
Capsaicin and Its Effect on Exercise Performance, Fatigue and Inflammation after Exercise


 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects and General Procedures
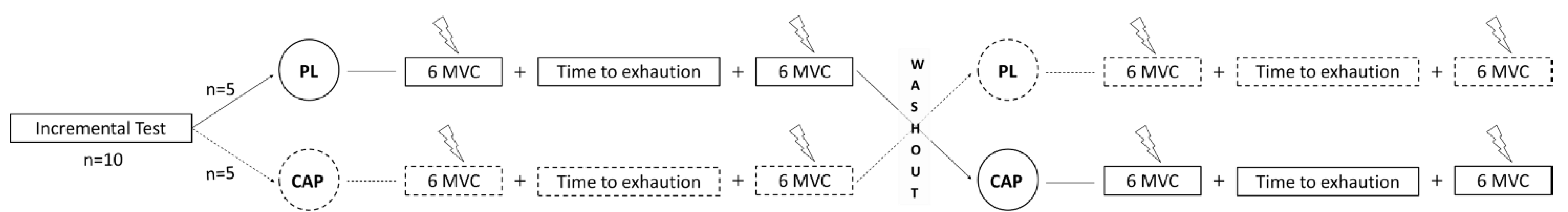
2.2. Experimental Design
2.3. Cardiorespiratory Exercise Responses
2.4. Assessment of Neuromuscular Function and Fatigue
2.5. Microvascular Oxygenation
2.6. Salivary Analysis
2.7. Biochemical Analysis of Capsules
2.8. Statistical Analysis
3. Results
3.1. Participant Characteristics
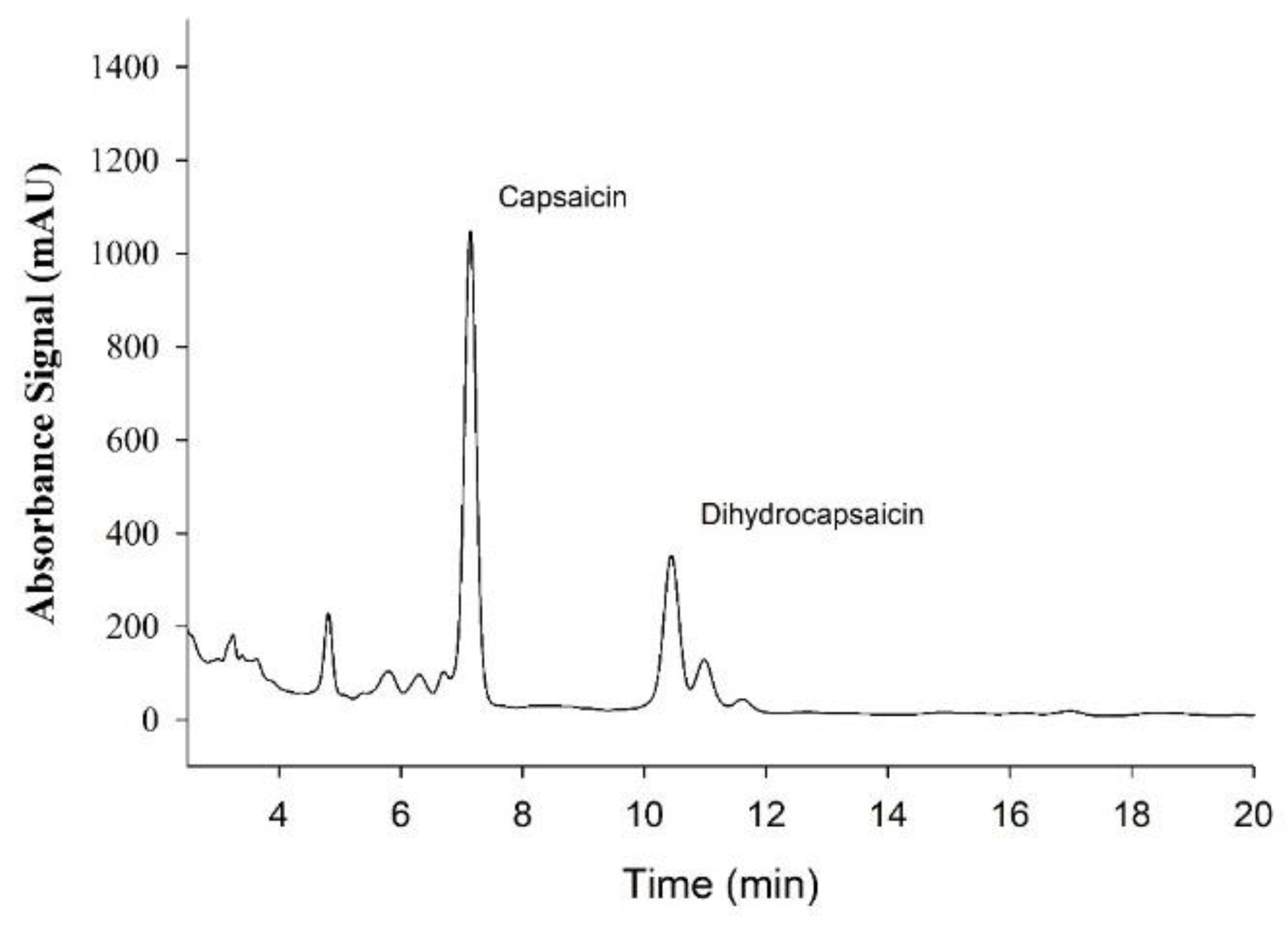
3.2. Supplement Analysis
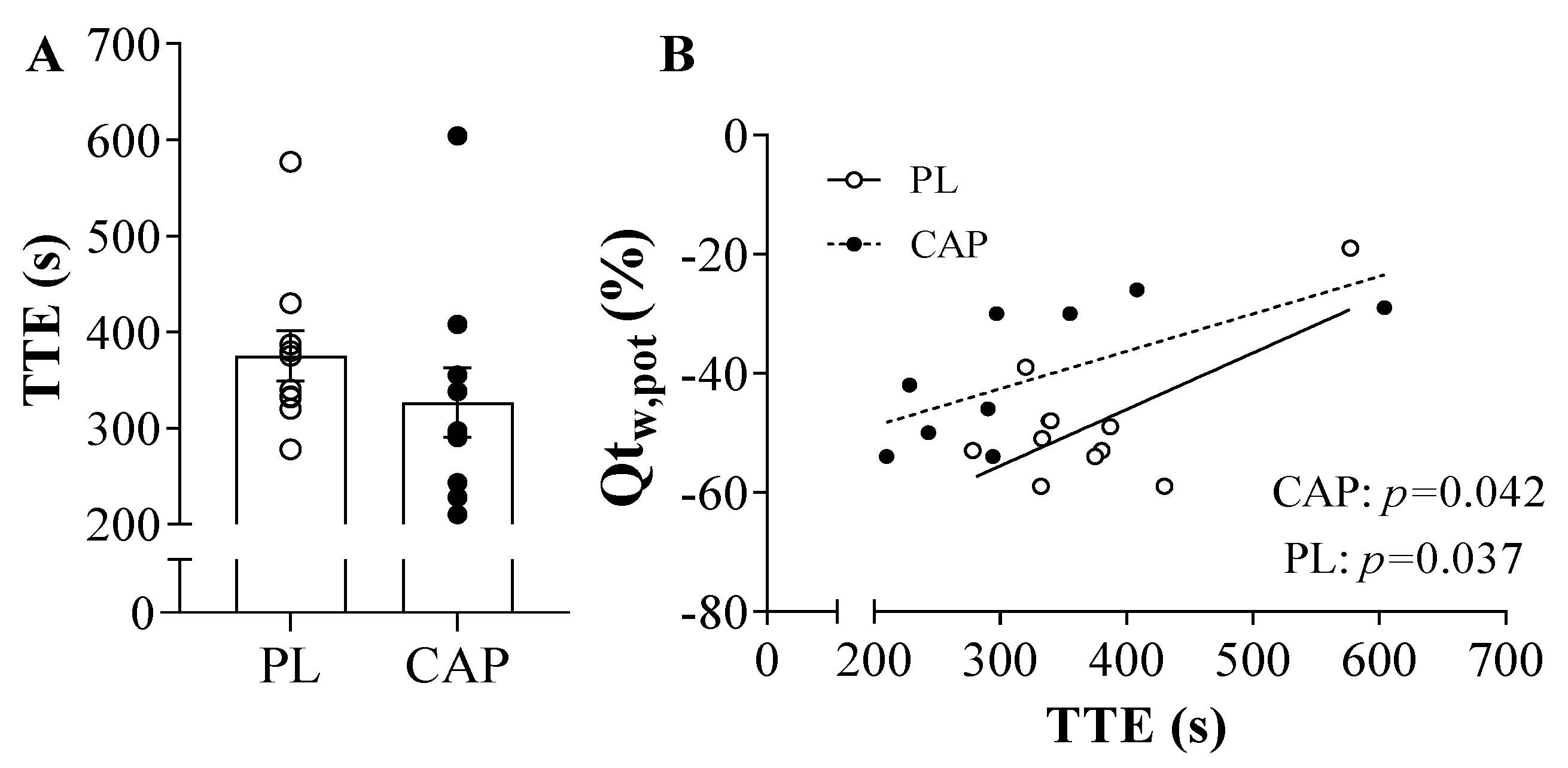
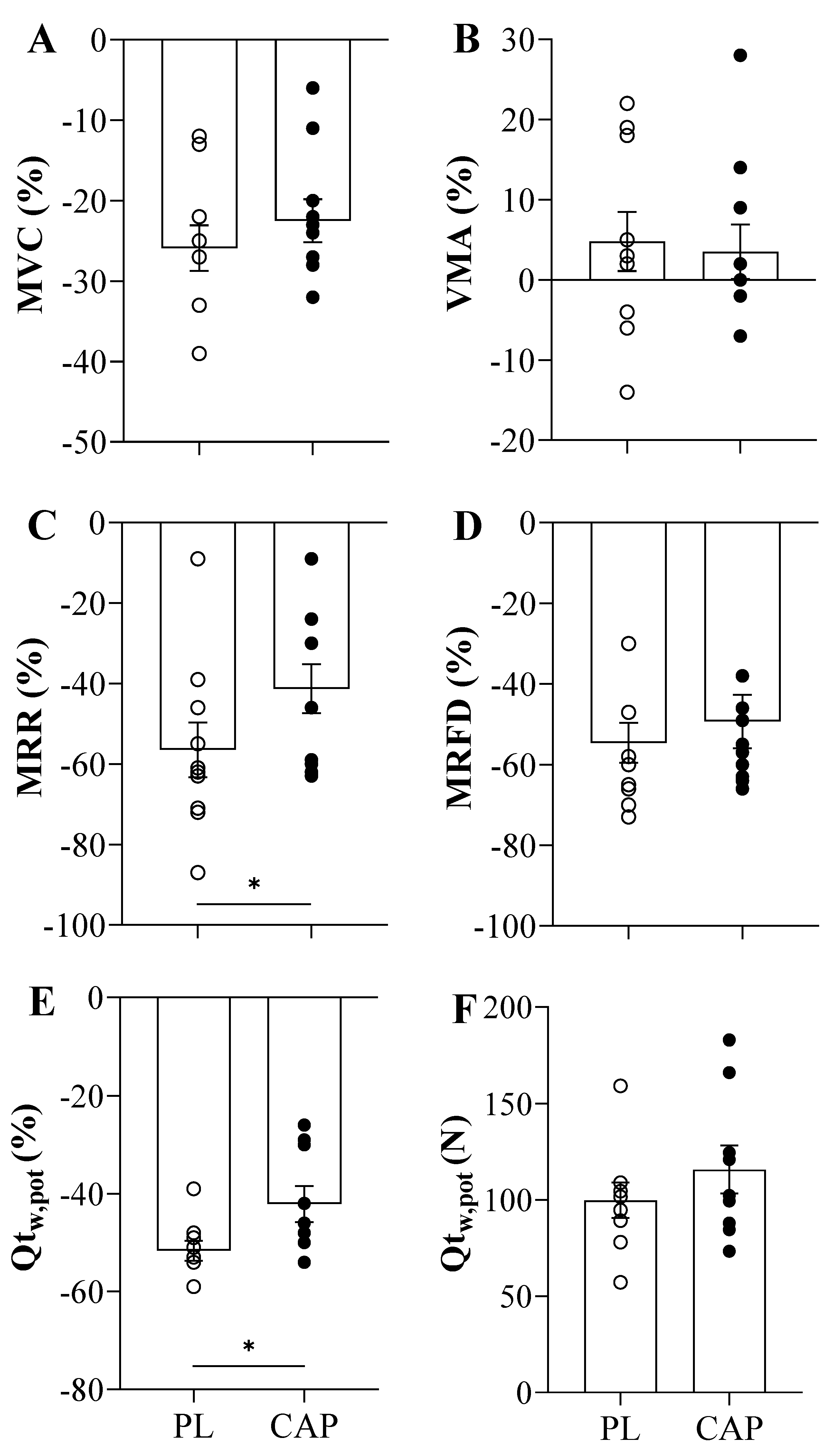
3.3. Exercise Performance, Neuromuscular Function and Fatigue
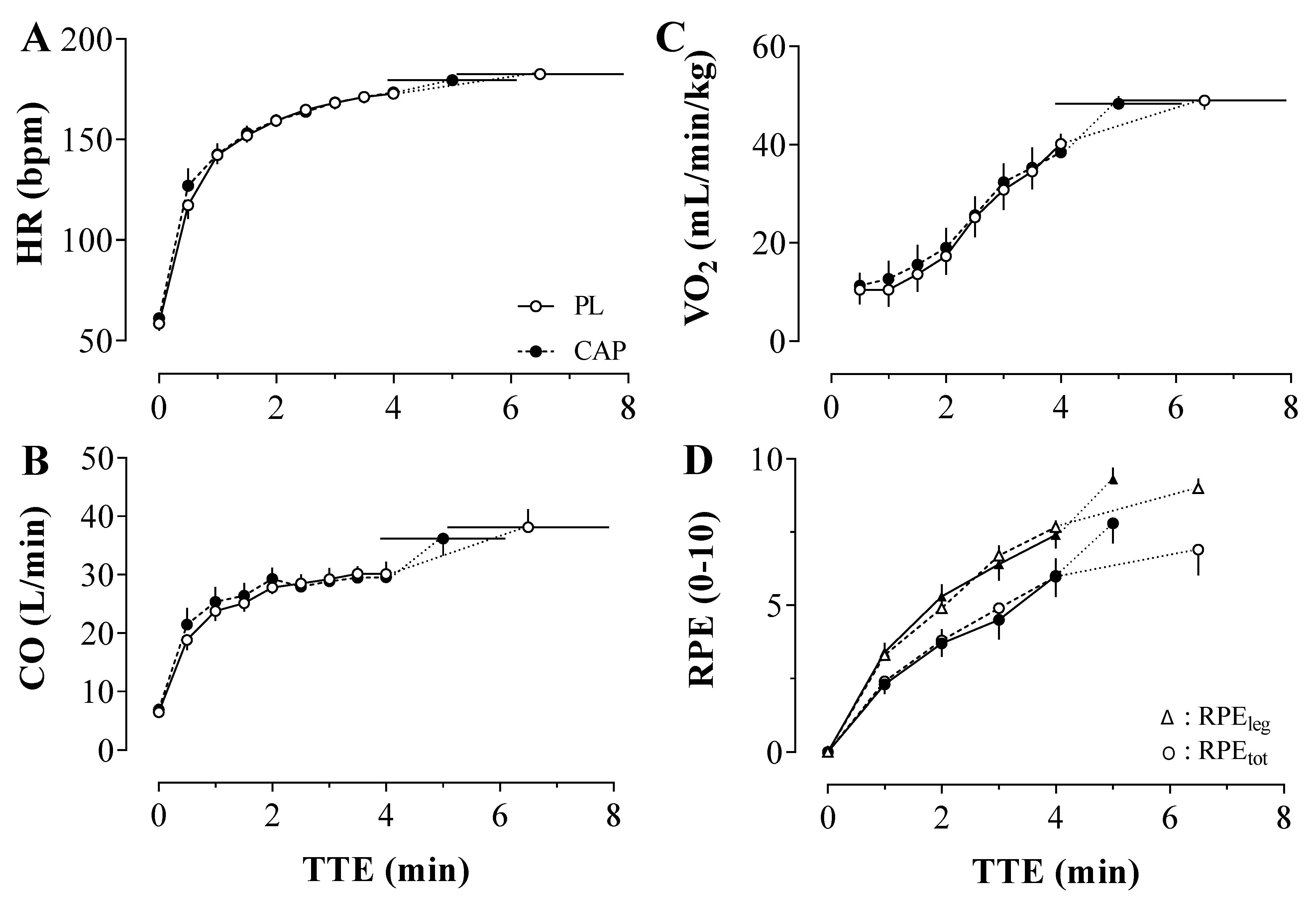
3.4. Microvascular Oxygenation during the TTE
3.5. Central Hemodynamics, Ventilation and Perceived Exertion during the TTE
3.6. Stress and Proinflammatory Biomarkers
4. Discussion
4.1. CAP and Exercise Performance
4.2. CAP and Exercise-Induced Neuromuscular Fatigue
4.3. CAP and the Physiological Response to Exercise
4.4. CAP and Neuroinflammatory Indices
4.5. Limitations of the Study
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vyklický, L.; Nováková-Tousová, K.; Benedikt, J.; Samad, A.; Touska, F.; Vlachová, V. Calcium-dependent desensitization of vanilloid receptor TRPV1: A mechanism possibly involved in analgesia induced by topical application of capsaicin. Physiol. Res. 2008, 57 (Suppl. 3), S59–S68. [Google Scholar] [CrossRef] [PubMed]
- Starowicz, K.; Cristino, L.; Di Marzo, V. TRPV1 receptors in the central nervous system: Potential for previously unforeseen therapeutic applications. Curr. Pharm. Des. 2008, 14, 42–54. [Google Scholar] [CrossRef]
- Kopanitsa, M.V.; Panchenko, V.A.; Magura, E.I.; Lishko, P.V.; Krishtal, O.A. Capsaicin blocks Ca2+ channels in isolated rat trigeminal and hippocampal neurones. Neuroreport 1995, 6, 2338–2340. [Google Scholar] [CrossRef]
- Sim, J.H.; Kim, Y.C.; Kim, S.J.; Lee, S.J.; Suh, S.H.; Jun, J.Y.; So, I.; Kim, K.W. Capsaicin inhibits the voltage-operated calcium channels intracellularly in the antral circular myocytes of guinea-pig stomach. Life Sci. 2001, 68, 2347–2360. [Google Scholar] [CrossRef]
- Southall, M.D.; Li, T.; Gharibova, L.S.; Pei, Y.; Nicol, G.D.; Travers, J.B. Activation of epidermal vanilloid receptor-1 induces release of proinflammatory mediators in human keratinocytes. J. Pharmacol. Exp. Ther. 2003, 304, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Basu, S.; Srivastava, P. Immunological role of neuronal receptor vanilloid receptor 1 expressed on dendritic cells. Proc. Natl. Acad. Sci. USA 2005, 102, 5120–5125. [Google Scholar] [CrossRef]
- Almeida, V.; Peres, F.F.; Levin, R.; Suiama, M.A.; Calzavara, M.B.; Zuardi, A.W.; Hallak, J.E.; Crippa, J.A.; Abílio, V.C. Effects of cannabinoid and vanilloid drugs on positive and negative-like symptoms on an animal model of schizophrenia: The SHR strain. Schizophr. Res. 2014, 153, 150–159. [Google Scholar] [CrossRef]
- Afari, N.; Buchwald, D. Chronic fatigue syndrome: A review. Am. J. Psychiatry 2003, 160, 221–236. [Google Scholar] [CrossRef]
- Hsu, C.L.; Yen, G.C. Effects of capsaicin on induction of apoptosis and inhibition of adipogenesis in 3T3-L1 cells. J. Agric. Food Chem. 2007, 55, 1730–1736. [Google Scholar] [CrossRef]
- Sarvaiya, K.; Goswami, S. Investigation of the effects of vanilloids in chronic fatigue syndrome. Brain Res. Bull. 2016, 127, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, F.; Inoue, N.; Yazawa, S.; Kawada, T.; Inoue, K.; Fushiki, T. Effects of CH-19 sweet, a non-pungent cultivar of red pepper, in decreasing the body weight and suppressing body fat accumulation by sympathetic nerve activation in humans. Biosci. Biotechnol. Biochem. 2006, 70, 2824–2835. [Google Scholar] [CrossRef]
- Ohnuki, K.; Moritani, T.; Ishihara, K.; Fushiki, T. Capsaicin increases modulation of sympathetic nerve activity in rats: Measurement using power spectral analysis of heart rate fluctuations. Biosci. Biotechnol. Biochem. 2001, 65, 638–643. [Google Scholar] [CrossRef]
- Ralevic, V.; Kendall, D.A.; Randall, M.D.; Zygmunt, P.M.; Movahed, P.; Hogestatt, E.D. Vanilloid receptors on capsaicin-sensitive sensory nerves mediate relaxation to methanandamide in the rat isolated mesenteric arterial bed and small mesenteric arteries. Br. J. Pharm. 2000, 130, 1483–1488. [Google Scholar] [CrossRef]
- Vanner, S. Mechanism of action of capsaicin on submucosal arterioles in the guinea pig ileum. Am. J. Physiol. 1993, 265, G51–G55. [Google Scholar] [CrossRef]
- Wang, L.H.; Luo, M.; Wang, Y.; Galligan, J.J.; Wang, D.H. Impaired vasodilation in response to perivascular nerve stimulation in mesenteric arteries of TRPV1-null mutant mice. J. Hypertens. 2006, 24, 2399–2408. [Google Scholar] [CrossRef]
- Bratz, I.N.; Dick, G.M.; Tune, J.D.; Edwards, J.M.; Neeb, Z.P.; Dincer, U.D.; Sturek, M. Impaired capsaicin-induced relaxation of coronary arteries in a porcine model of the metabolic syndrome. Am. J. Physiol. Heart Circ. Physiol. 2008, 294, H2489–H2496. [Google Scholar] [CrossRef] [PubMed]
- Hogaboam, C.M.; Wallace, J.L. Inhibition of platelet aggregation by capsaicin. An effect unrelated to actions on sensory afferent neurons. Eur. J. Pharm. 1991, 202, 129–131. [Google Scholar] [CrossRef]
- Kumar, P.; Chand, S.; Chandra, P.; Maurya, P.K. Influence of Dietary Capsaicin on Redox Status in Red Blood Cells during Human Aging. Adv. Pharm. Bull. 2015, 5, 583–586. [Google Scholar] [CrossRef] [PubMed][Green Version]
- McCarty, M.F.; DiNicolantonio, J.J.; O’Keefe, J.H. Capsaicin may have important potential for promoting vascular and metabolic health. Open Heart 2015, 2, e000262. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Luo, Z.; Ma, S.; Wong, W.T.; Ma, L.; Zhong, J.; He, H.; Zhao, Z.; Cao, T.; Yan, Z.; et al. Activation of TRPV1 by dietary capsaicin improves endothelium-dependent vasorelaxation and prevents hypertension. Cell Metab. 2010, 12, 130–141. [Google Scholar] [CrossRef]
- Hsu, Y.J.; Huang, W.C.; Chiu, C.C.; Liu, Y.L.; Chiu, W.C.; Chiu, C.H.; Chiu, Y.S.; Huang, C.C. Capsaicin Supplementation Reduces Physical Fatigue and Improves Exercise Performance in Mice. Nutrients 2016, 8, 648. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.M.; Kawada, T.; Ishihara, K.; Inoue, K.; Fushiki, T. Increase in swimming endurance capacity of mice by capsaicin-induced adrenal catecholamine secretion. Biosci. Biotechnol. Biochem. 1997, 61, 1718–1723. [Google Scholar] [CrossRef]
- Oh, T.W.; Oh, T.W.; Ohta, F. Dose-dependent effect of capsaicin on endurance capacity in rats. Br. J. Nutr. 2003, 90, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Oh, T.W.; Ohta, F. Capsaicin increases endurance capacity and spares tissue glycogen through lipolytic function in swimming rats. J. Nutr. Sci. Vitaminol. 2003, 49, 107–111. [Google Scholar] [CrossRef]
- Kazuya, Y.; Tonson, A.; Pecchi, E.; Dalmasso, C.; Vilmen, C.; Fur, Y.L.; Bernard, M.; Bendahan, D.; Giannesini, B. A single intake of capsiate improves mechanical performance and bioenergetics efficiency in contracting mouse skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E1110–E1119. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Ma, L.; Zhao, Z.; He, H.; Yang, D.; Feng, X.; Ma, S.; Chen, X.; Zhu, T.; Cao, T.; et al. TRPV1 activation improves exercise endurance and energy metabolism through PGC-1α upregulation in mice. Cell Res. 2012, 22, 551–564. [Google Scholar] [CrossRef] [PubMed]
- Faraut, B.; Giannesini, B.; Matarazzo, V.; Marqueste, T.; Dalmasso, C.; Rougon, G.; Cozzone, P.J.; Bendahan, D. Downregulation of uncoupling protein-3 in vivo is linked to changes in muscle mitochondrial energy metabolism as a result of capsiate administration. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E1474–E1482. [Google Scholar] [CrossRef] [PubMed]
- de Freitas, M.C.; Billaut, F.; Panissa, V.L.G.; Rossi, F.E.; Figueiredo, C.; Caperuto, E.C.; Lira, F.S. Capsaicin supplementation increases time to exhaustion in high-intensity intermittent exercise without modifying metabolic responses in physically active men. Eur. J. Appl. Physiol. 2019, 119, 971–979. [Google Scholar] [CrossRef]
- de Freitas, M.C.; Cholewa, J.M.; Freire, R.V.; Carmo, B.A.; Bottan, J.; Bratfich, M.; Della Bandeira, M.P.; Gonçalves, D.C.; Caperuto, E.C.; Lira, F.S.; et al. Acute Capsaicin Supplementation Improves Resistance Training Performance in Trained Men. J. Strength Cond. Res. 2018, 32, 2227–2232. [Google Scholar] [CrossRef]
- de Freitas, M.C.; Cholewa, J.M.; Gobbo, L.A.; de Oliveira, J.V.N.S.; Lira, F.S.; Rossi, F.E. Acute Capsaicin Supplementation Improves 1500-m Running Time-Trial Performance and Rate of Perceived Exertion in Physically Active Adults. J. Strength Cond. Res. 2018, 32, 572–577. [Google Scholar] [CrossRef]
- von Ah Morano, A.E.; Padilha, C.S.; Soares, V.A.M.; Andrade Machado, F.; Hofmann, P.; Rossi, F.E.; Lira, F.S. Capsaicin Analogue Supplementation Does Not Improve 10 km Running Time-Trial Performance in Male Amateur Athletes: A Randomized, Crossover, Double-Blind and Placebo-Controlled Study. Nutrients 2020, 13, 34. [Google Scholar] [CrossRef] [PubMed]
- Opheim, M.N.; Rankin, J.W. Effect of capsaicin supplementation on repeated sprinting performance. J. Strength Cond. Res. 2012, 26, 319–326. [Google Scholar] [CrossRef]
- Steensberg, A.; van Hall, G.; Osada, T.; Sacchetti, M.; Saltin, B.; Klarlund Pedersen, B. Production of interleukin-6 in contracting human skeletal muscles can account for the exercise-induced increase in plasma interleukin-6. J. Physiol. 2000, 529 Pt 1, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Cannon, J.G.; Fielding, R.A.; Fiatarone, M.A.; Orencole, S.F.; Dinarello, C.A.; Evans, W.J. Increased interleukin 1 beta in human skeletal muscle after exercise. Am. J. Physiol. 1989, 257, R451–R455. [Google Scholar] [CrossRef] [PubMed]
- Roerink, M.E.; van der Schaaf, M.E.; Dinarello, C.A.; Knoop, H.; van der Meer, J.W. Interleukin-1 as a mediator of fatigue in disease: A narrative review. J. Neuroinflamm. 2017, 14, 16. [Google Scholar] [CrossRef]
- Granger, D.A.; Kivlighan, K.T.; el-Sheikh, M.; Gordis, E.B.; Stroud, L.R. Salivary alpha-amylase in biobehavioral research: Recent developments and applications. Ann. N. Y. Acad. Sci. 2007, 1098, 122–144. [Google Scholar] [CrossRef] [PubMed]
- Jacks, D.E.; Sowash, J.; Anning, J.; McGloughlin, T.; Andres, F. Effect of exercise at three exercise intensities on salivary cortisol. J. Strength Cond. Res. 2002, 16, 286–289. [Google Scholar]
- Kobayashi, M.; Watanabe, K.; Yokoyama, S.; Matsumoto, C.; Hirata, M.; Tominari, T.; Inada, M.; Miyaura, C. Capsaicin, a TRPV1 Ligand, Suppresses Bone Resorption by Inhibiting the Prostaglandin E Production of Osteoblasts, and Attenuates the Inflammatory Bone Loss Induced by Lipopolysaccharide. ISRN Pharmacol. 2012, 2012, 439860. [Google Scholar] [CrossRef]
- Tang, J.; Luo, K.; Li, Y.; Chen, Q.; Tang, D.; Wang, D.; Xiao, J. Capsaicin attenuates LPS-induced inflammatory cytokine production by upregulation of LXRα. Int. Immunopharmacol. 2015, 28, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Noreen, E.E.; Lemon, P.W. Reliability of air displacement plethysmography in a large, heterogeneous sample. Med. Sci. Sports Exerc. 2006, 38, 1505–1509. [Google Scholar] [CrossRef]
- Reyes-Escogido, M.D.L.; Gonzalez-Mondragon, E.G.; Vazquez-Tzompantzi, E. Chemical and Pharmacological Aspects of Capsaicin. Molecules 2011, 16, 1253–1270. [Google Scholar] [CrossRef]
- Crouter, S.E.; Antczak, A.; Hudak, J.R.; DellaValle, D.M.; Haas, J.D. Accuracy and reliability of the ParvoMedics TrueOne 2400 and MedGraphics VO2000 metabolic systems. Eur. J. Appl. Physiol. 2006, 98, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Richard, R.; Lonsdorfer-Wolf, E.; Charloux, A.; Doutreleau, S.; Buchheit, M.; Oswald-Mammosser, M.; Lampert, E.; Mettauer, B.; Geny, B.; Lonsdorfer, J. Non-invasive cardiac output evaluation during a maximal progressive exercise test, using a new impedance cardiograph device. Eur. J. Appl. Physiol. 2001, 85, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Laginestra, F.G.; Amann, M.; Kirmizi, E.; Giuriato, G.; Barbi, C.; Ruzzante, F.; Pedrinolla, A.; Martignon, C.; Tarperi, C.; Schena, F.; et al. Electrically induced quadriceps fatigue in the contralateral leg impairs ipsilateral knee extensors performance. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2021, 320, R747–R756. [Google Scholar] [CrossRef]
- Amann, M.; Venturelli, M.; Ives, S.J.; McDaniel, J.; Layec, G.; Rossman, M.J.; Richardson, R.S. Peripheral fatigue limits endurance exercise via a sensory feedback-mediated reduction in spinal motoneuronal output. J. Appl. Physiol. (1985) 2013, 115, 355–364. [Google Scholar] [CrossRef]
- Giuriato, G.; Gundersen, A.; Verma, S.; Pelletier, E.; Bakewell, B.; Ives, S.J. The Effects of Chest Wall Loading on Perceptions of Fatigue, Exercise Performance, Pulmonary Function, and Muscle Perfusion. Sports 2020, 8, 3. [Google Scholar] [CrossRef]
- Ives, S.J.; Amann, M.; Venturelli, M.; Witman, M.A.; Groot, H.J.; Wray, D.W.; Morgan, D.E.; Stehlik, J.; Richardson, R.S. The Mechanoreflex and Hemodynamic Response to Passive Leg Movement in Heart Failure. Med. Sci. Sports Exerc. 2016, 48, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Barrett-O’Keefe, Z.; Ives, S.J.; Trinity, J.D.; Morgan, G.; Rossman, M.J.; Donato, A.J.; Runnels, S.; Morgan, D.E.; Gmelch, B.S.; Bledsoe, A.D.; et al. Taming the “sleeping giant”: The role of endothelin-1 in the regulation of skeletal muscle blood flow and arterial blood pressure during exercise. Am. J. Physiol. Heart Circ. Physiol. 2013, 304, H162–H169. [Google Scholar] [CrossRef]
- Hill, C.A.; Thompson, M.W.; Ruell, P.A.; Thom, J.M.; White, M.J. Sarcoplasmic reticulum function and muscle contractile character following fatiguing exercise in humans. J. Physiol. 2001, 531, 871–878. [Google Scholar] [CrossRef] [PubMed]
- Leppik, J.A.; Aughey, R.J.; Medved, I.; Fairweather, I.; Carey, M.F.; McKenna, M.J. Prolonged exercise to fatigue in humans impairs skeletal muscle Na+-K+-ATPase activity, sarcoplasmic reticulum Ca2+ release, and Ca2+ uptake. J. Appl. Physiol. (1985) 2004, 97, 1414–1423. [Google Scholar] [CrossRef] [PubMed]
- Allen, D.G.; Lamb, G.D.; Westerblad, H. Impaired calcium release during fatigue. J. Appl. Physiol. (1985) 2008, 104, 296–305. [Google Scholar] [CrossRef]
- Pingle, S.C.; Matta, J.A.; Ahern, G.P. Capsaicin Receptor: TRPV1 A Promiscuous TRP Channel. In Transient Receptor Potential (TRP) Channels; Flockerzi, V., Nilius, B., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 155–171. [Google Scholar]
- Santulli, G.; Lewis, D.; des Georges, A.; Marks, A.R.; Frank, J. Ryanodine Receptor Structure and Function in Health and Disease. In Membrane Protein Complexes: Structure and Function; Harris, J.R., Boekema, E.J., Eds.; Springer: Singapore, 2018; pp. 329–352. [Google Scholar]
- Rosa, A.; Deiana, M.; Casu, V.; Paccagnini, S.; Appendino, G.; Ballero, M.; Dessí, M.A. Antioxidant activity of capsinoids. J. Agric. Food Chem. 2002, 50, 7396–7401. [Google Scholar] [CrossRef]
- Vidal, K.; Robinson, N.; Ives, S.J. Exercise performance and physiological responses: The potential role of redox imbalance. Physiol. Rep. 2017, 5, e13225. [Google Scholar] [CrossRef] [PubMed]
- Paschalis, V.; Theodorou, A.A.; Kyparos, A.; Dipla, K.; Zafeiridis, A.; Panayiotou, G.; Vrabas, I.S.; Nikolaidis, M.G. Low vitamin C values are linked with decreased physical performance and increased oxidative stress: Reversal by vitamin C supplementation. Eur. J. Nutr. 2016, 55, 45–53. [Google Scholar] [CrossRef]
- Hureau, T.J.; Weavil, J.C.; Sidhu, S.K.; Thurston, T.S.; Reese, V.R.; Zhao, J.; Nelson, A.D.; Birgenheier, N.M.; Richardson, R.S.; Amann, M. Ascorbate attenuates cycling exercise-induced neuromuscular fatigue but fails to improve exertional dyspnea and exercise tolerance in COPD. J. Appl. Physiol. (1985) 2021, 130, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Rossman, M.J.; Garten, R.S.; Groot, H.J.; Reese, V.; Zhao, J.; Amann, M.; Richardson, R.S. Ascorbate infusion increases skeletal muscle fatigue resistance in patients with chronic obstructive pulmonary disease. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, R1163–R1170. [Google Scholar] [CrossRef]
- Varadharaj, S.; Kelly, O.J.; Khayat, R.N.; Kumar, P.S.; Ahmed, N.; Zweier, J.L. Role of Dietary Antioxidants in the Preservation of Vascular Function and the Modulation of Health and Disease. Front. Cardiovasc. Med. 2017, 4, 64. [Google Scholar] [CrossRef] [PubMed]
- Hoheisel, U.; Reinöhl, J.; Unger, T.; Mense, S. Acidic pH and capsaicin activate mechanosensitive group IV muscle receptors in the rat. Pain 2004, 110, 149–157. [Google Scholar] [CrossRef]
- Sidhu, S.K.; Weavil, J.C.; Mangum, T.S.; Jessop, J.E.; Richardson, R.S.; Morgan, D.E.; Amann, M. Group III/IV locomotor muscle afferents alter motor cortical and corticospinal excitability and promote central fatigue during cycling exercise. Clin. Neurophysiol. 2017, 128, 44–55. [Google Scholar] [CrossRef]
- Liao, S.F.; Korivi, M.; Tsao, J.P.; Huang, C.C.; Chang, C.C.; Cheng, I.S. Effect of Capsinoids Supplementation on Fat Oxidation and Muscle Glycogen Restoration During Post-exercise Recovery in Humans. Curr. Pharm. Des. 2021, 27, 981–988. [Google Scholar] [CrossRef]
- Budde, H.; Machado, S.; Ribeiro, P.; Wegner, M. The cortisol response to exercise in young adults. Front. Behav. Neurosci. 2015, 9, 13. [Google Scholar] [CrossRef] [PubMed]
- Kirschbaum, C.; Hellhammer, D.H. Salivary cortisol in psychoneuroendocrine research: Recent developments and applications. Psychoneuroendocrinology 1994, 19, 313–333. [Google Scholar] [CrossRef]
- Choi, Y.J.; Kim, J.Y.; Yoo, S.B.; Lee, J.H.; Jahng, J.W. Repeated oral administration of capsaicin increases anxiety-like behaviours with prolonged stress-response in rats. J. Biosci. 2013, 38, 561–571. [Google Scholar] [CrossRef]
- Bosch, J.A.; Ring, C.; de Geus, E.J.; Veerman, E.C.; Amerongen, A.V. Stress and secretory immunity. Int. Rev. Neurobiol. 2002, 52, 213–253. [Google Scholar] [CrossRef]
- Magaña-Barajas, E.; Buitimea-Cantúa, G.V.; Hernández-Morales, A.; Torres-Pelayo, V.D.R.; Vázquez-Martínez, J.; Buitimea-Cantúa, N.E. In vitro α-amylase and α-glucosidase enzyme inhibition and antioxidant activity by capsaicin and piperine from Capsicum chinense and Piper nigrum fruits. J. Environ. Sci. Health Part. B Pestic. Food Contam. Agric. Wastes 2021, 56, 282–291. [Google Scholar] [CrossRef]
- Ives, S.J.; Blegen, M.; Coughlin, M.A.; Redmond, J.; Matthews, T.; Paolone, V. Salivary estradiol, interleukin-6 production, and the relationship to substrate metabolism during exercise in females. Eur. J. Appl. Physiol. 2011, 111, 1649–1658. [Google Scholar] [CrossRef] [PubMed]
- Seki, N.; Shirasaki, H.; Kikuchi, M.; Himi, T. Capsaicin induces the production of IL-6 in human upper respiratory epithelial cells. Life Sci. 2007, 80, 1592–1597. [Google Scholar] [CrossRef] [PubMed]
- de Moura, E.S.V.E.L.; Cholewa, J.M.; Jäger, R.; Zanchi, N.E.; de Freitas, M.C.; de Moura, R.C.; Barros, E.M.L.; Antunes, B.M.; Caperuto, E.C.; Ribeiro, S.L.G.; et al. Chronic capsiate supplementation increases fat-free mass and upper body strength but not the inflammatory response to resistance exercise in young untrained men: A randomized, placebo-controlled and double-blind study. J. Int. Soc. Sports Nutr. 2021, 18, 50. [Google Scholar] [CrossRef]
- Kikuchi, H.; Oguri, G.; Yamamoto, Y.; Takano, N.K.; Tanaka, T.; Takahashi, M.; Nakamura, F.; Yamasoba, T.; Komuro, I.; Obi, S.; et al. Thermo-Sensitive Transient Receptor Potential Vanilloid (TRPV) Channels Regulate IL-6 Expression in Mouse Adipocytes. Cardiovasc. Pharmacol. 2015, 4. [Google Scholar] [CrossRef]
- Ghanemi, A.; St-Amand, J. Interleukin-6 as a “metabolic hormone”. Cytokine 2018, 112, 132–136. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Mean ± SD |
|---|---|
| Age (years) | 22.3 ± 3.6 |
| Height (cm) | 182 ± 10 |
| Weight (kg) | 81.3 ± 11.5 |
| Fat Mass (%) | 11.7 ± 1.5 |
| Peak Aerobic Power (Watts) | 340 ± 21 |
| Peak Oxygen Consumption (VO2peak, mL/kg/min) | 49.5 ± 8.1 |
| Baseline | Exercise | 10 min Post | 15 min Post | |||||
|---|---|---|---|---|---|---|---|---|
| PL | CAP | PL | CAP | PL | CAP | PL | CAP | |
| Cortisol (μg/dL) | 0.3 ± 0.2 | 0.3 ± 0.2 | 0.2 ± 0.0 | 0.3 ± 0.1 | 0.3 ± 0.2 | 0.3 ± 0.2 | 0.4 ± 0.2 | 0.4 ± 0.2 |
| α-amylase (U/mL) | 31 ± 36 | 15 ± 10 | 74 ± 41 | 57 ± 67 | 42 ± 29 | 40 ± 28 | 38 ± 27 | 22 ± 8 |
| IL-6 (pg/mL) | 10 ± 7 | 17 ± 14 * | 7 ± 5 | 13 ± 7 * | 8 ± 9 | 10 ± 7 | 8 ± 4 | 9 ± 5 |
| IL-1β (pg/mL) | 20 ± 18 | 21 ± 29 | - | - | 35 ± 40 | 15 ± 11 | 32 ± 28 | 15 ± 10 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giuriato, G.; Venturelli, M.; Matias, A.; Soares, E.M.K.V.K.; Gaetgens, J.; Frederick, K.A.; Ives, S.J. Capsaicin and Its Effect on Exercise Performance, Fatigue and Inflammation after Exercise. Nutrients 2022, 14, 232. https://doi.org/10.3390/nu14020232
Giuriato G, Venturelli M, Matias A, Soares EMKVK, Gaetgens J, Frederick KA, Ives SJ. Capsaicin and Its Effect on Exercise Performance, Fatigue and Inflammation after Exercise. Nutrients. 2022; 14(2):232. https://doi.org/10.3390/nu14020232
Chicago/Turabian StyleGiuriato, Gaia, Massimo Venturelli, Alexs Matias, Edgard M. K. V. K. Soares, Jessica Gaetgens, Kimberley A. Frederick, and Stephen J. Ives. 2022. "Capsaicin and Its Effect on Exercise Performance, Fatigue and Inflammation after Exercise" Nutrients 14, no. 2: 232. https://doi.org/10.3390/nu14020232
APA StyleGiuriato, G., Venturelli, M., Matias, A., Soares, E. M. K. V. K., Gaetgens, J., Frederick, K. A., & Ives, S. J. (2022). Capsaicin and Its Effect on Exercise Performance, Fatigue and Inflammation after Exercise. Nutrients, 14(2), 232. https://doi.org/10.3390/nu14020232