
Resveratrol and SIRT1: Antiaging Cornerstones for Oocytes?



{kind=link}
{kind=link}
Abstract
1. Introduction
2. Interaction between SIRT1 and Resveratrol
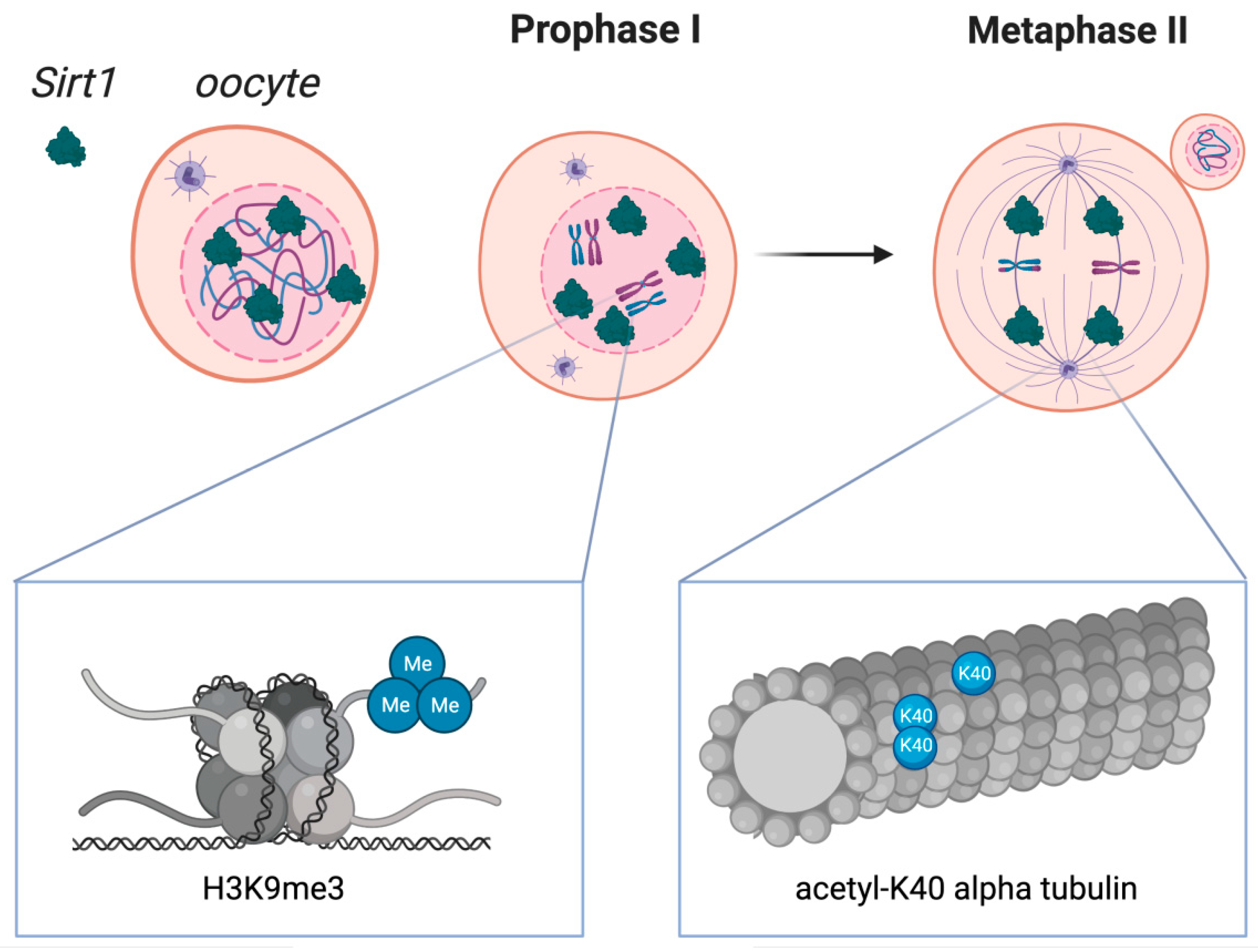
3. SIRT1 and Quality and Competence of Oocytes
4. SIRT1 and Aging
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mastenbroek, S.; de Wert, G.; Adashi, E.Y. The Imperative of Responsible Innovation in Reproductive Medicine. N. Engl. J. Med. 2021, 385, 2096–2100. [Google Scholar] [CrossRef] [PubMed]
- van der Reest, J.; Nardini Cecchino, G.; Haigis, M.C.; Kordowitzki, P. Mitochondria: Their Relevance during Oocyte Ageing. Ageing Res. Rev. 2021, 70, 101378. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, N.; Sato, Y.; Kawagoe, Y.; Shimizu, T.; Kawamura, K. Short-Term Resveratrol Treatment Restored the Quality of Oocytes in Aging Mice. Aging 2022, 14, 5628. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, R.; Caponnetto, A.; Caringella, A.M.; Cortone, A.; Ferrara, C.; Smirni, S.; Iannitti, R.; Purrello, M.; D’Amato, G.; Fioretti, B. Resveratrol Treatment Induces Mito-MiRNome Modification in Follicular Fluid from Aged Women with a Poor Prognosis for In Vitro Fertilization Cycles. Antioxidants 2022, 11, 1019. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Dudley, J.I.; Das, D.K. Dose-Dependency of Resveratrol in Providing Health Benefits. Dose Response 2010, 8, 478–500. [Google Scholar] [CrossRef]
- Posadino, A.M.; Cossu, A.; Giordo, R.; Zinellu, A.; Sotgia, S.; Vardeu, A.; Hoa, P.T.; Nguyen, L.H.V.; Carru, C.; Pintus, G. Resveratrol Alters Human Endothelial Cells Redox State and Causes Mitochondrial-Dependent Cell Death. Food Chem. Toxicol. 2015, 78, 10–16. [Google Scholar] [CrossRef]
- Singh, C.K.; Chhabra, G.; Ndiaye, M.A.; Garcia-Peterson, L.M.; Mack, N.J.; Ahmad, N. The Role of Sirtuins in Antioxidant and Redox Signaling. Antioxid. Redox Signal. 2018, 28, 643–661. [Google Scholar] [CrossRef]
- Jia, B.-Y.; Xiang, D.-C.; Shao, Q.-Y.; Zhang, B.; Liu, S.-N.; Hong, Q.-H.; Quan, G.-B.; Wu, G.-Q. Inhibitory Effects of Astaxanthin on Postovulatory Porcine Oocyte Aging in Vitro. Sci. Rep. 2020, 10, 20217. [Google Scholar] [CrossRef]
- Martín-Ramírez, R.; González-Fernández, R.; Hernández, J.; Martín-Vasallo, P.; Palumbo, A.; Ávila, J. Celastrol and Melatonin Modify SIRT1, SIRT6 and SIRT7 Gene Expression and Improve the Response of Human Granulosa-Lutein Cells to Oxidative Stress. Antioxidants 2021, 10, 1871. [Google Scholar] [CrossRef]
- Li, R.; Li, E.; Kamili, G.; Ou, S.; Yang, D. Effect of Resveratrol on Superovulation in Mice. Biomed. Pharmacother. 2022, 146, 112565. [Google Scholar] [CrossRef]
- Nie, J.; Sui, L.; Zhang, H.; Zhang, H.; Yan, K.; Yang, X.; Lu, S.; Lu, K.; Liang, X. Mogroside V Protects Porcine Oocytes from in Vitro Ageing by Reducing Oxidative Stress through SIRT1 Upregulation. Aging 2019, 11, 8362. [Google Scholar] [CrossRef] [PubMed]
- Di Emidio, G.; Falone, S.; Artini, P.G.; Amicarelli, F.; D’Alessandro, A.M.; Tatone, C. Mitochondrial Sirtuins in Reproduction. Antioxidants 2021, 10, 1047. [Google Scholar] [CrossRef] [PubMed]
- Wasielak-Politowska, M.; Kordowitzki, P. Chromosome Segregation in the Oocyte: What Goes Wrong during Aging. Int. J. Mol. Sci. 2022, 23, 2880. [Google Scholar] [CrossRef]
- Dvoran, M.; Nemcova, L.; Kalous, J. An Interplay between Epigenetics and Translation in Oocyte Maturation and Embryo Development: Assisted Reproduction Perspective. Biomedicines 2022, 10, 1689. [Google Scholar] [CrossRef] [PubMed]
- Ou, X.; Lee, M.R.; Huang, X.; Messina-Graham, S.; Broxmeyer, H.E. SIRT1 Positively Regulates Autophagy and Mitochondria Function in Embryonic Stem Cells Under Oxidative Stress. Stem. Cells 2014, 32, 1183–1194. [Google Scholar] [CrossRef] [PubMed]
- Osum, M.; Serakinci, N. Impact of Circadian Disruption on Health; SIRT1 and Telomeres. DNA Repair 2020, 96, 102993. [Google Scholar] [CrossRef] [PubMed]
- Giannakou, M.E.; Partridge, L. The Interaction between FOXO and SIRT1: Tipping the Balance towards Survival. Trends Cell Biol. 2004, 14, 408–412. [Google Scholar] [CrossRef]
- Richardson, B. Impact of Aging on DNA Methylation. Ageing Res. Rev. 2003, 2, 245–261. [Google Scholar] [CrossRef]
- Davenport, A.M.; Huber, F.M.; Hoelz, A. Structural and Functional Analysis of Human SIRT1. J. Mol. Biol. 2014, 426, 526–541. [Google Scholar] [CrossRef]
- Dai, H.; Case, A.W.; Riera, T.V.; Considine, T.; Lee, J.E.; Hamuro, Y.; Zhao, H.; Jiang, Y.; Sweitzer, S.M.; Pietrak, B.; et al. Crystallographic Structure of a Small Molecule SIRT1 Activator-Enzyme Complex. Nat. Commun. 2015, 6, 7645. [Google Scholar] [CrossRef]
- Gertz, M.; Nguyen, G.T.T.; Fischer, F.; Suenkel, B.; Schlicker, C.; Fränzel, B.; Tomaschewski, J.; Aladini, F.; Becker, C.; Wolters, D. A Molecular Mechanism for Direct Sirtuin Activation by Resveratrol. PLoS ONE 2012, 7, e49761. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Yang, N. Multiscale Landscape of Molecular Mechanism of SIRT1 Activation by STACs. Phys. Chem. Chem. Phys. 2020, 22, 826–837. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zeng, S.X.; Zhang, Y.; Zhang, Y.; Ding, D.; Ye, Q.; Meroueh, S.O.; Lu, H. A Small Molecule Inauhzin Inhibits SIRT1 Activity and Suppresses Tumour Growth through Activation of P53. EMBO Mol. Med. 2012, 4, 298–312. [Google Scholar] [CrossRef]
- Zhao, X.; Allison, D.; Condon, B.; Zhang, F.; Gheyi, T.; Zhang, A.; Ashok, S.; Russell, M.; MacEwan, I.; Qian, Y.; et al. The 2.5 Å Crystal Structure of the SIRT1 Catalytic Domain Bound to Nicotinamide Adenine Dinucleotide (NAD+) and an Indole (EX527 Analogue) Reveals a Novel Mechanism of Histone Deacetylase Inhibition. J. Med. Chem. 2013, 56, 963–969. [Google Scholar] [CrossRef] [PubMed]
- Borra, M.T.; Smith, B.C.; Denu, J.M. Mechanism of Human SIRT1 Activation by Resveratrol. J. Biol. Chem. 2005, 280, 17187–17195. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.; Wang, M.; Qiu, X.; Liu, D.; Jiang, H.; Yang, N.; Xu, R.-M. Structural Basis for Allosteric, Substrate-Dependent Stimulation of SIRT1 Activity by Resveratrol. Genes Dev. 2015, 29, 1316–1325. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Lu, J.; Zhan, L.; Wang, M.; Shi, R.; Yuan, X.; Gao, X.; Liu, X.; Zang, J.; Liu, W. Resveratrol-Induced Sirt1 Phosphorylation by LKB1 Mediates Mitochondrial Metabolism. J. Biol. Chem. 2021, 297, 100929. [Google Scholar] [CrossRef]
- Sirotkin, A.V. Effects of Resveratrol on Female Reproduction: A Review. Phytother. Res. 2021, 35, 5502–5513. [Google Scholar] [CrossRef]
- Kratz, E.M.; Kokot, I.; Dymicka-Piekarska, V.; Piwowar, A. Sirtuins—The New Important Players in Women’s Gynecological Health. Antioxidants 2021, 10, 84. [Google Scholar] [CrossRef]
- Yao, L.-N.; Zhang, T.-F.; Lin, W.-Q.; Jiang, N.; Cao, H.-F.; Li, H.; Qian, J.-H. Value of Serum and Follicular Fluid Sirtuin (SIRT)1 and SIRT2 Protein Levels in Predicting the Outcome of Assisted Reproduction. Ann. Transl. Med. 2021, 9, 343. [Google Scholar] [CrossRef]
- Ferreira, A.F.; Machado-Simões, J.; Soares, M.; Sousa, A.P.; Ramalho-Santos, J.; Almeida-Santos, T. Spatiotemporal Dynamics of SIRT 1, 2 and 3 during in Vitro Maturation of Bovine Oocytes. Theriogenology 2022, 186, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Chen, B.; Cui, H.; Gao, H.; Gao, M.; Tao, C. Dynamic Alterations in H4K12 Acetylation during Meiotic Maturation and after Parthenogenetic Activation of Mouse Oocytes. Zygote 2020, 28, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Nevoral, J.; Landsmann, L.; Stiavnicka, M.; Hosek, P.; Moravec, J.; Prokesova, S.; Rimnacova, H.; Koutna, E.; Klein, P.; Hoskova, K. Epigenetic and Non-Epigenetic Mode of SIRT1 Action during Oocyte Meiosis Progression. J. Anim. Sci. Biotechnol. 2019, 10, 1–12. [Google Scholar] [CrossRef]
- Zhang, L.; Ma, R.; Hu, J.; Ding, X.; Xu, Y. Sirtuin Inhibition Adversely Affects Porcine Oocyte Meiosis. PLoS ONE 2015, 10, e0132941. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, H.; Hamatani, T.; Kamijo, S.; Iwai, M.; Kobanawa, M.; Ogawa, S.; Miyado, K.; Tanaka, M. Impact of Oxidative Stress on Age-Associated Decline in Oocyte Developmental Competence. Front. Endocrinol. 2019, 10, 811. [Google Scholar] [CrossRef] [PubMed]
- Doroftei, B.; Ilie, O.-D.; Cojocariu, R.-O.; Ciobica, A.; Maftei, R.; Grab, D.; Anton, E.; McKenna, J.; Dhunna, N.; Simionescu, G. Minireview Exploring the Biological Cycle of Vitamin B3 and Its Influence on Oxidative Stress: Further Molecular and Clinical Aspects. Molecules 2020, 25, 3323. [Google Scholar] [CrossRef]
- Zhang, M.; Lu, Y.; Chen, Y.; Zhang, Y.; Xiong, B. Insufficiency of Melatonin in Follicular Fluid Is a Reversible Cause for Advanced Maternal Age-Related Aneuploidy in Oocytes. Redox Biol. 2020, 28, 101327. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Fu, B.; Xiong, Y.; Xu, S.; Liu, J.; Zaky, M.Y.; Qiu, D.; Wu, H. Folic Acid Ameliorates the Declining Quality of Sodium Fluoride-Exposed Mouse Oocytes through the Sirt1/Sod2 Pathway. Aging Dis. 2022, 13, 1471–1487. [Google Scholar] [CrossRef]
- Xing, X.; Zhang, J.; Wu, T.; Zhang, J.; Wang, Y.; Su, J.; Zhang, Y. SIRT1 Reduces Epigenetic and Non-Epigenetic Changes to Maintain the Quality of Postovulatory Aged Oocytes in Mice. Exp. Cell Res. 2021, 399, 112421. [Google Scholar] [CrossRef]
- John, G.B.; Gallardo, T.D.; Shirley, L.J.; Castrillon, D.H. Foxo3 Is a PI3K-Dependent Molecular Switch Controlling the Initiation of Oocyte Growth. Dev. Biol. 2008, 321, 197–204. [Google Scholar] [CrossRef]
- Kirillova, A.; Smitz, J.E.J.; Sukhikh, G.T.; Mazunin, I. The Role of Mitochondria in Oocyte Maturation. Cells 2021, 10, 2484. [Google Scholar] [CrossRef] [PubMed]
- Lima, T.; Li, T.Y.; Mottis, A.; Auwerx, J. Pleiotropic Effects of Mitochondria in Aging. Nat. Aging 2022, 2, 199–213. [Google Scholar] [CrossRef]
- Kong, D.; Yao, G.; Bai, Y.; Yang, G.; Xu, Z.; Kong, Y.; Fan, H.; He, Q.; Sun, Y. Expression of Sirtuins in Ovarian Follicles of Postnatal Mice. Mol. Reprod. Dev. 2020, 87, 1097–1108. [Google Scholar] [CrossRef] [PubMed]
- Zheng, B.; Meng, J.; Zhu, Y.; Ding, M.; Zhang, Y.; Zhou, J. Melatonin Enhances SIRT1 to Ameliorate Mitochondrial Membrane Damage by Activating PDK1/Akt in Granulosa Cells of PCOS. J. Ovarian. Res. 2021, 14, 152. [Google Scholar] [CrossRef]
- Furat Rencber, S.; Kurnaz Ozbek, S.; Eraldemır, C.; Sezer, Z.; Kum, T.; Ceylan, S.; Guzel, E. Effect of Resveratrol and Metformin on Ovarian Reserve and Ultrastructure in PCOS: An Experimental Study. J. Ovarian Res. 2018, 11, 1–16. [Google Scholar] [CrossRef]
- Tan, X.W.; You, W.; Song, E.L.; Zhao, H.B.; Liu, X.M.; Wang, H.Z.; Liu, G.F.; Cheng, H.J.; Liu, Y.F.; Wan, F.C. Effect of SIRT1 on Cellular Apoptosis and Steroidogenesis in Bovine Ovarian Granulosa Cells in Vitro. Livest. Sci. 2015, 180, 257–262. [Google Scholar] [CrossRef]
- Szymanska, M.; Manthe, S.; Shrestha, K.; Girsh, E.; Harlev, A.; Meidan, R. The CAMP Pathway Promotes Sirtuin-1 Expression in Human Granulosa-Lutein Cells. Reprod. Biol. 2020, 20, 273–281. [Google Scholar] [CrossRef]
- Sirotkin, A.V.; Dekanova, P.; Harrath, A.H. FSH, Oxytocin and IGF-I Regulate the Expression of Sirtuin 1 in Porcine Ovarian Granulosa Cells. Physiol. Res. 2020, 69, 461–466. [Google Scholar] [CrossRef]
- Azami, S.H.; Nazarian, H.; Abdollahifar, M.A.; Eini, F.; Farsani, M.A.; Novin, M.G.; Azami, S.H.; Nazarian, H.; Abdollahifar, M.A.; Eini, F.; et al. The Antioxidant Curcumin Postpones Ovarian Aging in Young and Middle-Aged Mice. Reprod. Fertil. Dev. 2020, 32, 292–303. [Google Scholar] [CrossRef]
- Gorczyca, G.; Wartalski, K.; Tabarowski, Z.; Duda, M. Effects of Vinclozolin Exposure on the Expression and Activity of SIRT1 and SIRT6 in the Porcine Ovary. J. Physiol. Pharm. 2019, 70, 153–165. [Google Scholar]
- Liu, Y.; Gao, J. Reproductive Aging: Biological Pathways and Potential Interventive Strategies. J. Genet. Genom. 2022, S1673-8527(22)00182-5. [Google Scholar] [CrossRef] [PubMed]
- Shvedunova, M.; Akhtar, A. Modulation of Cellular Processes by Histone and Non-Histone Protein Acetylation. Nat. Rev. Mol. Cell Biol. 2022, 23, 329–349. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Pickett, H.A. Targeting Telomeres: Advances in Telomere Maintenance Mechanism-Specific Cancer Therapies. Nat. Rev. Cancer 2022, 22, 515–532. [Google Scholar] [CrossRef] [PubMed]
- Córdova-Oriz, I.; Chico-Sordo, L.; Varela, E. Telomeres, Aging and Reproduction. Curr. Opin. Obstet. Gynecol. 2022, 34, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Revy, P.; Kannengiesser, C.; Bertuch, A.A. Genetics of Human Telomere Biology Disorders. Nat. Rev. Genet. 2022, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Dai, S.; Luo, X.; Zhu, J.; Li, F.; Liu, J.; Yao, G.; Sun, Y. Melatonin Attenuates Postovulatory Oocyte Dysfunction by Regulating SIRT1 Expression. Reproduction 2018, 156, 81–92. [Google Scholar] [CrossRef]
- Zhuan, Q.; Li, J.; Du, X.; Zhang, L.; Meng, L.; Cheng, K.; Zhu, S.; Hou, Y.; Fu, X. Nampt Affects Mitochondrial Function in Aged Oocytes by Mediating the Downstream Effector FoxO3a. J. Cell. Physiol. 2022, 237, 647–659. [Google Scholar] [CrossRef]
- Xiong, X.; Wang, Y.; Li, J.; Xiong, Y.; Zi, X.; Qiu, X. Effects of SIRT1 on the in Vitro Maturation and Aging of Yak Oocytes. Acta Vet. Et Zootech. Sin. 2019, 50, 2440–2448. [Google Scholar]
- Iljas, J.D.; Wei, Z.; Homer, H.A. Sirt1 Sustains Female Fertility by Slowing Age-Related Decline in Oocyte Quality Required for Post-Fertilization Embryo Development. Aging Cell 2020, 19, e13204. [Google Scholar] [CrossRef]
- Palacios, J.A.; Herranz, D.; De Bonis, M.L.; Velasco, S.; Serrano, M.; Blasco, M.A. SIRT1 Contributes to Telomere Maintenance and Augments Global Homologous Recombination. J. Cell Biol. 2010, 191, 1299–1313. [Google Scholar] [CrossRef]
- Fang, E.F.; Scheibye-Knudsen, M.; Brace, L.E.; Kassahun, H.; SenGupta, T.; Nilsen, H.; Mitchell, J.R.; Croteau, D.L.; Bohr, V.A. Defective Mitophagy in XPA via PARP-1 Hyperactivation and NAD+/SIRT1 Reduction. Cell 2014, 157, 882–896. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.P.; Price, N.L.; Ling, A.J.; Moslehi, J.J.; Montgomery, M.K.; Rajman, L.; White, J.P.; Teodoro, J.S.; Wrann, C.D.; Hubbard, B.P. Declining NAD+ Induces a Pseudohypoxic State Disrupting Nuclear-Mitochondrial Communication during Aging. Cell 2013, 155, 1624–1638. [Google Scholar] [CrossRef] [PubMed]
- De Bonis, M.L.; Ortega, S.; Blasco, M.A. SIRT1 Is Necessary for Proficient Telomere Elongation and Genomic Stability of Induced Pluripotent Stem Cells. Stem. Cell Rep. 2014, 2, 690–706. [Google Scholar] [CrossRef] [PubMed]
- Valerio, D.; Luddi, A.; De Leo, V.; Labella, D.; Longobardi, S.; Piomboni, P. SA1/SA2 Cohesion Proteins and SIRT1-NAD+ Deacetylase Modulate Telomere Homeostasis in Cumulus Cells and Are Eligible Biomarkers of Ovarian Aging. Hum. Reprod. 2018, 33, 887–894. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Cong, L.; Wang, Y.; Luo, X.; Li, H.; Wang, H.; Zhu, J.; Dai, S.; Jin, H.; Yao, G.; et al. Increasing Ovarian NAD+ Levels Improve Mitochondrial Functions and Reverse Ovarian Aging. Free Radic. Biol. Med. 2020, 156, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Liang, W.; Sun, Q.; Qiu, X.; Lin, Y.; Ge, X.; Jueraitetibaike, K.; Xie, M.; Zhou, J.; Huang, X. Sirt1/Nrf2 Pathway Is Involved in Oocyte Aging by Regulating Cyclin B1. Aging 2018, 10, 2991. [Google Scholar] [CrossRef]
- Zou, Y.; Hu, M.; Lee, J.; Nambiar, S.M.; Garcia, V.; Bao, Q.; Chan, J.Y.; Dai, G. Nrf2 Is Essential for Timely M Phase Entry of Replicating Hepatocytes during Liver Regeneration. Am. J. Physiol. -Gastrointest. Liver Physiol. 2015, 308, G262–G268. [Google Scholar] [CrossRef]
- Caldas, A.P.S.; Rocha, D.M.U.P.; Bressan, J.; Hermsdorff, H.H.M. Dietary Fatty Acids as Nutritional Modulators of Sirtuins: A Systematic Review. Nutr. Rev. 2021, 79, 235–246. [Google Scholar] [CrossRef]
- Ivanov, D.; Mazzoccoli, G.; Anderson, G.; Linkova, N.; Dyatlova, A.; Mironova, E.; Polyakova, V.; Kvetnoy, I.; Evsyukova, I.; Carbone, A.; et al. Melatonin, Its Beneficial Effects on Embryogenesis from Mitigating Oxidative Stress to Regulating Gene Expression. Int. J. Mol. Sci. 2021, 22, 5885. [Google Scholar] [CrossRef]
- Gao, Y.; Zhao, S.; Zhang, Y.; Zhang, Q. Melatonin Receptors: A Key Mediator in Animal Reproduction. Vet. Sci. 2022, 9, 309. [Google Scholar] [CrossRef]
- Guo, Y.; Sun, J.; Bu, S.; Li, B.; Zhang, Q.; Wang, Q.; Lai, D. Melatonin Protects against Chronic Stress-Induced Oxidative Meiotic Defects in Mice MII Oocytes by Regulating SIRT1. Cell Cycle 2020, 19, 1677–1695. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Li, Y.; Zhou, Y.; Yan, J.; Zhou, X.; Gao, Q.; Miao, Y.; Xiong, B. Nicotinamide Mononucleotide Supplementation Improves the Quality of Porcine Oocytes under Heat Stress. J. Anim. Sci. Biotechnol. 2022, 13, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.; Cui, Z.; Gao, Q.; Rui, R.; Xiong, B. Nicotinamide Mononucleotide Supplementation Reverses the Declining Quality of Maternally Aged Oocytes. Cell Rep. 2020, 32, 107987. [Google Scholar] [CrossRef] [PubMed]
- Ions, L.J.; Wakeling, L.A.; Bosomworth, H.J.; Hardyman, J.E.; Escolme, S.M.; Swan, D.C.; Valentine, R.A.; Mathers, J.C.; Ford, D. Effects of Sirt1 on DNA Methylation and Expression of Genes Affected by Dietary Restriction. Age 2013, 35, 1835–1849. [Google Scholar] [CrossRef] [PubMed]
- Wakeling, L.A.; Ions, L.J.; Ford, D. Could Sirt1-Mediated Epigenetic Effects Contribute to the Longevity Response to Dietary Restriction and Be Mimicked by Other Dietary Interventions? Age 2009, 31, 327–341. [Google Scholar] [CrossRef] [PubMed]
- O’Hagan, H.M.; Mohammad, H.P.; Baylin, S.B. Double Strand Breaks Can Initiate Gene Silencing and SIRT1-Dependent Onset of DNA Methylation in an Exogenous Promoter CpG Island. PLoS Genet. 2008, 4, e1000155. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grzeczka, A.; Kordowitzki, P. Resveratrol and SIRT1: Antiaging Cornerstones for Oocytes? Nutrients 2022, 14, 5101. https://doi.org/10.3390/nu14235101
Grzeczka A, Kordowitzki P. Resveratrol and SIRT1: Antiaging Cornerstones for Oocytes? Nutrients. 2022; 14(23):5101. https://doi.org/10.3390/nu14235101
Chicago/Turabian StyleGrzeczka, Arkadiusz, and Paweł Kordowitzki. 2022. "Resveratrol and SIRT1: Antiaging Cornerstones for Oocytes?" Nutrients 14, no. 23: 5101. https://doi.org/10.3390/nu14235101
APA StyleGrzeczka, A., & Kordowitzki, P. (2022). Resveratrol and SIRT1: Antiaging Cornerstones for Oocytes? Nutrients, 14(23), 5101. https://doi.org/10.3390/nu14235101