
Microbial and Chemical Profiles of Commercial Kombucha Products

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Kombucha Samples
2.2. Levels of Tea Catechins and Caffeine Analyzed by High Performance Liquid Chromatography (HPLC)
2.3. Gallic Acid Equivalent (GAE) Assay
2.4. Trolox Equivalent Antioxidant Capacity (TEAC) Assay
2.5. Ethanol Content ANALYZED by Gas Chromatography (GC)
2.6. pH Measurements
2.7. Shotgun Metagenomics Analysis
2.8. ITS Sequencing
2.9. Metabolomics
2.10. Statistical Analysis
3. Results
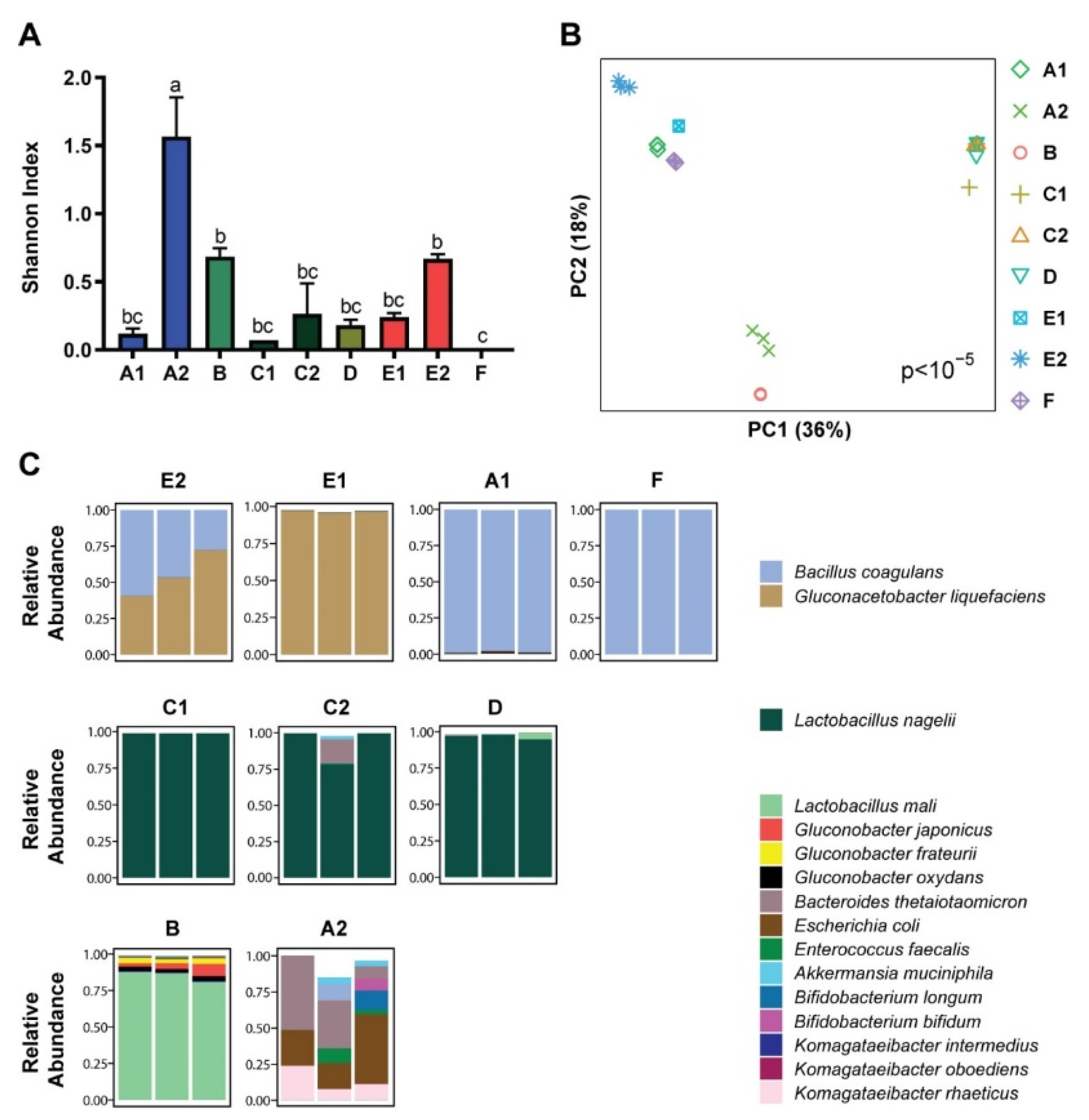
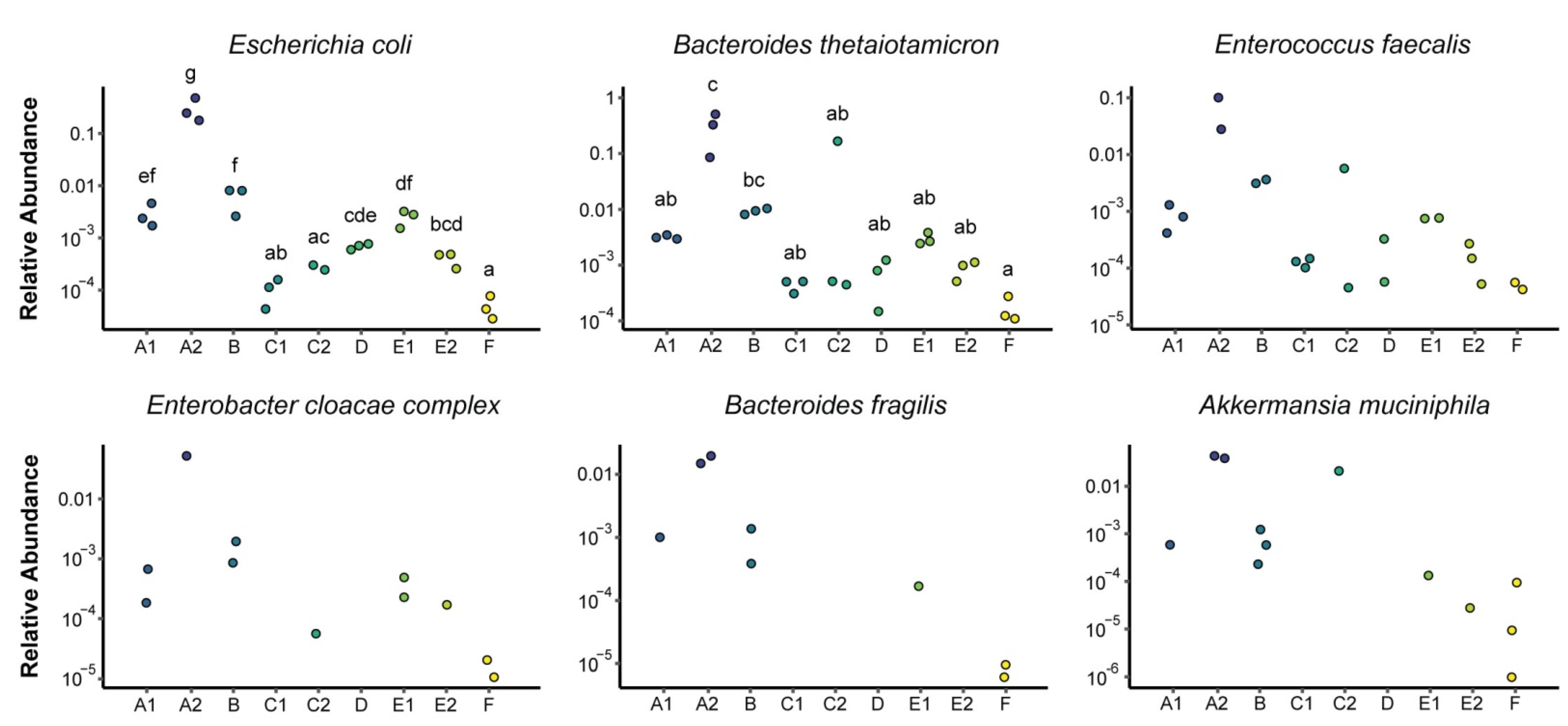
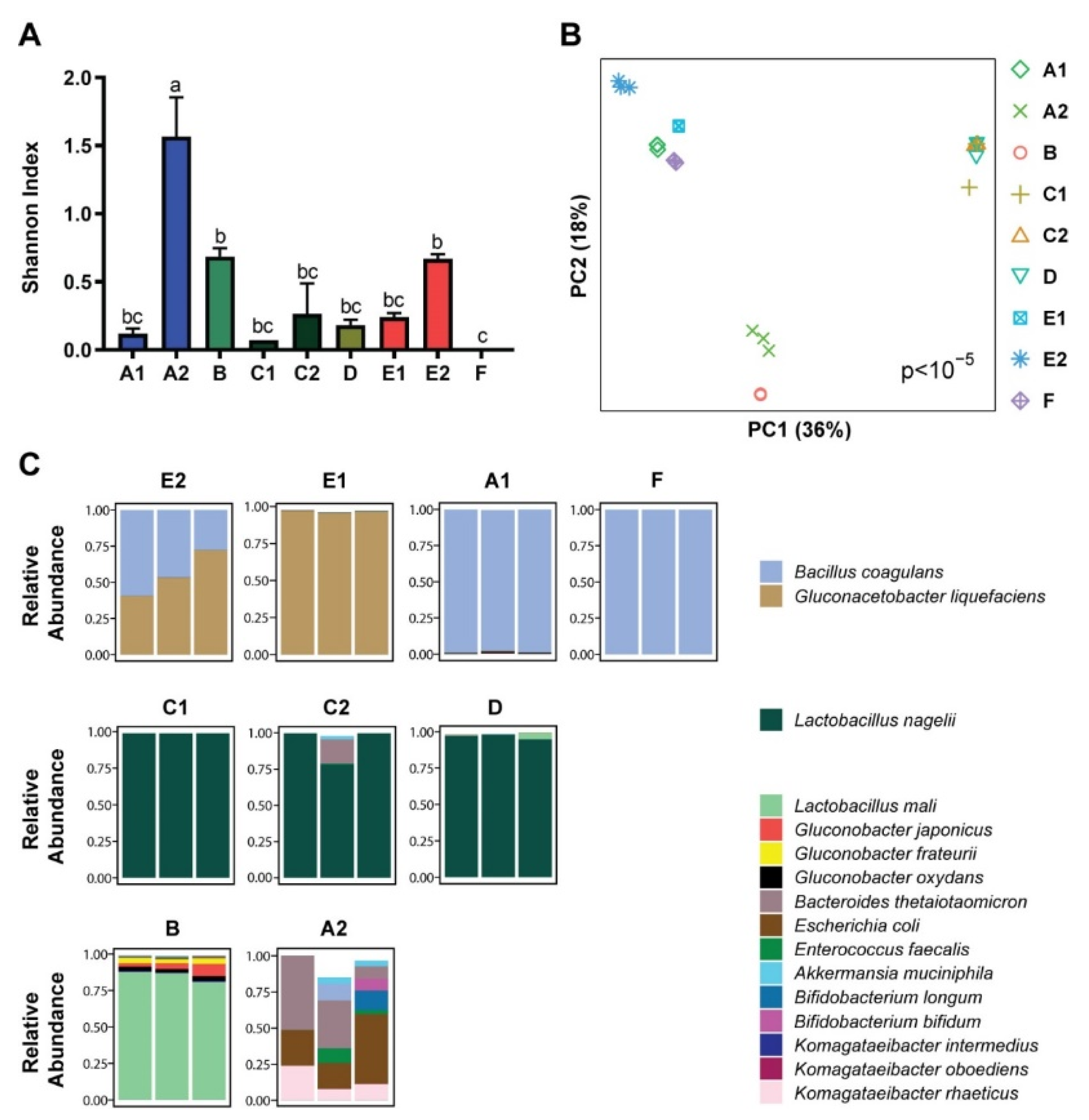
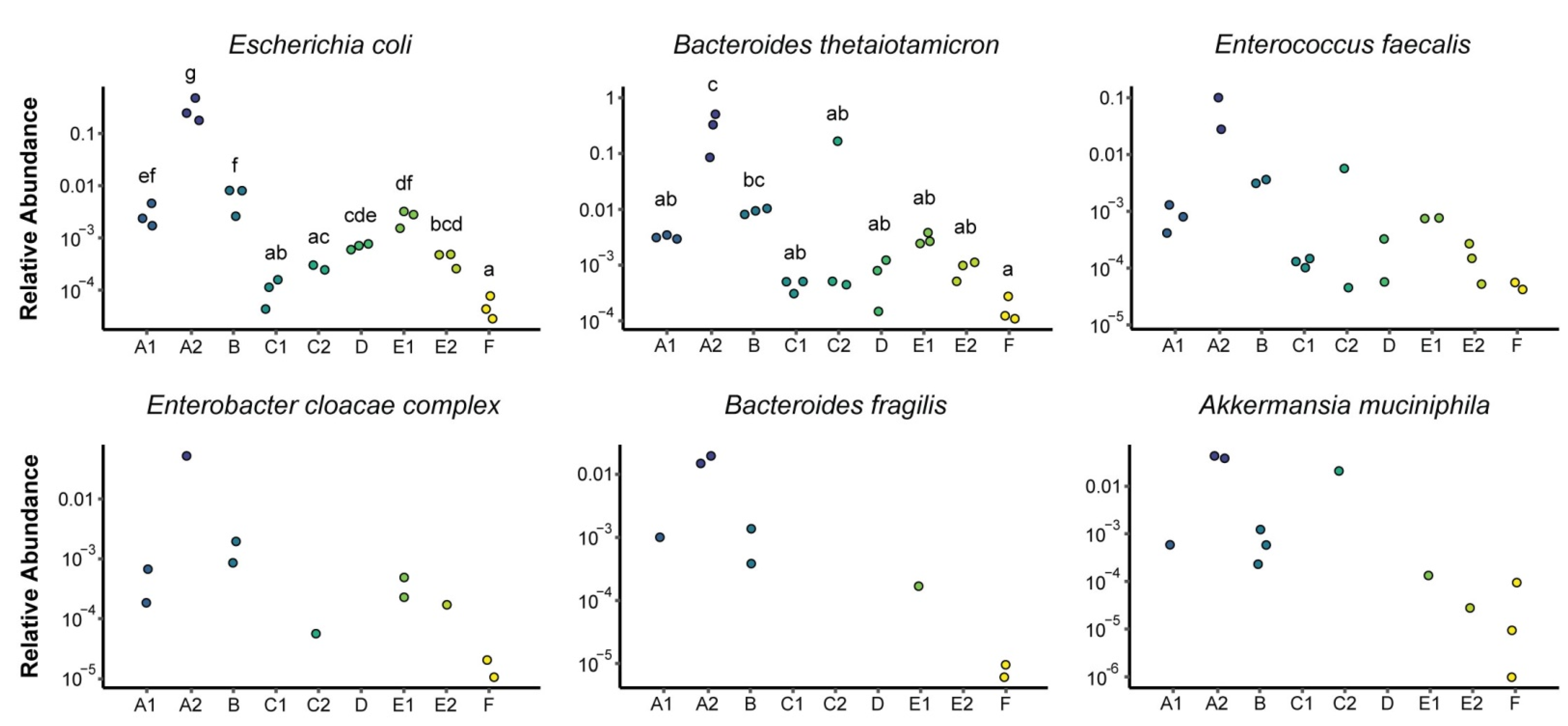
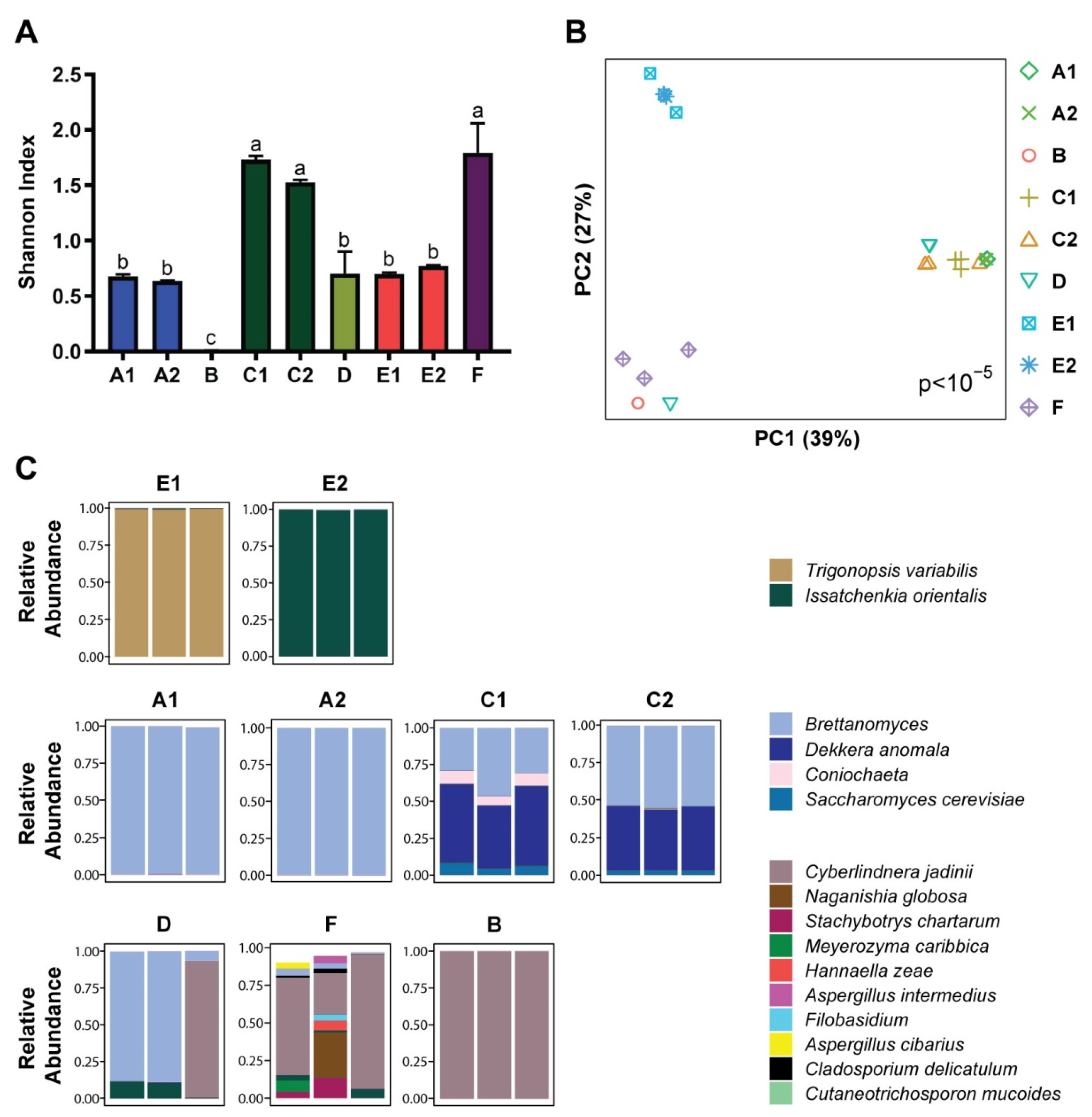
3.1. Microbial Composition
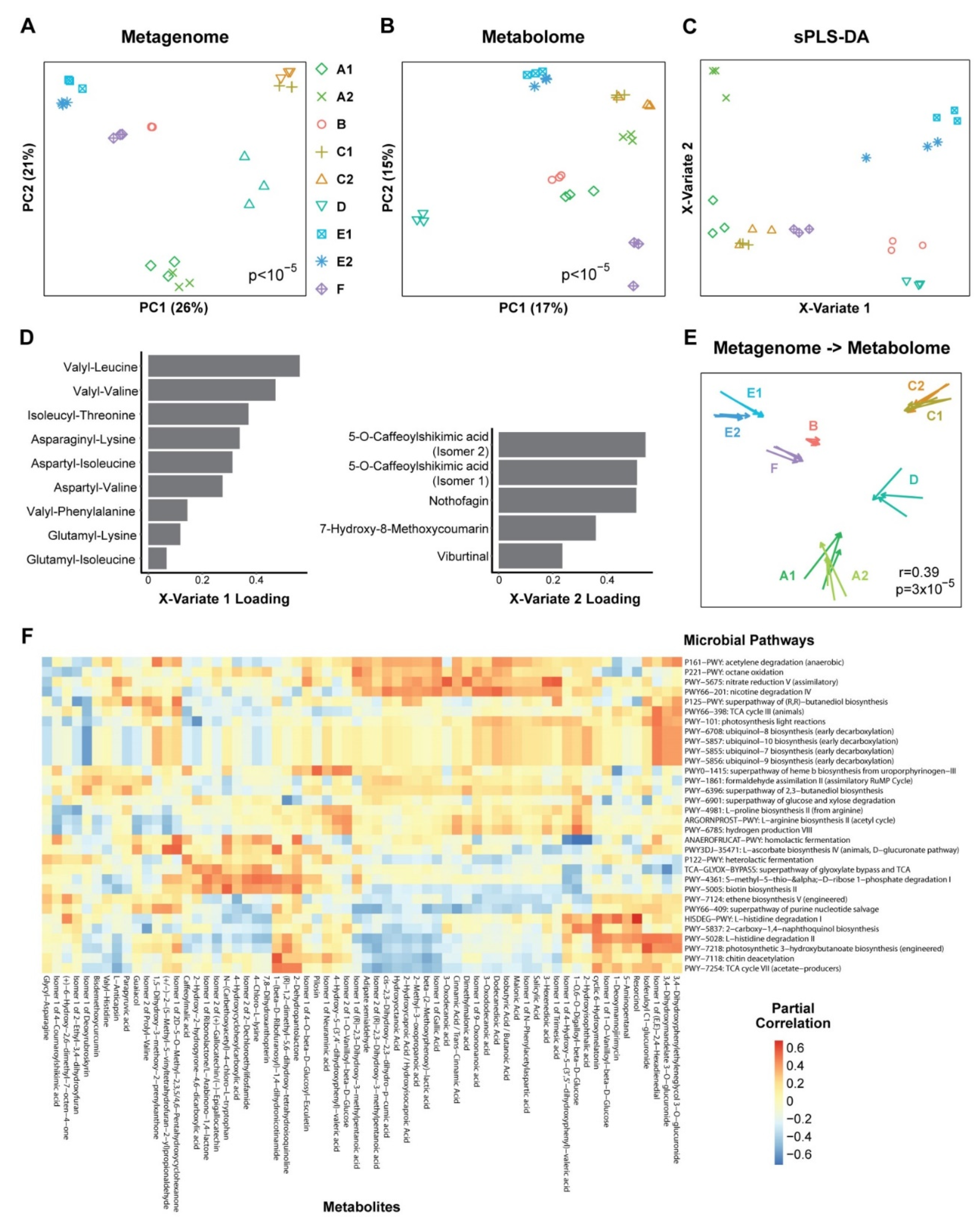
3.2. Metabolomics and Metagenomics Profiles
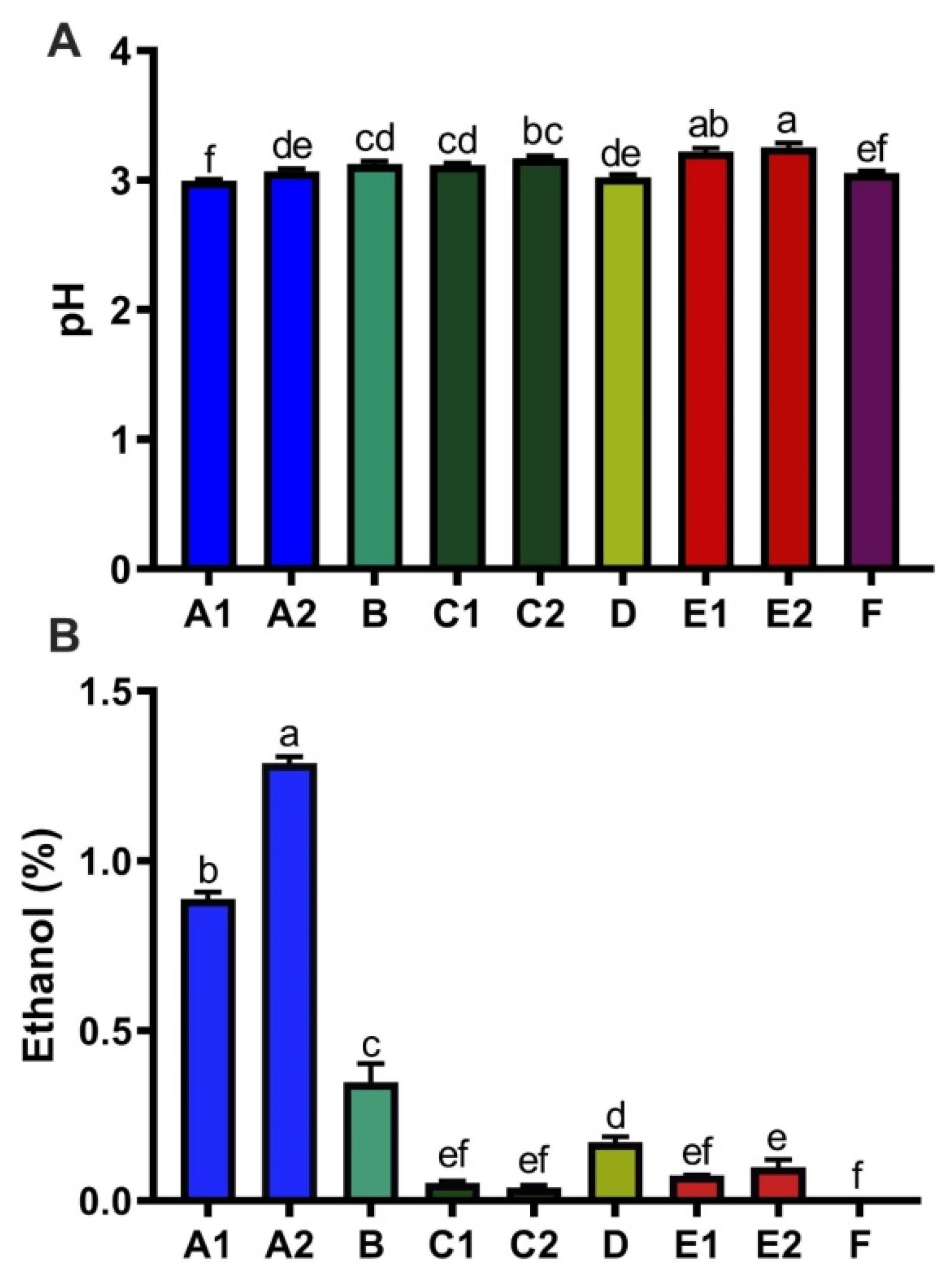
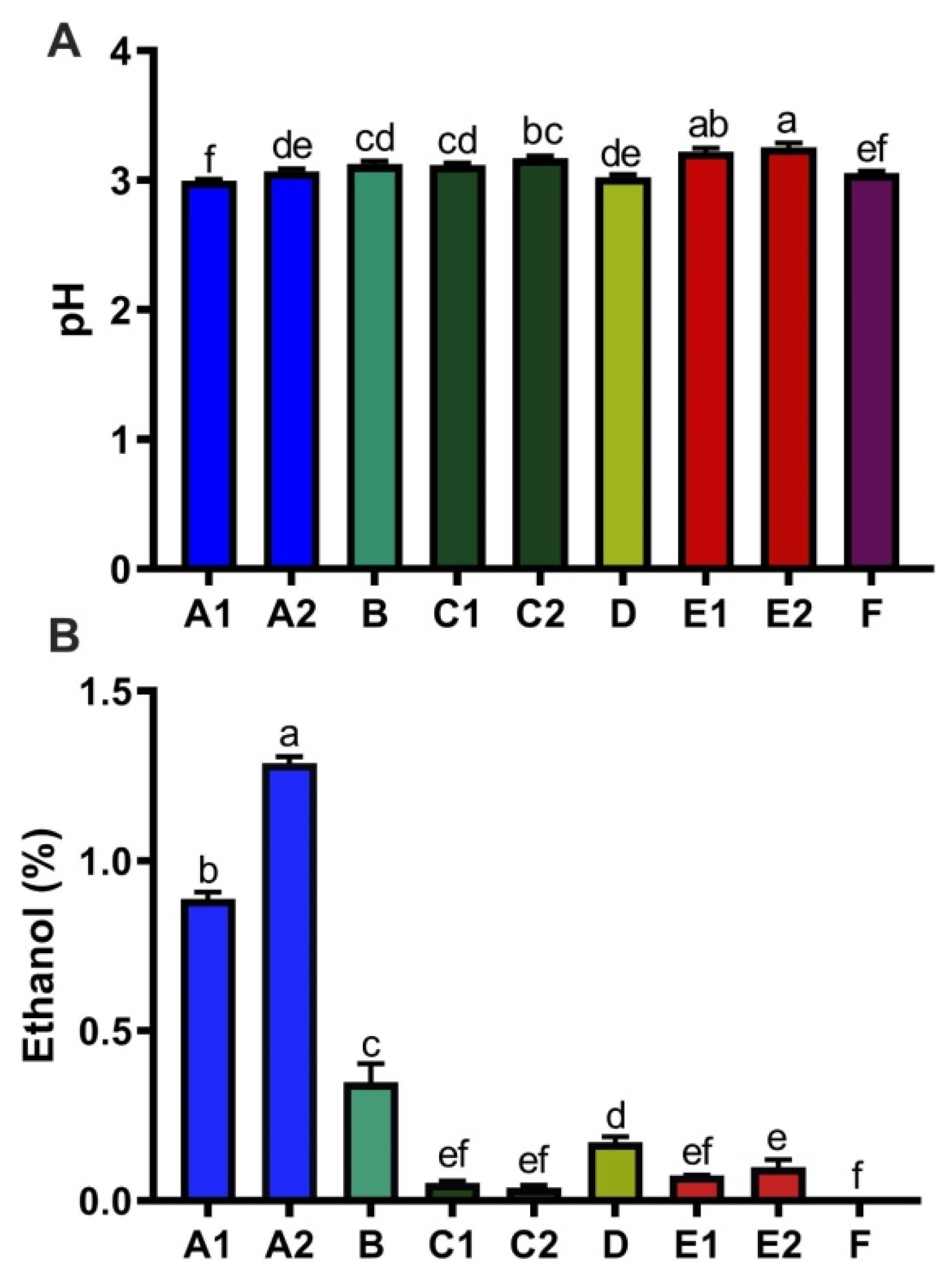
3.3. Ethanol, Acidity, and Caffeine Content of Kombucha
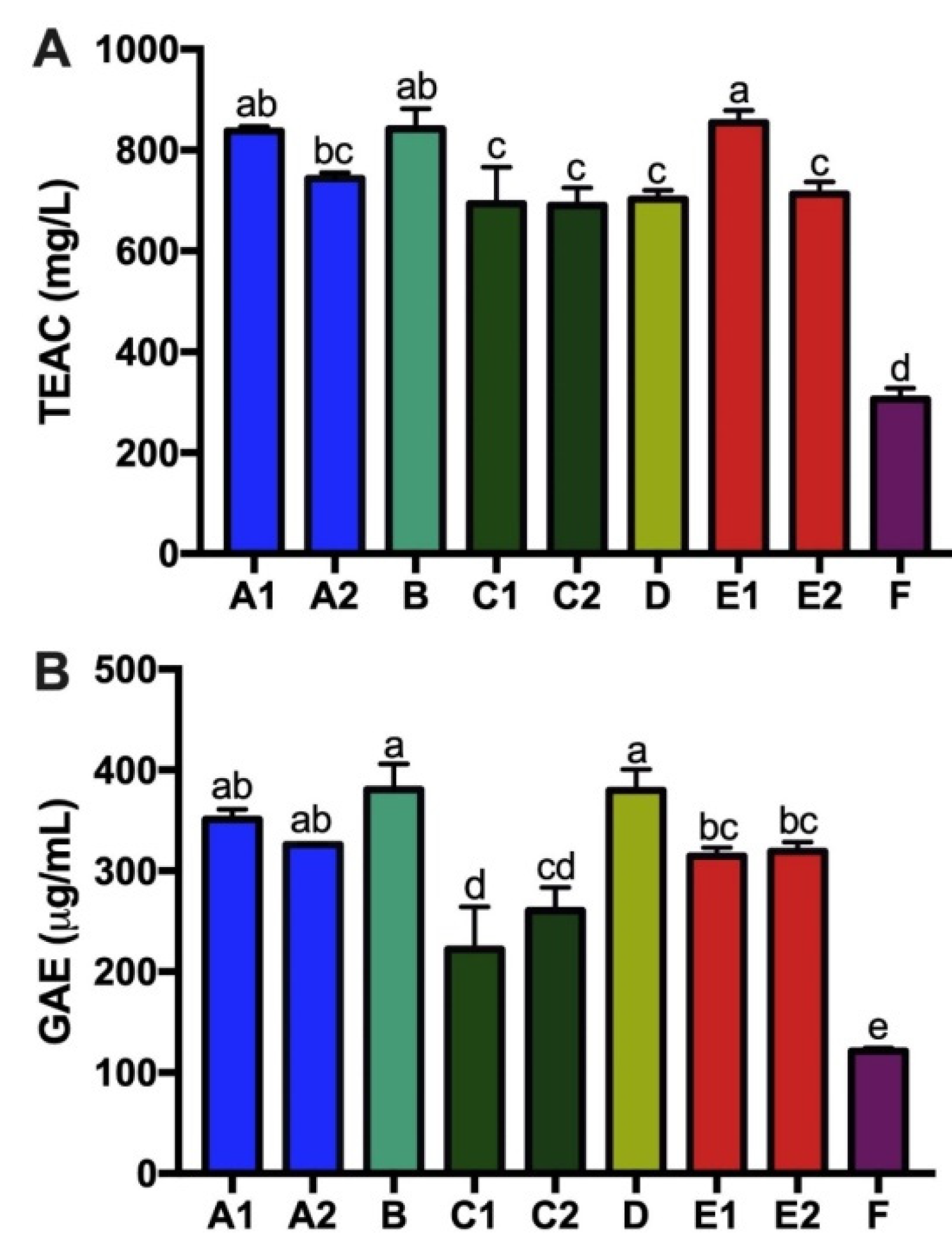
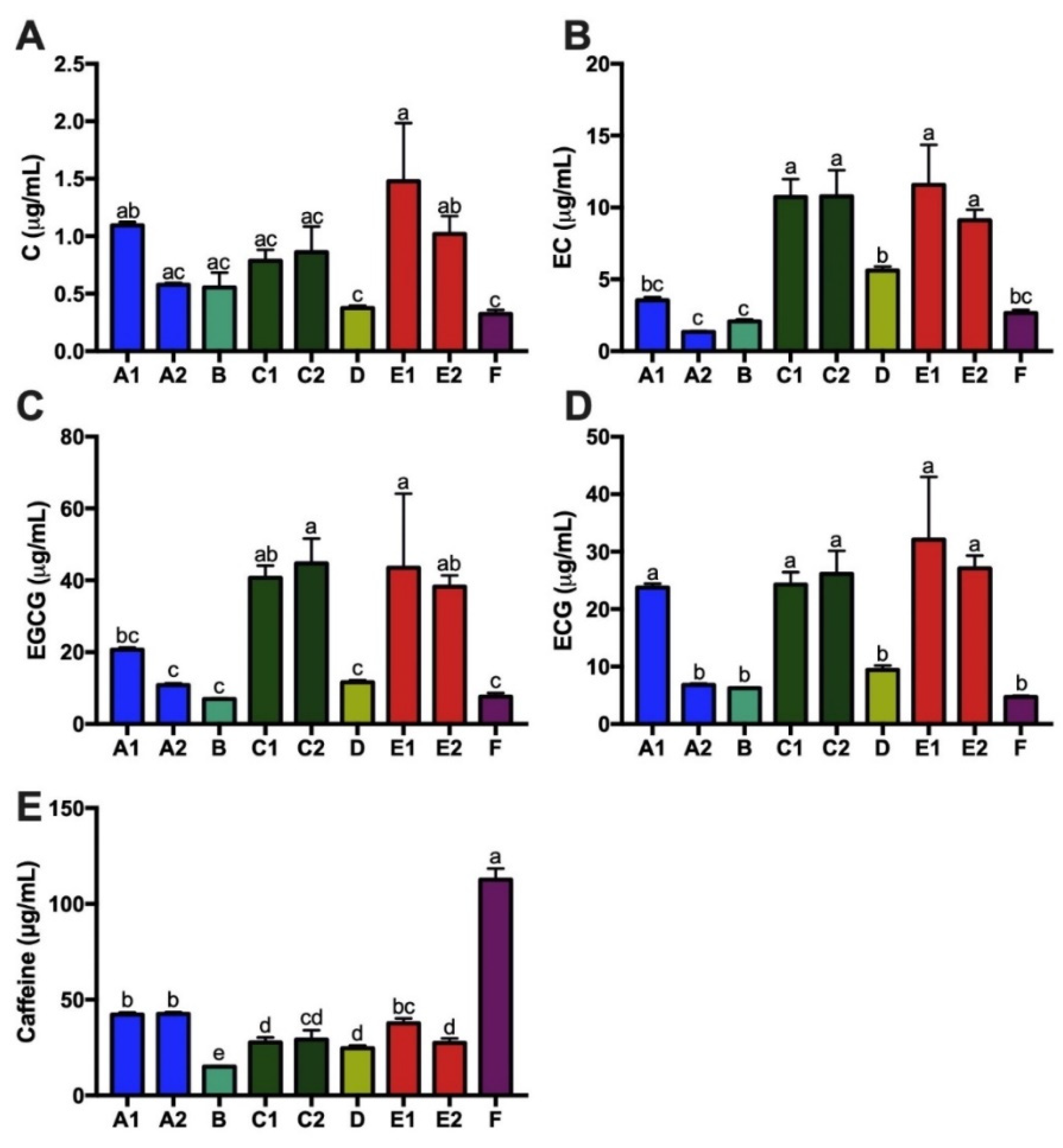
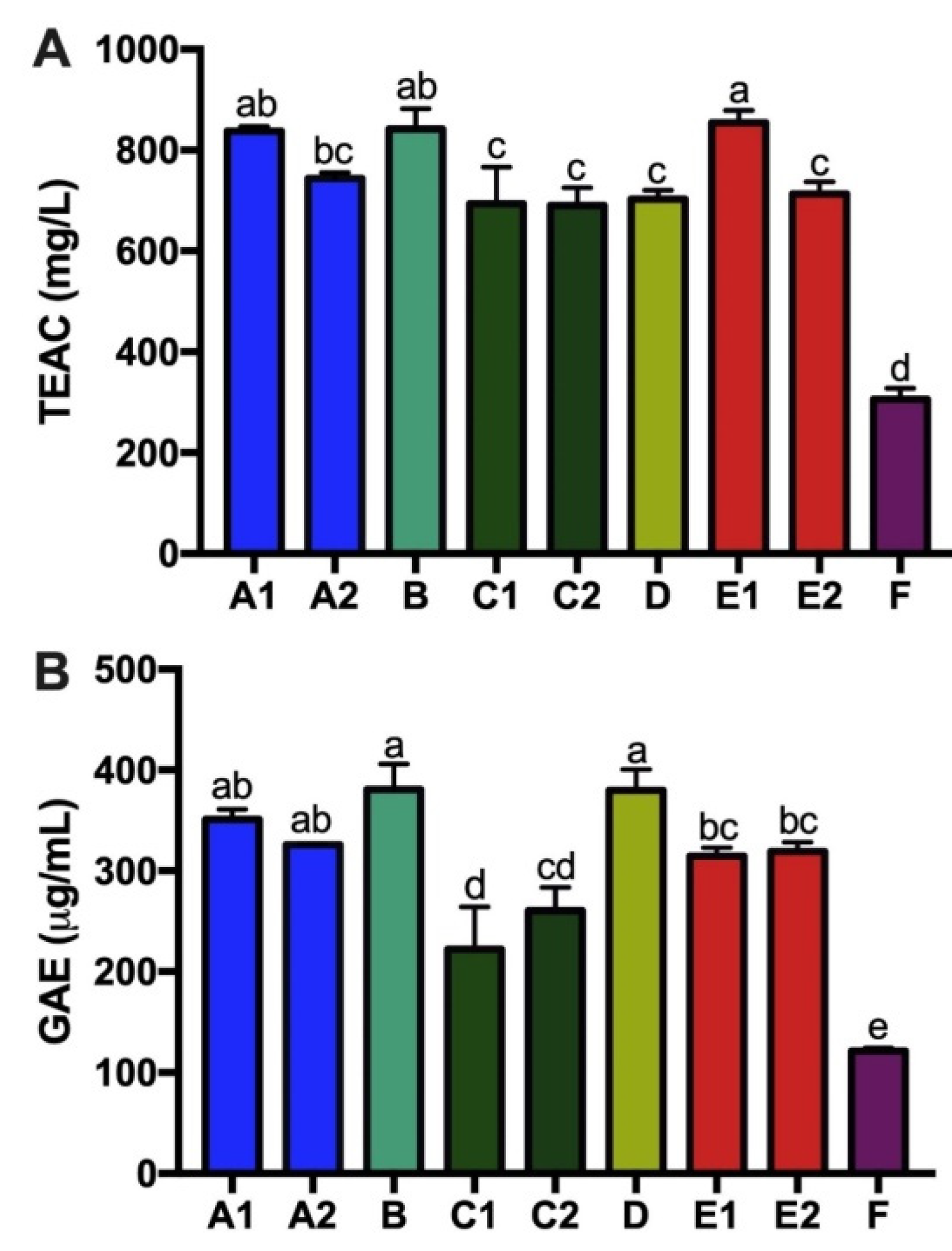
3.4. Tea Catechins, Antioxidant Capacity and Total Tea Polyphenols of Kombucha Products
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kim, J.; Adhikari, K. Current Trends in Kombucha: Marketing Perspectives and the Need for Improved Sensory Research. Beverages 2020, 6, 15. [Google Scholar] [CrossRef] [Green Version]
- Chakravorty, S.; Bhattacharya, S.; Chatzinotas, A.; Chakraborty, W.; Bhattacharya, D.; Gachhui, R. Kombucha tea fermentation: Microbial and biochemical dynamics. Int. J. Food Microbiol. 2016, 220, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Jayabalan, R.; Malbaša, R.V.; Lončar, E.S.; Vitas, J.S.; Sathishkumar, M. A Review on Kombucha Tea—Microbiology, Composition, Fermentation, Beneficial Effects, Toxicity, and Tea Fungus. Compr. Rev. Food Sci. Food Saf. 2014, 13, 538–550. [Google Scholar] [CrossRef]
- Vina, I.; Semjonovs, P.; Linde, R.; Denina, I. Current Evidence on Physiological Activity and Expected Health Effects of Kombucha Fermented Beverage. J. Med. Food 2014, 17, 179–188. [Google Scholar] [CrossRef] [Green Version]
- Mousavi, S.M.; Hashemi, S.A.; Zarei, M.; Gholami, A.; Lai, C.W.; Chiang, W.H.; Omidifar, N.; Bahrani, S.; Mazraedoost, S. Recent Progress in Chemical Composition, Production, and Pharmaceutical Effects of Kombucha Beverage: A Complementary and Alternative Medicine. Evid.-Based Complement. Altern. Med. 2020, 2020, 4397543. [Google Scholar] [CrossRef]
- Yang, D.-J.; Hwang, L.S.; Lin, J.-T. Effects of different steeping methods and storage on caffeine, catechins and gallic acid in bag tea infusions. J. Chromatogr. A 2007, 1156, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.S.; Tsai, Y.J.; Tsay, J.S.; Lin, J.K. Factors affecting the levels of tea polyphenols and caffeine in tea leaves. J. Agric. Food Chem. 2003, 51, 1864–1873. [Google Scholar] [CrossRef]
- Graham, H.N. Green tea composition, consumption, and polyphenol chemistry. Prev. Med. 1992, 21, 334–350. [Google Scholar] [CrossRef]
- Khan, N.; Mukhtar, H. Tea polyphenols for health promotion. Life Sci. 2007, 81, 519–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, N.; Mukhtar, H. Tea Polyphenols in Promotion of Human Health. Nutrients 2018, 11, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dufresne, C.; Farnworth, E. Tea, Kombucha, and health: A review. Food Res. Int. 2000, 33, 409–421. [Google Scholar] [CrossRef]
- Lee, K.W.; Lee, H.J.; Lee, C.Y. Antioxidant activity of black tea vs. green tea. J. Nutr. 2002, 132, 786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gramza-Michalowska, A. Caffeine in Tea Camellia Sinensis—Content, Absorption, Benefits and Risks of Consumption. J. Nutr. Health Aging 2014, 18, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Heber, D.; Zhang, Y.J.; Yang, J.P.; Ma, J.E.; Henning, S.M.; Li, Z.P. Green Tea, Black Tea, and Oolong Tea Polyphenols Reduce Visceral Fat and Inflammation in Mice Fed High-Fat, High-Sucrose Obesogenic Diets. J. Nutr. 2014, 144, 1385–1393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, E.W.C.; Soh, E.Y.; Tie, P.P.; Law, Y.P. Antioxidant and antibacterial properties of green, black, and herbal teas of Camellia sinensis. Pharm. Res. 2011, 3, 266–272. [Google Scholar] [CrossRef] [Green Version]
- Sengun, I.; Kirmizigul, A. Probiotic Potential of Kombucha. J. Clin. Gastroenterol. 2020, 54, S28. [Google Scholar]
- Vargas, B.K.; Fabricio, M.F.; Ayub, M.A.Z. Health effects and probiotic and prebiotic potential of Kombucha: A bibliometric and systematic review. Food Biosci. 2021, 44, 101332. [Google Scholar]
- Konuray, G.; Erginkaya, Z. Potential Use of Bacillus coagulans in the Food Industry. Foods 2018, 7, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanc, P.J. Characterization of the tea fungus metabolites. Biotechnol. Lett. 1996, 18, 139–142. [Google Scholar] [CrossRef]
- Villarreal-Soto, S.A.; Beaufort, S.; Bouajila, J.; Souchard, J.-P.; Taillandier, P. Understanding Kombucha Tea Fermentation: A Review. J. Food Sci. 2018, 83, 580–588. [Google Scholar] [CrossRef] [PubMed]
- SPINS TOTAL US $ SALES (MULO, CONVENIENCE, NATURAL) L52W ENDING 12.27.20. Available online: https://www.spins.com (accessed on 1 December 2021).
- Singleton, V.L.; Esau, P. Phenolic substances in grapes and wine, and their significance. Adv. Food Res. Suppl. 1969, 1, 1–261. [Google Scholar] [PubMed]
- Henning, S.M.; Zhang, Y.; Seeram, N.P.; Lee, R.P.; Wang, P.; Bowerman, S.; Heber, D. Antioxidant capacity and phytochemical content of herbs and spices in dry, fresh and blended herb paste form. Int. J. Food Sci. Nutr. 2011, 62, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Beghini, F.; McIver, L.J.; Blanco-Miguez, A.; Dubois, L.; Asnicar, F.; Maharjan, S.; Mailyan, A.; Manghi, P.; Scholz, M.; Thomas, A.M.; et al. Integrating taxonomic, functional, and strain-level profiling of diverse microbial communities with bioBakery 3. Elife 2021, 10, e65088. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Walters, W.; Hyde, E.R.; Berg-Lyons, D.; Ackermann, G.; Humphrey, G.; Parada, A.; Gilbert, J.A.; Jansson, J.K.; Caporaso, J.G.; Fuhrman, J.A.; et al. Improved Bacterial 16S rRNA Gene (V4 and V4-5) and Fungal Internal Transcribed Spacer Marker Gene Primers for Microbial Community Surveys. mSystems 2016, 1, e00009-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsson, R.H.; Larsson, K.H.; Taylor, A.F.S.; Bengtsson-Palme, J.; Jeppesen, T.S.; Schigel, D.; Kennedy, P.; Picard, K.; Glockner, F.O.; Tedersoo, L.; et al. The UNITE database for molecular identification of fungi: Handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res. 2019, 47, D259–D264. [Google Scholar] [CrossRef] [PubMed]
- Le Cao, K.A.; Boitard, S.; Besse, P. Sparse PLS discriminant analysis: Biologically relevant feature selection and graphical displays for multiclass problems. BMC Bioinform. 2011, 12, 253. [Google Scholar] [CrossRef] [Green Version]
- Xue, J.; Liu, P.; Guo, G.; Wang, W.; Zhang, J.; Wang, W.; Le, T.; Yin, J.; Ni, D.; Jiang, H. Profiling of dynamic changes in non-volatile metabolites of shaken black tea during the manufacturing process using targeted and non-targeted metabolomics analysis. LWT 2022, 156, 113010. [Google Scholar] [CrossRef]
- Joubert, E. HPLC quantification of the dihydrochalcones, aspalathin and nothofagin in rooibos tea (Aspalathus linearis) as affected by processing. Food Chem. 1996, 55, 403–411. [Google Scholar] [CrossRef]
- Godeau R., P.; Rossi J., C.; Fouraste, I. Methyl-4-formyl-7 cyclopenta(c)pyrane isolated by acid hydrolysis from Viburnum tinus. Phytochemistry 1977, 16, 604–605. [Google Scholar] [CrossRef]
- Barua, N.; Sharma, R.P.; Madhusudanan, K.P.; Thyagarajan, G.; Herz, W. Coumarins in Artemisia Caruifolia. Phytochemistry 1980, 9, 2. [Google Scholar]
- De Roos, J.; De Vuyst, L. Acetic acid bacteria in fermented foods and beverages. Curr. Opin. Biotechnol. 2018, 49, 115–119. [Google Scholar] [CrossRef] [PubMed]
- May, A.; Narayanan, S.; Alcock, J.; Varsani, A.; Maley, C.; Aktipis, A. Kombucha: A novel model system for cooperation and conflict in a complex multi-species microbial ecosystem. PeerJ 2019, 7, e7565. [Google Scholar] [CrossRef] [PubMed]
- Harrison, K.; Curtin, C. Microbial Composition of SCOBY Starter Cultures Used by Commercial Kombucha Brewers in North America. Microorganisms 2021, 9, 1060. [Google Scholar] [CrossRef]
- Nguyen, N.K.; Dong, N.T.; Nguyen, H.T.; Le, P.H. Lactic acid bacteria: Promising supplements for enhancing the biological activities of kombucha. Springerplus 2015, 4, 91. [Google Scholar] [CrossRef] [Green Version]
- Sreeramulu, G.; Zhu, Y.; Knol, W. Kombucha fermentation and its antimicrobial activity. J. Agric. Food Chem. 2000, 48, 2589–2594. [Google Scholar] [CrossRef]
- Bhattacharya, D.; Bhattacharya, S.; Patra, M.M.; Chakravorty, S.; Sarkar, S.; Chakraborty, W.; Koley, H.; Gachhui, R. Antibacterial Activity of Polyphenolic Fraction of Kombucha against Enteric Bacterial Pathogens. Curr. Microbiol. 2016, 73, 885–896. [Google Scholar] [CrossRef]
- Brewer, S.S.; Lowe, C.A.; Beuchat, L.R.; Ortega, Y.R. Survival of Salmonella and Shiga Toxin-Producing Escherichia coli and Changes in Indigenous Microbiota during Fermentation of Home-Brewed Kombucha. J. Food Prot. 2021, 84, 1366–1373. [Google Scholar] [CrossRef]
- Gaggìa, F.; Baffoni, L.; Galiano, M.; Nielsen, D.; Jakobsen, R.; Castro-Mejía, J.; Bosi, S.; Truzzi, F.; Musumeci, F.; Dinelli, G.; et al. Kombucha Beverage from Green, Black and Rooibos Teas: A Comparative Study Looking at Microbiology, Chemistry and Antioxidant Activity. Nutrients 2018, 11, 1. [Google Scholar] [CrossRef] [Green Version]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M.; et al. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: A proof-of-concept exploratory study. Nat. Med. 2019, 25, 1096–1103. [Google Scholar] [CrossRef]
- Mayser, P.; Fromme, S.; Leitzmann, C.; Grunder, K. The yeast spectrum of the ‘tea fungus Kombucha’. Mycoses 1995, 38, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Leung, L.K.; Su, Y.; Chen, R.; Zhang, Z.; Huang, Y.; Chen, Z.Y. Theaflavins in black tea and catechins in green tea are equally effective antioxidants. J. Nutr. 2001, 131, 2248–2251. [Google Scholar] [CrossRef] [PubMed]
- Dutta, H.; Paul, S.K. Kombucha Drink: Production, Quality, and Safety Aspects; Elsevier: Amsterdam, The Netherlands, 2019; pp. 259–288. [Google Scholar] [CrossRef]
- La Torre, C.; Fazio, A.; Caputo, P.; Plastina, P.; Caroleo, M.C.; Cannataro, R.; Cione, E. Effects of Long-Term Storage on Radical Scavenging Properties and Phenolic Content of Kombucha from Black Tea. Molecules 2021, 26, 5474. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Product | Ingredients Label | Nutrition (per 100 mL) | Expiration |
|---|---|---|---|
| A1 | Probiotics: kombucha culture, Bacillus Coagulans GBI-306086 (4 billion organisms), S. Boulardii (4 billion organisms), Lactobacillus bacteria (4 billion) Tea: black tea, green tea Flavors: kiwi juice, ginger juice | Calories: 10.6 Carbohydrate: 2.54 g, Added Sugar: 0 g Sodium: 2.11 mg Lactic acid: 21.1 mg, Acetic acid: 15.9 mg, Glucuronic acid: 296.0 mg, Gluconic acid: 137.4 mg | 5/6/2021 |
| A2 | Probiotics: kombucha culture, Lactobacillus bacteria (4 billion organisms), S. Boulardii (4 billion organisms) Tea: black tea, green tea Sugar: cane sugar | Calories: 12.5 Carbohydrate: 3.33 g, Added Sugar: 2.08 g Sodium: 2.08 g Lactic acid: 20.83 mg, Acetic acid: 15.6 mg, Glucuronic acid: 291.7 mg, Gluconic acid: 135.4 mg Polyphenols: 2.1 mg | 4/14/2021 |
| B | Probiotics: kombucha culture Tea: black tea Sugar: cane sugar Flavors: ginger, lemongrass, orange peel, spearmint, peppermint | Calories: 12.7 Carbohydrate: 2.56 g, Added Sugar: 1.83 g Sodium: 3.67 mg | 8/12/2021 |
| C1 | Probiotics: kombucha culture Tea: green tea Sugar: cane sugar Flavors: dried lemon, lemon verbena, ginger juice, ginger extract | Calories: 16.9 Carbohydrate: 4.10 g, Added Sugar: 3.14 g Sodium: 0 mg | 5/22/2021 |
| C2 | Probiotics: kombucha culture Tea: green tea Sugar: cane sugar Flavors: dried lemon, lemon verbena, ginger juice, ginger extract | Calories: 16.9 Carbohydrate: 3.94 g, Added Sugar: 3.38 g Sodium: 0 mg | 3/9/2021 |
| D | Probiotics: kombucha culture Sugar: cane sugar Flavors: ginger juice, black tea, green tea, lemon juice | Calories: 14.8 Carbohydrate: 2.96 g, Added Sugar: 1.69 g Sodium: 0 mg | 7/21/2021 |
| E1 | Probiotics: kombucha culture, Bacillus subtilis, vitamin B12 Tea: green tea, black tea Sugar: cane sugar Flavors: white grape juice, apple juice, ginger juice, lemon juice | Calories: 9.6 Carbohydrate: 2.41 g, Added Sugar: 0 g Sodium: 0 mg | 8/4/2021 |
| E2 | Probiotics: kombucha culture, Bacillus subtilis, Vitamin B12, Bacillus coagulans Tea: green tea, black tea Flavors: white grape juice, cane sugar, allulose syrup, monk fruit, ginger juice, natural flavors | Calories: 3.03 Carbohydrate: 3.03 g Added Sugar: 0 g Sodium: 0 mg | 8/24/2021 |
| F | Probiotics: kombucha culture, Bacillus coagulans MTCC 5856 Tea: black tea, green tea, black tea extract, green tea extract, black tea essence, green coffee bean extract (caffeine) Sugar: cane sugar Flavors: ginger extract, stevia leaf extract, sparkling water, natural flavor | Calories: 13.3 Carbohydrate: 3.56 g Added Sugar: 3.33 g Sodium: 3.33 mg Gluconic and acetic acid: 112.7 mg | 6/3/2021 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Lagishetty, V.; Kurnia, P.; Henning, S.M.; Ahdoot, A.I.; Jacobs, J.P. Microbial and Chemical Profiles of Commercial Kombucha Products. Nutrients 2022, 14, 670. https://doi.org/10.3390/nu14030670
Yang J, Lagishetty V, Kurnia P, Henning SM, Ahdoot AI, Jacobs JP. Microbial and Chemical Profiles of Commercial Kombucha Products. Nutrients. 2022; 14(3):670. https://doi.org/10.3390/nu14030670
Chicago/Turabian StyleYang, Jieping, Venu Lagishetty, Patrick Kurnia, Susanne M. Henning, Aaron I. Ahdoot, and Jonathan P. Jacobs. 2022. "Microbial and Chemical Profiles of Commercial Kombucha Products" Nutrients 14, no. 3: 670. https://doi.org/10.3390/nu14030670
APA StyleYang, J., Lagishetty, V., Kurnia, P., Henning, S. M., Ahdoot, A. I., & Jacobs, J. P. (2022). Microbial and Chemical Profiles of Commercial Kombucha Products. Nutrients, 14(3), 670. https://doi.org/10.3390/nu14030670