
Low-Dose Dietary Fish Oil Improves RBC Deformability without Improving Post-Transfusion Recovery in Mice

 , , , , ,
, , , , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Standards
2.2. Animal Experiments
2.3. In Vivo RBC Lifespan
2.4. Blood Collection and Storage
2.5. 24-h PTR
2.6. Flow Cytometry
2.7. Analysis of FAMEs by Gas Chromatography-Mass Spectrometry (GC-MS)
2.8. Analysis of MDA by GC-MS
2.9. RBC Deformability
2.10. Complete Blood Count (CBC)
2.11. Analysis of Free Long Chain-PUFAs (LC-PUFAs), Oxylipins, Phospholipids, and Lysophospholipids by LC-MS/MS
2.12. UHPLC-MS Metabolomics
2.13. Statistics
3. Results
3.1. Pilot PTR Study
3.1.1. Dietary Lipids Are Incorporated into the RBC Membrane in C57BL/6J Mice
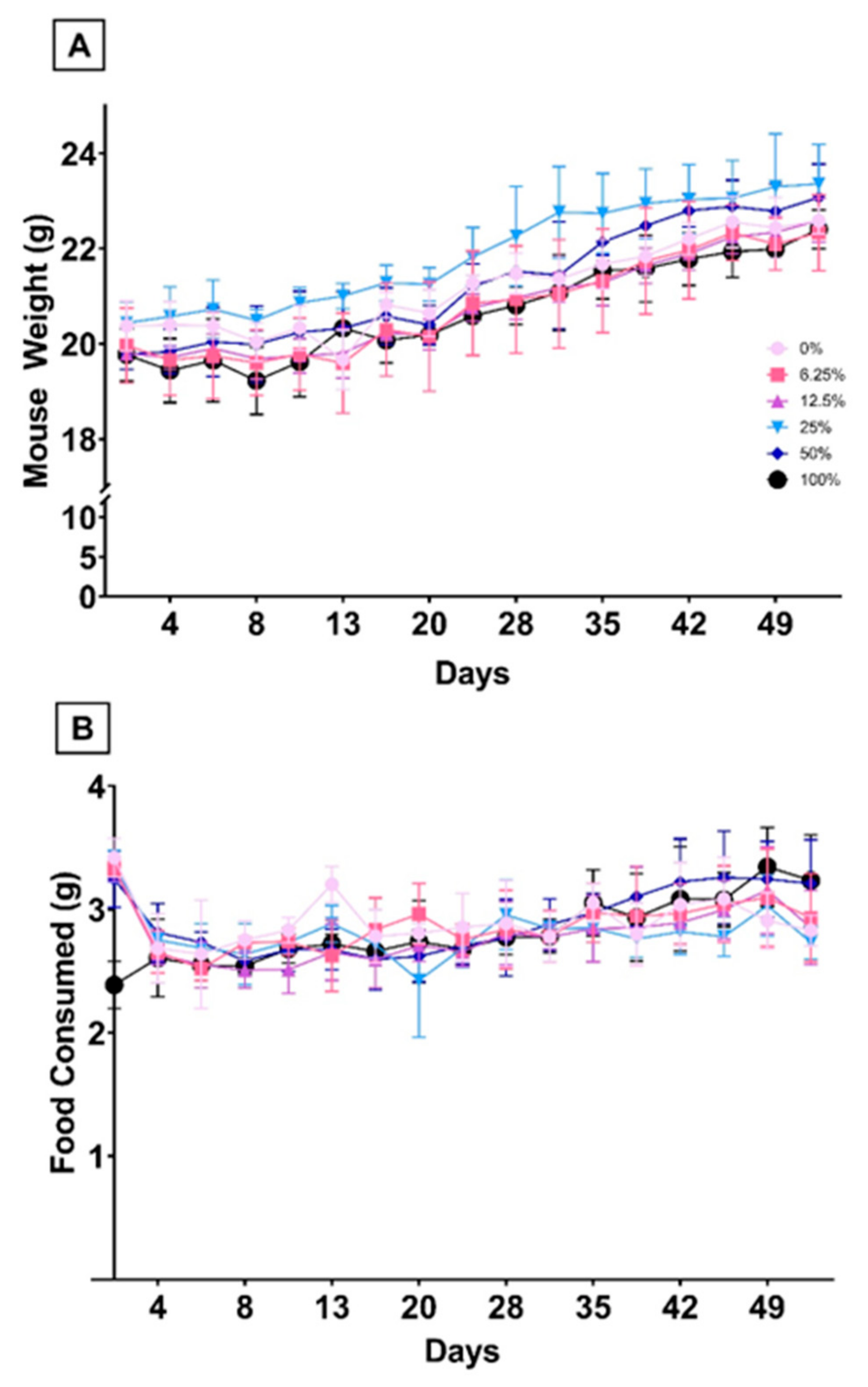
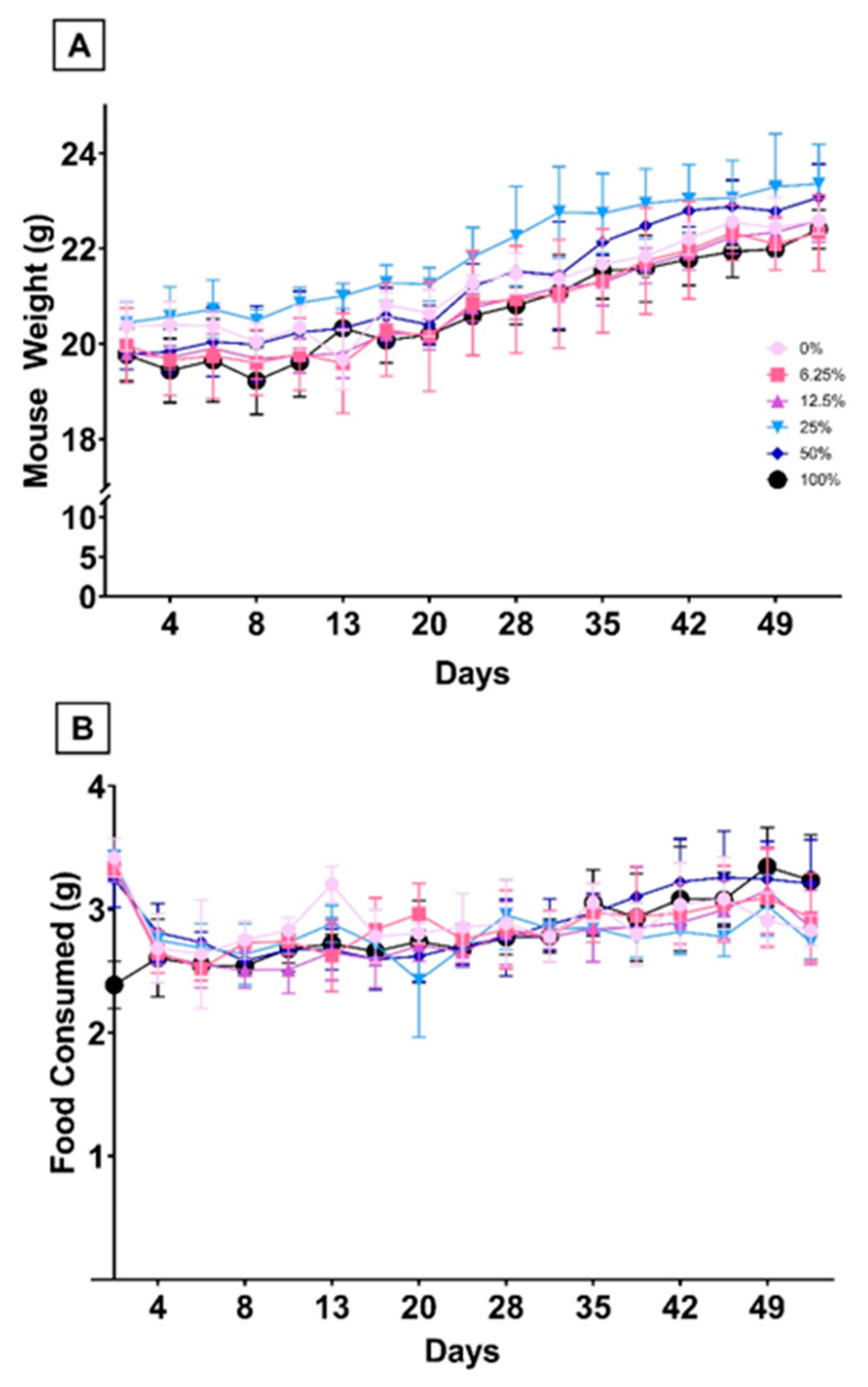
3.1.2. Dietary FO Was Not Associated with Differences in Hematological Parameters
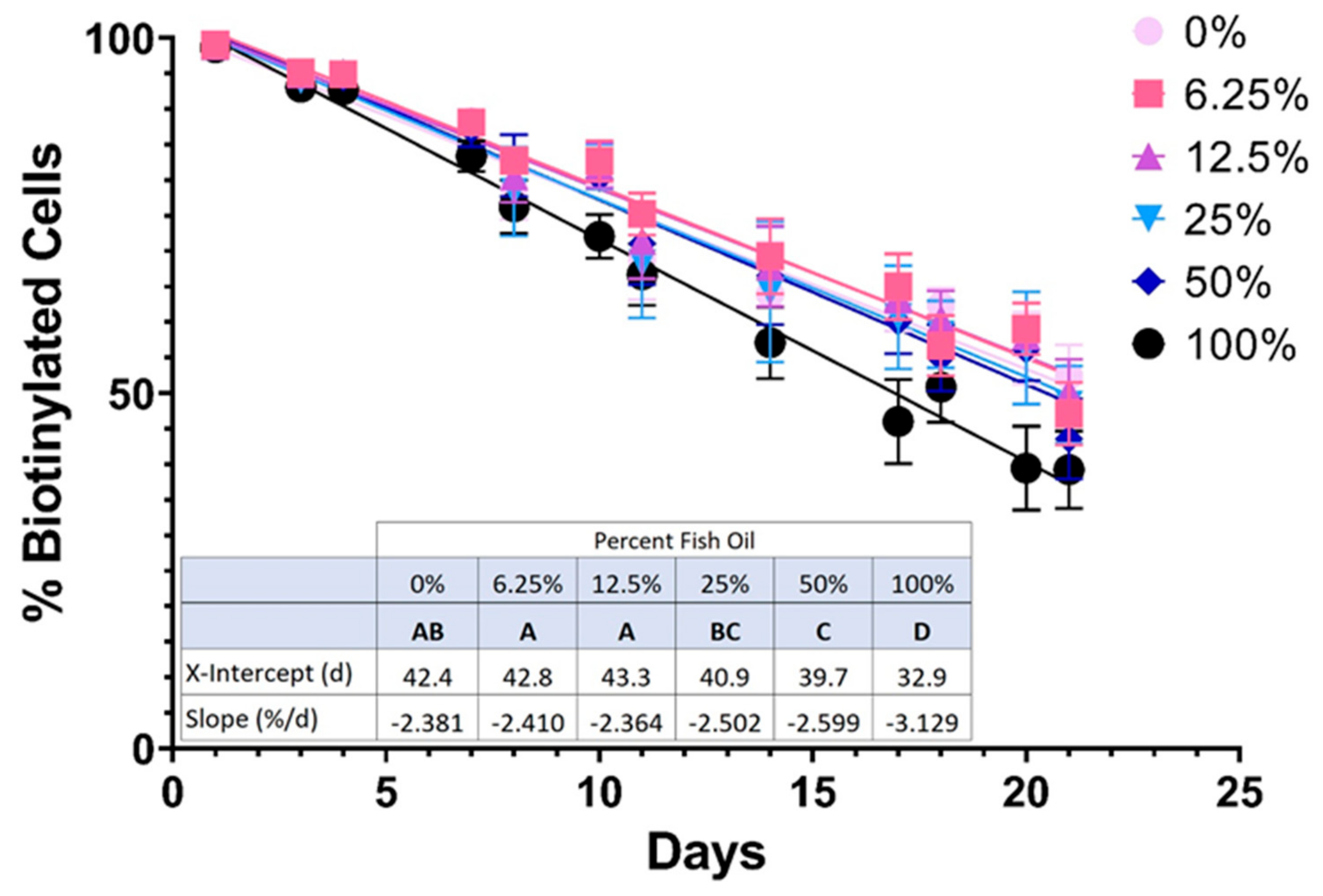
3.1.3. Higher Doses of FO Were Associated with Reduced RBC In Vivo Lifespan
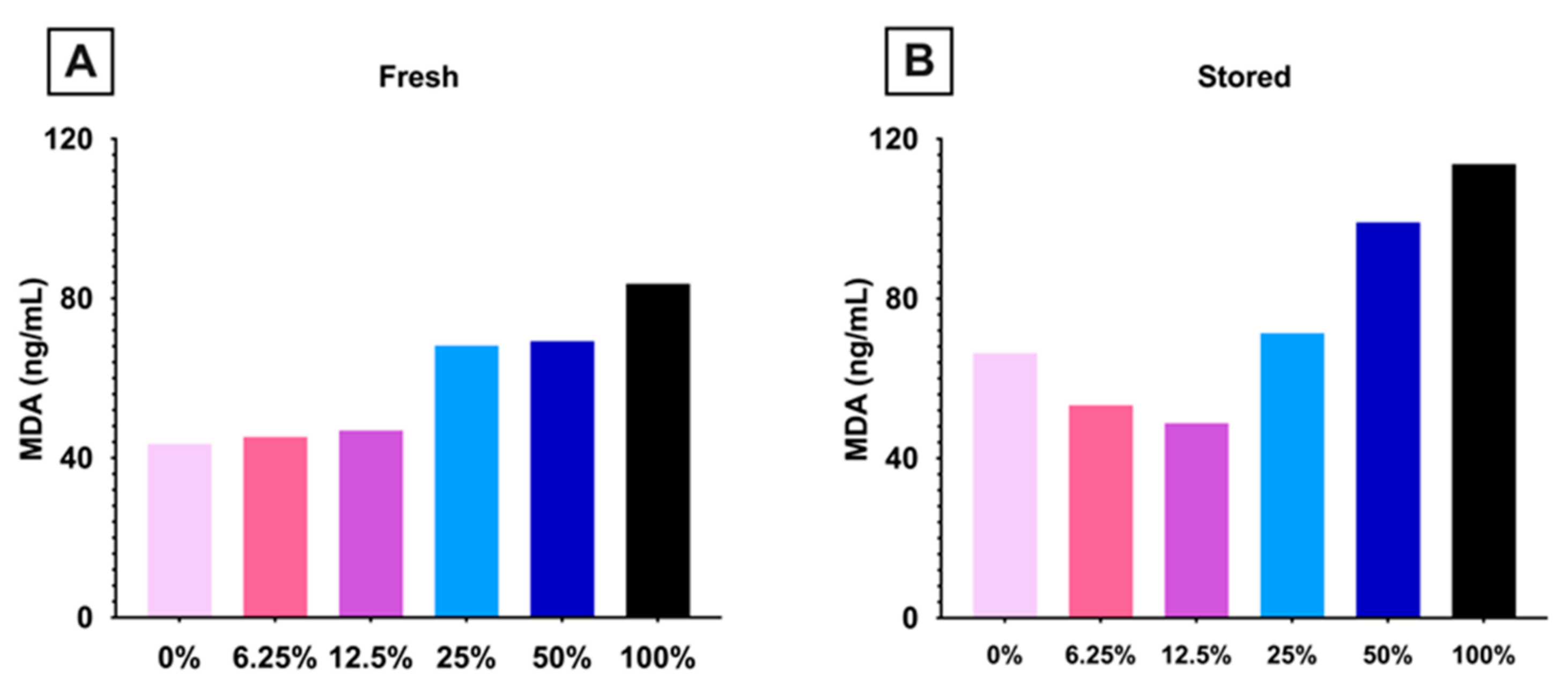
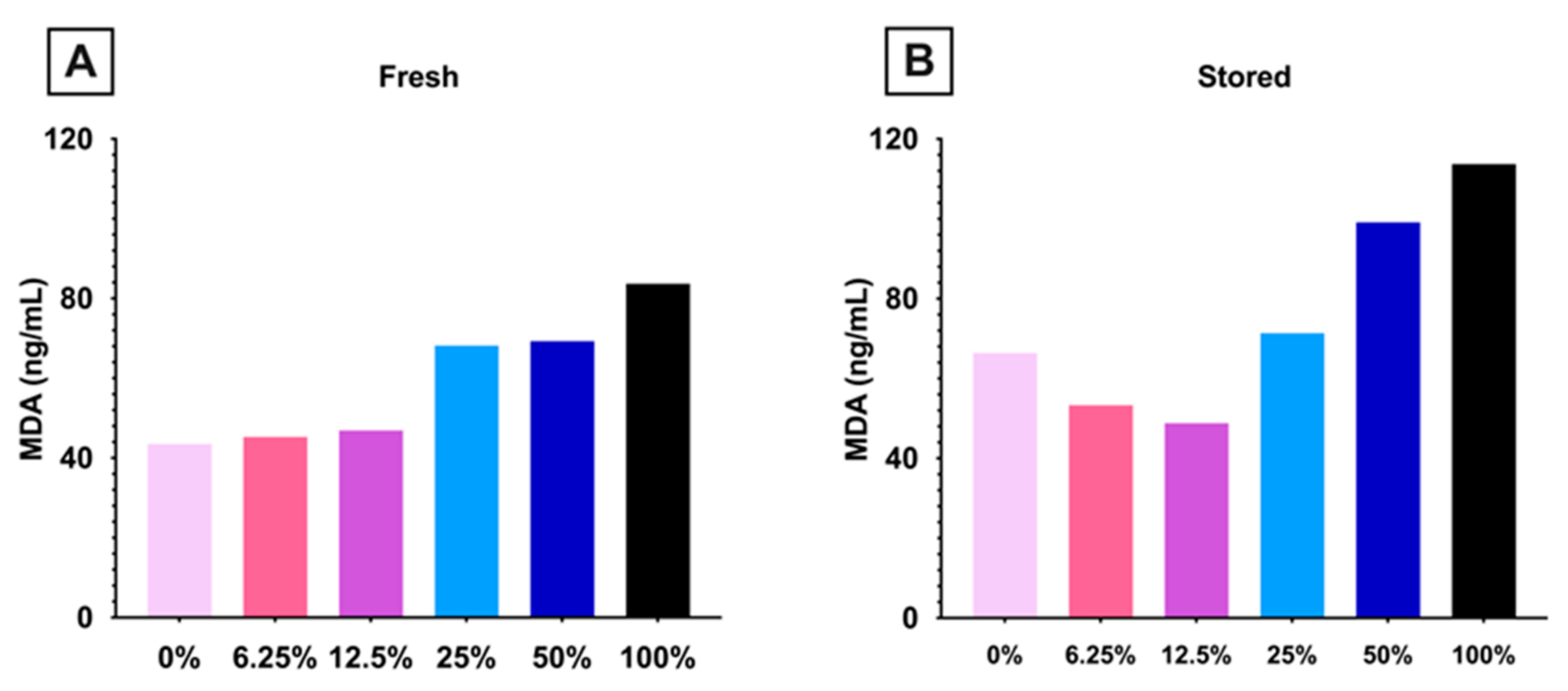
3.1.4. Lipid Peroxidation Increased in Mice Consuming High Dose FO
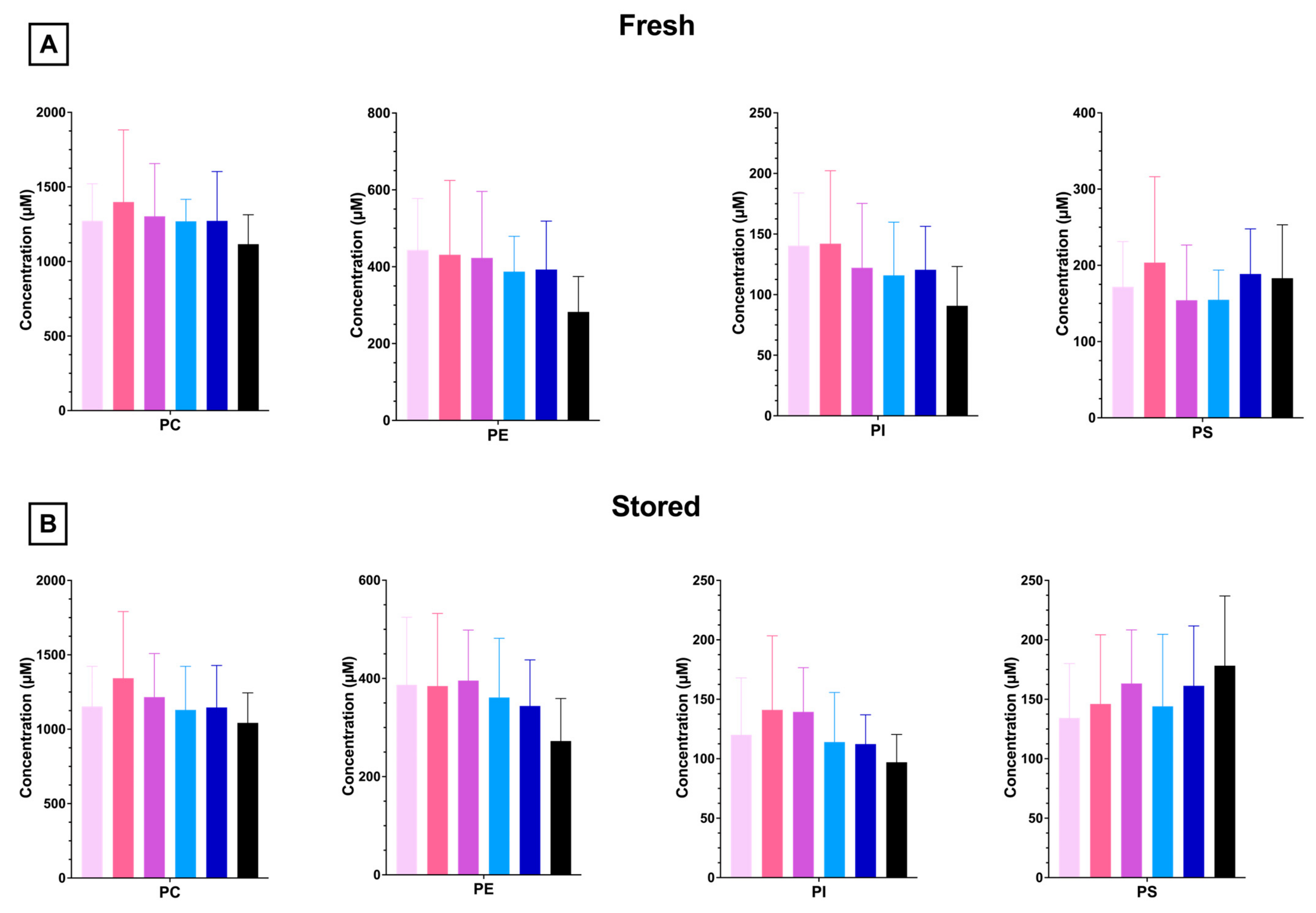
3.1.5. RBC PL and LPL Levels Are Unaffected by Dietary FO
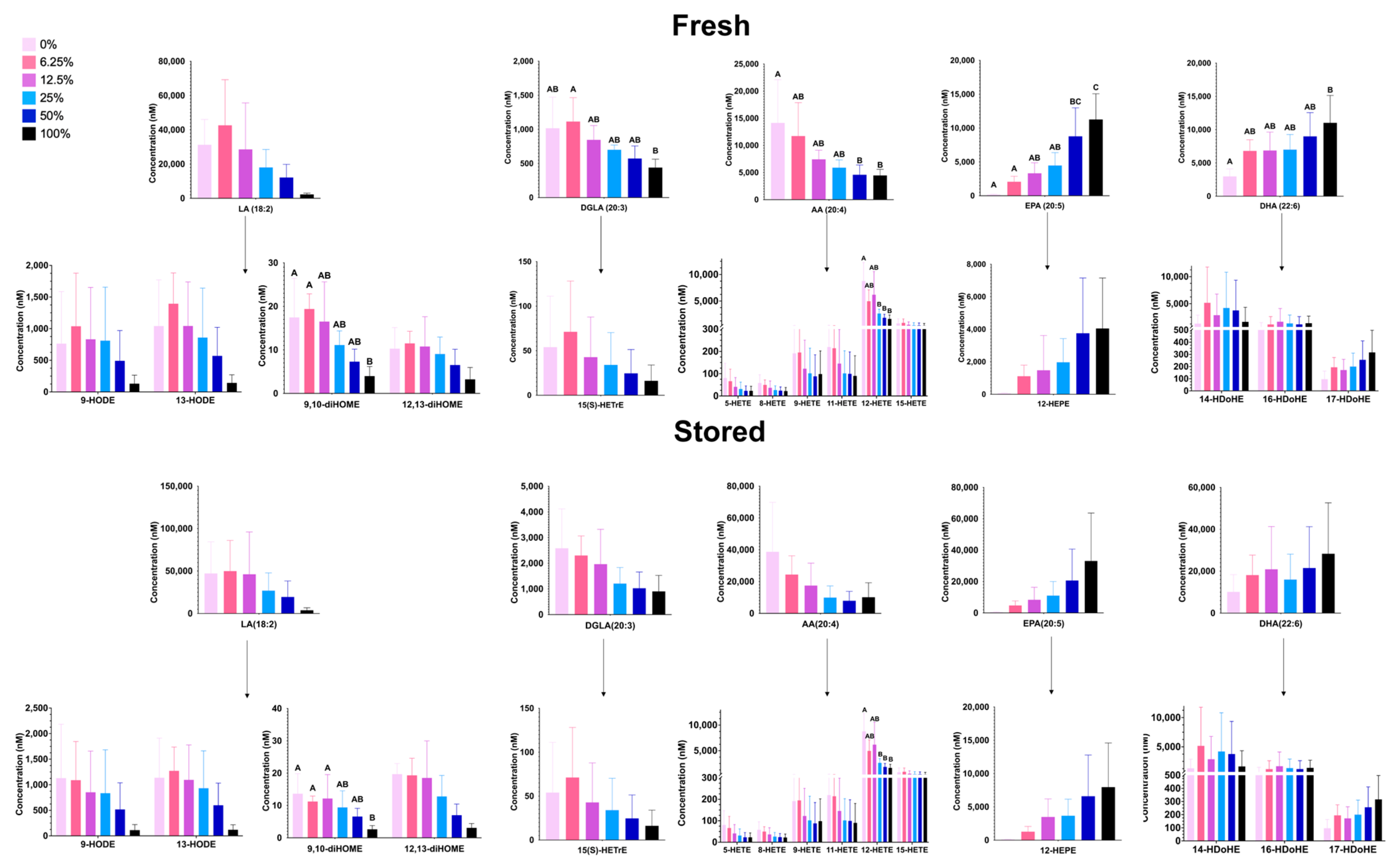
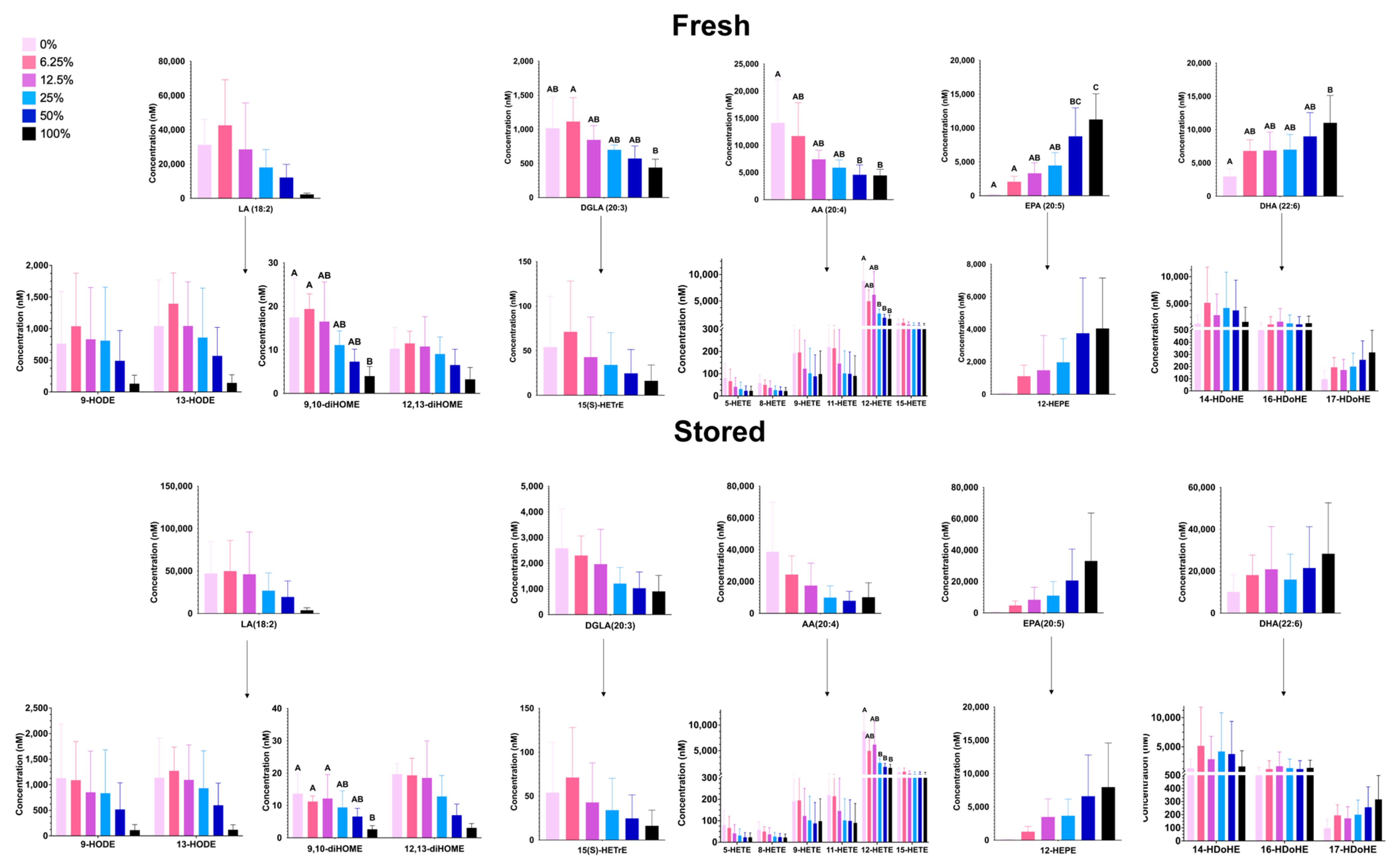
3.1.6. Free PUFAs and Oxylipins
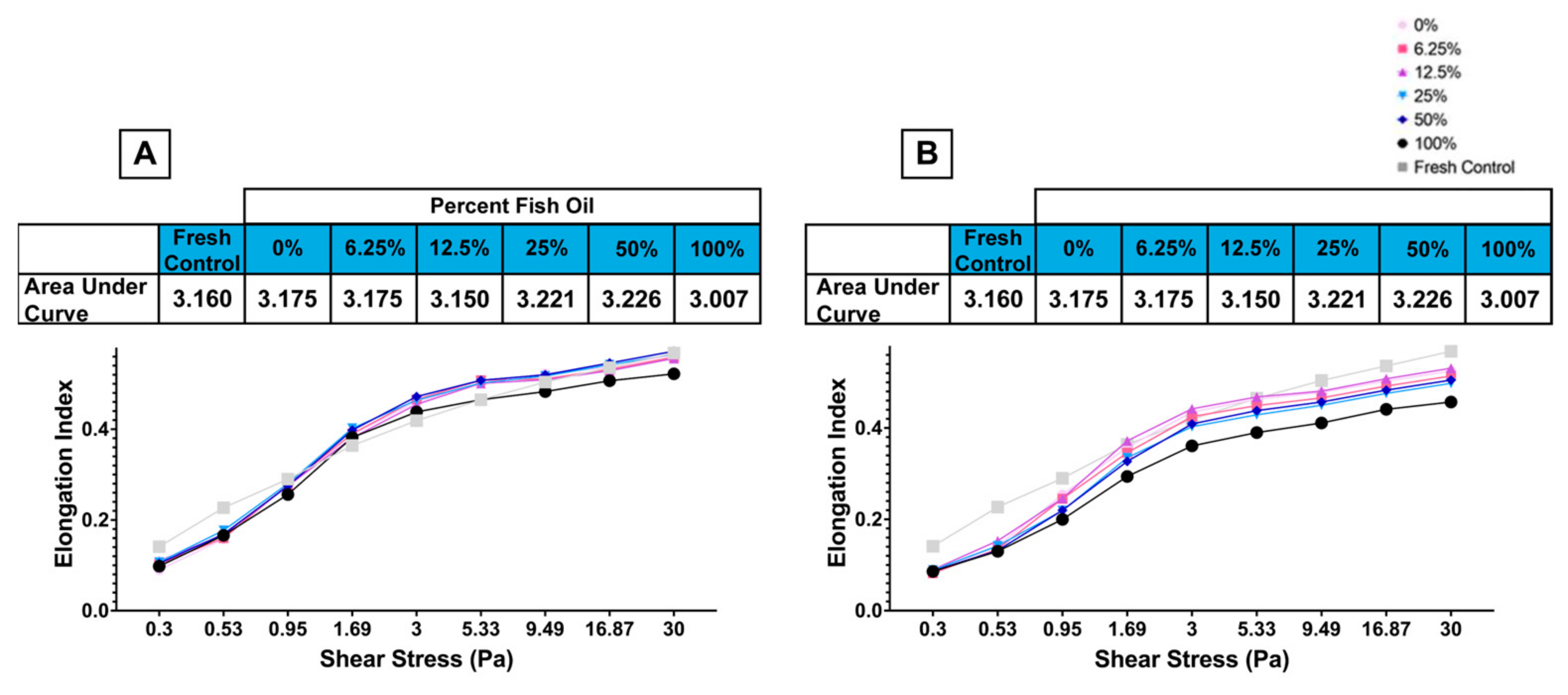
3.1.7. High Dose FO Is Associated with Reduced Deformability and Filterability
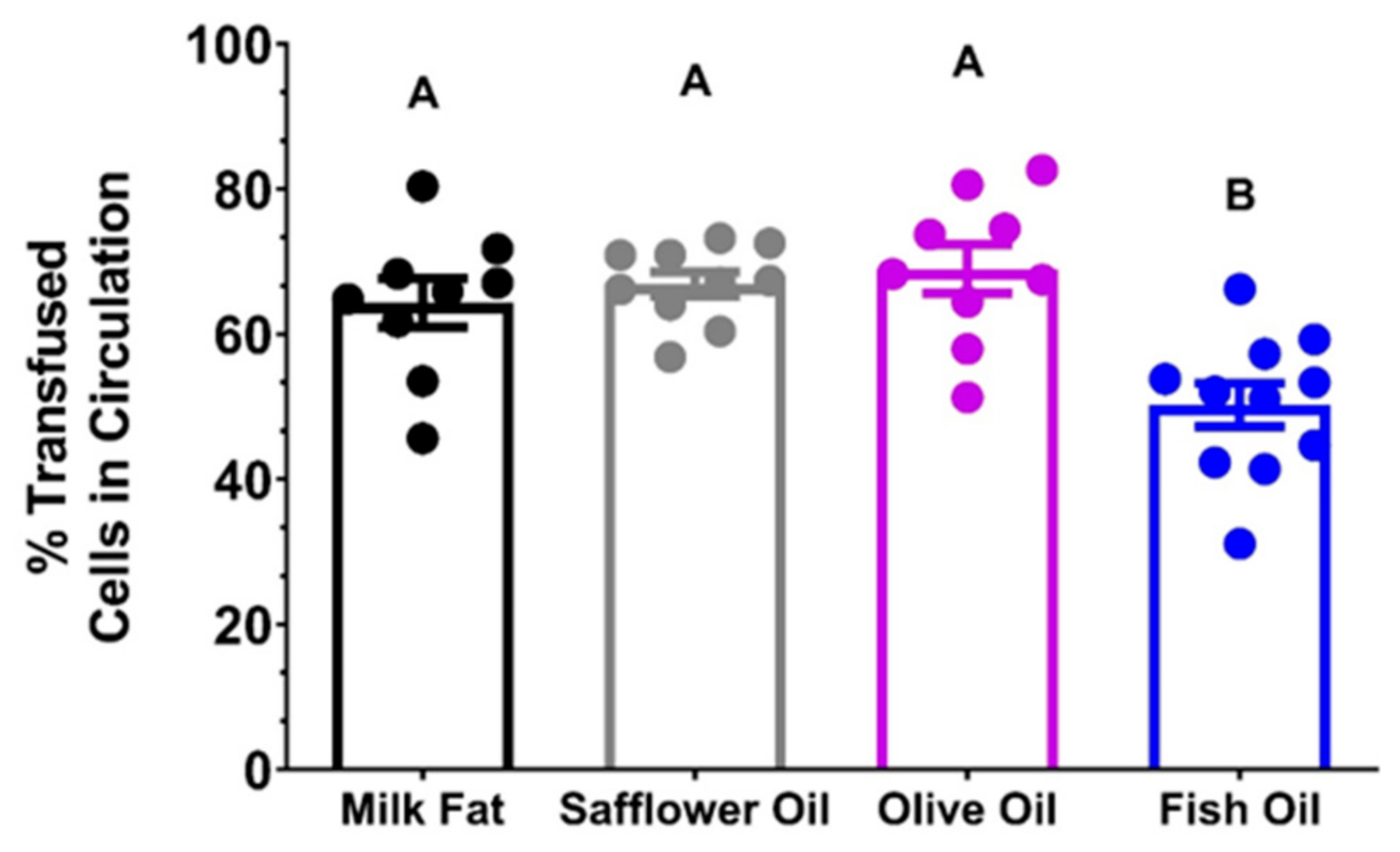
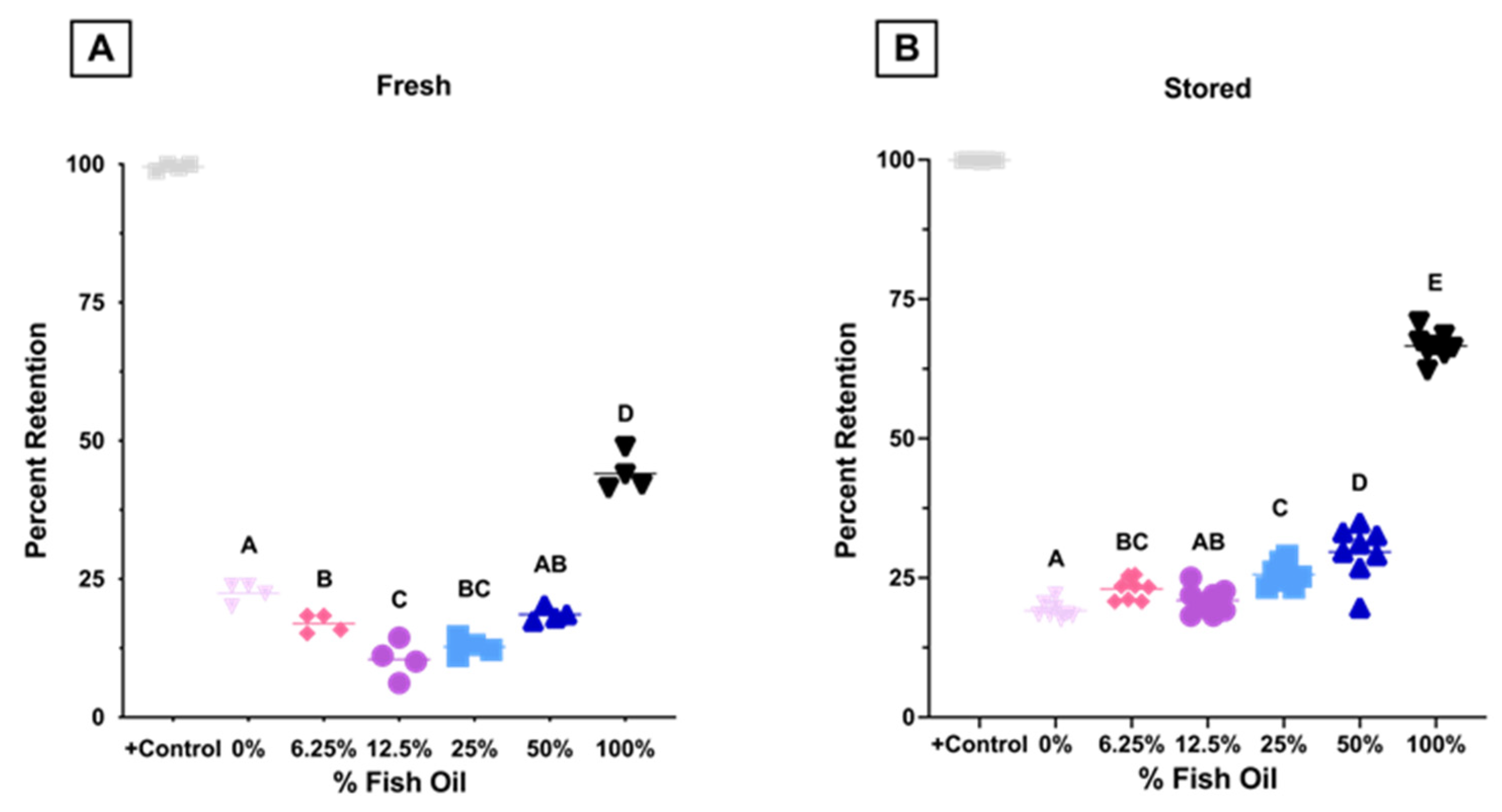
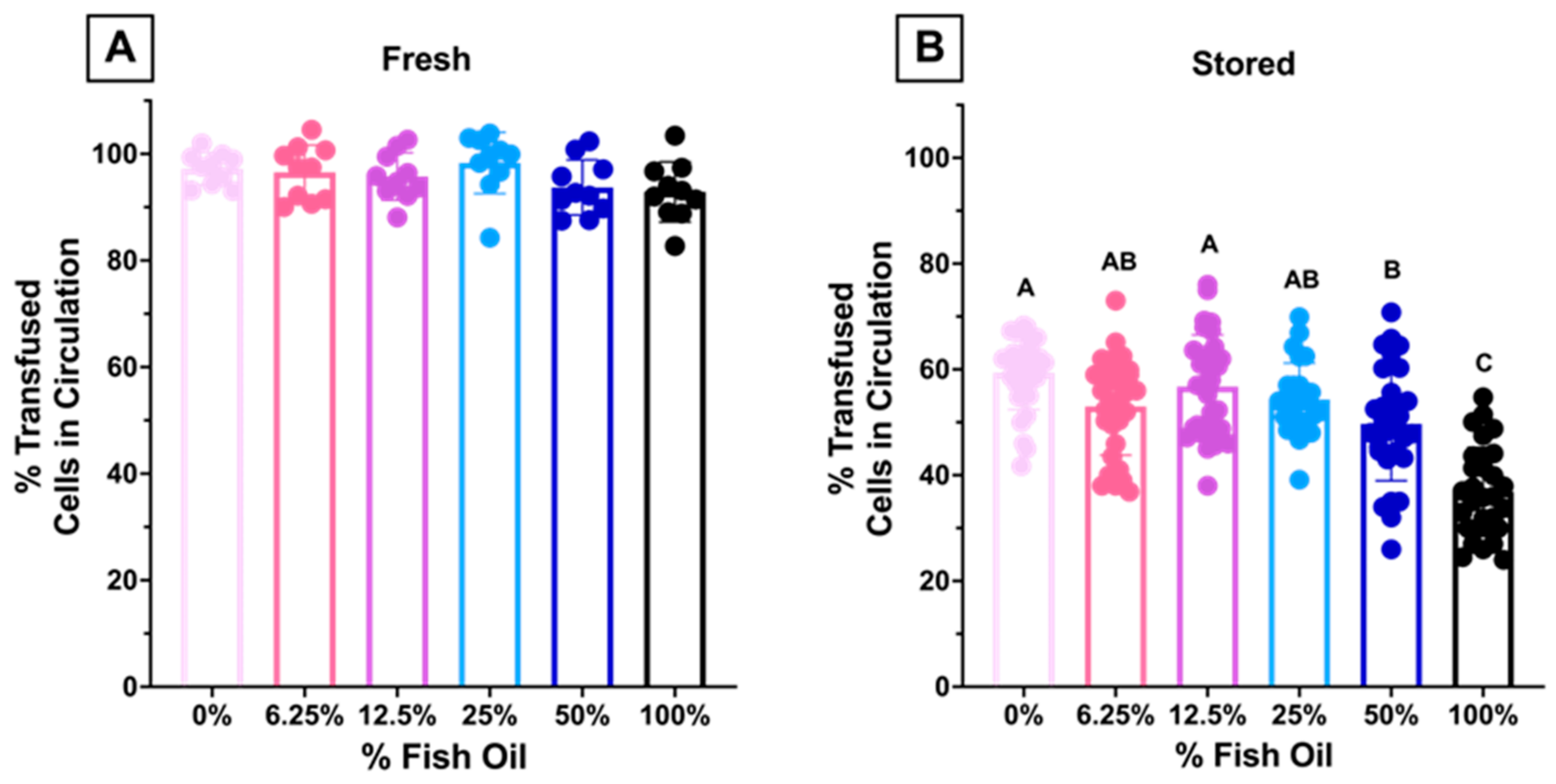
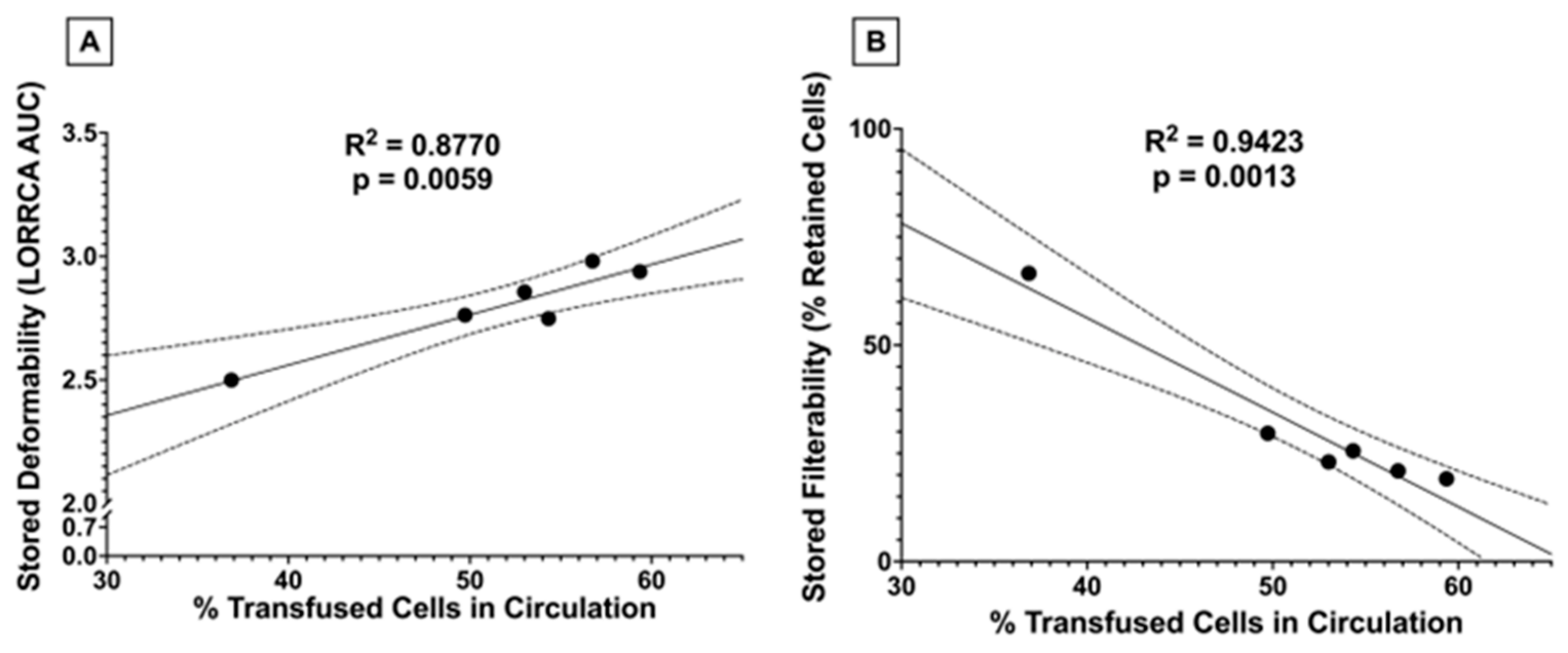
3.1.8. High Dose FO Diets Were Associated with Decreased 24 h PTR after Storage
3.1.9. Stored RBC 24 h PTR Correlates with Stored RBC Deformability and Filterability
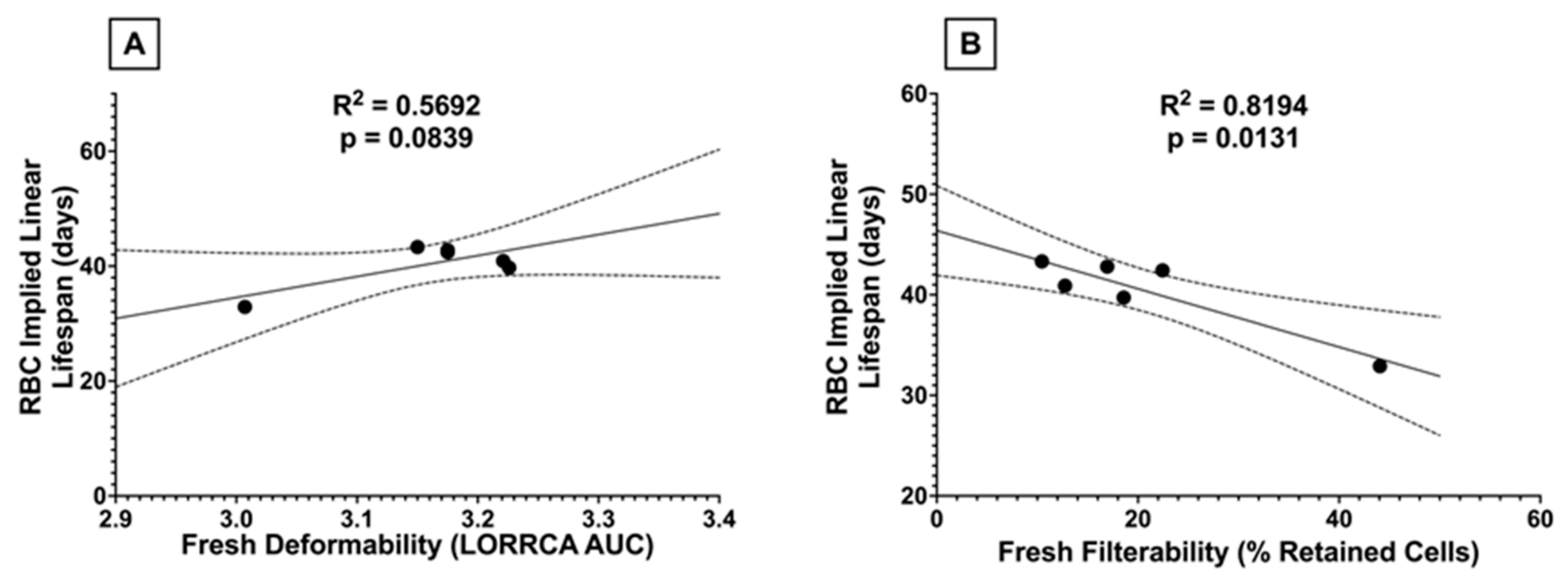
3.1.10. RBC Lifespan In Vivo Correlates with Fresh Filterability, but Not with Fresh Deformability
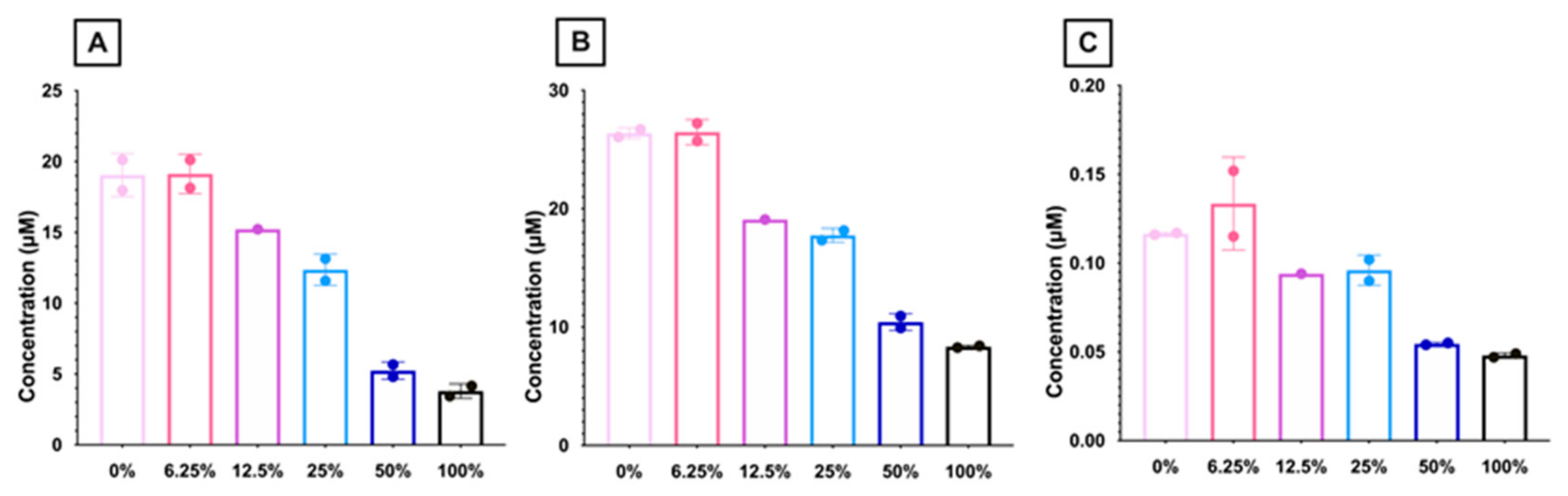
3.1.11. Carnitine Levels Decrease with Higher Dose FO Diets
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Heinicke, K.; Wolfarth, B.; Winchenbach, P.; Biermann, B.; Schmid, A.; Huber, G.; Friedmann, B.; Schmidt, W. Blood volume and hemoglobin mass in elite athletes of different disciplines. Int. J. Sports Med. 2001, 22, 504–512. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.M.; Sapiano, M.R.P.; Savinkina, A.A.; Haass, K.A.; Baker, M.L.; Henry, R.A.; Berger, J.J.; Basavaraju, S.V. Slowing decline in blood collection and transfusion in the United States-2017. Transfusion 2020, 60 (Suppl. S2), S1–S9. [Google Scholar] [CrossRef] [PubMed]
- Roback, J.D. Perspectives on the impact of storage duration on blood quality and transfusion outcomes. Vox Sang. 2016, 111, 357–364. [Google Scholar] [CrossRef] [PubMed]
- U.S. Food and Drug Administration. Workshop on Red Cell Stored in Additive Solution Systems; U.S. Food and Drug Administration: Bethesda, MD, USA, 1985.
- Luten, M.; Roerdinkholder-Stoelwinder, B.; Schaap, N.P.; de Grip, W.J.; Bos, H.J.; Bosman, G.J. Survival of red blood cells after transfusion: A comparison between red cells concentrates of different storage periods. Transfusion 2008, 48, 1478–1485. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, R.; Katharia, R. Oxidative injury as contributory factor for red cells storage lesion during twenty eight days of storage. Blood Transfus. 2012, 10, 59–62. [Google Scholar] [PubMed]
- Yoshida, T.; Prudent, M.; D’Alessandro, A. Red blood cell storage lesion: Causes and potential clinical consequences. Blood Transfus. 2019, 17, 27–52. [Google Scholar] [CrossRef] [PubMed]
- Bosman, G.J.; Lasonder, E.; Luten, M.; Roerdinkholder-Stoelwinder, B.; Novotny, V.M.; Bos, H.; De Grip, W.J. The proteome of red cell membranes and vesicles during storage in blood bank conditions. Transfusion 2008, 48, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Hovav, T.; Yedgar, S.; Manny, N.; Barshtein, G. Alteration of red cell aggregability and shape during blood storage. Transfusion 1999, 39, 277–281. [Google Scholar] [CrossRef]
- La Celle, P.L. Alteration of deformability of the erythrocyte membrane in stored blood. Transfusion 1969, 9, 238–245. [Google Scholar] [CrossRef]
- Berezina, T.L.; Zaets, S.B.; Morgan, C.; Spillert, C.R.; Kamiyama, M.; Spolarics, Z.; Deitch, E.A.; Machiedo, G.W. Influence of storage on red blood cell rheological properties. J. Surg. Res. 2002, 102, 6–12. [Google Scholar] [CrossRef]
- Cartwright, I.J.; Pockley, A.G.; Galloway, J.H.; Greaves, M.; Preston, F.E. The effects of dietary omega-3 polyunsaturated fatty acids on erythrocyte membrane phospholipids, erythrocyte deformability and blood viscosity in healthy volunteers. Atherosclerosis 1985, 55, 267–281. [Google Scholar] [CrossRef] [PubMed]
- Dougherty, R.M.; Galli, C.; Ferro-Luzzi, A.; Iacono, J.M. Lipid and phospholipid fatty acid composition of plasma, red blood cells, and platelets and how they are affected by dietary lipids: A study of normal subjects from Italy, Finland, and the USA. Am. J. Clin. Nutr. 1987, 45, 443–455. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Ma, J.; Campos, H.; Hankinson, S.E.; Hu, F.B. Comparison between plasma and erythrocyte fatty acid content as biomarkers of fatty acid intake in US women. Am. J. Clin. Nutr. 2007, 86, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Stierman, B.; Gahche, J.J.; Potischman, N. Dietary Supplement Use among Adults: United States, 2017–2018; NCHS Data Brief. 2021. Available online: https://stacks.cdc.gov/view/cdc/101131 (accessed on 21 September 2023).
- Harris, W.S.; Kris-Etherton, P.M.; Harris, K.A. Intakes of long-chain omega-3 fatty acid associated with reduced risk for death from coronary heart disease in healthy adults. Curr. Atheroscler. Rep. 2008, 10, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Kris-Etherton, P.M.; Harris, W.S.; Appel, L.J.; Nutrition, C. Fish consumption, fish oil, omega-3 fatty acids, and cardiovascular disease. Arterioscler. Thromb. Vasc. Biol. 2003, 23, e20–e30. [Google Scholar] [CrossRef]
- Katan, M.B.; Deslypere, J.P.; van Birgelen, A.P.; Penders, M.; Zegwaard, M. Kinetics of the incorporation of dietary fatty acids into serum cholesteryl esters, erythrocyte membranes, and adipose tissue: An 18-month controlled study. J. Lipid Res. 1997, 38, 2012–2022. [Google Scholar] [CrossRef]
- Dantas, L.S.; Viviani, L.G.; Inague, A.; Piccirillo, E.; Rezende, L.; Ronsein, G.E.; Augusto, O.; Medeiros, M.H.G.; Amaral, A.T.D.; Miyamoto, S. Lipid aldehyde hydrophobicity affects apo-SOD1 modification and aggregation. Free Radic. Biol. Med. 2020, 156, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Matsunaga, A.; Oe, T. Inhibition effect of pyridoxamine on lipid hydroperoxide-derived modifications to human serum albumin. PLoS ONE 2018, 13, e0196050. [Google Scholar] [CrossRef]
- Alviz-Amador, A.; Galindo-Murillo, R.; Pineda-Alemán, R.; Pérez-González, H.; Rodríguez-Cavallo, E.; Vivas-Reyes, R.; Méndez-Cuadro, D. Development and benchmark to obtain AMBER parameters dataset for non-standard amino acids modified with 4-hydroxy-2-nonenal. Data Brief 2018, 21, 2581–2589. [Google Scholar] [CrossRef]
- Kim, C.Y.; Johnson, H.; Peltier, S.; Spitalnik, S.L.; Hod, E.A.; Francis, R.O.; Hudson, K.E.; Stone, E.F.; Gordy, D.E.; Fu, X.; et al. Deuterated Linoleic Acid Attenuates the RBC Storage Lesion in a Mouse Model of Poor RBC Storage. Front. Physiol. 2022, 13, 868578. [Google Scholar] [CrossRef]
- Li, J.; Pora, B.L.R.; Dong, K.; Hasjim, J. Health benefits of docosahexaenoic acid and its bioavailability: A review. Food Sci. Nutr. 2021, 9, 5229–5243. [Google Scholar] [CrossRef] [PubMed]
- Barber, A.A.; Bernheim, F. Lipid peroxidation: Its measurement, occurrence, and significance in animal tissues. Adv. Gerontol. Res. 1967, 2, 355–403. [Google Scholar] [PubMed]
- Dholakia, U.; Bandyopadhyay, S.; Hod, E.A.; Prestia, K.A. Determination of RBC Survival in C57BL/6 and C57BL/6-Tg(UBC-GFP) Mice. Comp. Med. 2015, 65, 196–201. [Google Scholar] [PubMed]
- Zimring, J.C.; Smith, N.; Stowell, S.R.; Johnsen, J.M.; Bell, L.N.; Francis, R.O.; Hod, E.A.; Hendrickson, J.E.; Roback, J.D.; Spitalnik, S.L. Strain-specific red blood cell storage, metabolism, and eicosanoid generation in a mouse model. Transfusion 2014, 54, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Bessis, M.; Mohandas, N.; Feo, C. Automated ektacytometry: A new method of measuring red cell deformability and red cell indices. Blood Cells 1980, 6, 315–327. [Google Scholar] [PubMed]
- Marin, M.; Roussel, C.; Dussiot, M.; Ndour, P.A.; Hermine, O.; Colin, Y.; Gray, A.; Landrigan, M.; Le Van Kim, C.; Buffet, P.A.; et al. Metabolic rejuvenation upgrades circulatory functions of red blood cells stored under blood bank conditions. Transfusion 2021, 61, 903–918. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Anderson, M.; Wang, Y.; Zimring, J.C. LC-MS/MS-MRM-Based Targeted Metabolomics for Quantitative Analysis of Polyunsaturated Fatty Acids and Oxylipins. Methods Mol. Biol. 2019, 1978, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Felcyn, J.R.; Odem-Davis, K.; Zimring, J.C. Bioactive lipids accumulate in stored red blood cells despite leukoreduction: A targeted metabolomics study. Transfusion 2016, 56, 2560–2570. [Google Scholar] [CrossRef]
- Medved, J.; Knott, B.M.; Tarrah, S.N.; Li, A.N.; Shah, N.; Moscovich, T.C.; Boscia, A.R.; Salazar, J.E.; Santhanakrishnan, M.; Hendrickson, J.E.; et al. The lysophospholipid-binding molecule CD1D is not required for the alloimmunization response to fresh or stored RBCs in mice despite RBC storage driving alterations in lysophospholipids. Transfusion 2021, 61, 2169–2178. [Google Scholar] [CrossRef]
- Reisz, J.A.; Zheng, C.; D’Alessandro, A.; Nemkov, T. Untargeted and Semi-targeted Lipid Analysis of Biological Samples Using Mass Spectrometry-Based Metabolomics. Methods Mol. Biol. 2019, 1978, 121–135. [Google Scholar] [CrossRef]
- Nemkov, T.; Reisz, J.A.; Gehrke, S.; Hansen, K.C.; D’Alessandro, A. High-Throughput Metabolomics: Isocratic and Gradient Mass Spectrometry-Based Methods. Methods Mol. Biol. 2019, 1978, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Papaioannou, T.G.; Stefanadis, C. Vascular wall shear stress: Basic principles and methods. Hellenic J. Cardiol. 2005, 46, 9–15. [Google Scholar] [PubMed]
- Ballermann, B.J.; Dardik, A.; Eng, E.; Liu, A. Shear stress and the endothelium. Kidney Int. Suppl. 1998, 67, S100–S108. [Google Scholar] [CrossRef] [PubMed]
- Zierenberg, O.; Grundy, S.M. Intestinal absorption of polyenephosphatidylcholine in man. J. Lipid Res. 1982, 23, 1136–1142. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F. Marine oils from seafood waste. In Maximising the Value of Marine By-Products; Woodhead Publishing Series in Food Science, Technology and Nutrition; Woodhead Publishing: Sawston, UK, 2007; pp. 258–278. [Google Scholar]
- Chistyakov, D.V.; Gavrish, G.E.; Goriainov, S.V.; Chistyakov, V.V.; Astakhova, A.A.; Azbukina, N.V.; Sergeeva, M.G. Oxylipin Profiles as Functional Characteristics of Acute Inflammatory Responses in Astrocytes Pre-Treated with IL-4, IL-10, or LPS. Int. J. Mol. Sci. 2020, 21, 1780. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.; Cendali, F.; Fu, X.; Gamboni, F.; Morrison, E.J.; Beirne, J.; Nemkov, T.; Antonelou, M.H.; Kriebardis, A.; Welsby, I.; et al. Fatty acid desaturase activity in mature red blood cells and implications for blood storage quality. Transfusion 2021, 61, 1867–1883. [Google Scholar] [CrossRef]
- Fisher, A.B. The phospholipase A2 activity of peroxiredoxin 6. J. Lipid Res. 2018, 59, 1132–1147. [Google Scholar] [CrossRef]
- Burke, J.E.; Dennis, E.A. Phospholipase A2 biochemistry. Cardiovasc. Drugs Ther. 2009, 23, 49–59. [Google Scholar] [CrossRef]
- Leal, J.K.F.; Adjobo-Hermans, M.J.W.; Bosman, G.J.C.G. Red Blood Cell Homeostasis: Mechanisms and Effects of Microvesicle Generation in Health and Disease. Front. Physiol. 2018, 9, 703. [Google Scholar] [CrossRef]
- Willekens, F.L.; Werre, J.M.; Groenen-Döpp, Y.A.; Roerdinkholder-Stoelwinder, B.; de Pauw, B.; Bosman, G.J. Erythrocyte vesiculation: A self-protective mechanism? Br. J. Haematol. 2008, 141, 549–556. [Google Scholar] [CrossRef]
- Waugh, R.E.; Narla, M.; Jackson, C.W.; Mueller, T.J.; Suzuki, T.; Dale, G.L. Rheologic properties of senescent erythrocytes: Loss of surface area and volume with red blood cell age. Blood 1992, 79, 1351–1358. [Google Scholar] [CrossRef] [PubMed]
- Willekens, F.L.; Werre, J.M.; Kruijt, J.K.; Roerdinkholder-Stoelwinder, B.; Groenen-Döpp, Y.A.; van den Bos, A.G.; Bosman, G.J.; van Berkel, T.J. Liver Kupffer cells rapidly remove red blood cell-derived vesicles from the circulation by scavenger receptors. Blood 2005, 105, 2141–2145. [Google Scholar] [CrossRef] [PubMed]
- Virmani, M.A.; Cirulli, M. The Role of l-Carnitine in Mitochondria, Prevention of Metabolic Inflexibility and Disease Initiation. Int. J. Mol. Sci. 2022, 23, 2717. [Google Scholar] [CrossRef] [PubMed]
- Houten, S.M.; Wanders, R.J.A.; Ranea-Robles, P. Metabolic interactions between peroxisomes and mitochondria with a special focus on acylcarnitine metabolism. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165720. [Google Scholar] [CrossRef] [PubMed]
- Ribas, G.S.; Vargas, C.R.; Wajner, M. L-carnitine supplementation as a potential antioxidant therapy for inherited neurometabolic disorders. Gene 2014, 533, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Toptas, B.; Baykal, A.; Yesilipek, A.; Isbir, M.; Kupesiz, A.; Yalcin, O.; Baskurt, O.K. L-carnitine deficiency and red blood cell mechanical impairment in beta-thalassemia major. Clin. Hemorheol. Microcirc. 2006, 35, 349–357. [Google Scholar] [PubMed]
- Tzounakas, V.L.; Anastasiadi, A.T.; Arvaniti, V.Z.; Lelli, V.; Fanelli, G.; Paronis, E.C.; Apostolidou, A.C.; Balafas, E.G.; Kostomitsopoulos, N.G.; Papageorgiou, E.G.; et al. Supplementation with uric and ascorbic acid protects stored red blood cells through enhancement of non-enzymatic antioxidant activity and metabolic rewiring. Redox Biol. 2022, 57, 102477. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Percent Fish Oil | ||||||
|---|---|---|---|---|---|---|
| Fat Source | 0% | 6.25% | 12.5 | 25% | 50% | 100% |
| Concentration in Diet (g/kg) | ||||||
| Milkfat | 47.3 | 44.3 | 41.3 | 35.4 | 23.6 | 0.0 |
| Olive Oil | 13.5 | 12.7 | 11.8 | 10.1 | 6.8 | 0.0 |
| Safflower Oil | 60.8 | 57.0 | 53.2 | 45.6 | 30.4 | 0.0 |
| Canola Oil | 13.5 | 12.7 | 11.8 | 10.1 | 6.8 | 0.0 |
| Menhaden Oil | 0.0 | 8.4 | 16.9 | 33.8 | 67.5 | 135.0 |
| A | Percent Fish Oil | ||||||
|---|---|---|---|---|---|---|---|
| Total Fatty Acid | 0% | 6.25% | 12.5% | 25% | 50% | 100% | |
| Concentration in Diet (µmol/g) | |||||||
| C12 | Lauric Acid | 10.3 ± 2.8 A | 8.2 ± 1.7 A | 9.9 ± 4.3 A | 4.8 ± 2.1 AB | 4.9 ± 0.8 B | 1.1 ± 0.1 B |
| C14 | Myristic Acid | 18.2 ± 3 | 16.2 ± 4.2 | 18.9 ± 5.6 | 14.9 ± 2 | 25.1 ± 8 | 26.3 ± 1.2 |
| C16 | Palmitic Acid | 154.3 ± 10.4 A | 129.6 ± 2 BC | 115.2 ± 10.8 B | 127.0 ± 3.9 BC | 134.3 ± 4.7 C | 125.7 ± 3.7 BC |
| C16:1 | Palmitoleic Acid | 0.7 ± 0.1 A | 1.2 ± 0.3 A | 2.0 ± 0.7 A | 2.4 ± 0.2 A | 6.6 ± 3.6 B | 0.7 ± 0.1 A |
| C18 | Stearic Acid | 62.6 ± 2.5 A | 50.6 ± 4.6 AB | 41.3 ± 9.7 B | 53.8 ± 1.6 AB | 48.2 ± 6.1 B | 46.6 ± 1.3 B |
| C18:1 | Oleic Acid | 85.3 ± 10.3 A | 72.7 ± 17.4 A | 70.8 ± 17.3 A | 56.5 ± 4.1 A | 57.2 ± 20.7 A | 14.8 ± 1 B |
| C18:2 | Linoleic Acid (LA) | 128.5 ± 14.1 A | 107.9 ± 22.7 AB | 106.4 ± 23.9 AB | 83.4 ± 6.8 AB | 75.8 ± 25.1 B | 11.5 ± 0.5 C |
| C18:3 | Alpha-Linoleic Acid (ALA) | 3.9 ± 0.4 | 3.6 ± 0.4 | 3.8 ± 0.5 | 4.0 ± 0.2 | 5.5 ± 1.6 | 4.9 ± 0.5 |
| C20 | Arachidic acid | 1.3 ± 0.1 A | 1.2 ± 0.2 A | 1.1 ± 0.1 AB | 1.2 ± 0.1 A | 1.2 ± 0.2 A | 0.8 ± 0 B |
| C20:1 | Eicosenoic Acid | 0.8 ± 0.1 A | 0.9 ± 0.1 A | 0.9 ± 0.1 A | 1.0 ± 0.1 A | 1.7 ± 0.5 B | 1.8 ± 0.1 B |
| C20:4 | Arachidonic Acid (AA) | 0.2 ± 0 A | 0.3 ± 0.1 A | 0.5 ± 0.1 A | 0.8 ± 0 A | 1.7 ± 0.7 B | 2.2 ± 0.3 B |
| C20:5 | Eicosapentaenoic Acid (EPA) | 0.1 ± 0 A | 2.1 ± 0.4 A | 4.4 ± 0.9 A | 8.3 ± 0.5 A | 22.0 ± 8.3 B | 28.5 ± 2.6 B |
| C22:1 | Erucic acid | 1.0 ± 0.1 | 1.3 ± 0.1 | 1.2 ± 0.3 | 1.1 ± 0.1 | 1.0 ± 0.3 | 0.9 ± 0.2 |
| C22:5 | Docosapentaenoic Acid (DPA) | 0.1 ± 0 A | 0.3 ± 0.1 A | 0.6 ± 0.1 A | 1.0 ± 0.1 A | 2.9 ± 1 B | 3.6 ± 0.4 B |
| C22:6 | Docosahexaenoic Acid (DHA) | 0.3 ± 0 A | 1.7 ± 0.3 A | 3.3 ± 0.7 A | 6.4 ± 0.7 A | 15.6 ± 6 B | 20.1 ± 2.1 B |
| B | Percent Fish Oil | ||||||
| Total Fatty Acid | 0% | 6.25% | 12.5% | 25% | 50% | 100% | |
| Concentration in Fresh RBCs (µmol/L) | |||||||
| C12 | Lauric Acid | 6.0 | 8.2 | 13.0 | 9.7 | 4.2 | 7.5 |
| C14 | Myristic Acid | 59.3 | 105.8 | 118.1 | 107.0 | 122.8 | 125.4 |
| C16 | Palmitic Acid | 6134.9 | 9189.9 | 9355.5 | 8425.4 | 7450.5 | 6640.1 |
| C16:1 | Palmitoleic Acid | 77.8 | 112.7 | 124.9 | 120.3 | 160.3 | 203.3 |
| C18 | Stearic Acid | 2446.2 | 3143.7 | 3247.2 | 2777.9 | 2221.2 | 2137.9 |
| C18:1 | Oleic Acid | 1787.3 | 2686.0 | 2779.4 | 2235.7 | 1662.9 | 1215.4 |
| C18:2 | Linoleic Acid (LA) | 2108.0 | 3309.4 | 3588.2 | 2662.9 | 1643.7 | 321.6 |
| C18:3 | Alpha-Linoleic Acid (ALA) | 12.4 | 16.8 | 22.9 | 17.7 | 21.0 | 16.7 |
| C20 | Arachidic acid | 40.7 | 60.9 | 58.9 | 58.0 | 41.6 | 35.9 |
| C20:1 | Eicosenoic Acid | 35.7 | 50.4 | 48.6 | 33.0 | 33.0 | 20.2 |
| C20:4 | Arachidonic Acid (AA) | 2539.9 | 2601.8 | 2134.7 | 1457.8 | 855.5 | 809.8 |
| C20:5 | Eicosapentaenoic Acid (EPA) | 10.4 | 241.8 | 510.9 | 800.6 | 1036.9 | 1187.3 |
| C22:1 | Erucic acid | 26.5 | 23.1 | 26.5 | 24.0 | 23.5 | 23.7 |
| C22:5 | Docosapentaenoic Acid (DPA) | 47.0 | 193.3 | 265.5 | 293.9 | 305.0 | 253.4 |
| C22:6 | Docosahexaenoic Acid (DHA) | 638.0 | 1380.5 | 1595.3 | 1574.0 | 1354.1 | 1299.3 |
| CBC Measurement | Units | Percent Fish Oil | |||||
|---|---|---|---|---|---|---|---|
| 0% | 6.25% | 12.5% | 25% | 50% | 100% | ||
| Hematocrit | (%) | 36.2% ± 2.0% | 35.4% ± 3.8% | 33.7% ± 2.1% | 37.5% ± 1.3% | 37.2% ± 2.7% | 36.1% ± 0.9% |
| RBC Count | (1012/L) | 9.0 ± 0.6 | 8.4 ± 0.7 | 8.2 ± 0.5 | 8.9 ± 0.3 | 8.9 ± 0.5 | 8.7 ± 0.2 |
| Hemoglobin | (g/dL) | 11.5 ± 0.6 | 11.2 ± 1.2 | 10.4 ± 0.7 | 11.6 ± 0.5 | 11.7 ± 0.7 | 11.0 ± 0.3 |
| RBC Distribution Width | (%) | 18.5% ± 1.5% | 17.9% ± 1.0% | 18.2% ± 1.6% | 17.4% ± 0.4% | 18.3% ± 0.7% | 18.8% ± 0.7% |
| Mean Corpuscular Volume (MCV) | (fL) | 52.5 ± 4.5 | 51.2 ± 4.7 | 48.7 ± 1.4 | 50.1 ± 1.1 | 50.7 ± 3.0 | 50.1 ± 2.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, C.Y.; Larsen, H.J.; Spitalnik, S.L.; Hod, E.A.; Francis, R.O.; Hudson, K.E.; Gordy, D.E.; Stone, E.F.; Peltier, S.; Amireault, P.; et al. Low-Dose Dietary Fish Oil Improves RBC Deformability without Improving Post-Transfusion Recovery in Mice. Nutrients 2023, 15, 4456. https://doi.org/10.3390/nu15204456
Kim CY, Larsen HJ, Spitalnik SL, Hod EA, Francis RO, Hudson KE, Gordy DE, Stone EF, Peltier S, Amireault P, et al. Low-Dose Dietary Fish Oil Improves RBC Deformability without Improving Post-Transfusion Recovery in Mice. Nutrients. 2023; 15(20):4456. https://doi.org/10.3390/nu15204456
Chicago/Turabian StyleKim, Christopher Y., Hannah J. Larsen, Steven L. Spitalnik, Eldad A. Hod, Richard O. Francis, Krystalyn E. Hudson, Dominique E. Gordy, Elizabeth F. Stone, Sandy Peltier, Pascal Amireault, and et al. 2023. "Low-Dose Dietary Fish Oil Improves RBC Deformability without Improving Post-Transfusion Recovery in Mice" Nutrients 15, no. 20: 4456. https://doi.org/10.3390/nu15204456
APA StyleKim, C. Y., Larsen, H. J., Spitalnik, S. L., Hod, E. A., Francis, R. O., Hudson, K. E., Gordy, D. E., Stone, E. F., Peltier, S., Amireault, P., D’Alessandro, A., Zimring, J. C., Buehler, P. W., Fu, X., & Thomas, T. (2023). Low-Dose Dietary Fish Oil Improves RBC Deformability without Improving Post-Transfusion Recovery in Mice. Nutrients, 15(20), 4456. https://doi.org/10.3390/nu15204456