
Antioxidative and Anti-Inflammatory Activities of Rosebud Extracts of Newly Crossbred Roses
 ,
, 
Abstract

1. Introduction
2. Materials and Methods
2.1. Extraction of Rosebuds
2.2. Analysis of Antioxidant Ingredients
2.3. Measurement of Antioxidative Activities
2.3.1. Analysis of ABTS-Scavenging Activity
2.3.2. Analysis of DPPH-Scavenging Activity
2.3.3. Analysis of Correlation between Antioxidant Contents and Antioxidative Activities
2.4. Measurement of Anti-Inflammatory Activities
2.4.1. Analysis of NO-Inhibitory Activity
2.4.2. Analysis of Correlation between Antioxidant Contents and NO-Inhibitory Activities
2.5. Measurement of In Vitro Anti-Inflammatory Activity of PVRE
2.5.1. MTT Assay for Cytotoxicity
2.5.2. qPCR Analysis of mRNA Expressions of iNOS and COX-2
2.5.3. Chemical Analysis of NO and PGE2
2.6. Measurement of In Vivo Anti-Inflammatory Activity of PVRE
2.6.1. Animals
2.6.2. Experimental Design Using an Air-Pouch Inflammation Model
2.6.3. Analysis of Exudate Volume and Inflammation Cells
2.6.4. ELISA Analysis of Inflammatory Cytokines
2.6.5. Chemical Analysis of NO and PGE2
2.6.6. ImageJ Analysis of Dermal Blood Vessel Area
2.6.7. Microscopic Examination of Tissue Inflammation
2.7. Statistical Analysis
3. Results
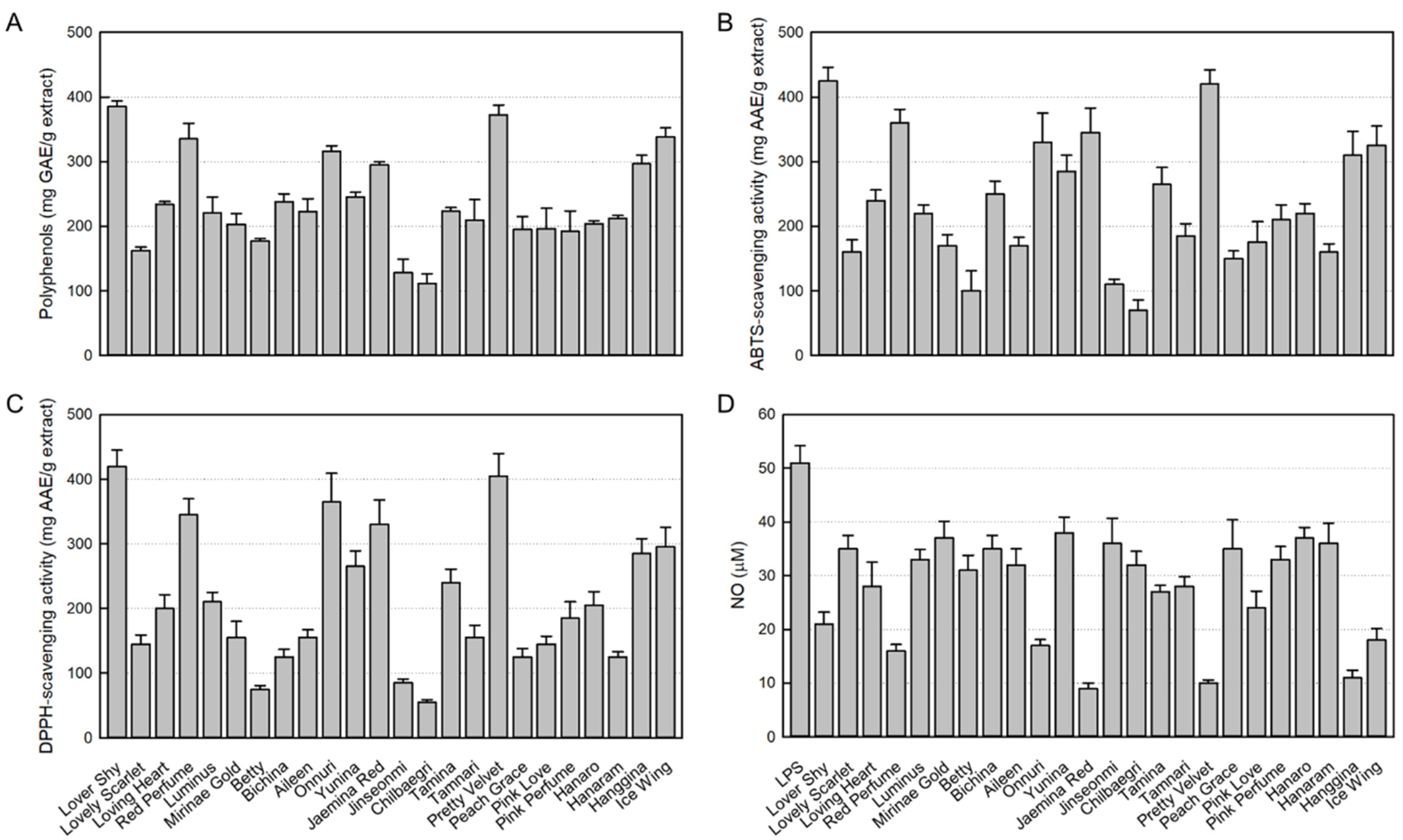
3.1. Antioxidant Contents in 24 Rosebud Extracts
3.2. Antioxidant Activities of 24 Rosebud Extracts
3.3. Inhibitory Activities of 24 Rosebud Extracts on NO Production
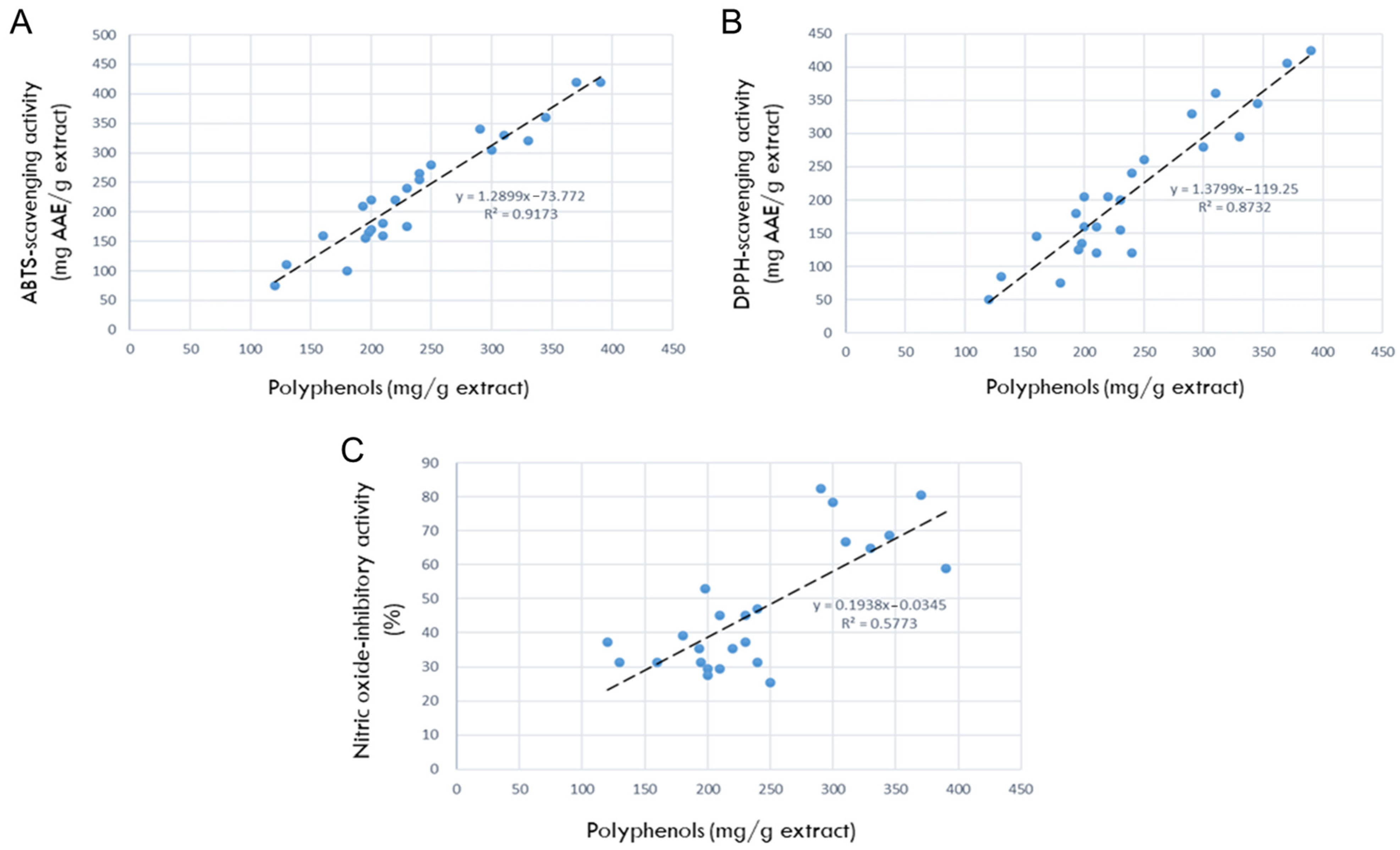
3.4. Correlation between Polyphenols and Antioxidative Activities
3.5. Correlation between Polyphenols and NO-Inhibitory Activities
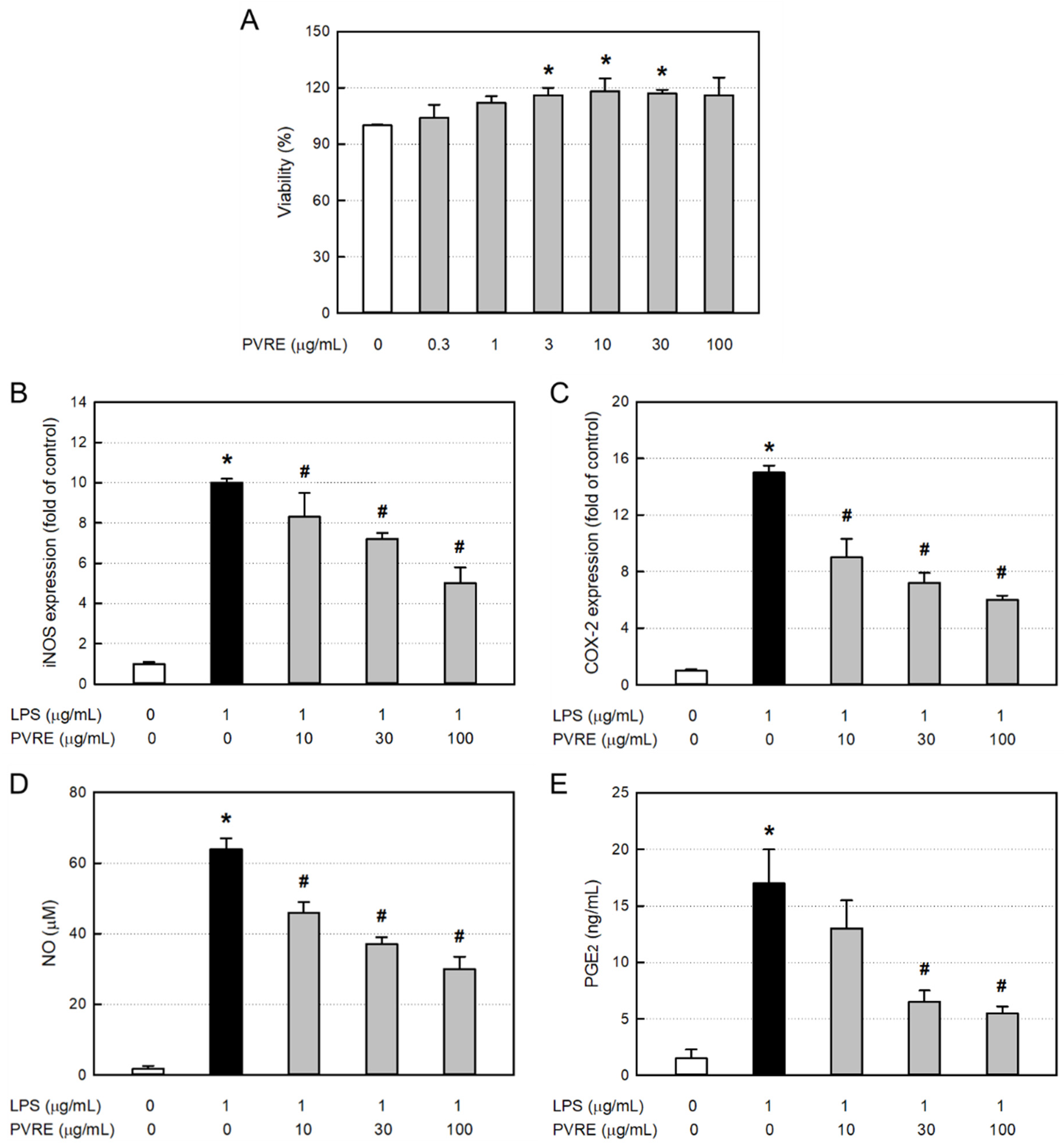
3.6. Cytotoxicity of PVRE in RAW 264.7 Cells
3.7. Effects on iNOS and COX-2 Expressions in RAW 264.7 Cells
3.8. Effects on NO and PGE2 Production in RAW 264.7 Cells
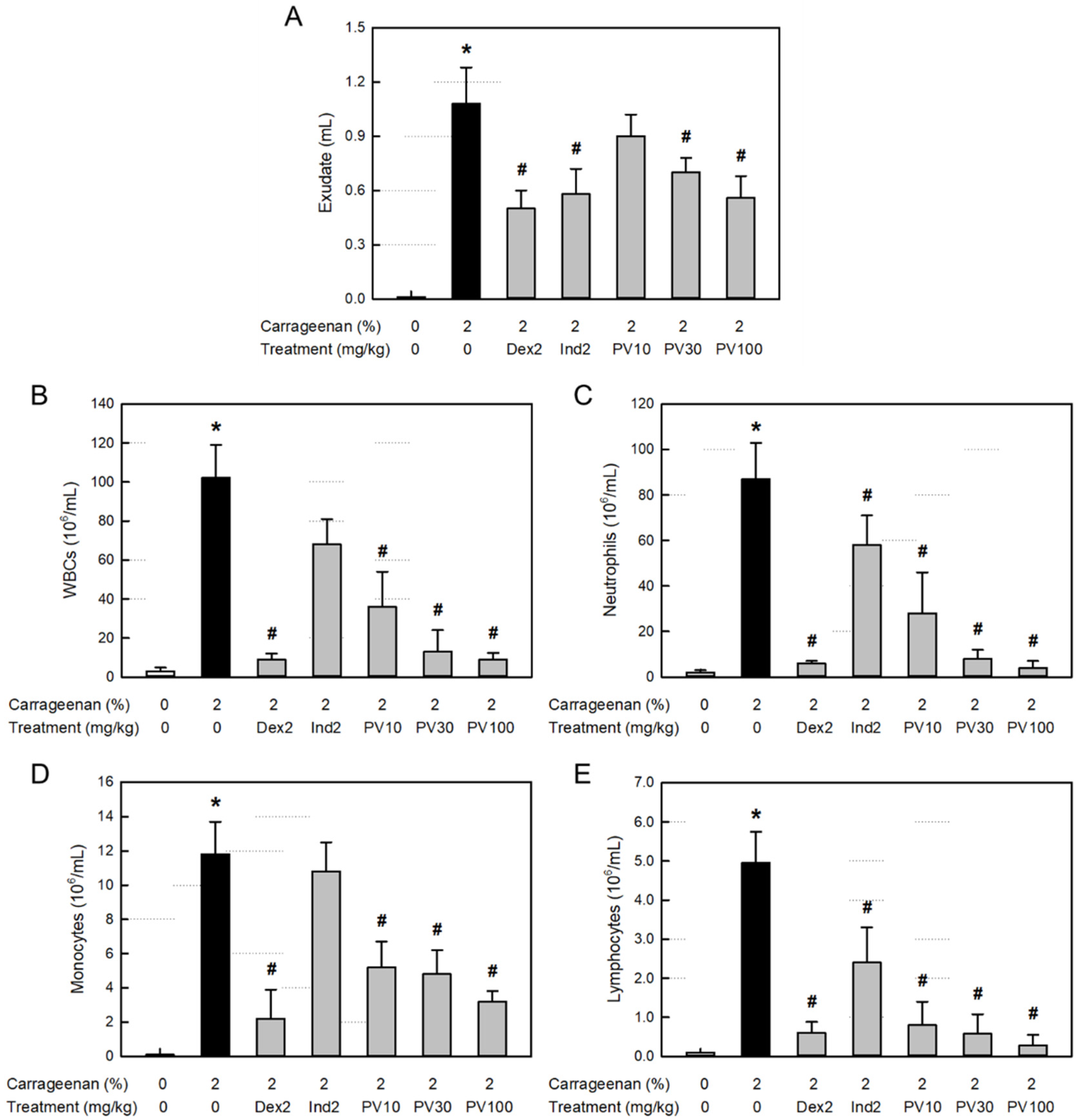
3.9. Effect on Exudation in Air-Pouch Inflammation
3.10. Effect on Inflammatory Cells in Exudate
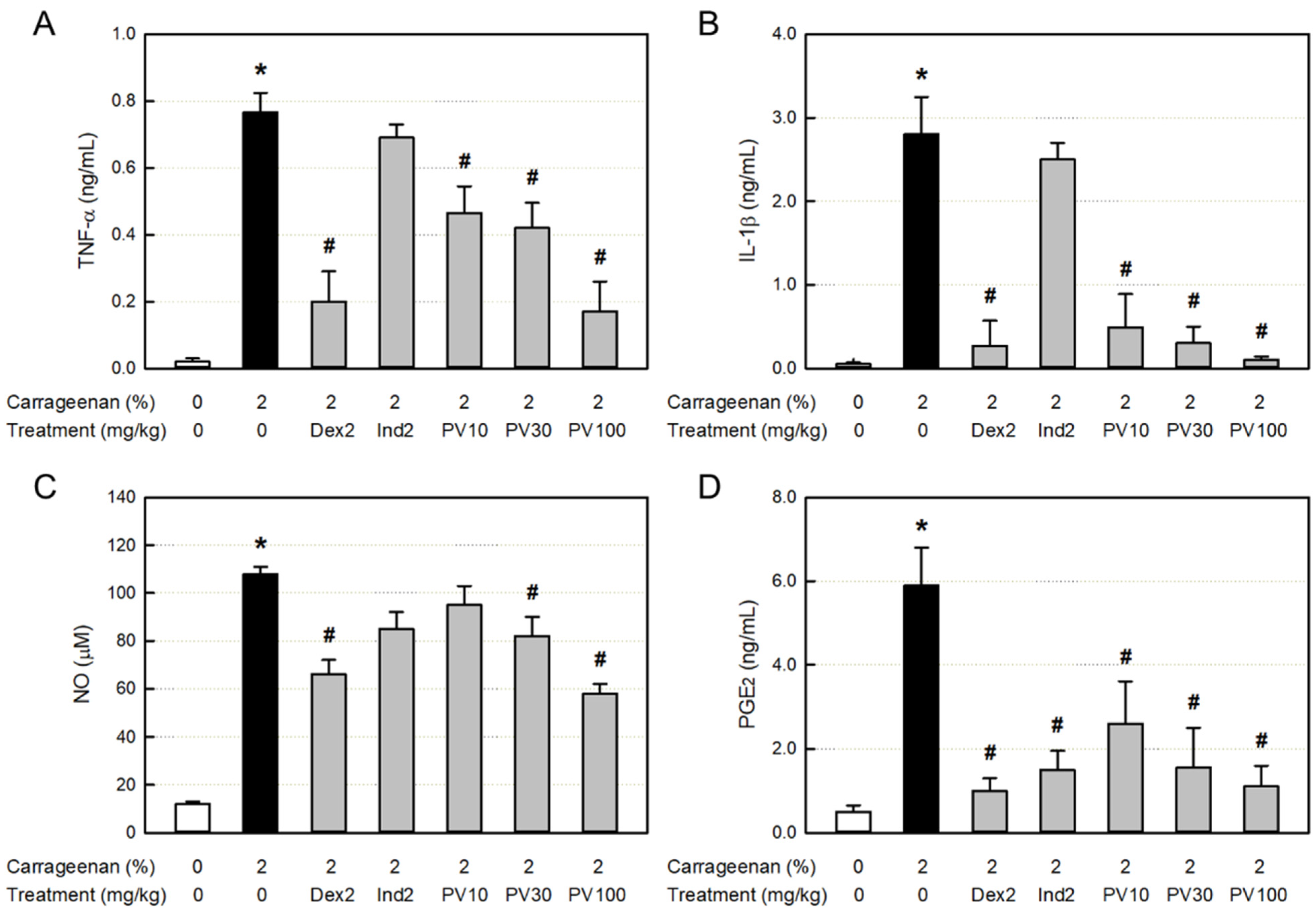
3.11. Effect on Inflammatory Cytokine Concentration in Exudate
3.12. Effects on NO and PGE2 Concentrations in Exudate
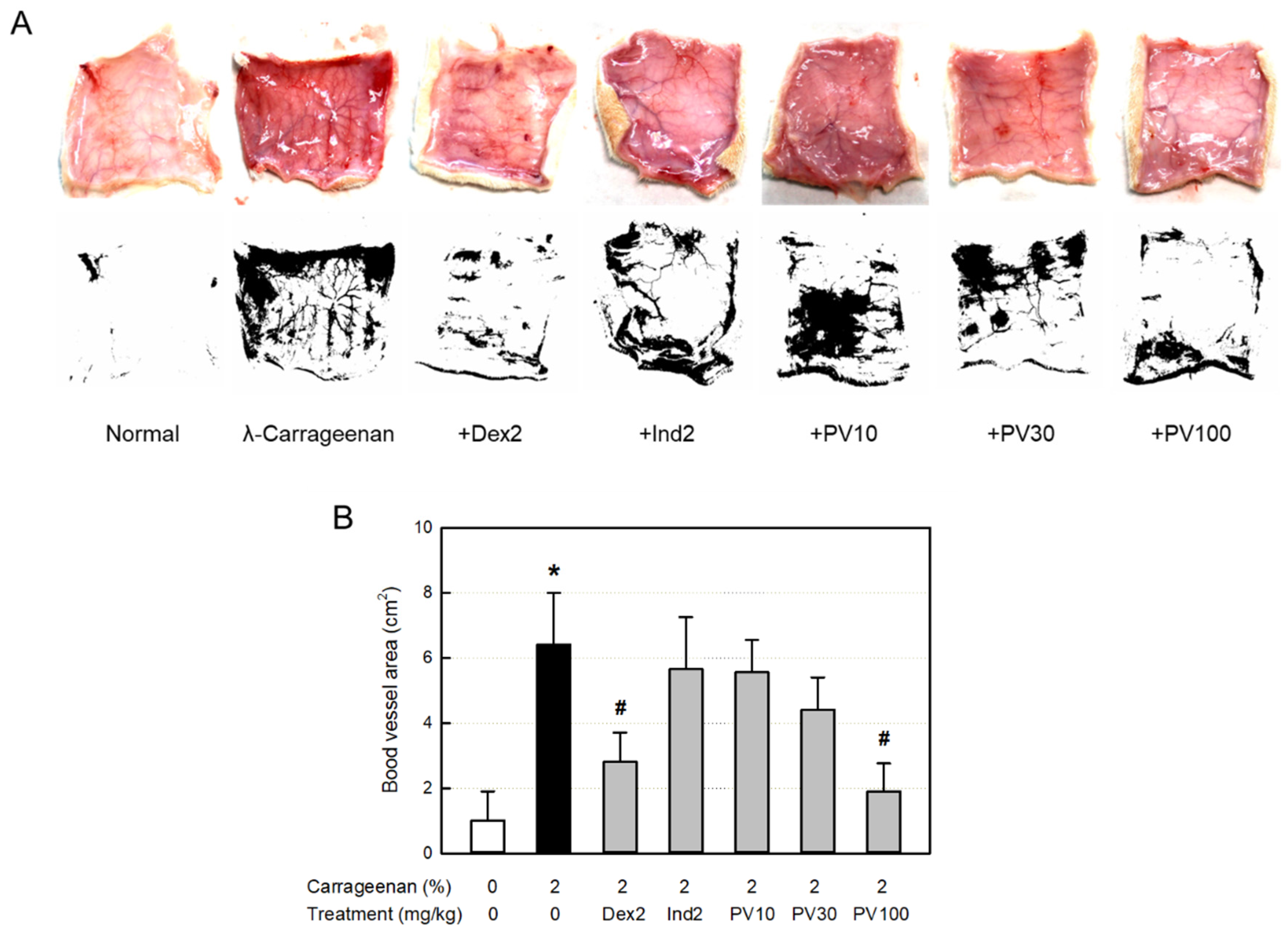
3.13. Effect on Blood Vessel Area
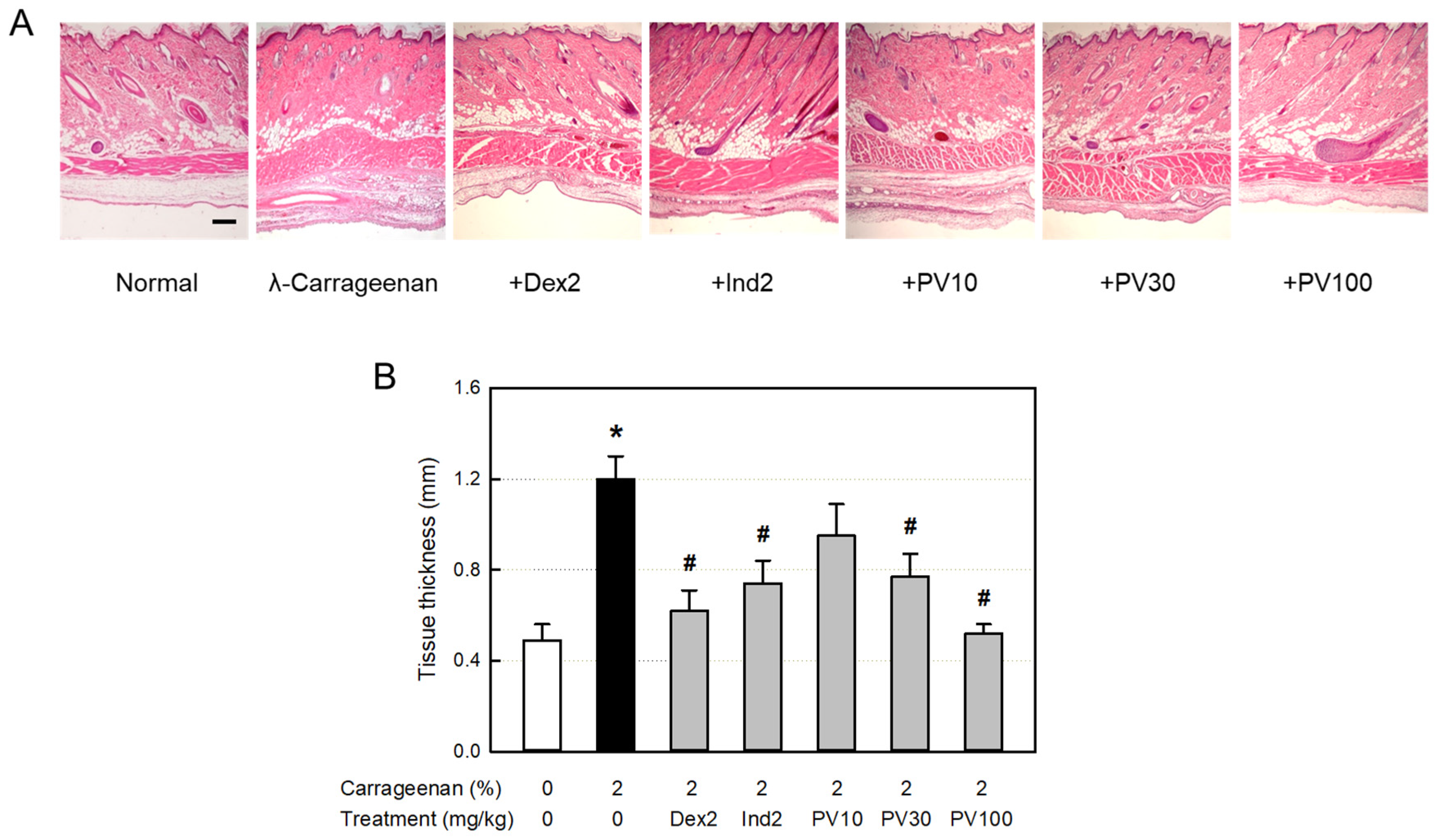
3.14. Microscopic Findings
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rani, V.; Deep, G.; Singh, R.K.; Palle, K.; Yadav, U.C.S. Oxidative stress and metabolic disorders: Pathogenesis and therapeutic strategies. Life Sci. 2016, 148, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.-H.; Dong, F.-X. The relevant targets of anti-oxidative stress: A review. J. Drug. Target. 2021, 29, 677–686. [Google Scholar] [CrossRef]
- Serhan, C.N.; Chiang, N.; Dalli, J. The resolution code of acute inflammation: Novel pro-resolving lipid mediators in resolution. Semin. Immunol. 2015, 27, 200–215. [Google Scholar] [CrossRef]
- Nonato, F.R.; Santana, D.G.; de Melo, F.M.; dos Santos, G.G.L.; Brustolim, D.; Camargo, E.A.; de Sousa, D.P.; Soares, M.B.P.; Villarreal, C.F. Anti-inflammatory properties of rose oxide. Int. Immunopharmacol. 2012, 14, 779–784. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.-T.; Wang, B.; Lv, L.-L.; Liu, B.-C. Extracellular vesicle-based Nanotherapeutics: Emerging frontiers in anti-inflammatory therapy. Theranostics 2020, 10, 8111–8129. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef]
- Schmid-Schönbein, G.W. Analysis of inflammation. Annu. Rev. Biomed. Eng. 2006, 8, 93–151. [Google Scholar] [CrossRef]
- Tsai, D.-S.; Huang, M.-H.; Tsai, J.-C.; Chang, Y.-S.; Chiu, Y.-J.; Lin, Y.-C.; Wu, L.-Y.; Peng, W.-H. Analgesic and Anti-Inflammatory Activities of Rosa taiwanensis Nakai in Mice. J. Med. Food 2015, 18, 592–600. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Su, W.W.; Wang, S.; Li, P.B. Naringgin inhibits chemokine production in an LPS-induced RAW 264.7 macrophage cell line. Mol. Med. Rep. 2012, 6, 1343–1350. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Ren, Y.; Lin, D.; Peng, S.; Zhong, B.; Ma, Z. The anti-inflammatory properties of Citrus wilsonii Tananka extract in LPS-induced RAW 264.7 and primary mouse bone marrow-derived dendritic cells. Molecules 2017, 22, 1213. [Google Scholar] [CrossRef]
- Shin, S.; Joo, S.S.; Jeon, J.H.; Park, D.; Jang, M.J.; Kim, T.O.; Kim, H.K.; Hwang, B.Y.; Kim, K.Y.; Kim, Y.B. Anti-inflammatory effects of a Houttuynia cordata supercritical extract. J. Vet. Sci. 2010, 11, 273–275. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A. Anti-inflammatory Agents: Present and Future. Cell 2010, 140, 935–950. [Google Scholar] [CrossRef] [PubMed]
- Thomson, A.W.; Horne, C.H. Toxicity of various carrageenans in the mouse. Br. J. Exp. Pathol. 1976, 57, 455–459. [Google Scholar]
- Edwards, J.C.W.; Sedgwick, A.D.; Willoughby, D.A. The formation of a structure with the features of synovial lining by subcutaneous injection of air: Anin vivo tissue culture system. J. Pathol. 1981, 134, 147–156. [Google Scholar] [CrossRef]
- Vandooren, J.; Berghmans, N.; Dillen, C.; Van Aelst, I.; Ronsse, I.; Israel, L.L.; Rosenberger, I.; Kreuter, J.; Lellouche, J.P.; Michaeli, S.; et al. Intradermal air pouch leukocytosis as an in vivo test for nanoparticles. Int. J. Nanomed. 2013, 8, 4745–4756. [Google Scholar] [CrossRef]
- Claxson, A.; Grootveld, M.; Chander, C.; Earl, J.; Haycock, P.; Mantle, M.; Williams, S.; Silwood, C.; Blake, D. Examination of the metabolic status of rat air pouch inflammatory exudate by high field proton NMR spectroscopy. Biochim. Biophys. Acta (BBA)—Mol. Basis Dis. 1999, 1454, 57–70. [Google Scholar] [CrossRef]
- Nijampurkar, B.; Qureshi, F.; Jain, N.; Banerjee, T.; Kumar, A.; Parmar, H.S. Anti-Inflammatory Role of Thyroid Hormones on Rat Air Pouch Model of Inflammation. Inflamm. Allergy-Drug. Targets 2015, 14, 117–124. [Google Scholar] [CrossRef]
- Eteraf-Oskouei, T.; Shafiee-Khamneh, A.; Heshmati-Afshar, F.; Delazar, A. Anti-inflammatory and anti-angiogenesis effect of bee pollen methanolic extract using air pouch model of inflammation. Res. Pharm. Sci. 2020, 15, 66–75. [Google Scholar] [CrossRef]
- Fehrenbacher, J.C.; McCarson, K.E. Models of Inflammation: Carrageenan Air Pouch. Curr. Protoc. 2021, 1, e183. [Google Scholar] [CrossRef]
- Kim, J.M.; Park, S.H. Risk and benefit of steroid therapy. J. Korean Soc. Intern. Med. 2009, 77, 298–303. [Google Scholar]
- Eteraf-Oskouei, T.; Mirak, S.M.; Najafi, M. Anti-Inflammatory and Anti-Angiogenesis Effects of Verapamil on Rat Air Pouch Inflammation Model. Adv. Pharm. Bull. 2017, 7, 585–591. [Google Scholar] [CrossRef]
- Lee, S.Y.; Cho, S.S.; Li, Y.C.; Bae, C.S.; Park, K.M.; Park, D.H. Anti-inflammatory effect of Curcuma longa and Allium hookeri co-treatment via NF-κB and COX-2 pathways. Sci. Rep. 2020, 10, 5718. [Google Scholar] [CrossRef]
- Boukhatem, M.N.; Kameli, A.; Ferhat, M.A.; Saidi, F.; Mekarnia, M. Rose geranium essential oil as a source of new and safe anti-inflammatory drugs. Libyan J. Med. 2013, 8, 22520. [Google Scholar] [CrossRef]
- Ahn, J.-H.; Park, Y.-L.; Song, A.-Y.; Kim, W.-G.; Je, C.-Y.; Jung, D.-H.; Kim, Y.-J.; Oh, J.; Cho, J.-Y.; Kim, D.-J.; et al. Water extract of Artemisia scoparia Waldst. & Kitam suppresses LPS-induced cytokine production and NLRP3 inflammasome activation in macrophages and alleviates carrageenan-induced acute inflammation in mice. J. Ethnopharmacol. 2021, 268, 113606. [Google Scholar] [CrossRef] [PubMed]
- Mileva, M.; Ilieva, Y.; Jovtchev, G.; Gateva, S.; Zaharieva, M.M.; Georgieva, A.; Dimitrova, L.; Dobreva, A.; Angelova, T.; Vilhelmova-Ilieva, N.; et al. Rose flowers—A delicate perfume or a natural healer? Biomolecules 2021, 11, 127. [Google Scholar] [CrossRef]
- Gorji, A. Pharmacological treatment of headache using traditional Persian medicine. Trends Pharmacol. Sci. 2003, 24, 331–334. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, N.; Serafini, M.; Colombi, B.; Del Rio, D.; Salvatore, S.; Bianchi, M.; Brighenti, F. Total antioxidant capacity of plant foods, beverages and oils consumed in Italy assessed by three different in vitro assays. J. Nutr. 2003, 133, 2812–2819. [Google Scholar] [CrossRef] [PubMed]
- Cho, E.K.; Son, J.Y.; Kang, K.O. Antioxidant activities of rose, camellia and cockscomb flower extracts. Food Serv. Indust J. 2015, 11, 21–33. [Google Scholar]
- Lee, H.R.; Lee, J.M.; Choi, N.S.; Lee, J.M. The anti-oxidative and anti-microbial ability of ethanol extracts from Rosa hybrida. Korean J. Food Sci. Technol. 2003, 35, 373–378. [Google Scholar]
- Park, D.; Jeon, J.H.; Kwon, S.-C.; Shin, S.; Jang, J.Y.; Jeong, H.S.; Lee, D.I.; Kim, Y.-B.; Joo, S.S. Antioxidative activities of white rose flower extract and pharmaceutical advantages of its hexane fraction via free radical scavenging effects. Biochem. Cell. Biol. 2009, 87, 943–952. [Google Scholar] [CrossRef]
- Seong, H.; Heo, J.; Lee, K.H.; Lee, Y.B.; Kim, Y.B.; Han, N.S. Enhancing the Antioxidant Activities of Wines by Addition of White Rose Extract. J. Microbiol. Biotechnol. 2017, 27, 1602–1608. [Google Scholar] [CrossRef] [PubMed]
- Konczak, I.; Zhang, W. Anthocyanins—more than nature’s colours. J. Biomed. Biotechnol. 2004, 2004, 239–240. [Google Scholar] [CrossRef]
- Zkan, G.; Sağdiç, O.; Baydar, N.G.; Baydar, H. Anti-oxidant and anti-bacterial activities of Rosa damascena flower extracts. Food Sci. Technol. Int. 2004, 10, 277–281. [Google Scholar]
- Park, D.; Shin, K.; Choi, Y.; Guo, H.; Cha, Y.; Kim, S.-H.; Han, N.S.; Joo, S.S.; Choi, J.K.; Lee, Y.B.; et al. Antimicrobial activities of ethanol and butanol fractions of white rose petal extract. Regul. Toxicol. Pharmacol. 2016, 76, 57–62. [Google Scholar] [CrossRef]
- Jeon, J.H.; Kwon, S.-C.; Park, D.; Shin, S.; Jeong, J.-H.; Park, S.-Y.; Hwang, S.-Y.; Kim, Y.-B.; Joo, S.S. Anti-allergic effects of white rose petal extract and anti-atopic properties of its hexane fraction. Arch. Pharmacal Res. 2009, 32, 823–830. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.-K.; Guo, H.; Choi, J.-K.; Jang, S.-K.; Shin, K.; Cha, Y.-S.; Choi, Y.; Seo, D.-W.; Lee, Y.-B.; Joo, S.-S.; et al. Extraction conditions of white rose petals for the inhibition of enzymes related to skin aging. Lab. Anim. Res. 2015, 31, 148–152. [Google Scholar] [CrossRef]
- Yang, G.; Park, D.; Lee, S.H.; Bae, D.-K.; Yang, Y.-H.; Kyung, J.; Kim, D.; Choi, E.-K.; Hong, J.T.; Jeong, H.-S.; et al. Neuroprotective effects of a butanol fraction of Rosa hybrida petals in a middle cerebral artery occlusion model. Biomol. Ther. 2013, 21, 454–461. [Google Scholar] [CrossRef] [PubMed]
- Yon, J.-M.; Kim, Y.-B.; Park, D. The ethanol fraction of white rose petal extract abrogates excitotoxicity-induced neuronal damage in vivo and in vitro through inhibition of oxidative stress and proinflammation. Nutrients 2018, 10, 1375. [Google Scholar] [CrossRef]
- Choi, S.Y.; Kim, M.Y.; Lee, Y.J.; Choi, E.-K.; Kim, Y.-B.; Lee, J.; Jeong, H.S. Antioxidant activities and functional components of some rose flower cultivars. J. Korean Soc. Food Sci. Nutr. 2019, 48, 494–500. [Google Scholar] [CrossRef]
- Dewanto, V.; Wu, X.; Liu, R.H. Processed sweet corn has higher antioxidant activity. J. Agric. Food Chem. 2002, 50, 4959–4964. [Google Scholar] [CrossRef]
- Choi, Y.; Lee, S.M.; Chun, J.; Lee, H.B.; Lee, J. Influence of heat treatment on the antioxidant activities and polyphenolic compounds of shiitake (Lentinus edodes) mushroom. Food Chem. 2006, 99, 381–387. [Google Scholar] [CrossRef]
- Hwang, I.G.; Woo, K.S.; Kim, T.M.; Kim, D.J.; Yang, M.H.; Jeong, H.S. Change of physicochemical characteristics of Korean pear (Pyrus pyrifolia Nakai) juice with heat treatment conditions. Korean J. Food Sci. Technol. 2006, 38, 334–342. [Google Scholar]
- Hwang, M.R.; Kim, H.E.; Park, D.K.; Heu, Y.C.; Lee, H.J.; Kang, N.J. Induction of oxidative stress and activation of antioxidant enzymes by infection of powdery mildew in cucurbita plants. J. Agric. Life Sci. 2013, 47, 75–81. [Google Scholar]
- Que, F.; Mao, L.; Zhu, C.; Xie, G. Anti-oxidant properties of Chinese yellow wine, its concentrate and volatiles. LWT-Food Sci. Technol. 2006, 39, 111–117. [Google Scholar] [CrossRef]
- Del Prado-Audelo, M.L.; Cortés, H.; Caballero-Florán, I.H.; González-Torres, M.; Escutia-Guadarrama, L.; Bernal-Chávez, S.A.; Giraldo-Gomez, D.M.; Magaña, J.J.; Leyva-Gómez, G. Therapeutic applications of terpenes on inflammatory diseases. Front. Pharmacol. 2021, 12, 704197. [Google Scholar] [CrossRef]
- Teixeira de Moraes Costa, M.M.; Penha de Oliveira, S.H.; Gomes-Filho, J.E. Mechanism of calcium hydroxide-induced neutrophil migration into air-pouch cavity. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endodontol. 2008, 105, 814–821. [Google Scholar] [CrossRef]
- Perez, D.A.; Vago, J.P.; Athayde, R.M.; Reis, A.C.; Teixeira, M.M.; Sousa, L.P.; Pinho, V. Switching off Key Signaling Survival Molecules to Switch on the Resolution of Inflammation. Mediat. Inflamm. 2014, 2014, 829851. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.M.; An, J.X. Cytokines, inflammation, and pain. Int. Anesth. Clin. 2009, 45, 27–37. [Google Scholar] [CrossRef]
- Winyard, P.G.; Willoughby, D. Inflammation protocols. In Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2003; p. 225. [Google Scholar]
- Wojdasiewicz, P.; Poniatowski, L.A.; Szukiewicz, D. The Role of Inflammatory and Anti-Inflammatory Cytokines in the Pathogenesis of Osteoarthritis. Mediat. Inflamm. 2014, 2014, 561459. [Google Scholar] [CrossRef]
- Cinelli, M.A.; Do, H.T.; Miley, G.P.; Silverman, R.B. Inducible nitric oxide synthase: Regulation, structure, and inhibition. Med. Res. Rev. 2019, 40, 158–189. [Google Scholar] [CrossRef]
- Tian, Y.; Zhou, S.; Takeda, R.; Okazaki, K.; Sekita, M.; Sakamoto, K. Anti-inflammatory activities of amber extract in lipopolysaccharide-induced RAW 264.7 macrophages. Biomed. Pharmacother. 2021, 141, 111854. [Google Scholar] [CrossRef] [PubMed]
- Agard, M.; Asakrah, S.; Morici, L.A. PGE2 suppression of innate immunity during mucosal bacterial infection. Front. Cell. Infect. Microbiol. 2013, 3, 45. [Google Scholar] [CrossRef]
- Shin, S.; Jeon, J.H.; Park, D.; Jang, J.Y.; Joo, S.S.; Hwang, B.Y.; Choe, S.Y.; Kim, Y.-B. Anti-inflammatory effects of an ethanol extract of Angelica gigas in a carrageenan-air pouch inflammation model. Exp. Anim. 2009, 58, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Goppelt-Struebe, M.; Wolter, D.; Resch, K. Glucocorticoids inhibit prostaglandin synthesis not only at the level of phospholipase A2 but also at the level of cyclo-oxygenase/PGE isomerase. Br. J. Pharmacol. 1989, 98, 1287–1295. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Sequence 5′-3′ | Temperature |
|---|---|---|
| iNOS | Forward: CAGGATCCAGTGGTCCAACC Reverse: CGTACCGGATGAGCTGTGAA | 60 °C 59 °C |
| COX-2 | Forward: GTACAAGCAGTGGCAAAGGC Reverse: ACGAGGTTTTTCCACCAGCA | 60 °C 60 °C |
| GAPDH | Forward: GACCTCATGGCCTACATGGC Reverse: GCCCCTCCTGTTATTATGGGG | 60 °C 59 °C |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.; Kim, I.-J.; Seong, H.-R.; Noh, C.H.; Park, S.; Kim, T.M.; Jeong, H.S.; Kim, K.Y.; Kim, S.T.; Yuk, H.-G.; et al. Antioxidative and Anti-Inflammatory Activities of Rosebud Extracts of Newly Crossbred Roses. Nutrients 2023, 15, 2376. https://doi.org/10.3390/nu15102376
Wang C, Kim I-J, Seong H-R, Noh CH, Park S, Kim TM, Jeong HS, Kim KY, Kim ST, Yuk H-G, et al. Antioxidative and Anti-Inflammatory Activities of Rosebud Extracts of Newly Crossbred Roses. Nutrients. 2023; 15(10):2376. https://doi.org/10.3390/nu15102376
Chicago/Turabian StyleWang, Cuicui, In-Jeong Kim, Hye-Rim Seong, Chan Ho Noh, Sangryong Park, Tae Myoung Kim, Heon Sang Jeong, Ka Young Kim, Seung Tae Kim, Hyun-Gyun Yuk, and et al. 2023. "Antioxidative and Anti-Inflammatory Activities of Rosebud Extracts of Newly Crossbred Roses" Nutrients 15, no. 10: 2376. https://doi.org/10.3390/nu15102376
APA StyleWang, C., Kim, I.-J., Seong, H.-R., Noh, C. H., Park, S., Kim, T. M., Jeong, H. S., Kim, K. Y., Kim, S. T., Yuk, H.-G., Kwon, S.-C., Choi, E.-K., & Kim, Y.-B. (2023). Antioxidative and Anti-Inflammatory Activities of Rosebud Extracts of Newly Crossbred Roses. Nutrients, 15(10), 2376. https://doi.org/10.3390/nu15102376