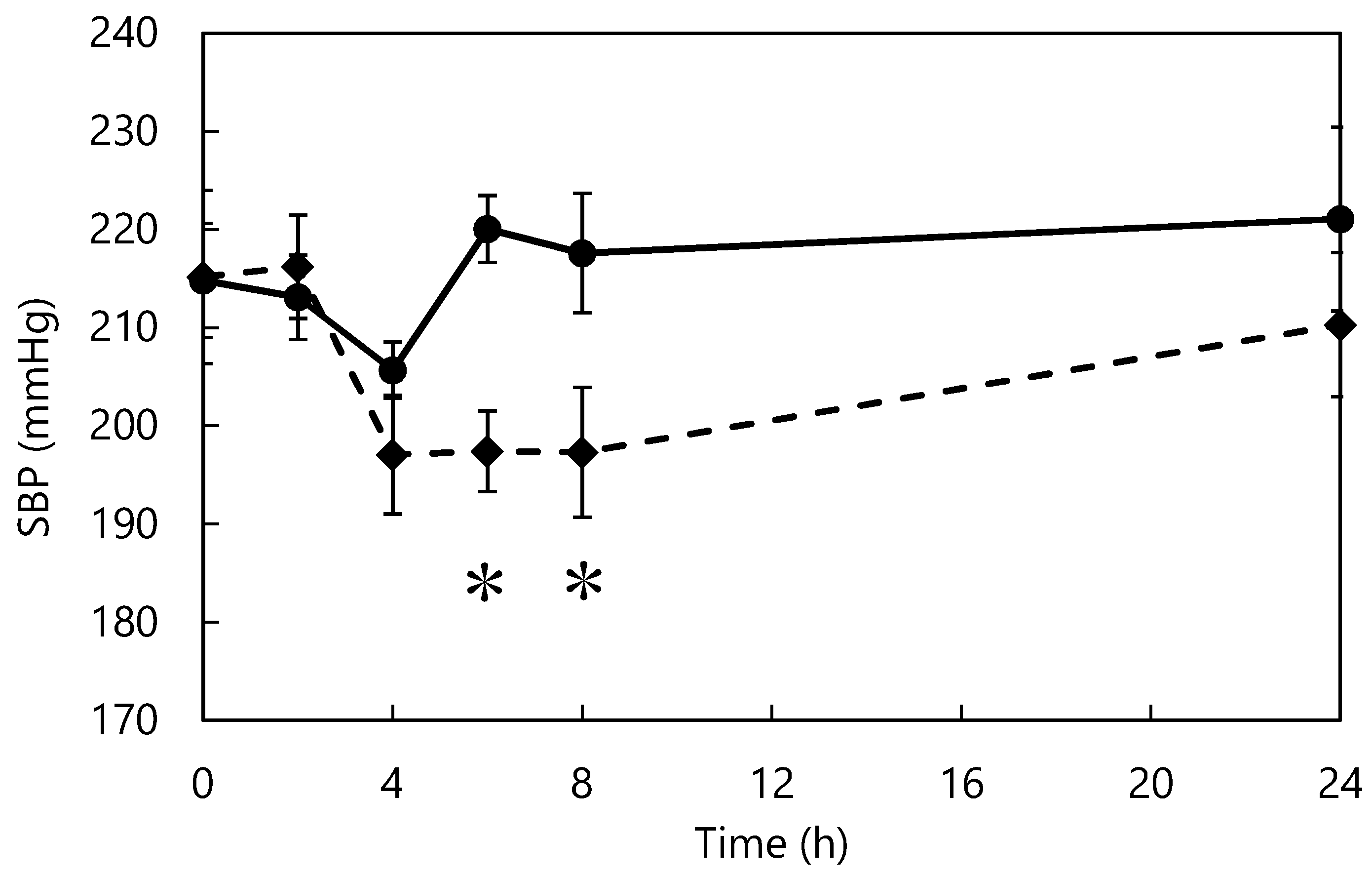
2.3. Long-Term Administration Study
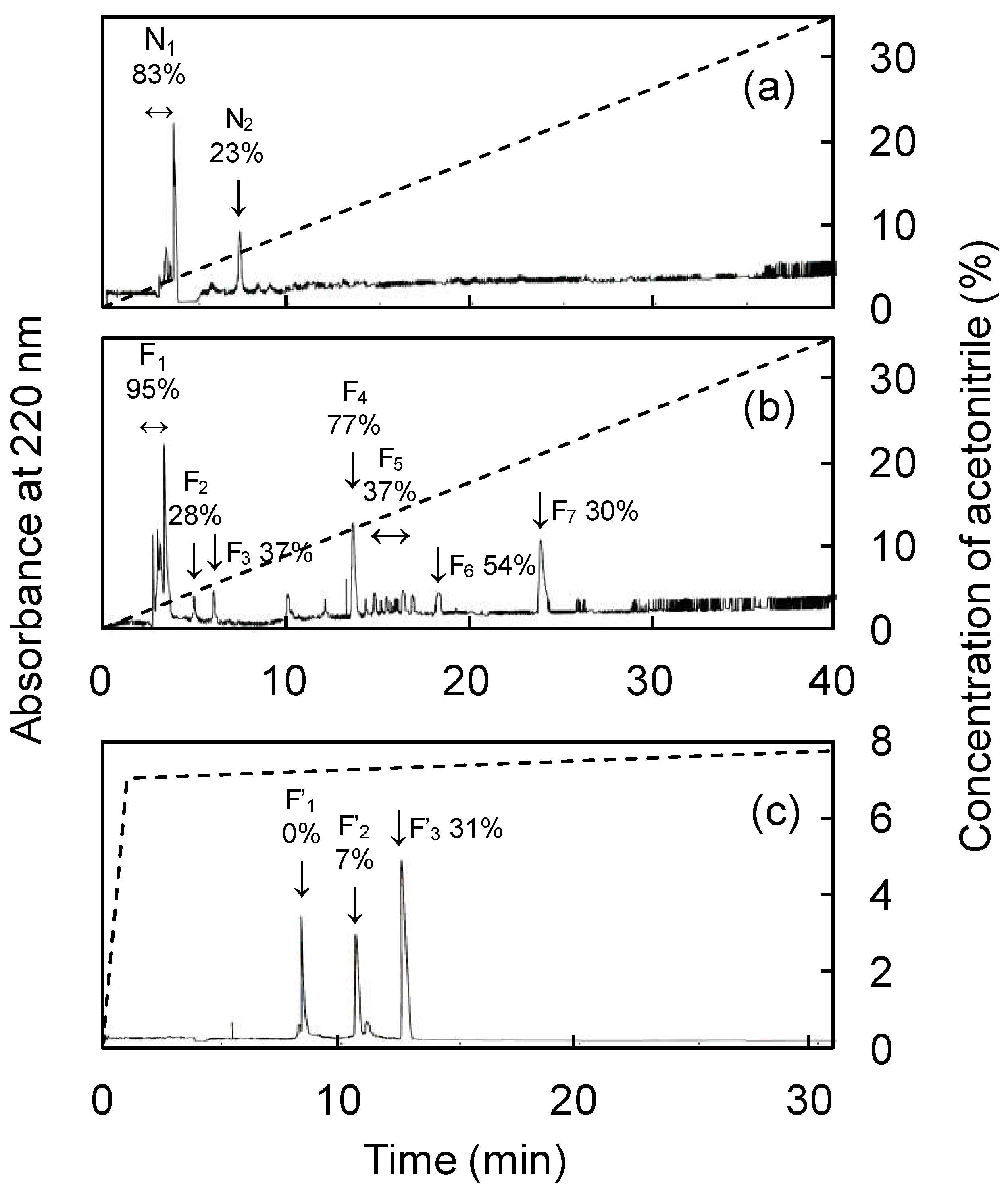
2.4. Purification of ACE Inhibitor Components
2.5. Identification of ACE Inhibitors
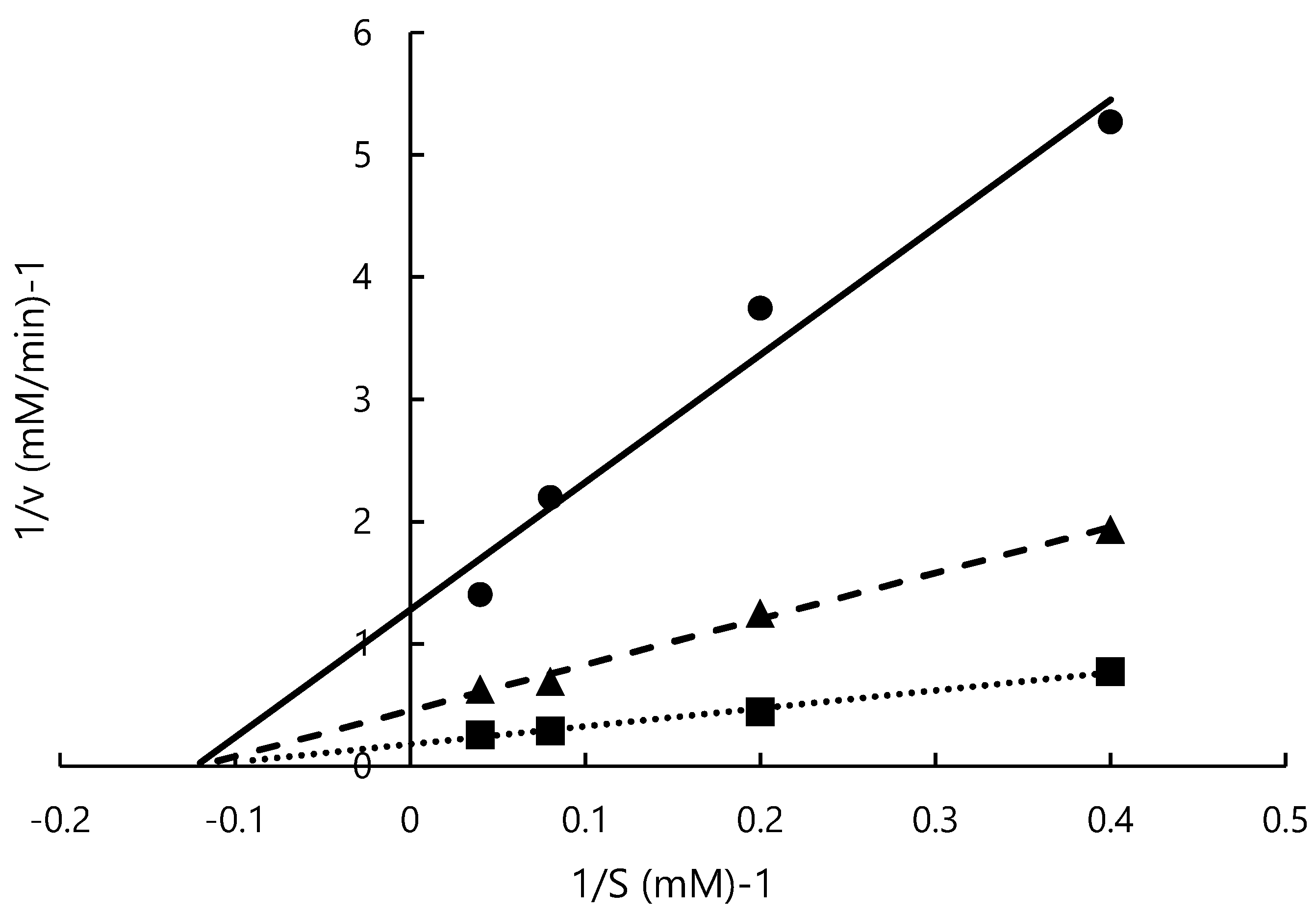
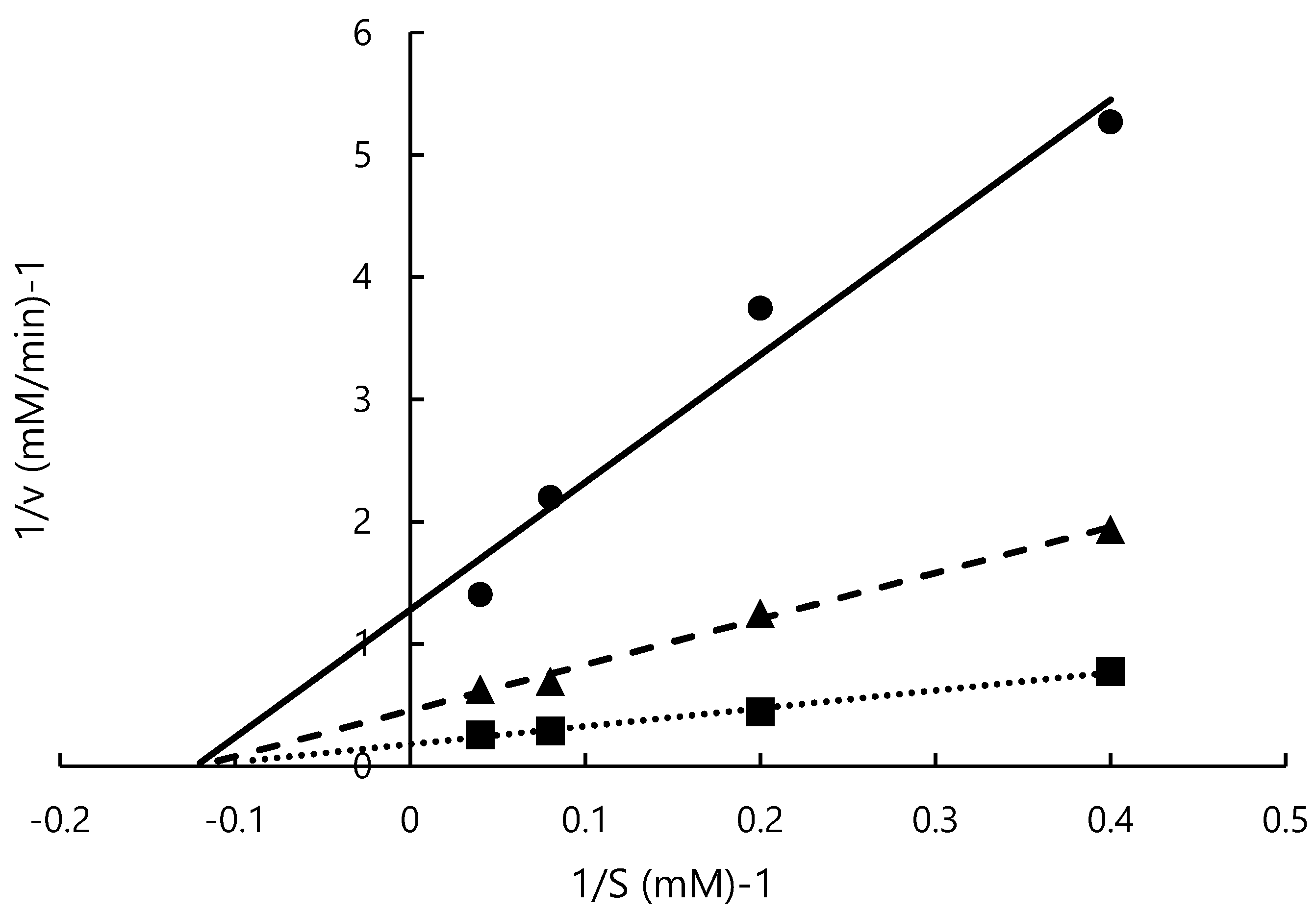
2.6. Estimation of Mode of Inhibition
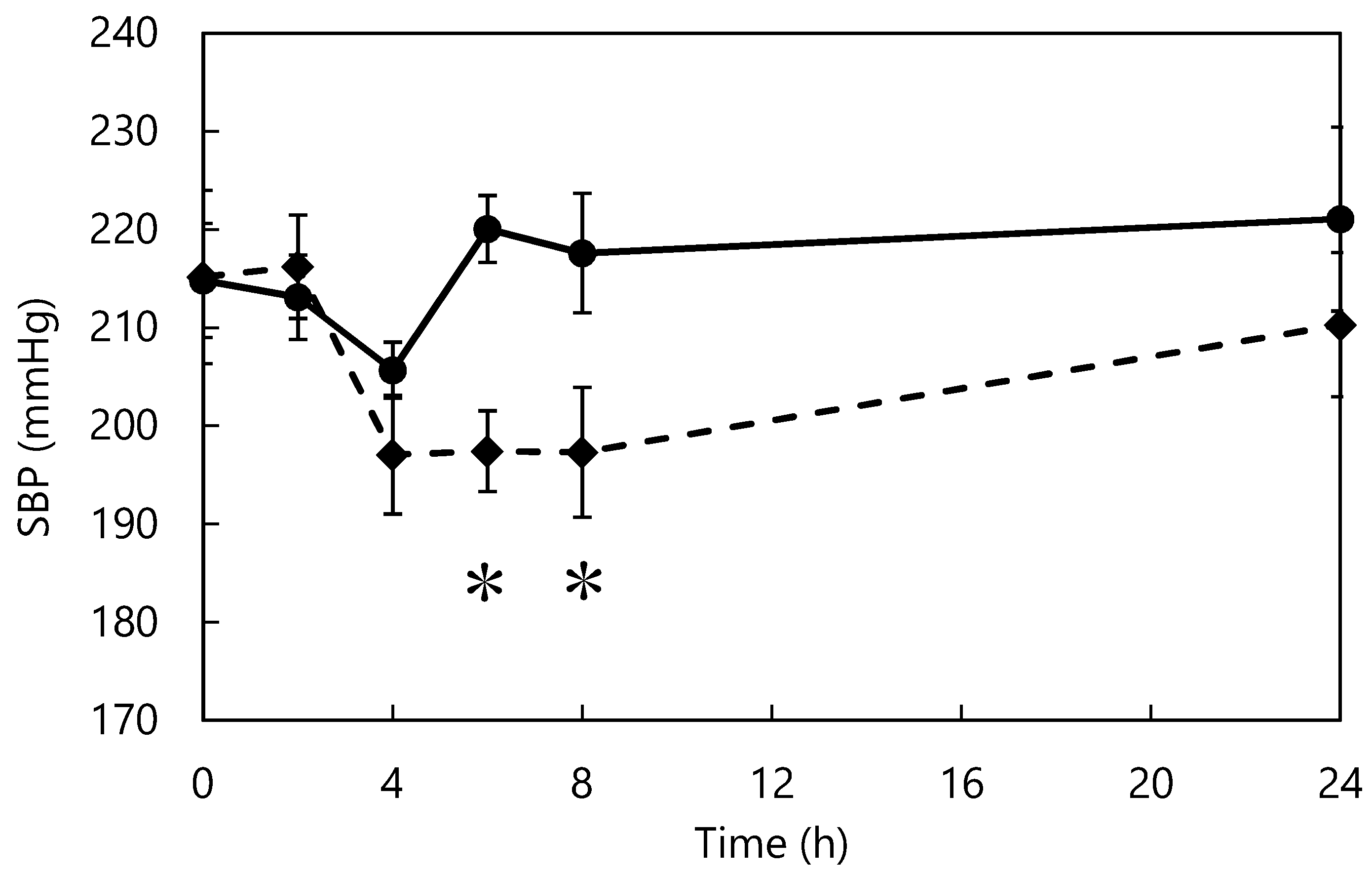
2.7. Single-Dose ACE Inhibitor Study
2.8. Statistical Analysis
3. Results & Discussion
3.1. Long-Term Dosing Study
3.2. Purification of ACE inhibitors
3.3. Identification of ACE Inhibitors
3.4. Estimation of Mode of Inhibition
3.5. Single-Dose Study of ACE Inhibitors in SHRs
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sadh, P.K.; Duhan, S.; Duhan, J.S. Agro-Industrial wastes and their utilization using solid state fermentation: A Review. Bioresour. Bioprocess. 2018, 5, 1. [Google Scholar] [CrossRef]
- Roodt-Wilding, R. Abalone ranching: A review on genetic considerations. Aquacult. Res. 2007, 38, 1229–1241. [Google Scholar] [CrossRef]
- FAO. FAO Yearbook of Fishery and Aquaculture Statistics; FAO: Rome, Italy, 2021; pp. 118–119. [Google Scholar]
- Je, J.Y.; Park, S.Y.; Hwang, J.Y.; Ahn, C.B. Amino acid composition and in vitro antioxidant and cytoprotective activity of abalone viscera hydrolysate. J. Funct. Foods 2015, 16, 94–103. [Google Scholar] [CrossRef]
- Sun, L.; Zhu, B.; Li, D.; Wang, L.; Dong, X.; Murata, Y.; Xing, R.; Dong, Y. Purification and bioactivity of a sulphated polysaccharide conjugate from viscera of abalone Haliotis discus Hannai Ino. Food Agric. Immunol. 2010, 21, 15–26. [Google Scholar] [CrossRef]
- Zhu, B.W.; Wang, L.S.; Zhou, D.Y.; Li, D.M.; Sun, L.M.; Yang, J.F.; Wu, H.T.; Zhou, X.Q.; Tada, M. Antioxidant activity of sulphated polysaccharide conjugates from abalone (Haliotis discus Hannai Ino). Eur. Food Res. Technol. 2008, 227, 1663–1668. [Google Scholar] [CrossRef]
- Hu, Y.; Yang, J.; He, C.; Wei, H.; Wu, G.; Xiong, H.; Ma, Y. Fractionation and purification of antioxidant peptides from abalone viscera by a combination of Sephadex G-15 and Toyopearl HW-40F chromatography. Int. J. Food Sci. Technol. 2022, 57, 1218–1225. [Google Scholar] [CrossRef]
- Viana, M.T.; López, L.M.; García-Esquivel, Z.; Mendez, E. The use of silage made from fish and abalone viscera as an ingredient in abalone feed. Aquaculture 1996, 140, 87–98. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Møller, L.L.H.; Larsen, T.O.; Kumar, R.; Arnau, J. Safety of the fungal workhorses of industrial biotechnology: Update on the mycotoxin and secondary metabolite potential of Aspergillus niger, Aspergillus oryzae, and Trichoderma Reesei. Appl. Microbiol. Biotechnol. 2018, 102, 9481–9515. [Google Scholar] [CrossRef]
- He, B.; Hu, Z.; Ma, L.; Li, H.; Ai, M.; Han, J.; Zeng, B. Transcriptome analysis of different growth stages of Aspergillus oryzae reveals dynamic changes of distinct classes of genes during growth. BMC Microbiol. 2018, 18, 12. [Google Scholar] [CrossRef]
- Papagianni, M. Fungal morphology and metabolite production in submerged mycelial processes. Biotechnol. Adv. 2004, 22, 189–259. [Google Scholar] [CrossRef]
- Zareian, M.; Oskoueian, E.; Majdinasab, M.; Forghani, B. Production Og GABA-enriched idli with ACE inhibitory and antioxidant properties using Aspergillus oryzae: The antihypertensive effects in spontaneously hypertensive rats. Food Funct. 2020, 11, 4304–4313. [Google Scholar] [CrossRef]
- Xu, J.; Jin, F.; Hao, J.; Regenstein, J.M.; Wang, F. Preparation of soy sauce by walnut meal fermentation: Composition, antioxidant properties, and angiotensin-converting enzyme inhibitory activities. Food Sci. Nutr. 2020, 8, 1665–1676. [Google Scholar] [CrossRef] [PubMed]
- Kjeldsen, S.E. Hypertension and cardiovascular risk: General aspects. Pharmacol. Res. 2018, 129, 95–99. [Google Scholar] [CrossRef] [PubMed]
- WHO. Guideline for the Pharmacological Treatment of Hypertension in Adults; WHO: Geneva, Switzerland, 2021; p. 1.
- Pihlanto-Leppaè, A. Bioactive peptides derived from bovine whey proteins: Opioid and ace-inhibitory peptides. Food Sci. Technol. 2001, 11, 347–356. [Google Scholar] [CrossRef]
- Chakraborty, R.; Roy, S. Angiotensin-converting enzyme inhibitors from plants: A review of their diversity, modes of action, prospects, and concerns in the management of diabetes-centric complications. J. Integr. Med. 2021, 19, 478–492. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Yu, J.; Xu, W.; Shuang, Q. Purification and characterization of angiotensin-I-converting enzyme inhibitory peptides isolated from whey proteins of milk fermented with Lactobacillus Plantarum QS670. J. Dairy Sci. 2020, 103, 4919–4928. [Google Scholar] [CrossRef]
- Chen, J.; Yu, X.; Chen, Q.; Wu, Q.; He, Q. Screening and mechanisms of novel angiotensin-I-converting enzyme inhibitory peptides from rabbit meat proteins: A combined in silico and in vitro study. Food Chem. 2022, 370, 131070. [Google Scholar] [CrossRef]
- Rendón-Rosales, M.Á.; Torres-Llanez, M.J.; Mazorra-Manzano, M.A.; González-Córdova, A.F.; Hernández-Mendoza, A.; Vallejo-Cordoba, B. In vitro and in silico evaluation of multifunctional properties of bioactive synthetic peptides identified in milk fermented with Lactococcus lactis NRRL B-50571 and NRRL B-50572. LWT 2022, 154, 112581. [Google Scholar] [CrossRef]
- Oh, J.Y.; Je, J.G.; Lee, H.G.; Kim, E.A.; Kang, S.I.; Lee, J.S.; Jeon, Y.J. Anti-hypertensive activity of novel peptides identified from olive flounder (Paralichthys olivaceus) surimi. Foods 2020, 9, 647. [Google Scholar] [CrossRef]
- Fujimura, Y.; Shimura, M.; Nagai, H.; Hamada-Sato, N. Evaluation of angiotensin-converting enzyme-inhibitory activity in abalone viscera fermented by Lactobacillus casei 001. J. Funct. Foods 2021, 82, 104474. [Google Scholar] [CrossRef]
- Yamanushi, M.; Shimura, M.; Nagai, H.; Hamada-Sato, N. Antihypertensive effects of abalone viscera fermented with Lactiplantibacillus pentosus SN001 via angiotensin-converting enzyme inhibition. Food Chem. X 2022, 13, 100239. [Google Scholar] [CrossRef] [PubMed]
- Liao, P.; Lan, X.; Liao, D.; Sun, L.; Zhou, L.; Sun, J.; Tong, Z. Isolation and characterization of angiotensin I-converting enzyme (ACE) inhibitory peptides from the enzymatic hydrolysate of Carapax Trionycis (the shell of the turtle Pelodiscus sinensis). J Agric. Food Chem. 2018, 66, 7015–7022. [Google Scholar] [CrossRef] [PubMed]
- Jama, H.A.; Muralitharan, R.R.; Xu, C.; O’Donnell, J.A.; Bertagnolli, M.; Broughton, R.S.B.; Head, G.A.; Marques, F.Z. Rodent models of hypertension. Br. J. Pharmacol. 2022, 179, 918–937. [Google Scholar] [CrossRef]
- He, H.L.; Liu, D.; Ma, C.B. Review on the angiotensin-I-converting enzyme (ACE) inhibitor peptides from marine proteins. Appl. Biochem. Biotechnol. 2013, 169, 738–749. [Google Scholar] [CrossRef] [PubMed]
- Pinto, Y.M.; Paul, M.; Ganten, D. Lessons from rat models of hypertension: From Goldblatt to genetic engineering. Cardiovasc. Res. 1998, 39, 77–88. [Google Scholar] [CrossRef]
- Manoharan, S.; Shuib, A.S.; Abdullah, N.; Ashrafzadeh, A.; Kabir, N. Gly-Val-Arg, an angiotensin-I-converting enzyme inhibitory tripeptide ameliorates hypertension on spontaneously hypertensive rats. Process Biochem. 2018, 69, 224–232. [Google Scholar] [CrossRef]
- Lye, H.S.; Kuan, C.Y.; Ewe, J.A.; Fung, W.Y.; Liong, M.T. The improvement of hypertension by probiotics: Effects on cholesterol, diabetes, renin, and phytoestrogens. Int. J. Mol. Sci. 2009, 27, 3755–3775. [Google Scholar] [CrossRef]
- Clarke, R.; Frost, C.; Collins, R.; Appleby, P.; Peto, R. Dietary lipids and blood cholesterol: Quantitative meta-analysis of metabolic ward studies. BMJ 1997, 314, 112–117. [Google Scholar] [CrossRef]
- Karimi, S.; Soofiani, N.M.; Lundh, T.; Mahboubi, A.; Kiessling, A.; Taherzadeh, M.J. Evaluation of filamentous fungal biomass cultivated on vinasse as an alternative nutrient source of fish feed: Protein, lipid, and mineral composition. Fermentation 2019, 5, 99. [Google Scholar] [CrossRef]
- Golovinskaia, O.; Wang, C.K. The hypoglycemic potential of phenolics from functional foods and their mechanisms. Food Sci. Hum. Wellness 2023, 12, 986–1007. [Google Scholar] [CrossRef]
- Glazunova, O.A.; Moiseenko, K.v.; Savinova, O.S.; Fedorova, T.V. In vitro and in vivo antihypertensive effect of milk fermented with different strains of common starter Lactic Acid Bacteria. Nutrients 2022, 14, 5357. [Google Scholar] [CrossRef] [PubMed]
- Chai, K.F.; Voo, A.Y.H.; Chen, W.N. Bioactive peptides from food fermentation: A comprehensive review of their sources, bioactivities, applications, and future development. Compr. Rev. Food Sci. Food Saf. 2020, 19, 3825–3885. [Google Scholar] [CrossRef]
- Soleymanzadeh, N.; Mirdamadi, S.; Mirzaei, M.; Kianirad, M. Novel β-casein derived antioxidant and ACE-Inhibitory active peptide from camel milk fermented by Leuconostoc Lactis PTCC1899: Identification and molecular docking. Int. Dairy J. 2019, 97, 201–208. [Google Scholar] [CrossRef]
- Moslehishad, M.; Ehsani, M.R.; Salami, M.; Mirdamadi, S.; Ezzatpanah, H.; Naslaji, A.N.; Moosavi-Movahedi, A.A. The comparative assessment of ACE-Inhibitory and antioxidant activities of peptide fractions obtained from fermented camel and bovine milk by Lactobacillus Rhamnosus PTCC 1637. Int. Dairy J. 2013, 29, 82–87. [Google Scholar] [CrossRef]
- Sitanggang, A.B.; Sumitra, J.; Budijanto, S. Continuous production of tempe-based bioactive peptides using an automated enzymatic membrane reactor. Innov. Food Sci. Emerg. Technol. 2021, 68, 102639. [Google Scholar] [CrossRef]
- Li, Z.; Wang, B.; Chi, C.; Gong, Y.; Luo, H.; Ding, G. Influence of average molecular weight on antioxidant and functional properties of cartilage collagen hydrolysates from Sphyrna lewini, Dasyatis akjei and Raja porosa. Food Res. Int. 2013, 51, 283–293. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, T.K.; Yong, H.I.; Cha, J.Y.; Song, K.M.; Lee, H.G.; Je, J.G.; Kang, M.C.; Choi, Y.S. Peptides inhibiting angiotensin-I-converting enzyme: Isolation from flavourzyme hydrolysate of protaetia brevitarsis larva protein and identification. Food Chem. 2023, 399, 133897. [Google Scholar] [CrossRef]
- O’Brien, C.; Mahoney, C.; Tharion, W.J.; Sils, I.v.; Castellani, J.W. Dietary tyrosine benefits cognitive and psychomotor performance during body cooling. Physiol. Behav. 2007, 90, 301–307. [Google Scholar] [CrossRef]
- Zhang, F.; Wang, Z.; Xu, S. Macroporous resin purification of grass carp fish (Ctenopharyngodon idella) scale peptides with in vitro angiotensin-I converting enzyme (ACE) inhibitory ability. Food Chem. 2009, 117, 387–392. [Google Scholar] [CrossRef]
- Ahn, C.B.; Jeon, Y.J.; Kim, Y.T.; Je, J.Y. Angiotensin I converting enzyme (ACE) inhibitory peptides from salmon byproduct protein hydrolysate by alcalase hydrolysis. Process Biochem. 2012, 47, 2240–2245. [Google Scholar] [CrossRef]
- Sun, S.; Xu, X.; Sun, X.; Zhang, X.; Chen, X.; Xu, N. Preparation and identification of ACE inhibitory peptides from the marine macroalga Ulva intestinalis. Mar. Drugs 2019, 17, 179. [Google Scholar] [CrossRef] [PubMed]
- Song, C.C.; Qiao, B.W.; Zhang, Q.; Wang, C.X.; Fu, Y.H.; Zhu, B.W. Study on the domain selective inhibition of angiotensin-converting enzyme (ACE) by food-derived tyrosine-containing dipeptides. J. Food Biochem. 2021, 45, e13779. [Google Scholar] [CrossRef] [PubMed]
- Rudolph, S.; Lunow, D.; Kaiser, S.; Henle, T. Identification and quantification of ACE-Inhibiting peptides in enzymatic hydrolysates of plant proteins. Food Chem. 2017, 224, 19–25. [Google Scholar] [CrossRef]
- Li, Y.; Sadiq, F.A.; Liu, T.J.; Chen, J.C.; He, G.Q. Purification and identification of novel peptides with inhibitory effect against angiotensin I-converting enzyme and optimization of process conditions in milk fermented with the yeast Kluyveromyces marxianus. J. Funct. Foods 2015, 16, 278–288. [Google Scholar] [CrossRef]
- Gu, X.; Hou, Y.K.; Li, D.; Wang, J.Z.; Wang, F.J. Separation, purification, and identification of angiotensin I-converting enzyme inhibitory peptides from walnut (Juglans Regia L.) hydrolyzate. Int. J. Food Prop. 2015, 18, 266–276. [Google Scholar] [CrossRef]
- Suetsuna, K. Rapid communication isolation and characterization of angiotensin I-converting enzyme inhibitor dipeptides derived from allium sativum L (Garlic). J. Nutr. Biochem. 1997, 9, 415–419. [Google Scholar] [CrossRef]
- Girgih, A.T.; He, R.; Aluko, R.E. Kinetics and molecular docking studies of the inhibitions of angiotensin converting enzyme and renin activities by hemp seed (Cannabis Sativa L.) peptides. J. Agric. Food Chem. 2014, 62, 4135–4144. [Google Scholar] [CrossRef]
- Chiou, T.K.; Lai, M.M.; Shiau, C.Y. Seasonal variations of chemical constituents in the muscle and viscera of small abalone fed different diets. Fish. Sci. 2001, 67, 146–156. [Google Scholar] [CrossRef]
- Zhang, Q.; Song, C.; Zhao, J.; Shi, X.; Sun, M.; Liu, J.; Fu, Y.; Jin, W.; Zhu, B. Separation and Characterization of Antioxidative and Angiotensin Converting Enzyme Inhibitory Peptide from Jellyfish Gonad Hydrolysate. Molecules 2018, 23, 94. [Google Scholar] [CrossRef]
- Lee, S.H.; Qian, Z.J.; Kim, S.K. A novel angiotensin I converting enzyme inhibitory peptide from tuna frame protein hydrolysate and its antihypertensive effect in spontaneously hypertensive rats. Food Chem 2010, 118, 96–102. [Google Scholar] [CrossRef]
- Balti, R.; Bougatef, A.; Sila, A.; Guillochon, D.; Dhulster, P.; Nedjar-Arroume, N. Nine novel angiotensin I-converting enzyme (ACE) inhibitory peptides from cuttlefish (Sepia Officinalis) muscle protein hydrolysates and antihypertensive effect of the potent active peptide in spontaneously hypertensive rats. Food Chem. 2015, 170, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Li, G.H.; Le, G.W.; Shi, Y.H.; Shrestha, S. Angiotensin I-converting enzyme inhibitory peptides derived from food proteins and their physiological and pharmacological effects. Nutr. Res. 2004, 24, 469–486. [Google Scholar] [CrossRef]
- Bossy, J.; Guidoux, R.; Milon, H. Development of hypertension in spontaneously hypertensive rats fed L-tyrosine-supplemented diets. Z Ernahrungswiss 1983, 22, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Hosokawa, T.; Yamaguchi, T.; Nakano, T.; Muramoto, K.; Kahara, T.; Funayama, K.; Kobayashi, A.; Nakano, T. Angiotensin I-converting enzyme inhibitory peptides derived from Wakame (Undaria pinnatifida) and their antihypertensive effect in spontaneously hypertensive rats. J. Agric. Food Chem. 2002, 50, 6245–6252. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Wu, K.; Wu, J. Pea-derived tripeptide LRW fails to reduce blood pressure in spontaneously hypertensive rats due to its low gastrointestinal stability and transepithelial permeability. Food Biosci. 2022, 49, 101964. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Ingredients (g/kg) | Control | Unfermented | Fermented |
|---|---|---|---|
| β-Cornstarch | 392 | 345.5 | 345.5 |
| Milk casein | 195 | 195 | 195 |
| α-Cornstarch | 132 | 132 | 132 |
| Sucrose | 100 | 100 | 100 |
| Soybean oil | 70 | 70 | 70 |
| Cellulose | 50 | 50 | 50 |
| AIN-76 Mineral mixture | 35 | 35 | 35 |
| AIN-76A Vitamin mixture | 12.5 | 12.5 | 12.5 |
| L-Cysteine | 3 | 3 | 3 |
| TBHQ | 0.014 | 0.014 | 0.014 |
| NaCl | 10 | 6.5 | 6.5 |
| Abalone viscera | 0 | 50 | 50 |
| Test Group | Control | Unfermented | Fermented |
|---|---|---|---|
| Kidney (g) | 2.56 ± 0.13 | 2.55 ± 0.14 | 2.47 ± 0.08 |
| Liver (g) | 10.06 ± 0.47 | 10.34 ± 0.43 | 10.26 ± 0.38 |
| Test Group | Control | Unfermented | Fermented |
|---|---|---|---|
| Glucose (mg/dL) | 119.0 ± 14.4 | 114.7 ± 32.7 | 133.6 ± 23.1 |
| Triglycerides (mg/dL) | 69.1 ± 16.2 | 78.9 ± 13.8 | 74.0 ± 14.4 |
| Total cholesterol (mg/dL) | 76.0 ± 14.5 | 84.0 ± 11.3 | 74.4 ± 8.8 |
| HDL-cholesterol (mg/dL) | 43.6 ± 2.9 | 42.1 ± 7.6 | 43.6 ± 7.2 |
| Alanine aminotransferase activity (IU/L) | 25.3 ± 2.4 | 24.6 ± 1.2 | 24.3 ± 2.2 |
| Aspartate aminotransferase activity (IU/L) | 33.0 ± 7.4 | 34.4 ± 7.1 | 31.1 ± 5.9 |
| MWCO (kDa) | <3 | 3~10 | 10~30 | 30~100 | 100< | |
|---|---|---|---|---|---|---|
| Fermented | IC50 (mg/mL) | 0.36 | 0.41 | 0.58 | 0.42 | 0.38 |
| Fraction weight (mg) | 451 | 42 | 18 | 10 | 24 | |
| Unfermented | IC50 (mg/mL) | 0.68 | 0.49 | 0.85 | N.D. | 1.35 |
| Fraction weight (mg) | 230 | 14 | 35 | N.D. | 36 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iwamoto, N.; Sasaki, A.; Maizawa, T.; Hamada-Sato, N. Abalone Viscera Fermented with Aspergillus oryzae 001 Prevents Pressure Elevation by Inhibiting Angiotensin Converting Enzyme. Nutrients 2023, 15, 947. https://doi.org/10.3390/nu15040947
Iwamoto N, Sasaki A, Maizawa T, Hamada-Sato N. Abalone Viscera Fermented with Aspergillus oryzae 001 Prevents Pressure Elevation by Inhibiting Angiotensin Converting Enzyme. Nutrients. 2023; 15(4):947. https://doi.org/10.3390/nu15040947
Chicago/Turabian StyleIwamoto, Natsumi, Asahi Sasaki, Tomoaki Maizawa, and Naoko Hamada-Sato. 2023. "Abalone Viscera Fermented with Aspergillus oryzae 001 Prevents Pressure Elevation by Inhibiting Angiotensin Converting Enzyme" Nutrients 15, no. 4: 947. https://doi.org/10.3390/nu15040947
APA StyleIwamoto, N., Sasaki, A., Maizawa, T., & Hamada-Sato, N. (2023). Abalone Viscera Fermented with Aspergillus oryzae 001 Prevents Pressure Elevation by Inhibiting Angiotensin Converting Enzyme. Nutrients, 15(4), 947. https://doi.org/10.3390/nu15040947