
Investigation of the Effect of Curcumin on Protein Targets in NAFLD Using Bioinformatic Analysis

 ,
,  ,
,  and
and 
Abstract
1. Introduction
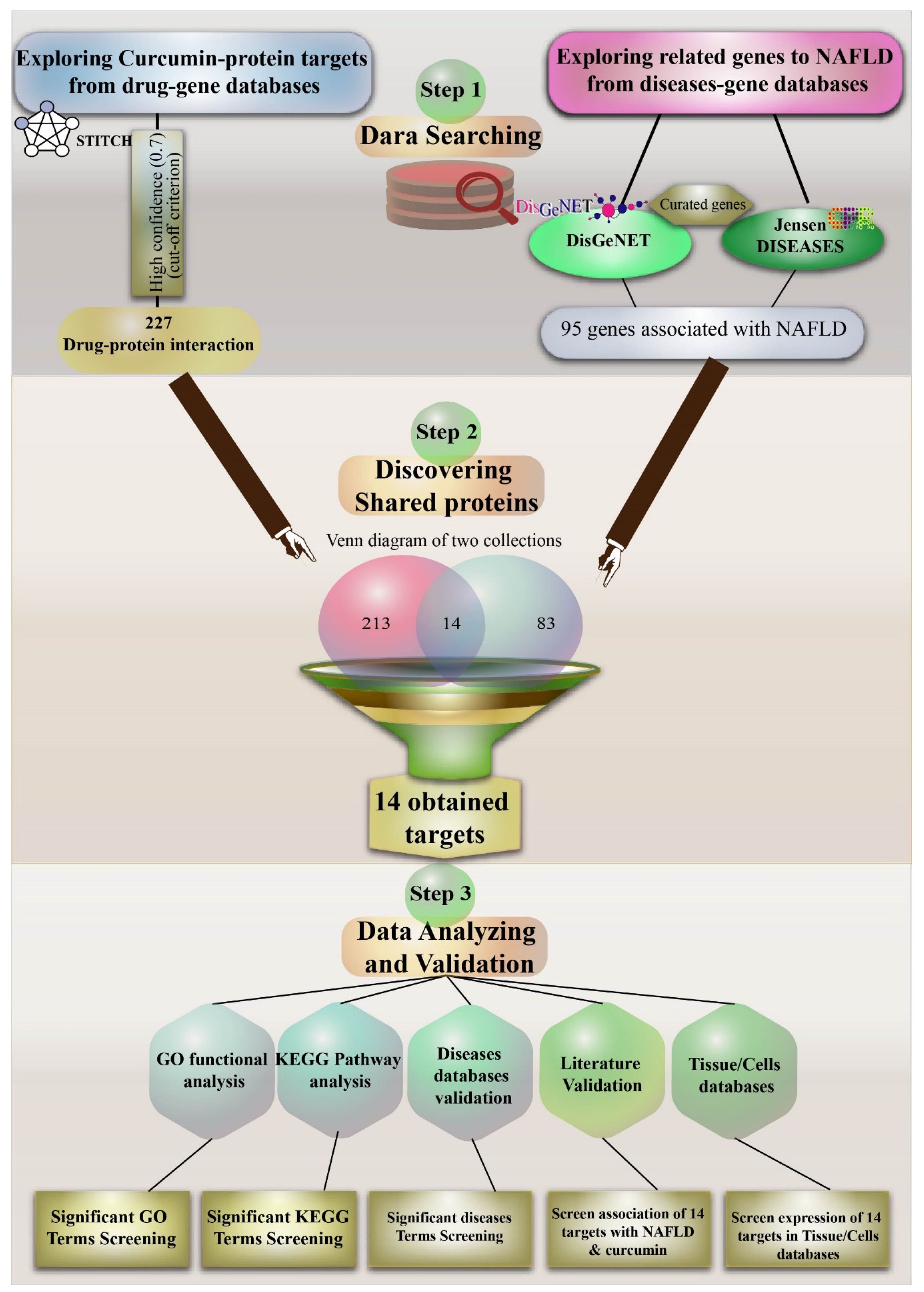
2. Methods
2.1. Curcumin and Target Search
2.2. Exploring Important NAFLD Genes in DISEASES and DisGeNET Databases
2.3. Venn Diagram to Obtain Important Curcumin Interaction Protein Targets in NAFLD
2.4. Gene Ontology Pathway Enrichment Analyses for Target Proteins of Curcumin
3. Results
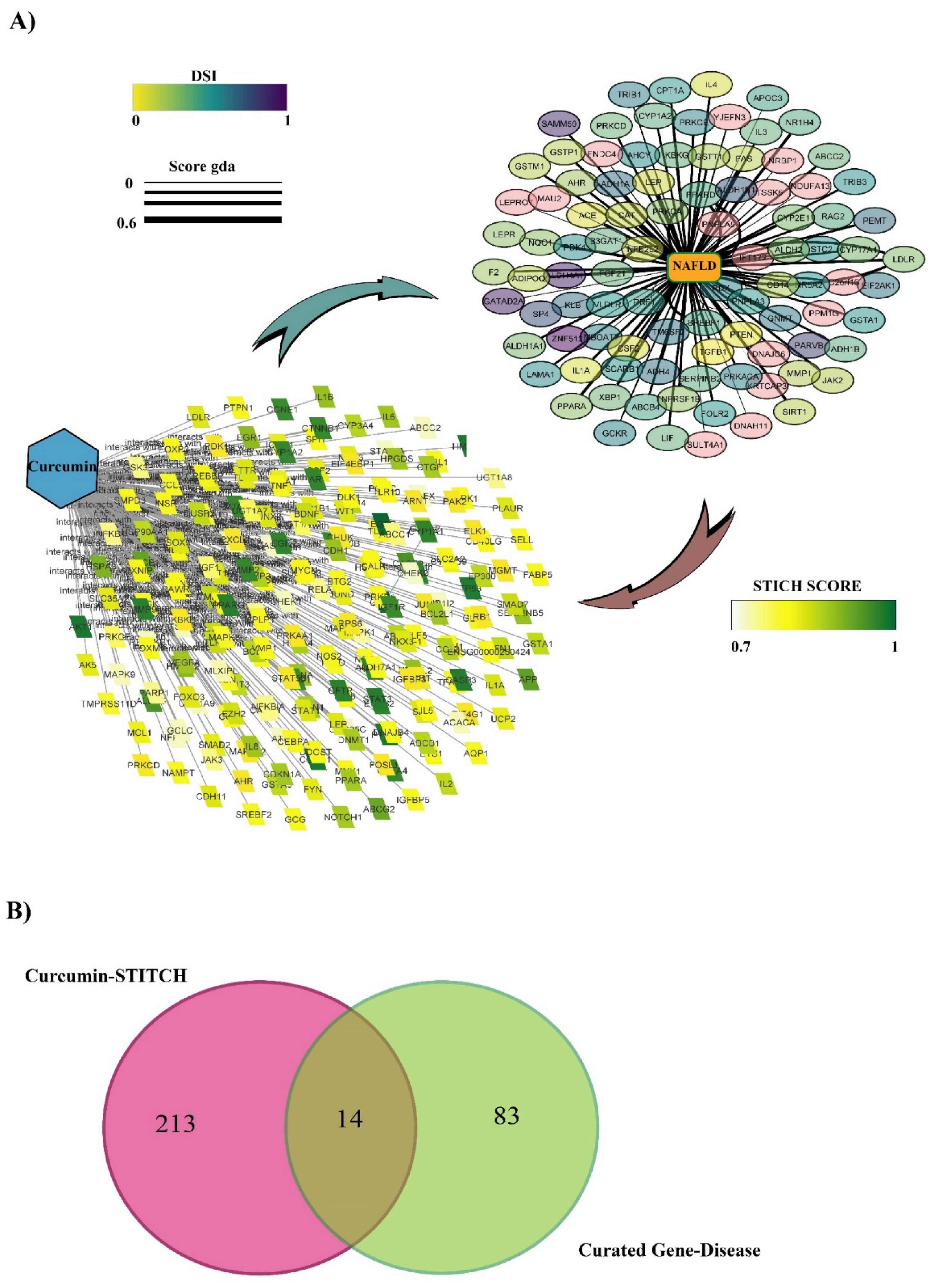
3.1. Protein Target Interaction with Curcumin in the STITCH Database
3.2. Discovering Curated NAFLD Genes
3.3. The Overlap of Curcumin Targets on the STITCH and Curated NAFLD Genes Were Visualized Using a Venn Diagram
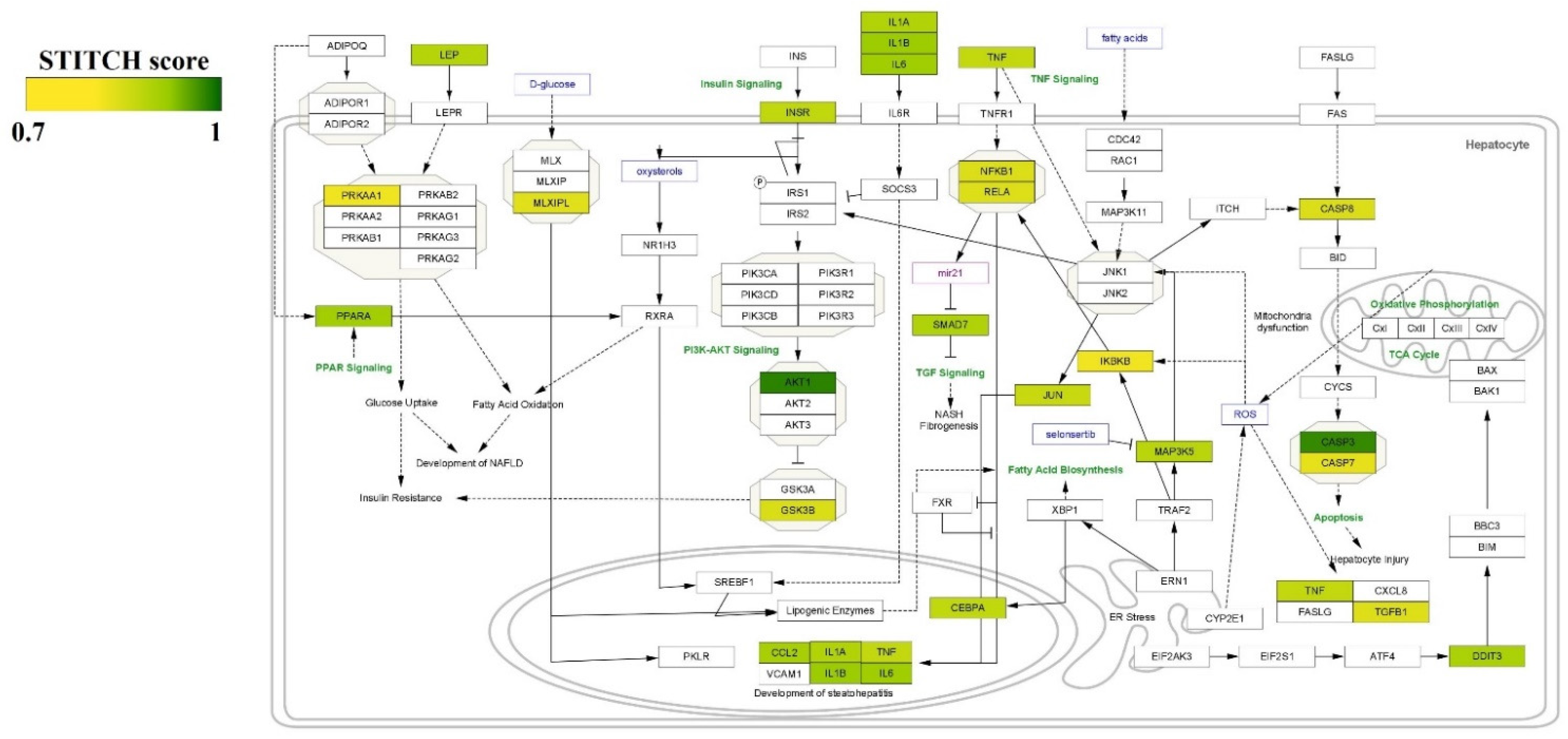
3.4. GO and KEGG Enrichment Analyses of Protein Targets of Curcumin
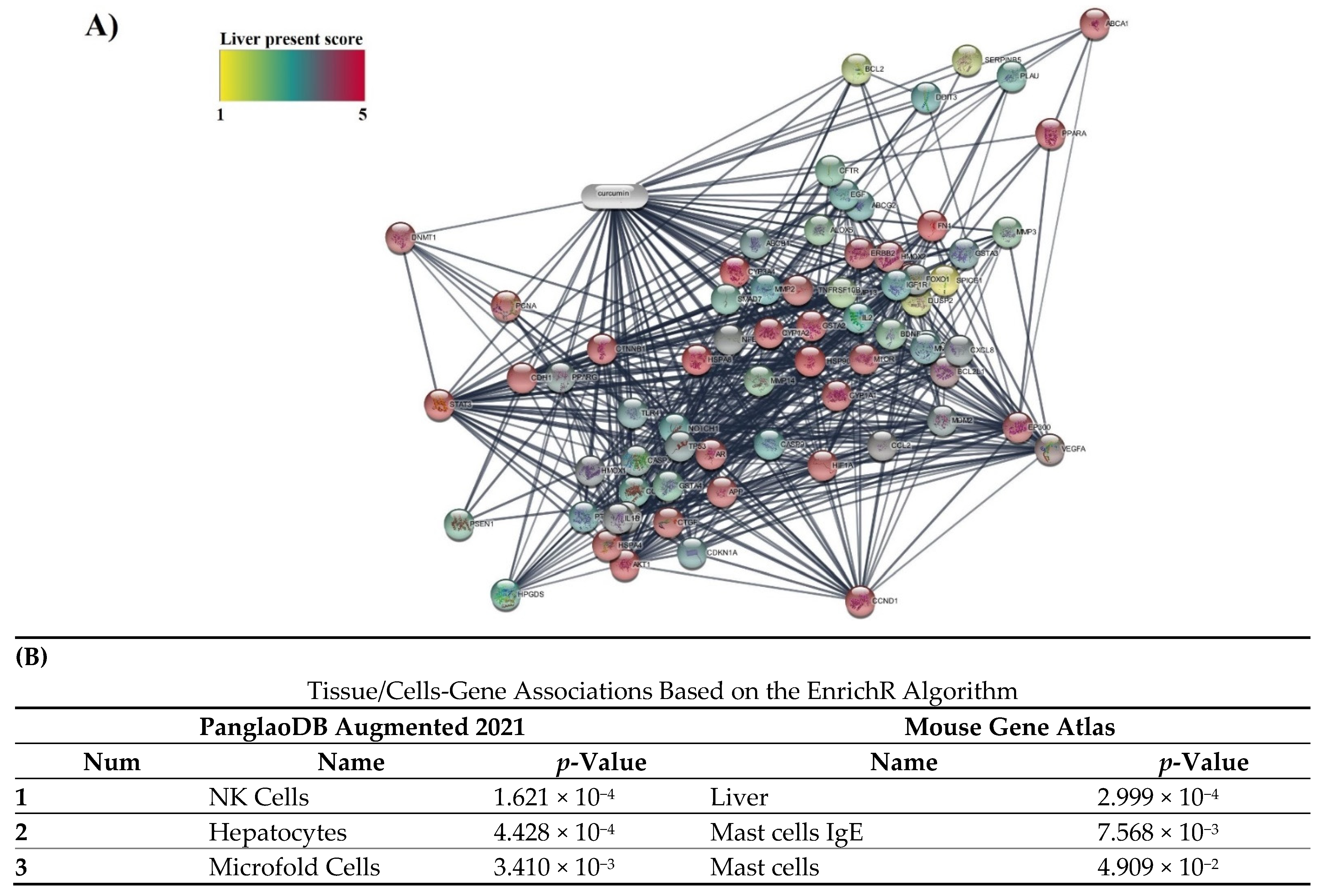
3.5. Specificity of 14 Obtained Protein Targets with Cells/Liver Tissue and Fatty Liver Disease in Databases
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Murag, S.; Ahmed, A.; Kim, D. Recent Epidemiology of Nonalcoholic Fatty Liver Disease. Gut Liver 2021, 15, 206–216. [Google Scholar] [CrossRef] [PubMed]
- Manne, V.; Handa, P.; Kowdley, K.V. Pathophysiology of Nonalcoholic Fatty Liver Disease/Nonalcoholic Steatohepatitis. Clin. Liver Dis. 2018, 22, 23–37. [Google Scholar] [CrossRef] [PubMed]
- Mansour-Ghanaei, F.; Joukar, F.; Mobaraki, S.N.; Mavaddati, S.; Hassanipour, S.; Sepehrimanesh, M. Prevalence of non-alcoholic fatty liver disease in patients with diabetes mellitus, hyperlipidemia, obesity and polycystic ovary syndrome: A cross-sectional study in north of Iran. Diabetes Metab. Syndr. Clin. Res. Rev. 2019, 13, 1591–1596. [Google Scholar] [CrossRef] [PubMed]
- Targher, G.; Byrne, C.D.; Tilg, H. NAFLD and increased risk of cardiovascular disease: Clinical associations, pathophysiological mechanisms and pharmacological implications. Gut 2020, 69, 1691–1705. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global epidemiology of nonalcoholic fatty liver disease-Meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology 2016, 64, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Mundi, M.S.; Velapati, S.; Patel, J.; Kellogg, T.A.; Abu Dayyeh, B.K.; Hurt, R.T. Evolution of NAFLD and Its Management. Nutr. Clin. Pract. 2020, 35, 72–84. [Google Scholar] [CrossRef]
- Sookoian, S.; Pirola, C.J. Precision medicine in nonalcoholic fatty liver disease: New therapeutic insights from genetics and systems biology. Clin. Mol. Hepatol. 2020, 26, 461–475. [Google Scholar] [CrossRef]
- Nobili, V.; Alisi, A.; Valenti, L.; Miele, L.; Feldstein, A.E.; Alkhouri, N. NAFLD in children: New genes, new diagnostic modalities and new drugs. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 517–530. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, T.; Li, J.; Wang, S.; Qiu, F.; Yu, H.; Zhang, Y.; Wang, T. Effects of Natural Products on Fructose-Induced Nonalcoholic Fatty Liver Disease (NAFLD). Nutrients 2017, 9, 96. [Google Scholar] [CrossRef]
- Reimer, K.C.; Wree, A.; Roderburg, C.; Tacke, F. New drugs for NAFLD: Lessons from basic models to the clinic. Hepatol. Int. 2020, 14, 8–23. [Google Scholar] [CrossRef]
- Afshari, A.R.; Jalili-Nik, M.; Abbasinezhad-Moud, F.; Javid, H.; Karimi, M.; Mollazadeh, H.; Jamialahmadi, T.; Sathyapalan, T.; Sahebkar, A. Anti-tumor Effects of Curcuminoids in Glioblastoma Multiforme: An Updated Literature Review. Curr. Med. Chem. 2021, 28, 8116–8138. [Google Scholar] [CrossRef] [PubMed]
- Gorabi, A.M.; Kiaie, N.; Hajighasemi, S.; Jamialahmadi, T.; Majeed, M.; Sahebkar, A. The Effect of Curcumin on the Differentiation of Mesenchymal Stem Cells into Mesodermal Lineage. Molecules 2019, 24, 4029. [Google Scholar] [CrossRef] [PubMed]
- Mohajeri, M.; Bianconi, V.; Ávila-Rodriguez, M.F.; Barreto, G.E.; Jamialahmadi, T.; Pirro, M.; Sahebkar, A. Curcumin: A phytochemical modulator of estrogens and androgens in tumors of the reproductive system. Pharmacol. Res. 2020, 156, 104765. [Google Scholar] [CrossRef] [PubMed]
- Alidadi, M.; Jamialahmadi, T.; Cicero, A.F.; Bianconi, V.; Pirro, M.; Banach, M.; Sahebkar, A. The potential role of plant-derived natural products in improving arterial stiffness: A review of dietary intervention studies. Trends Food Sci. Technol. 2020, 99, 426–440. [Google Scholar] [CrossRef]
- Farhood, B.; Mortezaee, K.; Goradel, N.H.; Khanlarkhani, N.; Salehi, E.; Nashtaei, M.S.; Najafi, M.; Sahebkar, A. Curcumin as an anti-inflammatory agent: Implications to radiotherapy and chemotherapy. J. Cell. Physiol. 2019, 234, 5728–5740. [Google Scholar] [CrossRef] [PubMed]
- Ganjali, S.; Blesso, C.N.; Banach, M.; Pirro, M.; Majeed, M.; Sahebkar, A. Effects of curcumin on HDL functionality. Pharmacol. Res. 2017, 119, 208–218. [Google Scholar] [CrossRef]
- Mortezaee, K.; Salehi, E.; Mirtavoos-Mahyari, H.; Motevaseli, E.; Najafi, M.; Farhood, B.; Rosengren, R.J.; Sahebkar, A. Mechanisms of apoptosis modulation by curcumin: Implications for cancer therapy. J. Cell. Physiol. 2019, 234, 12537–12550. [Google Scholar] [CrossRef]
- Panahi, Y.; Ghanei, M.; Bashiri, S.; Hajihashemi, A.; Sahebkar, A. Short-term Curcuminoid Supplementation for Chronic Pulmonary Complications due to Sulfur Mustard Intoxication: Positive Results of a Randomized Double-blind Placebo-controlled Trial. Drug Res. 2014, 65, 567–573. [Google Scholar] [CrossRef]
- Parsamanesh, N.; Moossavi, M.; Bahrami, A.; Butler, A.E.; Sahebkar, A. Therapeutic potential of curcumin in diabetic complications. Pharmacol. Res. 2018, 136, 181–193. [Google Scholar] [CrossRef]
- Sahebkar, A.; Henrotin, Y. Analgesic Efficacy and Safety of Curcuminoids in Clinical Practice: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Pain Med. 2016, 17, 1192–1202. [Google Scholar] [CrossRef]
- Shakeri, A.; Cicero, A.F.G.; Panahi, Y.; Mohajeri, M.; Sahebkar, A. Curcumin: A naturally occurring autophagy modulator. J. Cell. Physiol. 2019, 234, 5643–5654. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Harsha, C.; Parama, D.; Girisa, S.; Daimary, U.D.; Mao, X.; Kunnumakkara, A.B. Current clinical developments in curcumin-based therapeutics for cancer and chronic diseases. Phytotherapy Res. 2021, 35, 6768–6801. [Google Scholar] [CrossRef] [PubMed]
- Amato, A.; Caldara, G.-F.; Nuzzo, D.; Baldassano, S.; Picone, P.; Rizzo, M.; Mulè, F.; Di Carlo, M. NAFLD and Atherosclerosis Are Prevented by a Natural Dietary Supplement Containing Curcumin, Silymarin, Guggul, Chlorogenic Acid and Inulin in Mice Fed a High-Fat Diet. Nutrients 2017, 9, 492. [Google Scholar] [CrossRef] [PubMed]
- Cicero, A.F.G.; Sahebkar, A.; Fogacci, F.; Bove, M.; Giovannini, M.; Borghi, C. Effects of phytosomal curcumin on anthropometric parameters, insulin resistance, cortisolemia and non-alcoholic fatty liver disease indices: A double-blind, placebo-controlled clinical trial. Eur. J. Nutr. 2020, 59, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Hariri, M.; Gholami, A.; Mirhafez, S.R.; Bidkhori, M.; Sahebkar, A. A pilot study of the effect of curcumin on epigenetic changes and DNA damage among patients with non-alcoholic fatty liver disease: A randomized, double-blind, placebo-controlled, clinical trial. Complement. Ther. Med. 2020, 51, 102447. [Google Scholar] [CrossRef] [PubMed]
- Jalali, M.; Mahmoodi, M.; Mosallanezhad, Z.; Jalali, R.; Imanieh, M.H.; Moosavian, S.P. The effects of curcumin supplementation on liver function, metabolic profile and body composition in patients with non-alcoholic fatty liver disease: A systematic review and meta-analysis of randomized controlled trials. Complement. Ther. Med. 2020, 48, 102283. [Google Scholar] [CrossRef] [PubMed]
- Leclercq, I.A.; Farrell, G.C.; Sempoux, C.; Peña, A.D.; Horsmans, Y. Curcumin inhibits NF-κB activation and reduces the severity of experimental steatohepatitis in mice. J. Hepatol. 2004, 41, 926–934. [Google Scholar] [CrossRef]
- Rahmani, S.; Asgary, S.; Askari, G.; Keshvari, M.; Hatamipour, M.; Feizi, A.; Sahebkar, A. Treatment of Non-alcoholic Fatty Liver Disease with Curcumin: A Randomized Placebo-controlled Trial. Phytother. Res. 2016, 30, 1540–1548. [Google Scholar] [CrossRef]
- Saberi-Karimian, M.; Keshvari, M.; Ghayour-Mobarhan, M.; Salehizadeh, L.; Rahmani, S.; Behnam, B.; Jamialahmadi, T.; Asgary, S.; Sahebkar, A. Effects of curcuminoids on inflammatory status in patients with non-alcoholic fatty liver disease: A randomized controlled trial. Complement. Ther. Med. 2020, 49, 102322. [Google Scholar] [CrossRef]
- Vizzutti, F.; Provenzano, A.; Galastri, S.; Milani, S.; Delogu, W.; Novo, E.; Caligiuri, A.; Zamara, E.; Arena, U.; Laffi, G.; et al. Curcumin limits the fibrogenic evolution of experimental steatohepatitis. Lab. Investig. 2010, 90, 104–115. [Google Scholar] [CrossRef]
- Bagherniya, M.; Nobili, V.; Blesso, C.N.; Sahebkar, A. Medicinal plants and bioactive natural compounds in the treatment of non-alcoholic fatty liver disease: A clinical review. Pharmacol. Res. 2018, 130, 213–240. [Google Scholar] [CrossRef] [PubMed]
- Likić, V.A.; McConville, M.J.; Lithgow, T.; Bacic, A. Systems Biology: The Next Frontier for Bioinformatics. Adv. Bioinform. 2010, 2010, 268925. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, M.; Szklarczyk, D.; Franceschini, A.; Campillos, M.; von Mering, C.; Jensen, L.J.; Beyer, A.; Bork, P. STITCH 2: An interaction network database for small molecules and proteins. Nucleic Acids Res. 2010, 38, D552–D556. [Google Scholar] [CrossRef] [PubMed]
- Piñero, J.; Ramírez-Anguita, J.M.; Saüch-Pitarch, J.; Ronzano, F.; Centeno, E.; Sanz, F.; Furlong, L.I. The DisGeNET knowledge platform for disease genomics: 2019 update. Nucleic Acids Res. 2020, 48, D845–D855. [Google Scholar] [CrossRef]
- Pletscher-Frankild, S.; Pallejà, A.; Tsafou, K.; Binder, J.X.; Jensen, L.J. DISEASES: Text mining and data integration of disease–gene associations. Methods 2015, 74, 83–89. [Google Scholar] [CrossRef]
- The Gene Ontology Consortium. gene ontology Consortium: Going forward. Nucleic Acids Res. 2015, 43, D1049–D1056. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef]
- Franzén, O.; Gan, L.-M.; Björkegren, J.L.M. PanglaoDB: A web server for exploration of mouse and human single-cell RNA sequencing data. Database 2019, 2019, baz046. [Google Scholar] [CrossRef]
- Eppig, J.T.; Richardson, J.E.; Kadin, J.A.; Ringwald, M.; Blake, J.A.; Bult, C.J. Mouse Genome Informatics (MGI): Reflecting on 25 years. Mamm. Genome 2015, 26, 272–284. [Google Scholar] [CrossRef] [PubMed]
- Byrne, C.D.; Targher, G. NAFLD: A multisystem disease. J. Hepatol. 2015, 62, S47–S64. [Google Scholar] [CrossRef] [PubMed]
- Margini, C.; Dufour, J.F. The story of HCC in NAFLD: From epidemiology, across pathogenesis, to prevention and treatment. Liver Int. J. 2016, 36, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, R.P.; Moore, M.; Moore, A.N.; Healy, J.C.; Roberts, M.D.; Rector, R.S.; Martin, J.S. Curcumin supplementation mitigates NASH development and progression in female Wistar rats. Physiol. Rep. 2018, 6, e13789. [Google Scholar] [CrossRef] [PubMed]
- Ferramosca, A.; Di Giacomo, M.; Zara, V. Antioxidant dietary approach in treatment of fatty liver: New insights and updates. World J. Gastroenterol. 2017, 23, 4146–4157. [Google Scholar] [CrossRef]
- Gheibi, S.; Ghaleh, H.E.G.; Motlagh, B.M.; Azarbayjani, A.F.; Zarei, L. Therapeutic effects of curcumin and ursodexycholic acid on non-alcoholic fatty liver disease. Biomed. Pharmacother. 2019, 115, 108938. [Google Scholar] [CrossRef] [PubMed]
- Kersten, S.; Stienstra, R. The role and regulation of the peroxisome proliferator activated receptor alpha in human liver. Biochimie 2017, 136, 75–84. [Google Scholar] [CrossRef]
- Lakhia, R.; Yheskel, M.; Flaten, A.; Quittner-Strom, E.B.; Holland, W.L.; Patel, V. PPARα agonist fenofibrate enhances fatty acid β-oxidation and attenuates polycystic kidney and liver disease in mice. Am. J. Physiol. Physiol. 2018, 314, F122–F131. [Google Scholar] [CrossRef]
- Karahashi, M.; Hoshina, M.; Yamazaki, T.; Sakamoto, T.; Mitsumoto, A.; Kawashima, Y.; Kudo, N. Fibrates Reduce Triacylglycerol Content by Upregulating Adipose Triglyceride Lipase in the Liver of Rats. J. Pharmacol. Sci. 2013, 123, 356–370. [Google Scholar] [CrossRef]
- Pan, S.-Y.; Yu, Q.; Zhang, Y.; Wang, X.-Y.; Sun, N.; Yu, Z.-L.; Ko, K.-M. Dietary Fructus Schisandrae extracts and fenofibrate regulate the serum/hepatic lipid-profile in normal and hypercholesterolemic mice, with attention to hepatotoxicity. Lipids Health Dis. 2012, 11, 120. [Google Scholar] [CrossRef]
- Pan, S.-Y.; Jia, Z.-H.; Zhang, Y.; Yu, Q.; Wang, X.-Y.; Sun, N.; Zhu, P.-L.; Yu, Z.-L.; Ko, K.-M. A Novel Mouse Model of Combined Hyperlipidemia Associated with Steatosis and Liver Injury by a Single-Dose Intragastric Administration of Schisandrin B/Cholesterol/Bile Salts Mixture. J. Pharmacol. Sci. 2013, 123, 110–119. [Google Scholar] [CrossRef]
- Naderali, E.K.; Fatani, S.; Itua, I.; Wong, C. The effects of diet-induced obesity on hepatocyte insulin signaling pathways and induction of non-alcoholic liver damage. Int. J. Gen. Med. 2011, 4, 211–219. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hong, X.Z.; Da Li, L.; Wu, L.M. Effects of Fenofibrate and Xuezhikang on Highfat Diet-induced Nonalcoholic Fatty Liver Disease. Clin. Exp. Pharmacol. Physiol. 2007, 34, 27–35. [Google Scholar] [CrossRef]
- Shiri-Sverdlov, R.; Wouters, K.; van Gorp, P.; Gijbels, M.J.; Noel, B.; Buffat, L.; Staels, B.; Maeda, N.; van Bilsen, M.; Hofker, M.H. Early diet-induced non-alcoholic steatohepatitis in APOE2 knock-in mice and its prevention by fibrates. J. Hepatol. 2006, 44, 732–741. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.R.; Giri, S.R.; Bhoi, B.; Trivedi, C.; Rath, A.; Rathod, R.; Ranvir, R.; Kadam, S.; Patel, H.; Swain, P.; et al. Dual PPARα/γ agonist saroglitazar improves liver histopathology and biochemistry in experimental NASH models. Liver Int. 2018, 38, 1084–1094. [Google Scholar] [CrossRef] [PubMed]
- Hamed, A.M.; El-Kharashi, O.A.; Boctor, S.S.; Abd-Elaziz, L.F. Potential involvement of PPAR α activation in diminishing the hepatoprotective effect of fenofibrate in NAFLD: Accuracy of non- invasive panel in determining the stage of liver fibrosis in rats. Biomed. Pharmacother. 2017, 85, 68–78. [Google Scholar] [CrossRef]
- Seo, Y.S.; Kim, J.H.; Jo, N.Y.; Choi, K.M.; Baik, S.H.; Park, J.-J.; Kim, J.S.; Byun, K.S.; Bak, Y.-T.; Lee, C.H.; et al. PPAR agonists treatment is effective in a nonalcoholic fatty liver disease animal model by modulating fatty-acid metabolic enzymes. J. Gastroenterol. Hepatol. 2006, 23, 102–109. [Google Scholar] [CrossRef]
- Kong, D.; Zhang, Z.; Chen, L.; Huang, W.; Zhang, F.; Wang, L.; Wang, Y.; Cao, P.; Zheng, S. Curcumin blunts epithelial-mesenchymal transition of hepatocytes to alleviate hepatic fibrosis through regulating oxidative stress and autophagy. Redox Biol. 2020, 36, 101600. [Google Scholar] [CrossRef]
- Zeng, K.; Tian, L.; Patel, R.; Shao, W.; Song, Z.; Manuel, J.; Ma, X.; McGilvray, I.; Weng, J.; Jin, T.; et al. Diet polyphenol curcumin stimulates hepatic Fgf21 production and restores its sensitivity in high fat diet fed male mice. Endocrinology 2017, 158, 277–292. [Google Scholar] [CrossRef][Green Version]
- Li, L.; Fu, J.; Sun, J.; Liu, D.; Chen, C.; Wang, H.; Hou, Y.; Xu, Y.; Pi, J. Is Nrf2-ARE a potential target in NAFLD mitigation? Curr. Opin. Toxicol. 2019, 13, 35–44. [Google Scholar] [CrossRef]
- Chowdhry, S.; Nazmy, M.H.; Meakin, P.; Dinkova-Kostova, A.; Walsh, S.V.; Tsujita, T.; Dillon, J.; Ashford, M.; Hayes, J.D. Loss of Nrf2 markedly exacerbates nonalcoholic steatohepatitis. Free Radic. Biol. Med. 2010, 48, 357–371. [Google Scholar] [CrossRef] [PubMed]
- Ramadori, P.; Drescher, H.; Erschfeld, S.; Schumacher, F.; Berger, C.; Fragoulis, A.; Schenkel, J.; Kensler, T.; Wruck, C.J.; Trautwein, C.; et al. Hepatocyte-specific Keap1 deletion reduces liver steatosis but not inflammation during non-alcoholic steatohepatitis development. Free Radic. Biol. Med. 2016, 91, 114–126. [Google Scholar] [CrossRef]
- Lee, L.-Y.; Köhler, U.A.; Zhang, L.; Roenneburg, E.; Werner, S.; Johnson, J.A.; Foley, D.P. Activation of the Nrf2-ARE pathway in hepatocytes protects against steatosis in nutritionally induced non-alcoholic steatohepatitis in mice. Toxicol. Sci. 2014, 142, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Park, J.S.; Lee, Y.S.; Han, J.; Lee, D.-K.; Kwon, S.W.; Han, D.H.; Lee, Y.-H.; Bae, S.H. SQSTM1/p62 activates NFE2L2/NRF2 via ULK1-mediated autophagic KEAP1 degradation and protects mouse liver from lipotoxicity. Autophagy 2020, 16, 1949–1973. [Google Scholar] [CrossRef]
- Ashrafizadeh, M.; Ahmadi, Z.; Mohamamdinejad, R.; Farkhondeh, T.; Samarghandian, S. Curcumin Activates the Nrf2 Pathway and Induces Cellular Protection Against Oxidative Injury. Curr. Mol. Med. 2020, 20, 116–133. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Zhang, Y.; Zhang, X.; Aa, J.; Wang, G.; Xie, Y. Curcumin regulates endogenous and exogenous metabolism via Nrf2-FXR-LXR pathway in NAFLD mice. Biomed. Pharmacother. 2018, 105, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Dai, C.; Liu, Q.; Li, J.; Qiu, J. Curcumin Attenuates on Carbon Tetrachloride-Induced Acute Liver Injury in Mice via Modulation of the Nrf2/HO-1 and TGF-β1/Smad3 Pathway. Molecules 2018, 23, 215. [Google Scholar] [CrossRef]
- Lu, C.; Xu, W.; Zheng, S. Nrf2 activation is required for curcumin to induce lipocyte phenotype in hepatic stellate cells. Biomed. Pharmacother. 2017, 95, 1–10. [Google Scholar] [CrossRef]
- Swellam, M.; Hamdy, N. Association of nonalcoholic fatty liver disease with a single nucleotide polymorphism on the gene encoding leptin receptor. IUBMB Life 2012, 64, 180–186. [Google Scholar] [CrossRef]
- Xu, D.; Huang, X.-D.; Luo, H.-S.; Yuan, J.-P.; Zhang, H.; Wu, J. Impaired activation of phosphatidylinositol 3-kinase by leptin in NAFLD: A novel mechanism of hepatic leptin resistance. World Chin. J. Dig. 2012, 20, 3095–3100. [Google Scholar] [CrossRef]
- Xu, D.; Huang, X.-D.; Yuan, J.-P.; Wu, J.; Fan, Y.; Luo, H.-S.; Yang, Y.-H. Impaired Activation of Phosphatidylinositol 3-Kinase by Leptin is a Novel Mechanism of Hepatic Leptin Resistance in NAFLD. Hepatogastroenterology 2011, 58, 1703–1707. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Zheng, S.; Chen, A. Curcumin Eliminates Leptin’s Effects on Hepatic Stellate Cell Activation via Interrupting Leptin Signaling. Endocrinology 2009, 150, 3011–3020. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.-H.; Liao, J.; Zhang, J.-Y.; Hao, X.-H.; Liang, C.; Wang, L.-H.; Xue, H.; Zhang, K.; Yan, G.-T. Localized leptin release may be an important mechanism of curcumin action after acute ischemic injuries. J. Trauma Acute Care Surg. 2013, 74, 1044–1051. [Google Scholar] [CrossRef] [PubMed]
- Nejati-Koshki, K.; Akbarzadeh, A.; Pourhassan-Moghaddam, M. Curcumin inhibits leptin gene expression and secretion in breast cancer cells by estrogen receptors. Cancer Cell Int. 2014, 14, 66. [Google Scholar] [CrossRef] [PubMed]
- Askari, S.; Salehi, R.; Zarghami, N.; Akbarzadeh, A.; Rahmati-Yamchi, M. The anticancer effects of biodegradable nanomagnetic dual natural components on the leptin gene expression in lung cancer. Artif. Cells Nanomed. Biotechnol. 2016, 44, 1753–1763. [Google Scholar] [CrossRef]
- Lu, S.; Zhao, H.; Zhou, Y.; Xu, F. Curcumin Affects Leptin-Induced Expression of Methionine Adenosyltransferase 2A in Hepatic Stellate Cells by Inhibition of JNK Signaling. Pharmacology 2021, 106, 426–434. [Google Scholar] [CrossRef]
- Floeth, M.; Elges, S.; Gerss, J.; Schwöppe, C.; Kessler, T.; Herold, T.; Wardelmann, E.; Berdel, W.E.; Lenz, G.; Mikesch, J.; et al. Low-density lipoprotein receptor (LDLR) is an independent adverse prognostic factor in acute myeloid leukaemia. Br. J. Haematol. 2021, 192, 494–503. [Google Scholar] [CrossRef]
- Álvarez-Amor, L.; Sierra, A.L.; Cárdenas, A.; López-Bermudo, L.; López-Beas, J.; Andújar, E.; Pérez-Alegre, M.; Gallego-Durán, R.; Varela, L.M.; Martin-Montalvo, A.; et al. Extra virgin olive oil improved body weight and insulin sensitivity in high fat diet-induced obese LDLr−/−.Leiden mice without attenuation of steatohepatitis. Sci. Rep. 2021, 11, 8250. [Google Scholar] [CrossRef]
- Schoemaker, M.H.; Kleemann, R.; Morrison, M.C.; Verheij, J.; Salic, K.; Van Tol, E.A.F.; Kooistra, T.; Wielinga, P.Y. A casein hydrolysate based formulation attenuates obesity and associated non-alcoholic fatty liver disease and atherosclerosis in LDLr-/-.Leiden mice. PLoS ONE 2017, 12, e0180648. [Google Scholar] [CrossRef]
- Li, L.; Zhang, G.-F.; Lee, K.; Lopez, R.; Previs, S.F.; Willard, B.; McCullough, A.; Kasumov, T. A Western diet induced NAFLD in LDLR−/− mice is associated with reduced hepatic glutathione synthesis. Free Radic. Biol. Med. 2016, 96, 13–21. [Google Scholar] [CrossRef]
- He, Y.; Yang, W.; Gan, L.; Liu, S.; Ni, Q.; Bi, Y.; Han, T.; Liu, Q.; Chen, H.; Hu, Y.; et al. Silencing HIF-1α aggravates non-alcoholic fatty liver disease in vitro through inhibiting PPAR-α/ANGPTL4 singling pathway. Gastroenterol. Hepatol. 2020, 44, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Tai, M.-H.; Chen, P.-K.; Chen, P.-Y.; Wu, M.-J.; Ho, C.-T.; Yen, J.-H. Curcumin enhances cell-surface LDLR level and promotes LDL uptake through downregulation of PCSK9 gene expression in HepG2 cells. Mol. Nutr. Food Res. 2014, 58, 2133–2145. [Google Scholar] [CrossRef] [PubMed]
- Dou, X.B.; Wo, X.D.; Fan, C.L.; Yu, Y.; Qian, Y.; Huang, X.L.; Ling, X. Effect of curcumin on the expression of low density lipoprotein receptor in HepG2 cell line. Chin. Pharm. J. 2007, 42, 572–575. [Google Scholar]
- Cai, Y.; Lu, D.; Zou, Y.; Zhou, C.; Liu, H.; Tu, C.; Li, F.; Zhang, S. Curcumin Protects Against Intestinal Origin Endotoxemia in Rat Liver Cirrhosis by Targeting PCSK9. J. Food Sci. 2017, 82, 772–780. [Google Scholar] [CrossRef]
- Corral, P.A.; Botello, J.F.; Xing, C. Design, synthesis, and enzymatic characterization of quinazoline-based CYP1A2 inhibitors. Bioorganic Med. Chem. Lett. 2020, 30, 126719. [Google Scholar] [CrossRef]
- Chiba, T.; Noji, K.; Shinozaki, S.; Suzuki, S.; Umegaki, K.; Shimokado, K. Diet-induced non-alcoholic fatty liver disease affects expression of major cytochrome P450 genes in a mouse model. J. Pharm. Pharmacol. 2016, 68, 1567–1576. [Google Scholar] [CrossRef]
- Jearapong, N.; Chatuphonprasert, W.; Jarukamjorn, K. Effect of tetrahydrocurcumin on the profiles of drug-metabolizing enzymes induced by a high fat and high fructose diet in mice. Chem.-Biol. Interact. 2015, 239, 67–75. [Google Scholar] [CrossRef]
- Fisher, C.D.; Lickteig, A.J.; Augustine, L.M.; Ranger-Moore, J.; Jackson, J.P.; Ferguson, S.S.; Cherrington, N.J. Hepatic Cytochrome P450 Enzyme Alterations in Humans with Progressive Stages of Nonalcoholic Fatty Liver Disease. Drug Metab. Dispos. 2009, 37, 2087–2094. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, W.-H.; Chen, B.-L.; Fan, L.; Han, Y.; Wang, G.; Hu, D.-L.; Tan, Z.-R.; Zhou, G.; Cao, S.; et al. Plant Polyphenol Curcumin Significantly Affects CYPIA2 and CYP2A6 Activity in Healthy, Male Chinese Volunteers. Ann. Pharmacother. 2010, 44, 1038–1045. [Google Scholar] [CrossRef]
- Abd-Elhakim, Y.M.; Moustafa, G.G.; El-Sharkawy, N.I.; Hussein, M.M.; Ghoneim, M.H.; El Deib, M.M. The ameliorative effect of curcumin on hepatic CYP1A1 and CYP1A2 genes dysregulation and hepatorenal damage induced by fenitrothion oral intoxication in male rats. Pestic. Biochem. Physiol. 2021, 179, 104959. [Google Scholar] [CrossRef]
- Wu, R.; Cui, X.; Dong, W.; Zhou, M.; Simms, H.H.; Wang, P. Suppression of hepatocyte CYP1A2 expression by Kupffer cells via AhR pathway: The central role of proinflammatory cytokines. Int. J. Mol. Med. 2006, 18, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Appiahopong, R.; Commandeur, J.; Vanvugtlussenburg, B.; Vermeulen, N.P. Inhibition of human recombinant cytochrome P450s by curcumin and curcumin decomposition products. Toxicology 2007, 235, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Sookoian, S.; Gianotti, T.F.; Rosselli, M.S.; Burgueño, A.L.; Castaño, G.O.; Pirola, C.J. Liver transcriptional profile of atherosclerosis-related genes in human nonalcoholic fatty liver disease. Atherosclerosis 2011, 218, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Elsayed, H.R.H.; El-Nablaway, M.; Othman, B.H.; Abdalla, A.M.; El Nashar, E.M.; Abd-Elmonem, M.M.; El-Gamal, R. Can Dasatinib Ameliorate the Hepatic changes, Induced by Long Term Western Diet, in Mice? Ann. Anat. Anat. Anz. 2021, 234, 151626. [Google Scholar] [CrossRef] [PubMed]
- Mikuriya, Y.; Tashiro, H.; Kuroda, S.; Nambu, J.; Kobayashi, T.; Amano, H.; Tanaka, Y.; Ohdan, H. Fatty liver creates a pro-metastatic microenvironment for hepatocellular carcinoma through activation of hepatic stellate cells. Int. J. Cancer 2014, 136, E3–E13. [Google Scholar] [CrossRef]
- Poniachik, J.; Csendes, A.; Díaz, J.C.; Rojas, J.; Burdiles, P.; Maluenda, F.; Smok, G.; Rodrigo, R.; Videla, L.A. Increased production of IL-1α and TNF-α in lipopolysaccharide-stimulated blood from obese patients with non-alcoholic fatty liver disease. Cytokine 2006, 33, 252–257. [Google Scholar] [CrossRef]
- Xu, Y.X.; Pindolia, K.; Janakiraman, N.; Noth, C.J.; Chapman, R.A.; Gautam, S.C. Curcumin, a compound with anti-inflammatory and anti-oxidant properties, down-regulates chemokine expression in bone marrow stromal cells. Exp. Hematol. 1997, 25, 413–422. [Google Scholar]
- Xu, Y.X.; Pindolia, K.R.; Janakiraman, N.; Chapman, R.A.; Gautam, S.C. Curcumin inhibits IL1α and TNFα induction of AP-1 and NF-kB DNA-binding activity in bone marrow stromal cells. Hematopathol. Mol. Hematol. 1997, 11, 49–62. [Google Scholar]
- Michaud, V.; Tran, M.; Pronovost, B.; Bouchard, P.; Bilodeau, S.; Alain, K.; Vadnais, B.; Franco, M.; Bélanger, F.; Turgeon, J. Impact of GSTA1 Polymorphisms on Busulfan Oral Clearance in Adult Patients Undergoing Hematopoietic Stem Cell Transplantation. Pharmaceutics 2019, 11, 440. [Google Scholar] [CrossRef]
- Oniki, K.; Hori, M.; Saruwatari, J.; Morita, K.; Kajiwara, A.; Sakata, M.; Mihara, S.; Ogata, Y.; Nakagawa, K. Interactive effects of smoking and glutathione S-transferase polymorphisms on the development of non-alcoholic fatty liver disease. Toxicol. Lett. 2013, 220, 143–149. [Google Scholar] [CrossRef]
- Jin, X.-L.; Wang, K.; Li, Q.; Tian, W.-L.; Xue, X.-F.; Wu, L.-M.; Hu, F.-L. Antioxidant and anti-inflammatory effects of Chinese propolis during palmitic acid-induced lipotoxicity in cultured hepatocytes. J. Funct. Foods 2017, 34, 216–223. [Google Scholar] [CrossRef]
- Miyata, M.; Funaki, A.; Fukuhara, C.; Sumiya, Y.; Sugiura, Y. Taurine attenuates hepatic steatosis in a genetic model of fatty liver disease. J. Toxicol. Sci. 2020, 45, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Miyata, M.; Matsushita, K.; Shindo, R.; Shimokawa, Y.; Sugiura, Y.; Yamashita, M. Selenoneine Ameliorates Hepatocellular Injury and Hepatic Steatosis in a Mouse Model of NAFLD. Nutrients 2020, 12, 1898. [Google Scholar] [CrossRef] [PubMed]
- Robin, S.K.D.; Ansari, M.; Uppugunduri, C.R.S. Spectrophotometric Screening for Potential Inhibitors of Cytosolic Glutathione S-Transferases. J. Vis. Exp. 2020, e61347. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Ellis, B.; Sharma, A. Role of alpha class glutathione transferases (GSTs) in chemoprevention: GSTA1 and A4 overexpressing human leukemia (HL60) cells resist sulforaphane and curcumin induced toxicity. Phytother. Res. 2011, 25, 563–568. [Google Scholar] [CrossRef]
- Odenthal, J.; van Heumen, B.W.H.; Roelofs, H.M.J.; Morsche, R.H.M.T.; Marian, B.; Nagengast, F.M.; Peters, W.H.M. The Influence of Curcumin, Quercetin, and Eicosapentaenoic Acid on the Expression of Phase II Detoxification Enzymes in the Intestinal Cell Lines HT-29, Caco-2, HuTu 80, and LT97. Nutr. Cancer 2012, 64, 856–863. [Google Scholar] [CrossRef]
- Qadri, Q.; Sameer, A.; Shah, Z.; Hamid, A.; Alam, S.; Manzoor, S.; Siddiqi, M. Genetic polymorphism of the glutathione-S-transferase P1 gene (GSTP1) and susceptibility to prostate cancer in the Kashmiri population. Genet. Mol. Res. 2011, 10, 3038–3045. [Google Scholar] [CrossRef]
- Hori, M.; Oniki, K.; Nakagawa, T.; Takata, K.; Mihara, S.; Marubayashi, T.; Nakagawa, K. Association between combinations of glutathione-S-transferaseM1, T1 and P1 genotypes and non-alcoholic fatty liver disease. Liver Int. 2009, 29, 164–168. [Google Scholar] [CrossRef]
- Kirpich, I.A.; Gobejishvili, L.N.; Homme, M.B.; Waigel, S.; Cave, M.; Arteel, G.; Barve, S.S.; McClain, C.J.; Deaciuc, I.V. Integrated hepatic transcriptome and proteome analysis of mice with high-fat diet-induced nonalcoholic fatty liver disease. J. Nutr. Biochem. 2011, 22, 38–45. [Google Scholar] [CrossRef]
- Prysyazhnyuk, V.; Rossokha, Z.; Gorovenko, N. Variation in particular biochemical indicators, cytokine and adipokine profiles of the blood, and the structural and functional parameters of the liver in patients with nonalcoholic fatty liver disease and different genotypes by the polymorphic locus A313G of the GSTP1 gene. Cytol. Genet. 2017, 51, 455–461. [Google Scholar] [CrossRef]
- Hashemi, M.; Eskandari, E.; Fazaeli, A.; Bahari, A.; Hashemzehi, N.-A.; Shafieipour, S.; Taheri, M.; Moazeni-Roodi, A.; Zakeri, Z.; Bakhshipour, A.; et al. Association of Genetic Polymorphisms of Glutathione-S-Transferase Genes (GSTT1, GSTM1, and GSTP1) and Susceptibility to Nonalcoholic Fatty Liver Disease in Zahedan, Southeast Iran. DNA Cell Biol. 2012, 31, 672–677. [Google Scholar] [CrossRef] [PubMed]
- Nishinaka, T.; Ichijo, Y.; Ito, M.; Kimura, M.; Katsuyama, M.; Iwata, K.; Miura, T.; Terada, T.; Yabe-Nishimura, C. Curcumin activates human glutathione S-transferase P1 expression through antioxidant response element. Toxicol. Lett. 2007, 170, 238–247. [Google Scholar] [CrossRef] [PubMed]
- Duvoix, A.; Morceau, F.; Delhalle, S.; Schmitz, M.; Schnekenburger, M.; Galteau, M.-M.; Dicato, M.; Diederich, M. Induction of apoptosis by curcumin: Mediation by glutathione S-transferase P1-1 inhibition. Biochem. Pharmacol. 2003, 66, 1475–1483. [Google Scholar] [CrossRef]
- Li, H.; Li, L. Relationship of GSTP1 lower expression and multidrug resistance reversing of curcumin on human colon carcinoma cells. Zhonghua Yi Xue Za Zhi 2015, 95, 2478–2482. [Google Scholar]
- Carrillo-Gálvez, A.B.; Gálvez-Peisl, S.; González-Correa, J.E.; Haro-Carrillo, M.; Ayllón, V.; Carmona-Sáez, P.; Ramos-Mejía, V.; Galindo-Moreno, P.; Cara, F.E.; Granados-Principal, S.; et al. GARP is a key molecule for mesenchymal stromal cell responses to TGF-β and fundamental to control mitochondrial ROS levels. STEM CELLS Transl. Med. 2020, 9, 636–650. [Google Scholar] [CrossRef]
- Subudhi, S.; Drescher, H.K.; Dichtel, L.E.; Bartsch, L.M.; Chung, R.T.; Hutter, M.M.; Gee, D.W.; Meireles, O.R.; Witkowski, E.R.; Gelrud, L.; et al. Distinct Hepatic Gene-Expression Patterns of NAFLD in Patients with Obesity. Hepatol. Commun. 2021, 6, 77–89. [Google Scholar] [CrossRef]
- Tang, M.; Jia, H.; Chen, S.; Yang, B.; Patpur, B.K.; Song, W.; Chang, Y.; Li, J.; Yang, C. Significance of MR/OPN/HMGB1 axis in NAFLD-associated hepatic fibrogenesis. Life Sci. 2021, 264, 118619. [Google Scholar] [CrossRef]
- Brown, K.D.; Shah, M.H.; Liu, G.-S.; Chan, E.C.; Crowston, J.G.; Peshavariya, H.M. Transforming Growth Factor ?1–Induced NADPH Oxidase-4 Expression and Fibrotic Response in Conjunctival Fibroblasts. Investig. Opthalmol. Vis. Sci. 2017, 58, 3011–3017. [Google Scholar] [CrossRef]
- Fu, X.-Y.; Zhang, D.-W.; Li, Y.-D.; Jian-Zhao, N.; Tang, Y.-Q.; Niu, J.-Z.; Li, Y. Curcumin Treatment Suppresses CCR7 Expression and the Differentiation and Migration of Human Circulating Fibrocytes. Cell. Physiol. Biochem. 2015, 35, 489–498. [Google Scholar] [CrossRef]
- Li, H.; Liu, B.; Li, P.; Feng, L.; Ma, H.; Xuan, S.; Cao, Y. Inhibitory effects of curcumin on inflammatory cytokines in rats with paraquat poisoning. Chin. J. Ind. Hyg. Occup. Dis. 2015, 33, 689–692. [Google Scholar]
- Gordon, M.Y.; Riley, G.P.; Watt, S.M.; Greaves, M.F. Compartmentalization of a haematopoietic growth factor (GM-CSF) by glycosaminoglycans in the bone marrow microenvironment. Nature 1987, 326, 403–405. [Google Scholar] [CrossRef] [PubMed]
- Tan-Garcia, A.; Lai, F.; Yeong, J.P.S.; Irac, S.E.; Ng, P.Y.; Msallam, R.; Lim, J.C.T.; Wai, L.-E.; Tham, C.Y.; Choo, S.P.; et al. Liver fibrosis and CD206+ macrophage accumulation are suppressed by anti-GM-CSF therapy. JHEP Rep. 2020, 2, 100062. [Google Scholar] [CrossRef]
- Xin, X.; Jin, Y.; Wang, X.; Cai, B.; An, Z.; Hu, Y.-Y.; Feng, Q. A Combination of Geniposide and Chlorogenic Acid Combination Ameliorates Nonalcoholic Steatohepatitis in Mice by Inhibiting Kupffer Cell Activation. BioMed Res. Int. 2021, 2021, 6615881. [Google Scholar] [CrossRef] [PubMed]
- de Almeida-Souza, C.B.; Antunes, M.M.; Carbonera, F.; Godoy, G.; da Silva, M.A.; Masi, L.N.; Visentainer, J.V.; Curi, R.; Bazotte, R.B. A High-Fat Diet Induces Lower Systemic Inflammation than a High-Carbohydrate Diet in Mice. Metab. Syndr. Relat. Disord. 2021, 19, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Morrison, M.C.; Kleemann, R.; Van Koppen, A.; Hanemaaijer, R.; Verschuren, L. Key Inflammatory Processes in Human NASH Are Reflected in Ldlr−/−.Leiden Mice: A Translational Gene Profiling Study. Front. Physiol. 2018, 9, 132. [Google Scholar] [CrossRef]
- Kobayashi, T.; Hashimoto, S.; Horie, T. Curcumin inhibition of Dermatophagoides farinea-induced interleukin-5 (IL-5) and granulocyte macrophage-colony stimulating factor (GM-CSF) production by lymphocytes from bronchial asthmatics. Biochem. Pharmacol. 1997, 54, 819–824. [Google Scholar] [CrossRef]
- Latimer, B.; Ekshyyan, O.; Nathan, N.; Moore-Medlin, T.; Rong, X.; Ma, X.; Khandelwal, A.; Christy, H.T.; Abreo, F.; McClure, G.; et al. Enhanced Systemic Bioavailability of Curcumin Through Transmucosal Administration of a Novel Microgranular Formulation. Anticancer Res. 2015, 35, 6411–6418. [Google Scholar]
- Mellor, H.; Parker, P. The extended protein kinase C superfamily. Biochem. J. 1998, 332, 281–292. [Google Scholar] [CrossRef]
- Jornayvaz, F.R.; Shulman, G.I. Diacylglycerol Activation of Protein Kinase Cε and Hepatic Insulin Resistance. Cell Metab. 2012, 15, 574–584. [Google Scholar] [CrossRef]
- ter Horst, K.W.; Gilijamse, P.W.; Versteeg, R.I.; Ackermans, M.T.; Nederveen, A.J.; la Fleur, S.E.; Romijn, J.A.; Nieuwdorp, M.; Zhang, D.; Samuel, V.T.; et al. Hepatic Diacylglycerol-Associated Protein Kinase Cε Translocation Links Hepatic Steatosis to Hepatic Insulin Resistance in Humans. Cell Rep. 2017, 19, 1997–2004. [Google Scholar] [CrossRef]
- Birkenfeld, A.L.; Shulman, G.I. Nonalcoholic fatty liver disease, hepatic insulin resistance, and type 2 Diabetes. Hepatology 2013, 59, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Pany, S.; You, Y.; Das, J. Curcumin Inhibits Protein Kinase Cα Activity by Binding to Its C1 Domain. Biochemistry 2016, 55, 6327–6336. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, B.; Huang, F.; Liu, B.; Xie, Y. Curcumin inhibits lipolysis via suppression of ER stress in adipose tissue and prevents hepatic insulin resistance. J. Lipid Res. 2016, 57, 1243–1255. [Google Scholar] [CrossRef] [PubMed]
- Jemnitz, K.; Heredi-Szabo, K.; Janossy, J.; Ioja, E.; Vereczkey, L.; Krajcsi, P. ABCC2/Abcc2: A multispecific transporter with dominant excretory functions. Drug Metab. Rev. 2010, 42, 402–436. [Google Scholar] [CrossRef] [PubMed]
- Sookoian, S.; Castaño, G.; Gianotti, T.F.; Gemma, C.; Pirola, C.J. Polymorphisms of MRP2 (ABCC2) are associated with susceptibility to nonalcoholic fatty liver disease. J. Nutr. Biochem. 2009, 20, 765–770. [Google Scholar] [CrossRef]
- Sakulterdkiat, T.; Srisomsap, C.; Udomsangpetch, R.; Svasti, J.; Lirdprapamongkol, K. Curcumin resistance induced by hypoxia in HepG2 cells is mediated by multidrug-resistance-associated proteins. Anticancer Res. 2012, 32, 5337–5342. [Google Scholar]
- Cho, C.; Yang, C.; Wu, C.; Ho, J.; Yu, C.; Wu, S.; Yu, D. The modulation study of multiple drug resistance in bladder cancer by curcumin and resveratrol. Oncol. Lett. 2019, 18, 6869–6876. [Google Scholar] [CrossRef]
- Cho, C.; Yu, C.; Wu, C.; Ho, J.; Yang, C.; Yu, D. Decreased drug resistance of bladder cancer using phytochemicals treatment. Kaohsiung J. Med Sci. 2021, 37, 128–135. [Google Scholar] [CrossRef]
- Bock, K.W. Modulation of aryl hydrocarbon receptor (AHR) and the NAD+-consuming enzyme CD38: Searches of therapeutic options for nonalcoholic fatty liver disease (NAFLD). Biochem. Pharmacol. 2020, 175, 113905. [Google Scholar] [CrossRef]
- Bock, K.W. Aryl hydrocarbon receptor (AHR)-mediated inflammation and resolution: Non-genomic and genomic signaling. Biochem. Pharmacol. 2020, 182, 114220. [Google Scholar] [CrossRef]
- Choi, H.; Chun, Y.-S.; Shin, Y.J.; Ye, S.K.; Kim, M.-S.; Park, J.-W. Curcumin attenuates cytochrome P450 induction in response to 2,3,7,8-tetrachlorodibenzo-p-dioxin by ROS-dependently degrading AhR and ARNT. Cancer Sci. 2008, 99, 2518–2524. [Google Scholar] [CrossRef] [PubMed]
- Nishiumi, S.; Yoshida, K.-I.; Ashida, H. Curcumin suppresses the transformation of an aryl hydrocarbon receptor through its phosphorylation. Arch. Biochem. Biophys. 2007, 466, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Lai, S.; Li, Y.; Kuang, Y.; Cui, H.; Yang, Y.; Sun, W.; Liu, K.; Chen, N.; Yan, Q.; Wen, L. PKCδ silencing alleviates saturated fatty acid induced ER stress by enhancing SERCA activity. Biosci. Rep. 2017, 37, BSR20170869. [Google Scholar] [CrossRef] [PubMed]
- Cai, C.; Song, X.; Yu, C. Identification of genes in hepatocellular carcinoma induced by non-alcoholic fatty liver disease. Cancer Biomarkers 2020, 29, 69–78. [Google Scholar] [CrossRef]
- Park, S.W.; Zhou, Y.; Lee, J.; Lee, J.; Ozcan, U. Sarco(endo)plasmic reticulum Ca 2+ -ATPase 2b is a major regulator of endoplasmic reticulum stress and glucose homeostasis in obesity. Proc. Natl. Acad. Sci. USA 2010, 107, 19320–19325. [Google Scholar] [CrossRef] [PubMed]
- Frangioudakis, G.; Burchfield, J.G.; Narasimhan, S.; Cooney, G.J.; Leitges, M.; Biden, T.J.; Schmitz-Peiffer, C. Diverse roles for protein kinase C δ and protein kinase C ε in the generation of high-fat-diet-induced glucose intolerance in mice: Regulation of lipogenesis by protein kinase C δ. Diabetologia 2009, 52, 2616–2620. [Google Scholar] [CrossRef]
- Bezy, O.; Tran, T.T.; Pihlajamäki, J.; Suzuki, R.; Emanuelli, B.; Winnay, J.; Mori, M.; Haas, J.; Biddinger, S.B.; Leitges, M.; et al. PKCδ regulates hepatic insulin sensitivity and hepatosteatosis in mice and humans. J. Clin. Investig. 2011, 121, 2504–2517. [Google Scholar] [CrossRef]
- Rushworth, S.A.; Ogborne, R.M.; Charalambos, C.A.; O’Connell, M.A. Role of protein kinase C δ in curcumin-induced antioxidant response element-mediated gene expression in human monocytes. Biochem. Biophys. Res. Commun. 2006, 341, 1007–1016. [Google Scholar] [CrossRef]
- Kunwar, A.; Narang, H.; Priyadarsini, K.I.; Krishna, M.; Pandey, R.; Sainis, K. Delayed activation of PKCδ and NFκB and higher radioprotection in splenic lymphocytes by copper (II)–Curcumin (1:1) complex as compared to curcumin. J. Cell. Biochem. 2007, 102, 1214–1224. [Google Scholar] [CrossRef]
- Hsu, H.-Y.; Chu, L.-C.; Hua, K.-F.; Chao, L.K. Heme oxygenase-1 mediates the anti-inflammatory effect of Curcumin within LPS-stimulated human monocytes. J. Cell. Physiol. 2008, 215, 603–612. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | UniProt | Gene_Full_Name | Protein Class | DSI g | Score Gda | STITCH | |||
|---|---|---|---|---|---|---|---|---|---|
| Score | Source | Action Type | Liver Tissue | ||||||
| CYP1A2 | P05177 | cytochrome P450 family 1 subfamily A member 2 | Enzyme | 0.49 | 0.30 | 0.94 | Curated databases | Inhibition | 4.78 |
| NFE2L2 | Q16236 | nuclear factor, erythroid 2 like 2 | Enzyme | 0.35 | 0.40 | 0.87 | Textmining | Activation | 3.65 |
| PPARA | Q07869 | peroxisome proliferator-activated receptor alpha | Nuclear receptor | 0.43 | 0.40 | 0.86 | Curated databases | Activation | 4.75 |
| GSTA1 | P08263 | glutathione S-transferase alpha 1 | Enzymes, Plasma proteins | 0.56 | 0.31 | 0.84 | Textmining | Activation/ Inhibition | 4.90 |
| IL1A | P01583 | interleukin 1 alpha | Plasma proteins, Transporters | 0.33 | 0.33 | 0.82 | Textmining | Inhibition | 2.77 |
| LEP | P41159 | leptin | Plasma protein | 0.34 | 0.40 | 0.82 | Textmining | Activation/Inhibition | 3.17 |
| LDLR | P01130 | low density lipoprotein receptor | Plasma protein | 0.44 | 0.37 | 0.82 | Textmining | Activation/Inhibition, Transcriptional regulator | 4.40 |
| CSF2 | P04141 | colony-stimulating factor 2 | Signaling | 0.33 | 0.30 | 0.81 | Textmining | Inhibition, Transcriptional regulator | 2.60 |
| GSTP1 | P09211 | glutathione S-transferase pi 1 | Enzyme | 0.38 | 0.33 | 0.79 | Textmining | Activation | 4.64 |
| PRKCE | Q02156 | protein kinase C epsilon | Kinase | 0.59 | 0.31 | 0.76 | Curated databases | Binding | 2.34 |
| TGFB1 | P01137 | transforming growth factor-beta 1 | Signaling | 0.28 | 0.34 | 0.73 | Curated databases | Inhibition, Transcriptional regulator | 3.01 |
| ABCC2 | Q92887 | ATP binding cassette subfamily C member 2 | Transporter | 0.48 | 0.32 | 0.72 | Curated databases | Inhibition | 4.78 |
| AHR | P35869 | aryl hydrocarbon receptor | Transcription factor | 0.41 | 0.33 | 0.70 | Textmining | Unspecific | 4.39 |
| PRKCD | Q05655 | protein kinase C delta | Kinase | 0.48 | 0.31 | 0.70 | Curated databases | Unspecific | 4.49 |
| Biological Process (GO) | ||||
| Accession | Pathway Description | Gene Count | p-Value | FDR |
| GO: 1901700 | response to oxygen-containing compound | 13 | 3.93 × 10−14 | 6.17 × 10−10 |
| GO: 0071396 | cellular response to lipid | 10 | 5.70 × 10−14 | 4.48 × 10−10 |
| GO: 0033993 | response to lipid | 11 | 1.57 × 10−13 | 8.25 × 10−10 |
| GO:0006950 | response to stress | 12 | 2.19 × 10−0.08 | 1.07 × 10−0.05 |
| GO:0019216 | regulation of lipid metabolic process | 6 | 4.75 × 10−0.08 | 2.07 × 10−0.05 |
| Molecular Function (GO) | ||||
| Pathway Description | Gene Count | p-Value | FDR | |
| GO:0004699 | calcium-independent protein kinase C activity | 2 | 2.57 × 10−0.06 | 1.25 × 10−0.02 |
| GO:0030546 | signaling receptor activator activity | 5 | 1.43 × 10−0.05 | 3.49 × 10−0.02 |
| GO: 0001221 | transcription coregulator binding | 3 | 2.22 × 10−0.05 | 3.61 × 10−0.02 |
| GO: 0030545 | signaling receptor regulator activity | 5 | 2.24 × 10−0.05 | 2.73 × 10−0.02 |
| Cellular Component (GO) | ||||
| Pathway Description | Gene Count | p-Value | FDR | |
| GO: 0036019 | endolysosome | 2 | 1.72 × 10−0.04 | 3.42 × 10−0.01 |
| GO: 0005615 | extracellular space | 8 | 1.11 × 10−0.06 | 1.10 × 10−0.03 |
| GO: 0034774 | secretory granule lumen | 3 | 1.23 × 10−0.03 | 8.16 × 10−0.01 |
| KEGG | ||||
|---|---|---|---|---|
| NUM | Pathway Name | Genes | Gene Count | p-Value |
| 1 | Chemical carcinogenesis | GSTP1, CYP1A2, GSTA1, AHR, PPARA | 5 | 4.285 × 10−7 |
| 2 | AGE-RAGE signaling pathway in diabetic complications | IL1A, TGFB1, PRKCE, PRKCD | 4 | 5.667 × 10−7 |
| 3 | Fluid shear stress and atherosclerosis | NFE2L2, IL1A, GSTP1, GSTA1 | 4 | 2.119 × 10−6 |
| 4 | Non-alcoholic fatty liver disease | IL1A, PPARA, TGFB1, LEP | 3 | 3.270 × 10−6 |
| 5 | Cytokine-cytokine receptor interaction | IL1A, CSF2, TGFB1, LEP | 4 | 5.667 × 10−7 |
| Disease–Gene Associations Based on EnrichR Algorithm | ||||
|---|---|---|---|---|
| Jensen DISEASES | DisGeNET | |||
| NUM | Name | p-Value | Name | p-Value |
| 1 | Arthritis | 6.740 × 10−6 | Non-alcoholic Fatty Liver Disease | 3.411 × 10−20 |
| 2 | Coronary artery disease | 9.900 × 10−6 | Nonalcoholic Steatohepatitis | 1.206 × 10−17 |
| 3 | Fatty liver disease | 2.607 × 10−5 | Hypertensive disease | 2.505 × 10−17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahmoudi, A.; Butler, A.E.; Majeed, M.; Banach, M.; Sahebkar, A. Investigation of the Effect of Curcumin on Protein Targets in NAFLD Using Bioinformatic Analysis. Nutrients 2022, 14, 1331. https://doi.org/10.3390/nu14071331
Mahmoudi A, Butler AE, Majeed M, Banach M, Sahebkar A. Investigation of the Effect of Curcumin on Protein Targets in NAFLD Using Bioinformatic Analysis. Nutrients. 2022; 14(7):1331. https://doi.org/10.3390/nu14071331
Chicago/Turabian StyleMahmoudi, Ali, Alexandra E. Butler, Muhammed Majeed, Maciej Banach, and Amirhossein Sahebkar. 2022. "Investigation of the Effect of Curcumin on Protein Targets in NAFLD Using Bioinformatic Analysis" Nutrients 14, no. 7: 1331. https://doi.org/10.3390/nu14071331
APA StyleMahmoudi, A., Butler, A. E., Majeed, M., Banach, M., & Sahebkar, A. (2022). Investigation of the Effect of Curcumin on Protein Targets in NAFLD Using Bioinformatic Analysis. Nutrients, 14(7), 1331. https://doi.org/10.3390/nu14071331