

Optimization of Se- and Zn-Enriched Mycelium of Lentinula edodes (Berk.) Pegler as a Dietary Supplement with Immunostimulatory Activity

 ,
,  , ,
, ,  , and
, and
Abstract
1. Introduction
- In the first step, we optimized the composition of the culture medium to obtain a L. edodes mycelia enriched in Se and Zn at the assumed concentrations;
- Then, we prepared mycelial extracts with defined polysaccharide, selenium, and zinc contents to test the expected biological activity;
- In the last stage of the work, we determined the effects of L. edodes mycelium extracts with different concentrations of selenium and zinc on human T cells activation.
2. Materials and Methods
2.1. Microorganism and Cultivation Media
Supplementation of Media and Growth Conditions in Mycelial Shake-Flask Cultures
2.2. Determination of Selenium and Zinc in L. edodes Mycelium
2.2.1. Microwave Mineralization Procedure
2.2.2. Determination of Zinc Content by High-Performance Ion Chromatography (HPIC)
2.2.3. Determination of the Selenium Content via Reverse-Phase High-Performance Liquid Chromatography (RPHPLC)
2.3. Determination of the Selenium and Zinc Content in Mycelial Water Extracts via Inductively Coupled Plasma Mass Spectrometry (ICP-MS)
2.4. Preparation of the Mycelial Extracts
- Mycelium cultured in medium not enriched with micronutrients (sample 0);
- Mycelium cultured in medium enriched with selenium only at a concentration of 0.8 mM (sample Se);
- Mycelium cultured in medium enriched with zinc only at a concentration of 0.8 mM (sample Zn);
- Mycelium cultured in medium enriched with selenium at a concentration of 0.8 mM and zinc at a concentration of 0.2 mM (sample Se/Zn);
- Mycelium enriched with selenium at a concentration of 0.2 mM and zinc at a concentration of 0.8 mM (sample Zn/Se).
2.5. Determination of the Carbohydrate Content in the Mycelial Extracts
2.6. Determination of the Effects of Mycelial Extracts on T Cell Activation
2.6.1. The Isolation, Culture, and Stimulation of PBMCs
2.6.2. Flow Cytometry
2.7. Statistical Analysis
3. Results
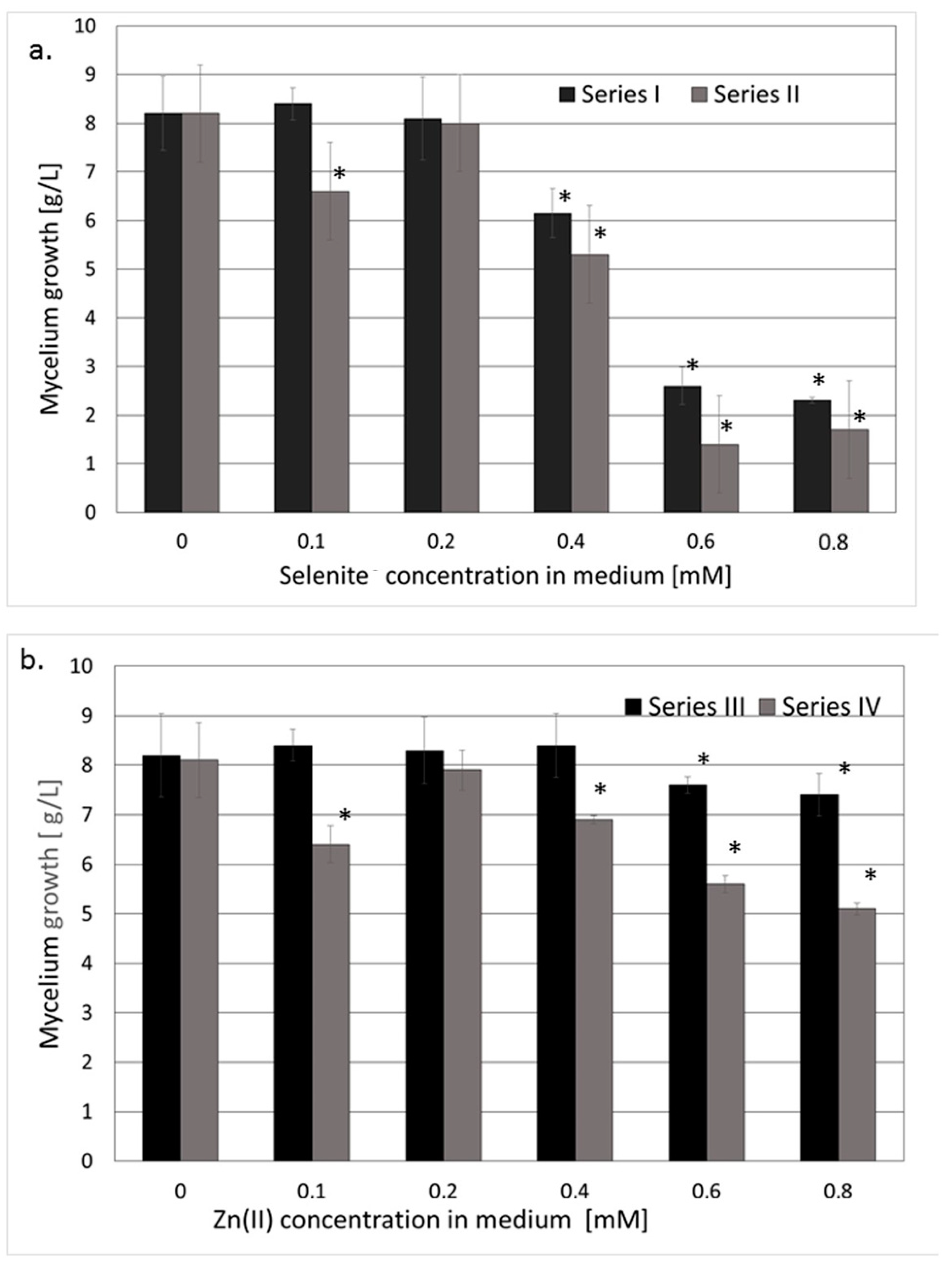
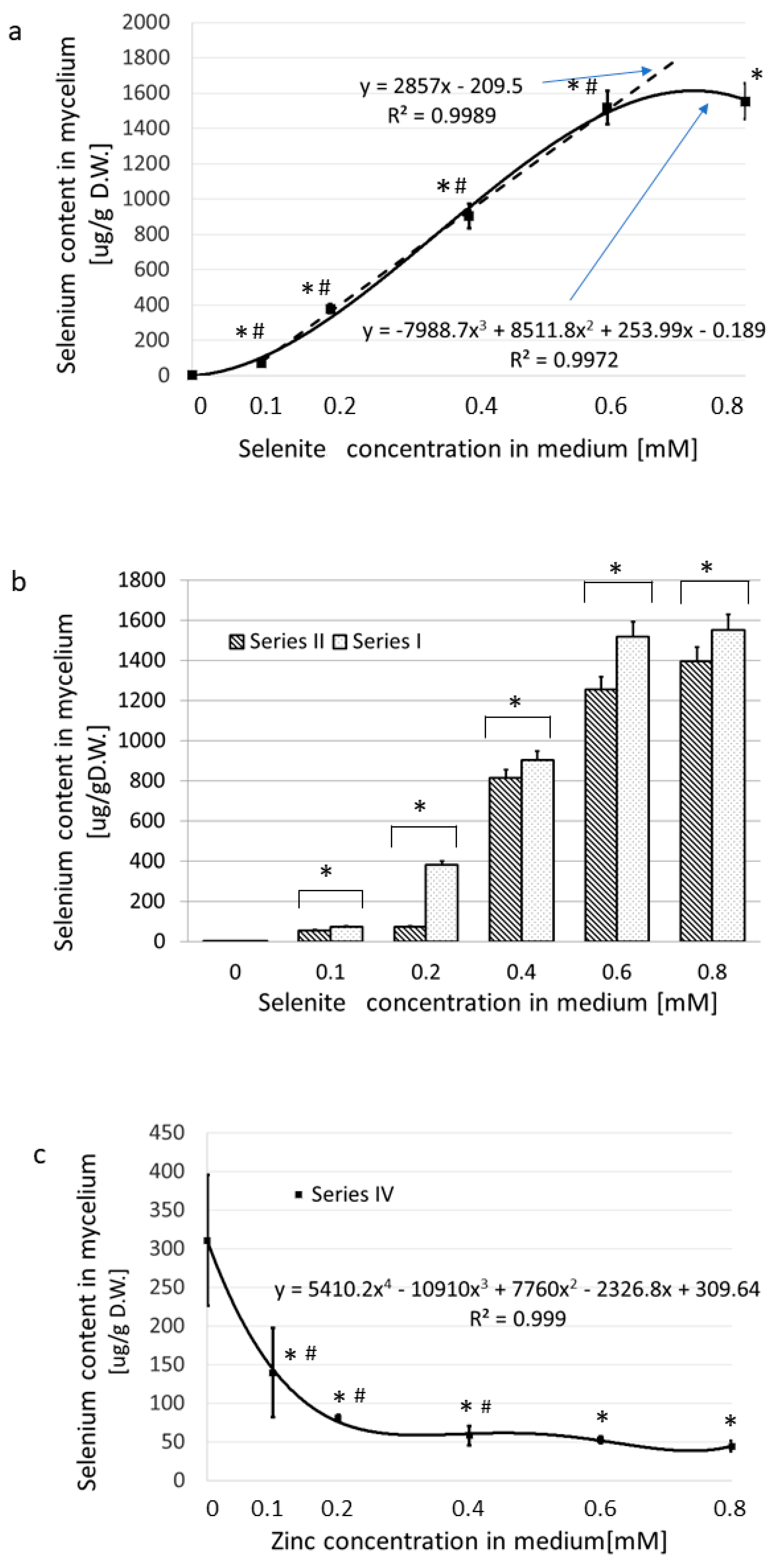
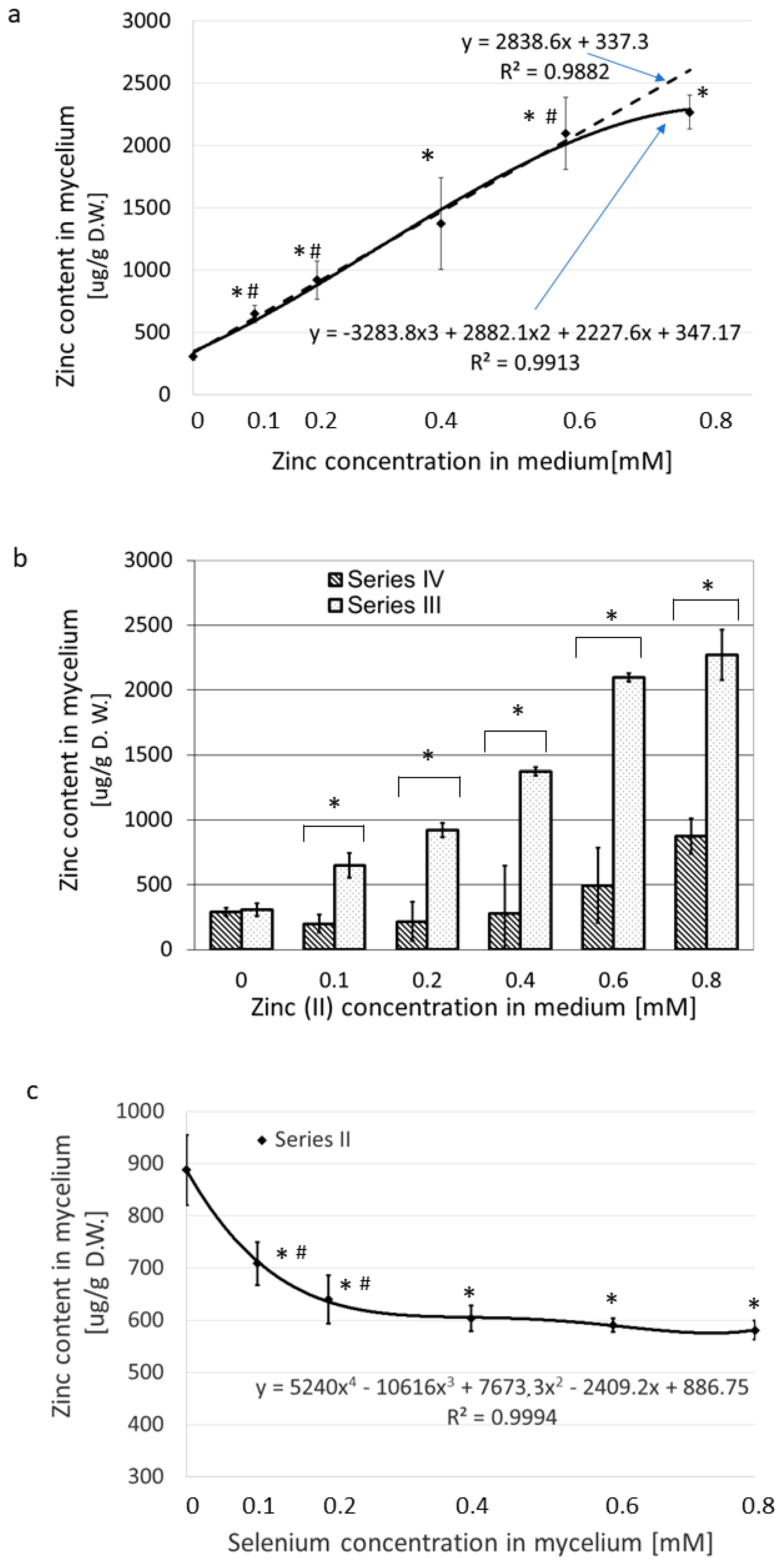
3.1. Accumulation of Se and Zn in the Mycelium of L. edodes
3.1.1. Reference Series and Mycelial Growth
3.1.2. The Accumulation of Se Exclusively versus in the Presence of Zn(II)
3.1.3. Zn(II) Accumulation Exclusively versus in the Presence of SeO32−
3.2. Composition of the Water Extracts
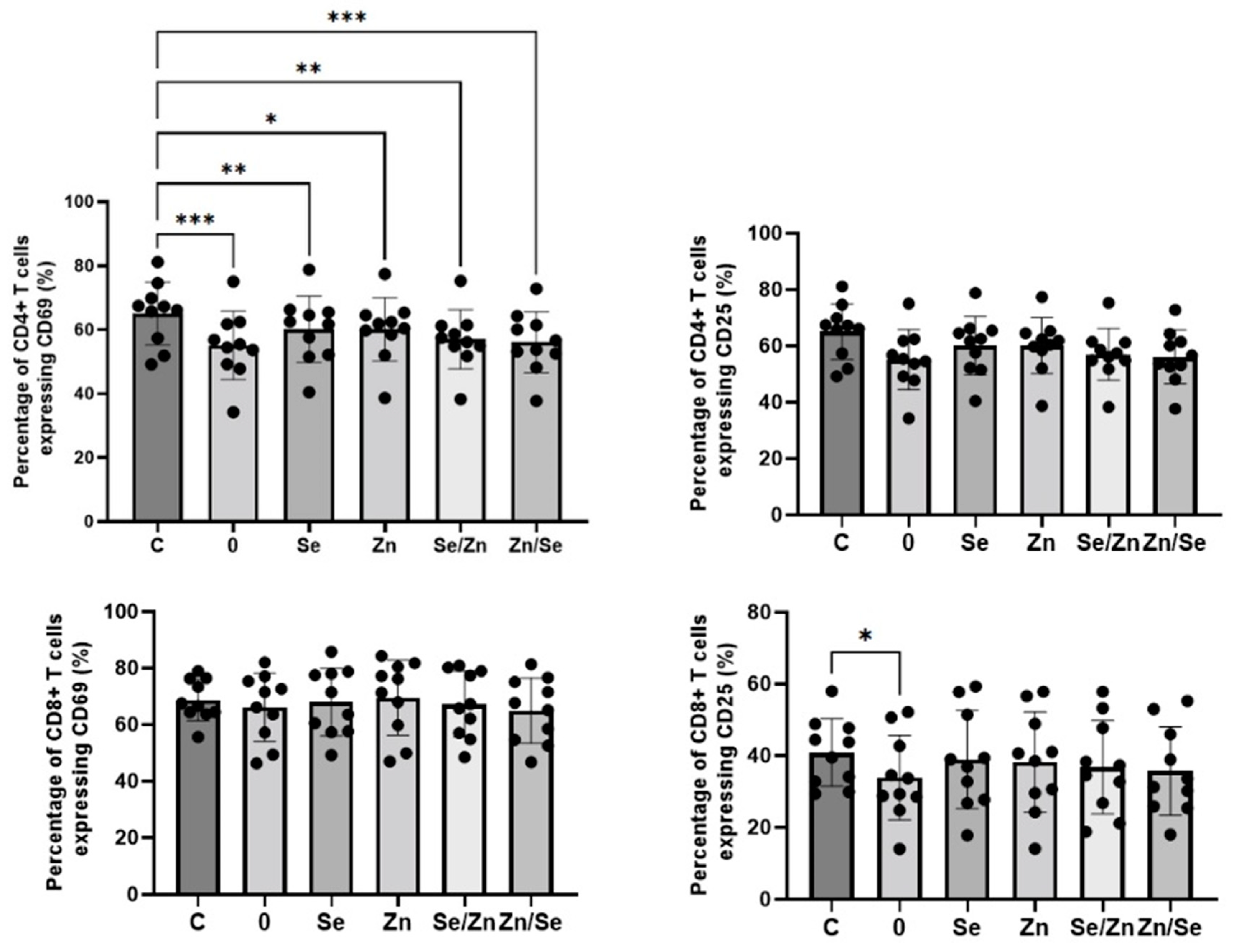
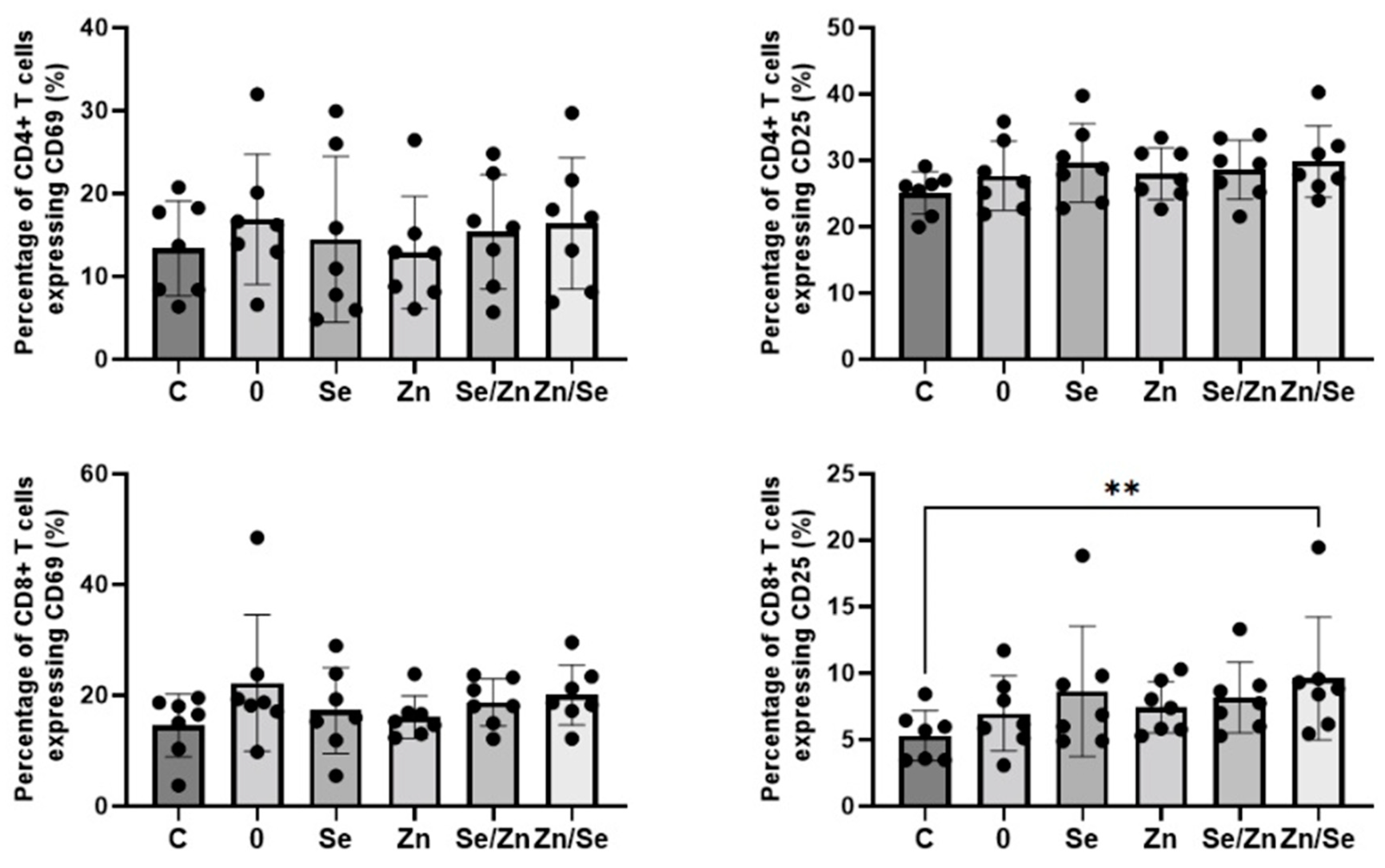
3.3. Immunological Activity of Mycelial Extracts
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jayawardena, R.; Sooriyaarachchi, P.; Chourdakis, M.; Jeewandara, C.; Ranasinghe, P. Enhancing Immunity in Viral Infections, with Special Emphasis on COVID-19: A Review. Diabetes Metab. Syndr. Clin. Res. Rev. 2020, 14, 367–382. [Google Scholar] [CrossRef] [PubMed]
- Baum, M.K.; Miguez-Burbano, M.J.; Campa, A.; Shor-Posner, G. Selenium and Interleukins in Persons Infected with Human Immunodeficiency Virus Type 1. J. Infect. Dis. 2000, 182, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Taylor, E.; Cox, A.; Zhao, L.; Ruzicka, J.; Bhat, A.; Zhang, W.; Nadimpalli, R.; Dean, R. Nutrition, HIV, and Drug Abuse: The Molecular Basis of a Unique Role for Selenium. J. Acquir. Immune Defic. Syndr. 2000, 25, S53–S61. [Google Scholar] [CrossRef] [PubMed]
- Diamond, A.M.; Hu, Y.J.; Mansur, D.B. Glutathione Peroxidase and Viral Replication: Implications for Viral Evolution and Chemoprevention. Biofactors 2001, 14, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Gómez, R.M.; Berría, M.I.; Levander, O.A. Host Selenium Status Selectively Influences Susceptibility to Experimental Viral Myocarditis. Biol. Trace Elem. Res. 2001, 80, 23–31. [Google Scholar] [CrossRef]
- Kalkan, A.; Bulut, V.; Avci, S.; Celik, I.; Bingol, N.K. Trace Elements in Viral Hepatitis. J. Trace Elem. Med. Biol. 2002, 16, 227–230. [Google Scholar] [CrossRef]
- Haraguchi, Y.; Sakurai, H.; Hussain, S.; Anner, B.M.; Hoshino, H. Inhibition of HIV-1 Infection by Zinc Group Metal Compounds. Antivir. Res. 1999, 43, 123–133. [Google Scholar] [CrossRef]
- Yuasa, K.; Naganuma, A.; Sato, K.; Ikeda, M.; Kato, N.; Takagi, H.; Mori, M. Zinc Is a Negative Regulator of Hepatitis C Virus RNA Replication. Liver Int. 2006, 26, 1111–1118. [Google Scholar] [CrossRef]
- Soriano-Garcia, M. Organoselenium Compounds as Potential Therapeutic and Chemopreventive Agents: A Review. Curr. Med. Chem. 2004, 11, 1657–1669. [Google Scholar] [CrossRef]
- Takahashi, K.; Suzuki, N.; Ogra, Y. Bioavailability Comparison of Nine Bioselenocompounds In Vitro and In Vivo. Int. J. Mol. Sci. 2017, 18, 506. [Google Scholar] [CrossRef]
- Schrauzer, G.N.; Surai, P.F. Selenium in Human and Animal Nutrition: Resolved and Unresolved Issues. A Partly Historical Treatise in Commemoration of the Fiftieth Anniversary of the Discovery of the Biological Essentiality of Selenium, Dedicated to the Memory of Klaus Schwarz (1914–1978) on the occasion of the thirtieth anniversary of his death. Crit. Rev. Biotechnol. 2009, 29, 2–9. [Google Scholar] [PubMed]
- Daragó, A.; Sapota, A.; Nasiadek, M.; Klimczak, M.; Kilanowicz, A. The Effect of Zinc and Selenium Supplementation Mode on Their Bioavailability in the Rat Prostate. Should Administration Be Joint or Separate? Nutrients 2016, 8, 601. [Google Scholar] [CrossRef] [PubMed]
- Federico, A.; Iodice, P.; Federico, P.; Del Rio, A.; Mellone, M.C.; Catalano, G.; Federico, P. Effects of Selenium and Zinc Supplementation on Nutritional Status in Patients with Cancer of Digestive Tract. Eur. J. Clin. Nutr. 2001, 55, 293–297. [Google Scholar] [CrossRef]
- Chang, S.-T.; Wasser, S.P. The Role of Culinary-Medicinal Mushrooms on Human Welfare with a Pyramid Model for Human Health. Int. J. Med. Mushrooms 2012, 14, 95–134. [Google Scholar] [CrossRef]
- Mizuno, M.; Nishitani, Y. Immunomodulating Compounds in Basidiomycetes. J. Clin. Biochem. Nutr. 2013, 52, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Wasser, S.P. Current Findings, Future Trends, and Unsolved Problems in Studies of Medicinal Mushrooms. Appl. Microbiol. Biotechnol. 2011, 89, 1323–1332. [Google Scholar] [CrossRef] [PubMed]
- Giavasis, I. Production of Microbial Polysaccharides for Use in Food. In Microbial Production of Food Ingredients, Enzymes and Nutraceuticals; Elsevier: Amsterdam, The Netherlands, 2013; pp. 413–468. [Google Scholar]
- Friedman, M. Mushroom Polysaccharides: Chemistry and Antiobesity, Antidiabetes, Anticancer, and Antibiotic Properties in Cells, Rodents, and Humans. Foods 2016, 5, 80. [Google Scholar] [CrossRef]
- Pethkar, A.V.; Paknikar, K.M. Recovery of Gold from Solutions Using Cladosporium Cladosporioides Biomass Beads. J. Biotechnol. 1998, 63, 121–136. [Google Scholar] [CrossRef]
- Borovička, J.; Řanda, Z. Distribution of Iron, Cobalt, Zinc and Selenium in Macrofungi. Mycol. Prog. 2007, 6, 249–259. [Google Scholar] [CrossRef]
- Da Silva, M.C.S.; Naozuka, J.; Da Luz, J.M.R.; De Assunão, L.S.; Oliveira, P.V.; Vanetti, M.C.D.; Bazzolli, D.M.S.; Kasuya, M.C.M. Enrichment of Pleurotus Ostreatus Mushrooms with Selenium in Coffee Husks. Food Chem. 2012, 131, 558–563. [Google Scholar] [CrossRef]
- Reczyński, W.; Muszyńska, B.; Opoka, W.; Smalec, A.; Sułkowska-Ziaja, K.; Malec, M. Comparative Study of Metals Accumulation in Cultured In Vitro Mycelium and Naturally Grown Fruiting Bodies of Boletus Badius and Cantharellus Cibarius. Biol. Trace Elem. Res. 2013, 153, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Savic, M.; Andjelkovic, I.; Duvnjak, D.; Matijasevic, D.; Avramovic, A.; Pesic-Mikulec, D.; Niksic, M. The Fungistatic Activity of Organic Selenium and Its Application to the Production of Cultivated Mushrooms Agaricus bisporus and Pleurotus spp. Arch. Biol. Sci. 2012, 64, 1455–1463. [Google Scholar] [CrossRef]
- Turlo, J.; Gutkowska, B.; Herold, F.; Blazewicz, A.; Kocjan, R. Relationship between Sodium Selenite Concentration in Culture Media and Heavy Metal Uptake by Lentinula edodes (Berk.) in Mycelial Cultures. Fresenius Environ. Bull. 2009, 18, 1035–1038. [Google Scholar]
- Turło, J.; Gutkowska, B.; Herold, F.; Klimaszewska, M.; Suchocki, P. Optimization of Selenium-Enriched Mycelium of Lentinula edodes (Berk.) Pegler as a Food Supplement. Food Biotechnol. 2010, 24, 180–196. [Google Scholar] [CrossRef]
- Malinowska, E.; Klimaszewska, M.; Strączek, T.; Schneider, K.; Kapusta, C.; Podsadni, P.; Łapienis, G.; Dawidowski, M.; Kleps, J.; Górska, S.; et al. Selenized Polysaccharides—Biosynthesis and Structural Analysis. Carbohydr. Polym. 2018, 198, 407–417. [Google Scholar] [CrossRef]
- Kaleta, B.; Górski, A.; Zagożdżon, R.; Cieślak, M.; Kaźmierczak-Barańska, J.; Nawrot, B.; Klimaszewska, M.; Malinowska, E.; Górska, S.; Turło, J. Selenium-Containing Polysaccharides from Lentinula edodes—Biological Activity. Carbohydr. Polym. 2019, 223, 115078. [Google Scholar] [CrossRef]
- Mihova, S.; Godjevargova, T. Biosorption of Heavy Metals from Aqueous Solutions. J. Int. Res. Pub 2001, 1, 1311. [Google Scholar]
- Yan, G.; Viraraghavan, T. Heavy-Metal Removal from Aqueous Solution by Fungus Mucor Rouxii. Water Res. 2003, 37, 4486–4496. [Google Scholar] [CrossRef]
- Eide, D.J. Homeostatic and Adaptive Responses to Zinc Deficiency in Saccharomyces cerevisiae. J. Biol. Chem. 2009, 284, 18565–18569. [Google Scholar] [CrossRef]
- Staats, C.C.; Kmetzsch, L.; Schrank, A.; Vainstein, M.H. Fungal Zinc Metabolism and Its Connections to Virulence. Front. Cell. Infect. Microbiol. 2013, 3, 65. [Google Scholar] [CrossRef]
- Gharieb, M.M. The Kinetics of 75[Se]-Selenite Uptake by Saccharomyces cerevisiae and the Vacuolization Response to High Concentrations. Mycol. Res. 2004, 108, 1415–1422. [Google Scholar] [CrossRef] [PubMed]
- Lazard, M.; Blanquet, S.; Fisicaro, P.; Labarraque, G.; Plateau, P. Uptake of Selenite by Saccharomyces cerevisiae Involves the High and Low Affinity Orthophosphate Transporters. J. Biol. Chem. 2010, 285, 32029–32037. [Google Scholar] [CrossRef] [PubMed]
- McDermott, J.R.; Rosen, B.P.; Liu, Z. Jen1p: A High Affinity Selenite Transporter in Yeast. Mol. Biol. Cell 2010, 21, 3934–3941. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, B.; Zhang, D.; Wu, Y. Selenium Uptake, Tolerance and Reduction in Flammulina Velutipes Supplied with Selenite. PeerJ 2016, 4, e1993. [Google Scholar] [CrossRef]
- Turło, J.; Gutkowska, B.; Kałucka, M.; Bujak, M. Accumulation of Zinc by the Lentinus edodes (Berk.) Mycelium Cultivated in Submerged Culture. Acta Pol. Pharm.—Drug Res. 2007, 64, 45–51. [Google Scholar]
- Clever, H.L.; Derrick, M.E.; Johnson, S.A. The Solubility of Some Sparingly Soluble Salts of Zinc and Cadmium in Water and in Aqueous Electrolyte Solutions. J. Phys. Chem. Ref. Data 1992, 21, 941–1004. [Google Scholar] [CrossRef]
- Banks, W.H. The Dissociation of the Selenates of Zinc and Cadmium in Water. J. Chem. Soc. Resumed 1934, 1010–1012. [Google Scholar] [CrossRef]
- Moriya, H.; Sekine, T. A Solvent-Extraction Study of Zinc (II) Complexes with Several Divalent Anions of Carboxylic and Inorganic Acids. Bull. Chem. Soc. Jpn. 1974, 47, 747–748. [Google Scholar] [CrossRef]
- Somer, G.; Yilmaz, Ü.T. Interference between Selenium and Some Trace Elements during Polarographic Studies and Its Elimination. Talanta 2005, 65, 598–602. [Google Scholar] [CrossRef]
- Feroci, G.; Badiello, R.; Fini, A. Interactions between Different Selenium Compounds and Zinc, Cadmium and Mercury. J. Trace Elem. Med. Biol. 2005, 18, 227–234. [Google Scholar] [CrossRef]
- Turło, J.; Gutkowska, B.; Herold, F.; Luczak, I. Investigation of the Kinetics of Selenium Accumulation by Lentinula edodes (Berk.) Mycelial Culture by Use of Reversed-Phase High-Performance Liquid Chromatography with Fluorimetric Detection. Acta Chromatogr. 2009, 21, 1–11. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Turło, J.; Gutkowska, B.; Malinowska, E. Relationship between the Selenium, Selenomethionine, and Selenocysteine Content of Submerged Cultivated Mycelium of Lentinula edodes (Berk.). Acta Chromatogr. 2007, 18, 36–48. [Google Scholar]
- Klimaszewska, M.; Górska, S.; Dawidowski, M.; Podsadni, P.; Turło, J. Biosynthesis of Se-Methyl-Seleno-l-Cysteine in Basidiomycetes Fungus Lentinula edodes (Berk.) Pegler. Springerplus 2016, 5, 733. [Google Scholar] [CrossRef]
- Rouse, B.T.; Sehrawat, S. Immunity and Immunopathology to Viruses: What Decides the Outcome? Nat. Rev. Immunol. 2010, 10, 514–526. [Google Scholar] [CrossRef]
- Shipkova, M.; Wieland, E. Surface Markers of Lymphocyte Activation and Markers of Cell Proliferation. Clin. Chim. Acta 2012, 413, 1338–1349. [Google Scholar] [CrossRef]
- Cibrián, D.; Sánchez-Madrid, F. CD69: From Activation Marker to Metabolic Gatekeeper. Eur. J. Immunol. 2017, 47, 946–953. [Google Scholar] [CrossRef]
- Shatrova, A.N.; Mityushova, E.V.; Vassilieva, I.O.; Aksenov, N.D.; Zenin, V.V.; Nikolsky, N.N.; Marakhova, I.I. Time-Dependent Regulation of IL-2R α-Chain (CD25) Expression by TCR Signal Strength and IL-2-Induced STAT5 Signaling in Activated Human Blood T Lymphocytes. PLoS ONE 2016, 11, e0167215. [Google Scholar] [CrossRef]
- Jubel, J.M.; Barbati, Z.R.; Burger, C.; Wirtz, D.C.; Schildberg, F.A. The Role of PD-1 in Acute and Chronic Infection. Front. Immunol. 2020, 11, 487. [Google Scholar] [CrossRef]
- Zou, C.; Du, Y.; Rashid, A.; Ram, H.; Savasli, E.; Pieterse, P.J.; Ortiz-Monasterio, I.; Yazici, A.; Kaur, C.; Mahmood, K.; et al. Simultaneous Biofortification of Wheat with Zinc, Iodine, Selenium, and Iron through Foliar Treatment of a Micronutrient Cocktail in Six Countries. J. Agric. Food Chem. 2019, 67, 8096–8106. [Google Scholar] [CrossRef]
- Roszczyk, A.; Zych, M.; Zielniok, K.; Krata, N.; Turło, J.; Klimaszewska, M.; Zagożdżon, R.; Kaleta, B. The Effect of Novel Selenopolysaccharide Isolated from Lentinula edodes Mycelium on Human T Lymphocytes Activation, Proliferation, and Cytokines Synthesis. Biomolecules 2022, 12, 1900. [Google Scholar] [CrossRef] [PubMed]
- Bayanati, M.; Al-Tawaha, A.R.; Al-Taey, D.; Al-Ghzawi, A.L.; Abu-Zaitoon, Y.M.; Shawaqfeh, S.; Al-Zoubi, O.; Al-Ramamneh, E.A.D.; Alomari, L.; Al-Tawaha, A.R.; et al. Interaction between Zinc and Selenium Bio-Fortification and Toxic Metals (Loid) Accumulation in Food Crops. Front. Plant Sci. 2022, 13, 1001992. [Google Scholar] [CrossRef] [PubMed]
- Koszinowski, U.H.; Reddehase, M.J.; Jonjic, S. The Role of CD4 and CD8 T Cells in Viral Infections. Curr. Opin. Immunol. 1991, 3, 471–475. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Mentzer, A.J.; Liu, G.; Yao, X.; Yin, Z.; Dong, D.; Dejnirattisai, W.; Rostron, T.; Supasa, P.; Liu, C.; et al. Broad and Strong Memory CD4+ and CD8+ T Cells Induced by SARS-CoV-2 in UK Convalescent Individuals Following COVID-19. Nat. Immunol. 2020, 21, 1336–1345. [Google Scholar] [CrossRef] [PubMed]
- Trickett, A.; Kwan, Y.L. T Cell Stimulation and Expansion Using Anti-CD3/CD28 Beads. J. Immunol. Methods 2003, 275, 251–255. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Series | Precursor | Concentration of Precursor [mM] | ||||
|---|---|---|---|---|---|---|
| I | SeO32− | 0.1 | 0.2 | 0.4 | 0.6 | 0.8 |
| Zn2+ | 0 | 0 | 0 | 0 | 0 | |
| II | SeO32− | 0.1 | 0.2 | 0.4 | 0.6 | 0.8 |
| Zn2+ | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | |
| III | Zn2+ | 0.1 | 0.2 | 0.4 | 0.6 | 0.8 |
| SeO32− | 0 | 0 | 0 | 0 | 0 | |
| IV | Zn2+ | 0.1 | 0.2 | 0.4 | 0.6 | 0.8 |
| SeO32− | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | |
| Parameter | Value |
|---|---|
| RF Power | 1550 W |
| Plasma argon flow rate | 15 L min−1 |
| Auxiliary argon flow rate | 0.9 L min−1 |
| Nebulizer argon flow rate | 1.03 L min−1 |
| Cell gas flow rate | 4.3 mL min−1 |
| Dwell time | 0.3 s |
| Sweeps | 100 |
| Number of readings per replicate | 3 |
| Conditions | Oxide 156/140 and doubly charged 70/140 < 2% |
| Sample Acronym | Molar Ratio of Medium Supplements | Concentration of Se and Zn in Mycelium | Concentration of Se, Zn, and Saccharides in Mycelial Water Extracts | ||||
|---|---|---|---|---|---|---|---|
| (mM) | (ug/g) | (ug/g) | (ug/g) | (mg/g) | |||
| SeO32− | Zn (II) | Se | Zn | Se | Zn | Saccharides | |
| 0 | 0 | 0 | 2.52 (0.17) | 298.10 (29.02) | 0.39 (0.06) | 92.65 (0.91) | 196.40 (6.91) |
| Se | 0.8 | 0 | 1553.91 (102.1) | 181.92 (19.46) | 203.60 (5.18) | 134.81 (4.42) | 139.64 (4.31) |
| Se/Zn | 0.8 | 0.2 | 1395.89 (16.86) | 591.25 (18.41) | 199.39 (7.99) | 152.04 (6.07) | 146.60 (2.90) |
| Zn/Se | 0.2 | 0.8 | 44.59 (6.72) | 874.94 (194.16) | 26.67 (0.56) | 212.42 (4.42) | 142.16 (9.72) |
| Zn | 0 | 0.8 | 3.01 (0.65) | 2370.32 (235.49) | 7.07 (0.09) | 393.15 (4.46) | 188.53 (4.64) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kałucka, M.; Roszczyk, A.; Klimaszewska, M.; Kaleta, B.; Drelich, E.; Błażewicz, A.; Górska-Jakubowska, S.; Malinowska, E.; Król, M.; Prus, A.M.; et al. Optimization of Se- and Zn-Enriched Mycelium of Lentinula edodes (Berk.) Pegler as a Dietary Supplement with Immunostimulatory Activity. Nutrients 2023, 15, 4015. https://doi.org/10.3390/nu15184015
Kałucka M, Roszczyk A, Klimaszewska M, Kaleta B, Drelich E, Błażewicz A, Górska-Jakubowska S, Malinowska E, Król M, Prus AM, et al. Optimization of Se- and Zn-Enriched Mycelium of Lentinula edodes (Berk.) Pegler as a Dietary Supplement with Immunostimulatory Activity. Nutrients. 2023; 15(18):4015. https://doi.org/10.3390/nu15184015
Chicago/Turabian StyleKałucka, Małgorzata, Aleksander Roszczyk, Marzenna Klimaszewska, Beata Kaleta, Ewelina Drelich, Anna Błażewicz, Sandra Górska-Jakubowska, Eliza Malinowska, Marek Król, Aleksandra Maria Prus, and et al. 2023. "Optimization of Se- and Zn-Enriched Mycelium of Lentinula edodes (Berk.) Pegler as a Dietary Supplement with Immunostimulatory Activity" Nutrients 15, no. 18: 4015. https://doi.org/10.3390/nu15184015
APA StyleKałucka, M., Roszczyk, A., Klimaszewska, M., Kaleta, B., Drelich, E., Błażewicz, A., Górska-Jakubowska, S., Malinowska, E., Król, M., Prus, A. M., Trześniowska, K., Wołczyńska, A., Dorożyński, P., Zagożdżon, R., & Turło, J. (2023). Optimization of Se- and Zn-Enriched Mycelium of Lentinula edodes (Berk.) Pegler as a Dietary Supplement with Immunostimulatory Activity. Nutrients, 15(18), 4015. https://doi.org/10.3390/nu15184015