
The Influence of Family History of Type 2 Diabetes on Metabolism during Submaximal Aerobic Exercise and in the Recovery Period in Postmenopausal Women

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
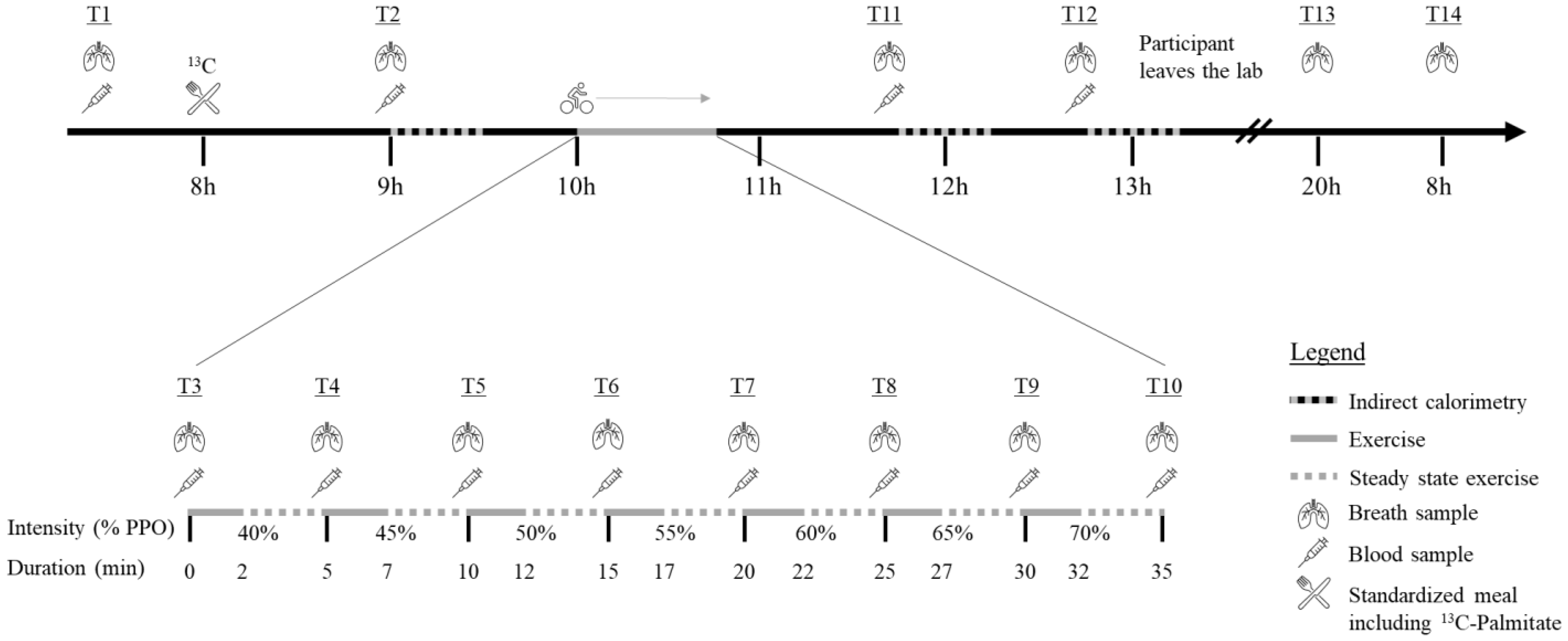
2.2. Protocol Overview
2.3. Anthropometrics and Body Composition
2.4. Indirect Calorimetry
2.5. Diet
2.6. Maximal Aerobic Capacity
2.7. Submaximal Exercise Testing
2.8. Blood Sampling
2.9. Biochemistry
2.10. Breath Sampling
2.11. Sources of Fat Oxidation
2.12. Statistics
3. Results
3.1. Population
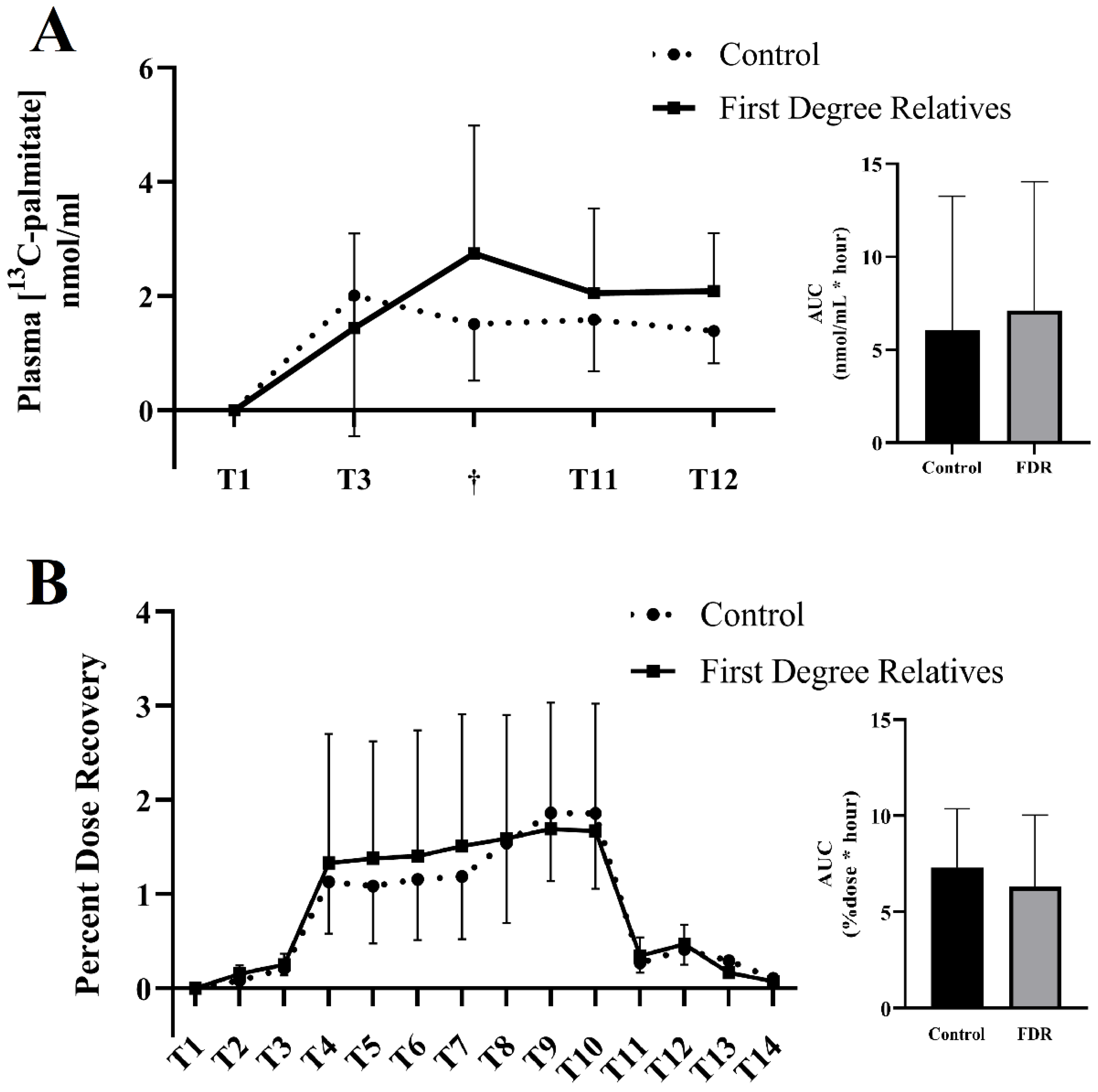
3.2. Primary Outcome: Expired & Plasmatic 13C-Palmitate
3.3. Secondary Outcomes: Blood Metabolites during Submaximal Exercise
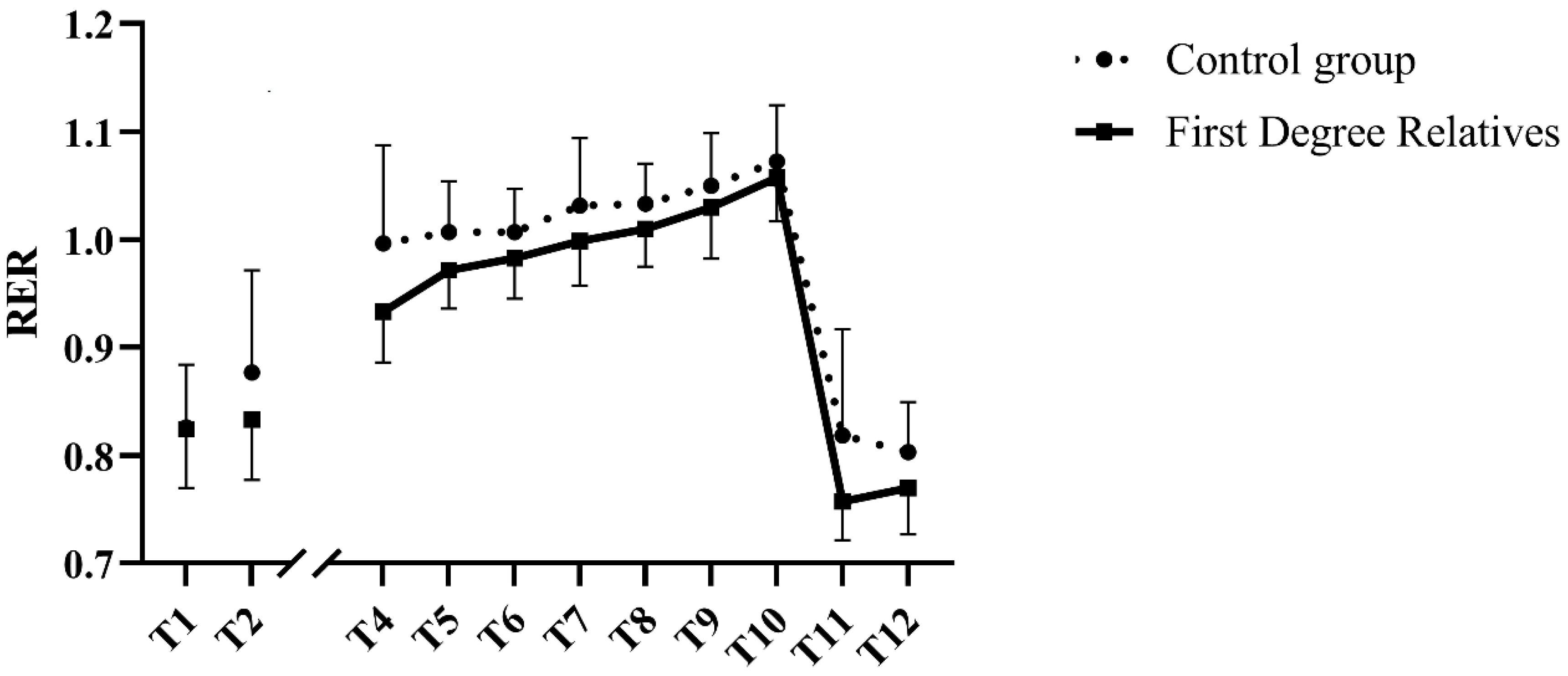
3.4. Submaximal Exercise Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hosseini, Z.; Whiting, S.J.; Vatanparast, H. Type 2 diabetes prevalence among Canadian adults—Dietary habits and sociodemographic risk factors. Appl. Physiol. Nutr. Metab. 2019, 44, 1099–1104. [Google Scholar] [CrossRef] [PubMed]
- Moccia, P.; Belda-Montesinos, R.; Monllor-Tormos, A.; Chedraui, P.; Cano, A. Body weight and fat mass across the menopausal transition: Hormonal modulators. Gynecol. Endocrinol. 2021, 38, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Wagner, R.; Thorand, B.; Osterhoff, M.A.; Müller, G.; Böhm, A.; Meisinger, C.; Kowall, B.; Rathmann, W.; Kronenberg, F.; Staiger, H.; et al. Family history of diabetes is associated with higher risk for prediabetes: A multicentre analysis from the German Center for Diabetes Research. Diabetologia 2013, 56, 2176–2180. [Google Scholar] [CrossRef] [PubMed]
- Nyholm, B.; Nielsen, M.F.; Kristensen, K.; Nielsen, S.; Østergård, T.; Pedersen, S.B.; Christiansen, T.; Richelsen, B.; Jensen, M.D.; Schmitz, O. Evidence of increased visceral obesity and reduced physical fitness in healthy insulin-resistant first-degree relatives of type 2 diabetic patients. Eur. J. Endocrinol. 2004, 150, 207–214. [Google Scholar] [CrossRef][Green Version]
- Henninger, J.; Rawshani, A.; Hammarstedt, A.; Eliasson, B. Metabolic characteristics of individuals at a high risk of type 2 diabetes—A comparative cross-sectional study. BMC Endocr. Disord. 2017, 17, 40. [Google Scholar] [CrossRef]
- Deng, L.; Liu, S.; Gong, Y.; Tian, H.; Liu, Y.; Song, J.; Ran, X.; Yu, H.; Zhang, X.; Long, Y.; et al. Increased Metabolic Disorders and Impaired Insulin Secretory Function in the First-Degree Relatives of Type 2 Diabetic Patients with Normal Glucose Tolerance. Metab. Syndr. Relat. Disord. 2016, 14, 431–436. [Google Scholar] [CrossRef]
- Baig, S.; Shabeer, M.; Rizi, E.P.; Agarwal, M.; Lee, M.H.; Ooi, D.S.Q.; Chia, C.; Aung, N.; Ng, G.; Teo, Y.; et al. Heredity of type 2 diabetes confers increased susceptibility to oxidative stress and inflammation. BMJ Open Diabetes Res. Care 2020, 8, e000945. [Google Scholar] [CrossRef]
- Ukropcova, B.; Sereda, O.; de Jonge, L.; Bogacka, I.; Nguyen, T.; Xie, H.; Bray, G.A.; Smith, S.R. Family History of Diabetes Links Impaired Substrate Switching and Reduced Mitochondrial Content in Skeletal Muscle. Diabetes 2007, 56, 720–727. [Google Scholar] [CrossRef]
- Russell, R.D.; Kraemer, R.R.; Nelson, A.G. Metabolic dysfunction in diabetic offspring: Deviations in metabolic flexibility. Med. Sci. Sports Exerc. 2013, 45, 8–15. [Google Scholar] [CrossRef]
- Allerton, T.D.; Irving, B.A.; Spielmann, G.; Primeaux, S.; Landin, D.; Nelson, A.; Johannsen, N.M. Metabolic flexibility is impaired in response to acute exercise in the young offspring of mothers with type 2 diabetes. Physiol. Rep. 2019, 7, e14189. [Google Scholar] [CrossRef]
- Befroy, D.E.; Petersen, K.F.; Dufour, S.; Mason, G.F.; de Graaf, R.A.; Rothman, D.L.; Shulman, G.I. Impaired Mitochondrial Substrate Oxidation in Muscle of Insulin-Resistant Offspring of Type 2 Diabetic Patients. Diabetes 2007, 56, 1376–1381. [Google Scholar] [CrossRef]
- Elgzyri, T.; Parikh, H.; Zhou, Y.; Nitert, M.D.; Rönn, T.; Segerström, Å.B.; Ling, C.; Franks, P.W.; Wollmer, P.; Eriksson, K.F.; et al. First-Degree Relatives of Type 2 Diabetic Patients Have Reduced Expression of Genes Involved in Fatty Acid Metabolism in Skeletal Muscle. J. Clin. Endocrinol. Metab. 2012, 97, E1332–E1337. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.; Pan, Y.; Zhong, T.; Zeng, Y.; Cheng, A.S. Effects of aerobic, resistance, and combined exercise on metabolic syndrome parameters and cardiovascular risk factors: A systematic review and network meta-analysis. Rev. Cardiovasc. Med. 2021, 22, 1523–1533. [Google Scholar] [CrossRef] [PubMed]
- Jelleyman, C.; Yates, T.; O’Donovan, G.; Gray, L.J.; King, J.A.; Khunti, K.; Davies, M.J. The effects of high-intensity interval training on glucose regulation and insulin resistance: A meta-analysis. Obes. Rev. 2015, 16, 942–961. [Google Scholar] [CrossRef] [PubMed]
- Amador, M.; Meza, C.A.; McAinch, A.J.; King, G.A.; Covington, J.D.; Bajpeyi, S. Exercise-Induced Improvements in Insulin Sensitivity Are Not Attenuated by a Family History of Type 2 Diabetes. Front. Endocrinol. 2020, 11, 120. [Google Scholar] [CrossRef] [PubMed]
- Barwell, N.D.; Malkova, D.; Moran, C.N.; Cleland, S.J.; Packard, C.J.; Zammit, V.A.; Gill, J.M.R. Exercise training has greater effects on insulin sensitivity in daughters of patients with type 2 diabetes than in women with no family history of diabetes. Diabetologia 2008, 51, 1912. [Google Scholar] [CrossRef]
- Ekman, C.; Elgzyri, T.; Ström, K.; Almgren, P.; Parikh, H.; Nitert, M.D.; Rönn, T.; Koivula, F.M.; Ling, C.; Tornberg, Å.B.; et al. Less pronounced response to exercise in healthy relatives to type 2 diabetic subjects compared with controls. J. Appl. Physiol. 2015, 119, 953–960. [Google Scholar] [CrossRef]
- Irving, B.A.; Short, K.R.; Nair, K.S.; Stump, C.S. Nine Days of Intensive Exercise Training Improves Mitochondrial Function but not Insulin Action in Adult Offspring of Mothers with Type 2 Diabetes. J. Clin. Endocrinol. Metab. 2011, 96, E1137–E1141. [Google Scholar] [CrossRef][Green Version]
- Zacharewicz, E.; Hesselink, M.K.C.; Schrauwen, P. Exercise counteracts lipotoxicity by improving lipid turnover and lipid droplet quality. J. Intern. Med. 2018, 284, 505–518. [Google Scholar] [CrossRef]
- Purdom, T.; Kravitz, L.; Dokladny, K.; Mermier, C. Understanding the factors that effect maximal fat oxidation. J. Int. Soc. Sports Nutr. 2018, 15, 3. [Google Scholar] [CrossRef]
- Goodpaster, B.H.; Wolfe, R.R.; Kelley, D.E. Effects of Obesity on Substrate Utilization during Exercise. Obes. Res. 2002, 10, 575–584. [Google Scholar] [CrossRef] [PubMed]
- Maltais, M.L.; Perreault, K.; Courchesne-Loyer, A.; Lagacé, J.C.; Barsalani, R.; Dionne, I.J. Effect of Resistance Training and Various Sources of Protein Supplementation on Body Fat Mass and Metabolic Profile in Sarcopenic Overweight Elderly Men: A Pilot Study. Int. J. Sport Nutr. Exerc. Metab. 2016, 26, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Goran, M.I.; Poehlman, E.T. Total energy expenditure and energy requirements in healthy elderly persons. Metabolism 1992, 41, 744–753. [Google Scholar] [CrossRef]
- Mercuro, G.; Saiu, F.; Deidda, M.; Mercuro, S.; Vitale, C.; Rosano, G.M. Impairment of physical exercise capacity in healthy postmenopausal women. Am. Heart J. 2006, 151, 923–927. [Google Scholar] [CrossRef]
- Stisen, A.B.; Stougaard, O.; Langfort, J.; Helge, J.W.; Sahlin, K.; Madsen, K. Maximal fat oxidation rates in endurance trained and untrained women. Eur. J. Appl. Physiol. 2006, 98, 497–506. [Google Scholar] [CrossRef]
- Williams, N. The Borg Rating of Perceived Exertion (RPE) scale. Occup. Med. 2017, 67, 404–405. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Chevalier, L.; Plourde, M. Comparison of pharmacokinetics of omega-3 fatty acid supplements in monoacylglycerol or ethyl ester in humans: A randomized controlled trial. Eur. J. Clin. Nutr. 2020, 75, 680–688. [Google Scholar] [CrossRef]
- Chevalier, L.; Vachon, A.; Plourde, M. Pharmacokinetics of Supplemental Omega-3 Fatty Acids Esterified in Monoglycerides, Ethyl Esters, or Triglycerides in Adults in a Randomized Crossover Trial. J. Nutr. 2021, 151, 1111–1118. [Google Scholar] [CrossRef]
- Goodman, K.J.; Brenna, J.T. High sensitivity tracer detection using high-precision gas chromatography-combustion isotope ratio mass spectrometry and highly enriched [U-13C]-labeled precursors. Anal. Chem. 1992, 64, 1088–1095. [Google Scholar] [CrossRef]
- Brossard, N.; Croset, M.; Normand, S.; Pousin, J.; Lecerf, J.; Laville, M.; Tayot, J.L.; Lagarde, M. Human plasma albumin transports [13C]docosahexaenoic acid in two lipid forms to blood cells. J. Lipid Res. 1997, 38, 1571–1582. [Google Scholar] [CrossRef]
- Plourde, M.; Chouinard-Watkins, R.; Vandal, M.; Zhang, Y.; Lawrence, P.; Brenna, J.T.; Cunnane, S.C. Plasma incorporation, apparent retroconversion and β-oxidation of 13C-docosahexaenoic acid in the elderly. Nutr. Metab. 2011, 8, 5. [Google Scholar] [CrossRef] [PubMed]
- Nadler, S.B.; Hidalgo, J.H.; Bloch, T. Prediction of blood volume in normal human adults. Surgery 1962, 51, 224–232. [Google Scholar]
- Freemantle, E.; Vandal, M.; Tremblay-Mercier, J.; Plourde, M.; Poirier, J.; Cunnane, S.C. Metabolic response to a ketogenic breakfast in the healthy elderly. J. Nutr. Health Aging 2009, 13, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Nellemann, B.; Søndergaard, E.; Jensen, J.; Pedersen, S.B.; Jessen, N.; Jørgensen, J.O.L.; Nielsen, S. Kinetics and utilization of lipid sources during acute exercise and acipimox. Am. J. Physiol. Metab. 2014, 307, E199–E208. [Google Scholar] [CrossRef]
- Sondergaard, E.; Rahbek, I.; Sørensen, L.P.; Christiansen, J.S.; Gormsen, L.C.; Jensen, M.D.; Nielsen, S. Effects of exercise on VLDL-triglyceride oxidation and turnover. Am. J. Physiol. Metab. 2011, 300, E939–E944. [Google Scholar] [CrossRef] [PubMed]
- Richter, E.A.; Sylow, L.; Hargreaves, M. Interactions between insulin and exercise. Biochem. J. 2021, 478, 3827–3846. [Google Scholar] [CrossRef]
- Eriksson, J.W.; Smith, U.; Waagstein, F.; Wysocki, M.; Jansson, P.A. Glucose turnover and adipose tissue lipolysis are insulin-resistant in healthy relatives of type 2 diabetes patients: Is cellular insulin resistance a secondary phenomenon? Diabetes 1999, 48, 1572–1578. [Google Scholar] [CrossRef]
- Dahlman, I.; Ryden, M.; Arner, P. Family history of diabetes is associated with enhanced adipose lipolysis: Evidence for the implication of epigenetic factors. Diabetes Metab. 2017, 44, 155–159. [Google Scholar] [CrossRef]
- Bianco, A.; Pomara, F.; Patti, A.; Thomas, E.; Petrucci, M.; Bellafiore, M.; Battaglia, G.; Paoli, A.; Palma, A. The surprising influence of family history to type 2 diabetes on anaerobic performance of young male élite athletes. SpringerPlus 2014, 3, 224. [Google Scholar] [CrossRef]
- Nyholm, B.; Qu, Z.; Kaal, A.; Pedersen, S.B.; Gravholt, C.H.; Andersen, J.L.; Saltin, B.; Schmitz, O. Evidence of an increased number of type IIb muscle fibers in insulin-resistant first-degree relatives of patients with NIDDM. Diabetes 1997, 46, 1822–1828. [Google Scholar] [CrossRef] [PubMed]
 = Indirect calorimetry;
= Indirect calorimetry;  = period of exercise;
= period of exercise;  = steady state exercise;
= steady state exercise;  = breath sample;
= breath sample;  = blood sample;
= blood sample;  = Standardized meal including 13C-Palmitate.
= Indirect calorimetry; = period of exercise; = steady state exercise; = breath sample; = blood sample; = Standardized meal including 13C-Palmitate.
= Standardized meal including 13C-Palmitate.
= Indirect calorimetry; = period of exercise; = steady state exercise; = breath sample; = blood sample; = Standardized meal including 13C-Palmitate.



{kind=link}
{kind=link}
{kind=link}
| Control (n = 10) | FDR (n = 9) | |
|---|---|---|
| Age (years) | 67.7 ± 4.7 | 67.6 ± 4.2 |
| Height (m) | 1.60 ± 0.07 | 1.58 ± 0.06 |
| Weight (kg) | 65.3 ± 6.4 | 65.6 ± 9.9 |
| BMI (kg/m2) | 25.6 ± 2.2 | 26.3 ± 4.2 |
| Waist Circumference (cm) | 101.9 ± 20.8 | 97.5 ± 8.8 |
| Fat-free mass (kg) | 36.4 ± 3.2 | 38.2 ± 4.6 |
| Fat mass (kg) | 26.4 ± 5.5 | 25.5 ± 5.4 |
| Fat mass (%) | 41.8 ± 5.5 | 39.8 ± 4.7 |
| Resting vital signs & metabolism | ||
| Resting Heart Rate (bpm) | 62.4 ± 8.0 | 62.89 ± 2.62 |
| Systolic blood pressure (mmHg) | 121.5 ± 6.6 | 122.7 ± 13.9 |
| Diastolic blood pressure (mmHg) | 75.0 ± 6.0 | 73.7 ± 7.6 |
| Resting metabolic rate (kcal/day) | 1099.5 ± 120.4 | 1151.1 ± 248.3 |
| Fasting blood samples | ||
| Fasting glucose (mmol/L) | 4.68 ± 0.46 | 4.57 ± 0.49 |
| Fasting insulin (pmol/L) | 36.67 ± 8.96 | 67.13 ± 43.30 |
| HbA1c (%) | 5.65 ± 0.26 | 5.47 ± 0.23 |
| HDL-cholesterol (mmol/L) | 1.94 ± 0.49 | 1.62 ± 0.42 |
| LDL-cholesterol (mmol/L) | 3.55 ± 1.02 | 3.63 ± 0.78 |
| Total Cholesterol (mmol/L) | 6.00 ± 1.06 | 5.82 ± 1.06 |
| Triglycerides (mmol/L) | 1.13 ± 0.46 | 1.26 ± 0.60 |
| Maximal aerobic capacity | ||
| Absolute V̇O2 peak (L/min) | 1.2 ± 0.3 | 1.3 ± 0.2 |
| Relative V̇O2 peak (ml/kg/min) | 18.7 ± 3.8 | 20.4 ± 2.7 |
| Peak Power Output (W) | 94.0 ± 21.7 | 95.6 ± 15.9 |
| Peak Heart Rate (bpm) | 153.6 ± 15.6 | 149.9 ± 9.5 |
| Pre-Exercise | Stage 1 | Stage 2 | Stage 3 | Stage 4 | Stage 5 | Stage 6 | Stage 7 | ||
|---|---|---|---|---|---|---|---|---|---|
| Glucose (mmol/L) | Control | 5.19 ± 1.02 | 5.42 ± 0.86 | 5.46 ± 0.68 | 5.36 ± 0.67 | 5.31 ± 0.61 | 5.01 ± 0.43 | 5.49 ± 0.92 | 5.26 ± 0.51 |
| FDR | 5.71 ± 0.81 | 5.89 ± 1.02 | 6.00 ± 0.92 | 5.44 ± 0.75 | 5.46 ± 0.96 | 5.54 ± 0.92 | 5.41 ± 1.01 | 5.68 ± 0.81 | |
| Lactate (mmol/L) a,b | Control | 1.30 ± 0.29 | 2.29 ± 0.68 | 3.02 ± 1.20 | 3.43 ± 0.94 | 4.01 ± 1.15 | 4.53 ± 1.21 | 5.70 ± 1.51 | 6.49 ± 1.30 |
| FDR | 1.55 ± 0.69 | 2.11 ± 0.57 | 2.86 ± 0.88 | 3.05 ± 0.73 | 3.66 ± 1.39 | 4.30 ± 1.59 | 4.92 ± 1.62 | 5.91 ± 2.18 | |
| NEFA (mmol/L) | Control | 0.37 ± 0.14 | 0.36 ± 0.17 | 0.41 ± 0.24 | 0.39 ± 0.19 | 0.31 ± 0.07 | 0.37 ± 0.14 | 0.36 ± 0.09 | 0.38 ± 0.08 |
| FDR | 0.40 ± 0.22 | 0.47 ± 0.26 | 0.35 ± 0.19 | 0.4 ± 0.27 | 0.41 ± 0.18 | 0.40 ± 0.18 | 0.47 ± 0.24 | 0.46 ± 0.21 | |
| Glycerol (μmol/L) a | Control | 80.31 ± 30.48 | 77.14 ± 20.73 | 90.81 ± 29.13 | 112.56 ± 40.70 | 98.65 ± 13.25 | - | 154.66 ± 28.57 | 157.22 ± 13.95 |
| FDR | 89.14 ± 46.53 | 99.12 ± 53.79 | 97.11 ± 43.24 | 99.61 ± 56.68 | 111.25 ± 58.32 | 107.57 ± 42.65 | 152.62 ± 78.65 | 171.19 ± 71.99 |
| Stage 1 | Stage 2 | Stage 3 | Stage 4 | Stage 5 | Stage 6 | Stage 7 | ||
|---|---|---|---|---|---|---|---|---|
| Heart Rate Reserve (%) a,b | Control | 56.3 ± 12.4 | 64.0 ± 12.7 | 70.9 ± 12.0 | 78.9 ± 12.3 | 85.8 ± 12.0 | 89.2 ± 8.0 | 95.0 ± 6.5 |
| FDR | 41.5 ± 8.7 | 48.8 ± 8.6 | 55.9 ± 10.0 | 64.7 ± 10.4 | 70.6 ± 10.3 | 78.0 ± 9.3 | 83.2 ± 9.9 | |
| Perceived exertion a | Control | 2.0 ± 1.0 | 2.5 ± 1.3 | 3.3 ± 1.7 | 3.2 ± 1.3 | 3.9 ± 1.1 | 3.9 ± 0.7 | 4.8 ± 1.1 |
| FDR | 1.4 ± 0.8 | 2.0 ± 1.4 | 2.1 ± 0.9 | 2.7 ± 0.8 | 3.3 ± 1.2 | 3.9 ± 1.2 | 4.3 ± 1.1 | |
| V̇O2 (L/min) a | Control | 0.88 ± 0.13 | 0.9 ± 0.2 | 0.93 ± 0.23 | 1.02 ± 0.28 | 1.14 ± 0.31 | 1.2 ± 0.35 | 1.36 ± 0.34 |
| FDR | 0.89 ± 0.13 | 0.91 ± 0.14 | 0.95 ± 0.12 | 1.01 ± 0.12 | 1.05 ± 0.15 | 1.1 ± 0.17 | 1.13 ± 0.16 | |
| V̇CO2 (L/min) a | Control | 0.87 ± 0.09 | 0.90 ± 0.17 | 0.93 ± 0.21 | 1.05 ± 0.26 | 1.17 ± 0.28 | 1.35 ± 0.26 | 1.41 ± 0.26 |
| FDR | 0.83 ± 0.14 | 0.88 ± 0.15 | 0.93 ± 0.14 | 1.01 ± 0.13 | 1.06 ± 0.16 | 1.13 ± 0.18 | 1.20 ± 0.18 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lagacé, J.-C.; Paquin, J.; Tremblay, R.; St-Martin, P.; Tessier, D.; Plourde, M.; Riesco, E.; Dionne, I.J. The Influence of Family History of Type 2 Diabetes on Metabolism during Submaximal Aerobic Exercise and in the Recovery Period in Postmenopausal Women. Nutrients 2022, 14, 4638. https://doi.org/10.3390/nu14214638
Lagacé J-C, Paquin J, Tremblay R, St-Martin P, Tessier D, Plourde M, Riesco E, Dionne IJ. The Influence of Family History of Type 2 Diabetes on Metabolism during Submaximal Aerobic Exercise and in the Recovery Period in Postmenopausal Women. Nutrients. 2022; 14(21):4638. https://doi.org/10.3390/nu14214638
Chicago/Turabian StyleLagacé, Jean-Christophe, Jasmine Paquin, Renaud Tremblay, Philippe St-Martin, Daniel Tessier, Mélanie Plourde, Eléonor Riesco, and Isabelle J. Dionne. 2022. "The Influence of Family History of Type 2 Diabetes on Metabolism during Submaximal Aerobic Exercise and in the Recovery Period in Postmenopausal Women" Nutrients 14, no. 21: 4638. https://doi.org/10.3390/nu14214638
APA StyleLagacé, J.-C., Paquin, J., Tremblay, R., St-Martin, P., Tessier, D., Plourde, M., Riesco, E., & Dionne, I. J. (2022). The Influence of Family History of Type 2 Diabetes on Metabolism during Submaximal Aerobic Exercise and in the Recovery Period in Postmenopausal Women. Nutrients, 14(21), 4638. https://doi.org/10.3390/nu14214638