
FTO Common Obesity SNPs Interact with Actionable Environmental Factors: Physical Activity, Sugar-Sweetened Beverages and Wine Consumption



Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. SNP Selection and Hardy–Weinberg Equilibrium (HWE)
2.3. Statistical Analyses
3. Results
3.1. Participants Characteristics
3.2. FTO SNPs Association and Obesity Risk
3.3. Gene Environment Interactions
3.3.1. Physical Activity Interaction with FTO on Obesity Risk
3.3.2. Wine Consumption Interaction with FTO Polymorphism in Determining Obesity Risk
3.3.3. SSBs Consumption Interaction with FTO on Obesity Risk
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Elks, C.E.; den Hoed, M.; Zhao, J.H.; Sharp, S.J.; Wareham, N.J.; Loos, R.J.; Ong, K.K. Variability in the heritability of body mass index: A systematic review and meta-regression. Front. Endocrinol. 2012, 3, 29. [Google Scholar] [CrossRef] [PubMed]
- Rankinen, T.; Zuberi, A.; Chagnon, Y.C.; Weisnagel, S.J.; Argyropoulos, G.; Walts, B.; Pérusse, L.; Bouchard, C. The Human Obesity Gene Map: The 2005 Update. Obesity 2006, 14, 529–644. [Google Scholar] [CrossRef] [PubMed]
- Engin, A. The Definition and Prevalence of Obesity and Metabolic Syndrome. Obes. Lipotoxicity 2017, 960, 1–17. [Google Scholar]
- Markwald, R.R.; Melanson, E.L.; Smith, M.R.; Higgins, J.; Perreault, L.; Eckel, R.H.; Wright, K.P., Jr. Impact of insufficient sleep on total daily energy expenditure, food intake, and weight gain. Proc. Natl. Acad. Sci. USA 2013, 110, 5695–5700. [Google Scholar] [CrossRef] [PubMed]
- Nicolaidis, S. Environment and obesity. Metab. Clin. Exp. 2019, 100, 153942. [Google Scholar] [CrossRef]
- Peters, T.; Ausmeier, K.; Dildrop, R.; Rüther, U. The mouse Fused toes (Ft) mutation is the result of a 1.6-Mb deletion including the entire Iroquois B gene cluster. Mamm. Genome 2002, 13, 186–188. [Google Scholar] [CrossRef]
- Fredriksson, R.; Hägglund, M.; Olszewski, P.K.; Stephansson, O.; Jacobsson, J.A.; Olszewska, A.M.; Levine, A.S.; Lindblom, J.; Schiöth, H.B. The obesity gene, FTO, is of ancient origin, up-regulated during food deprivation and expressed in neurons of feeding-related nuclei of the brain. Endocrinology 2008, 149, 2062–2071. [Google Scholar] [CrossRef]
- Gerken, T.; Girard, C.A.; Tung, Y.L.; Webby, C.J.; Saudek, V.; Hewitson, K.S.; Yeo, G.S.; McDonough, M.A.; Cunliffe, S.; McNeill, L.A.; et al. The Obesity-Associated FTO Gene Encodes a 2-Oxoglutarate–Dependent Nucleic Acid Demethylase. Science 2007, 318, 1469–1472. [Google Scholar] [CrossRef]
- Zhao, X.; Yang, Y.; Sun, B.; Zhao, Y.; Yang, Y. FTO and obesity: Mechanisms of association. Curr. Diab. Rep. 2014, 14, 486. [Google Scholar] [CrossRef]
- Meyre, D.; Delplanque, J.; Chèvre, J.; Lecoeur, C.; Lobbens, S.; Gallina, S.; Durand, E.; Vatin, V.; Degraeve, F.; Proença, C.; et al. Genome-wide association study for early-onset and morbid adult obesity identifies three new risk loci in European populations. Nat. Genet. 2009, 41, 157–159. [Google Scholar] [CrossRef]
- Frayling, T.M.; Timpson, N.J.; Weedon, M.N.; Zeggini, E.; Freathy, R.M.; Lindgren, C.M.; Perry, J.R.; Elliott, K.S.; Lango, H.; Rayner, N.W.; et al. A Common Variant in the FTO Gene Is Associated with Body Mass Index and Predisposes to Childhood and Adult Obesity. Science 2007, 316, 889. [Google Scholar] [CrossRef] [PubMed]
- Cecil, J.E.; Tavendale, R.; Watt, P.; Hetherington, M.M.; Palmer, C.N.A. An obesity-associated FTO gene variant and increased energy intake in children. N. Engl. J. Med. 2008, 359, 2558–2566. [Google Scholar] [CrossRef] [PubMed]
- Park, S.L.; Cheng, I.; Pendergrass, S.A.; Kucharska-Newton, A.M.; Lim, U.; Ambite, J.L.; Caberto, C.P.; Monroe, K.R.; Schumacher, F.; Hindorff, L.A.; et al. Association of the FTO Obesity Risk Variant rs8050136 With Percentage of Energy Intake from Fat in Multiple Racial/Ethnic Populations: The PAGE Study. Am. J. Epidemiol. 2013, 178, 780. [Google Scholar] [CrossRef] [PubMed]
- Wardle, J.; Llewellyn, C.; Sanderson, S.; Plomin, R. The FTO gene and measured food intake in children. Int. J. Obes. 2009, 33, 42–45. [Google Scholar] [CrossRef] [PubMed]
- Brunkwall, L.; Ericson, U.; Hellstrand, S.; Gullberg, B.; Orho-Melander, M.; Sonestedt, E. Genetic variation in the fat mass and obesity-associated gene (FTO) in association with food preferences in healthy adults. Food Nutr. Res. 2013, 57, 20028. [Google Scholar] [CrossRef] [PubMed]
- Loos, R.J.F.; Yeo, G.S.H. The bigger picture of FTO—The first GWAS-identified obesity gene. Nature reviews. Endocrinology 2014, 10, 51–61. [Google Scholar] [PubMed]
- Croteau-Chonka, D.C.; Marvelle, A.F.; Lange, E.M.; Lee, N.R.; Adair, L.S.; Lange, L.A.; Mohlke, K.L. Genome-wide association study of anthropometric traits and evidence of interactions with age and study year in Filipino women. Obesity 2011, 19, 1019–1027. [Google Scholar] [CrossRef]
- Hotta, K.; Nakata, Y.; Matsuo, T.; Kamohara, S.; Kotani, K.; Komatsu, R.; Itoh, N.; Mineo, I.; Wada, J.; Masuzaki, H.; et al. Variations in the FTO gene are associated with severe obesity in the Japanese. J. Hum. Genet. 2008, 53, 546–553. [Google Scholar] [CrossRef]
- Tan, J.T.; Dorajoo, R.; Seielstad, M.; Sim, X.L.; Ong, R.T.; Chia, K.S.; Wong, T.Y.; Saw, S.M.; Chew, S.K.; Aung, T.; et al. FTO Variants Are Associated With Obesity in the Chinese and Malay Populations in Singapore. Diabetes 2008, 57, 2851–2857. [Google Scholar] [CrossRef]
- Tan, L.; Zhu, H.; He, H.; Wu, K.; Li, J.; Chen, X.; Zhang, J.G.; Shen, H.; Tian, Q.; Krousel-Wood, M.; et al. Replication of 6 obesity genes in a meta-analysis of genome-wide association studies from diverse ancestries. PLoS ONE 2014, 9, e96149. [Google Scholar] [CrossRef]
- Dastgheib, S.A.; Bahrami, R.; Setayesh, S.; Salari, S.; Mirjalili, S.R.; Noorishadkam, M.; Sadeghizadeh-Yazdi, J.; Akbarian, E.; Neamatzadeh, H. Evidence from a meta-analysis for association of MC4R rs17782313 and FTO rs9939609 polymorphisms with susceptibility to obesity in children. Diabetes Metab. Syndr. 2021, 15, 102234. [Google Scholar] [CrossRef] [PubMed]
- Hester, J.M.; Wing, M.R.; Li, J.; Palmer, N.D.; Xu, J.; Hicks, P.J.; Roh, B.H.; Norris, J.M.; Wagenknecht, L.E.; Langefeld, C.D.; et al. Implication of European-derived adiposity loci in African Americans. Int. J. Obes. 2012, 36, 465–473. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Li, H.; Wu, Y.; Loos, R.J.F.; Hu, F.B.; Liu, Y.; Wang, J.; Yu, Z.; Lin, X. Variants in the fat mass- and obesity-associated (FTO) gene are not associated with obesity in a Chinese Han population. Diabetes 2008, 57, 264–268. [Google Scholar] [CrossRef] [PubMed]
- Kilpeläinen, T.O.; Qi, L.; Brage, S.; Sharp, S.J.; Sonestedt, E.; Demerath, E.; Ahmad, T.; Mora, S.; Kaakinen, M.; Sandholt, C.H.; et al. Physical activity attenuates the influence of FTO variants on obesity risk: A meta-analysis of 218,166 adults and 19,268 children. PLoS Med. 2011, 8, e1001116. [Google Scholar] [CrossRef]
- Kim, J.Y.; DeMenna, J.T.; Puppala, S.; Chittoor, G.; Schneider, J.; Duggirala, R.; Mandarino, L.J.; Shaibi, G.Q.; Coletta, D.K. Physical activity and FTO genotype by physical activity interactive influences on obesity. BMC Genet. 2016, 17, 47. [Google Scholar] [CrossRef]
- Qi, Q.; Chu, A.Y.; Kang, J.H.; Jensen, M.K.; Curhan, G.C.; Pasquale, L.R.; Ridker, P.M.; Hunter, D.J.; Willett, W.C.; Rimm, E.B.; et al. Sugar-Sweetened Beverages and Genetic Risk of Obesity. N. Engl. J. Med. 2012, 367, 1387–1396. [Google Scholar] [CrossRef]
- Haupt, A.; Thamer, C.; Staiger, H.; Tschritter, O.; Kirchhoff, K.; Machicao, F.; Häring, H.U.; Stefan, N.; Fritsche, A. Variation in the FTO gene influences food intake but not energy expenditure. Exp. Clin. Endocrinol. Diabetes 2009, 117, 194–197. [Google Scholar] [CrossRef]
- Xiang, L.; Wu, H.; Pan, A.; Patel, B.; Xiang, G.; Qi, L.; Kaplan, R.C.; Hu, F.; Wylie-Rosett, J.; Qi, Q. FTO genotype and weight loss in diet and lifestyle interventions: A systematic review and meta-analysis. Am. J. Clin. Nutr. 2016, 10, 1162–1170. [Google Scholar] [CrossRef]
- Weir, C.B.; Jan, A. BMI Classification Percentile and Cut Off Points; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Thorleifsson, G.; Walters, G.B.; Gudbjartsson, D.F.; Steinthorsdottir, V.; Sulem, P.; Helgadottir, A.; Styrkarsdottir, U.; Gretarsdottir, S.; Thorlacius, S.; Jonsdottir, I.; et al. Genome-wide association yields new sequence variants at seven loci that associate with measures of obesity. Nat. Genet. 2009, 41, 18–24. [Google Scholar] [CrossRef]
- Hinney, A.; Nguyen, T.T.; Scherag, A.; Friedel, S.; Brönner, G.; Müller, T.D.; Grallert, H.; Illig, T.; Wichmann, H.E.; Rief, W.; et al. Genome wide association (GWA) study for early onset extreme obesity supports the role of fat mass and obesity associated gene (FTO) variants. PLoS ONE 2007, 2, e1361. [Google Scholar] [CrossRef]
- Scuteri, A.; Sanna, S.; Chen, W.; Uda, M.; Albai, G.; Strait, J.; Najjar, S.; Nagaraja, R.; Orrú, M.; Usala, G.; et al. Genome-Wide Association Scan Shows Genetic Variants in the FTO Gene Are Associated with Obesity-Related Traits. PLoS Genet. 2007, 3, e115. [Google Scholar] [CrossRef] [PubMed]
- Willer, C.J.; Speliotes, E.K.; Loos, R.J.F.; Li, S.; Lindgren, C.M.; Heid, I.M.; Berndt, S.I.; Elliott, A.L.; Jackson, A.U.; Lamina, C.; et al. Six new loci associated with body mass index highlight a neuronal influence on body weight regulation. Nat. Genet. 2009, 41, 25–34. [Google Scholar] [PubMed]
- Qi, Q.; Kilpeläinen, T.O.; Downer, M.K.; Tanaka, T.; Smith, C.E.; Sluijs, I.; Sonestedt, E.; Chu, A.Y.; Renström, F.; Lin, X.; et al. FTO genetic variants, dietary intake and body mass index: Insights from 177 330 individuals. Hum. Mol. Genet. 2014, 23, 6961–6972. [Google Scholar] [CrossRef] [PubMed]
- MacArthur, J.; Bowler, E.; Cerezo, M.; Gil, L.; Hall, P.; Hastings, E.; Junkins, H.; McMahon, A.; Milano, A.; Morales, J.; et al. The new NHGRI-EBI Catalog of published genome-wide association studies (GWAS Catalog). Nucleic Acids Res. 2017, 45, D896–D901. [Google Scholar] [CrossRef]
- Kang, H. Sample size determination and power analysis using the G*Power software. J. Educ. Eval. Health Prof. 2021, 18, 17. [Google Scholar] [CrossRef]
- Rampersaud, E.; Mitchell, B.D.; Pollin, T.I.; Fu, M.; Shen, H.; O’Connell, J.R.; Ducharme, J.L.; Hines, S.; Sack, P.; Naglieri, R.; et al. Physical activity and the association of common FTO gene variants with body mass index and obesity. Arch. Intern. Med. 2008, 168, 1791–1797. [Google Scholar] [CrossRef]
- Liaw, Y.; Liaw, Y.; Lan, T. Physical Activity Might Reduce the Adverse Impacts of the FTO Gene Variant rs3751812 on the Body Mass Index of Adults in Taiwan. Genes 2019, 10, 354. [Google Scholar] [CrossRef]
- Jonsson, A.; Renström, F.; Lyssenko, V.; Brito, E.C.; Isomaa, B.; Berglund, G.; Nilsson, P.M.; Groop, L.; Franks, P.W. Assessing the effect of interaction between an FTO variant (rs9939609) and physical activity on obesity in 15,925 Swedish and 2,511 Finnish adults. Diabetologia 2009, 52, 1334–1338. [Google Scholar] [CrossRef][Green Version]
- Leońska-Duniec, A.; Jastrzębski, Z.; Zarębska, A.; Maciejewska, A.; Ficek, K.; Cięszczyk, P. Assessing effect of interaction between the FTO A/T polymorphism (rs9939609) and physical activity on obesity-related traits. J. Sport Health Sci. 2018, 7, 459–464. [Google Scholar] [CrossRef]
- Andreasen, C.H.; Stender-Petersen, K.L.; Mogensen, M.S.; Torekov, S.S.; Wegner, L.; Andersen, G.; Nielsen, A.L.; Albrechtsen, A.; Borch-Johnsen, K.; Rasmussen, S.S.; et al. Low physical activity accentuates the effect of the FTO rs9939609 polymorphism on body fat accumulation. Diabetes 2008, 57, 95–101. [Google Scholar] [CrossRef]
- Vimaleswaran, K.S.; Li, S.; Zhao, J.H.; Luan, J.; Bingham, S.A.; Khaw, K.; Ekelund, U.; Wareham, N.J.; Loos, R.J. Physical activity attenuates the body mass index-increasing influence of genetic variation in the FTO gene. Am. J. Clin. Nutr. 2009, 90, 425–428. [Google Scholar] [CrossRef] [PubMed]
- Hubacek, J.A.; Adamkova, V.; Dlouha, D.; Jirsa, M.; Šperl, J.; Tönjes, A.; Kovacs, P.; Pikhart, H.; Peasey, A.; Bobak, M. Fat mass and obesity-associated (fto) gene and alcohol intake. Addiction 2012, 107, 1185–1186. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, X.; Luo, X.; Zeng, M.; Zuo, L.; Wang, K. Genetic variants in the fat mass- and obesity-associated (FTO) gene are associated with alcohol dependence. J. Mol. Neurosci. 2013, 51, 416–424. [Google Scholar] [CrossRef] [PubMed]
- DeSalvo, K.B.; Olson, R.; Casavale, K.O. Dietary Guidelines for Americans. JAMA 2016, 315, 457–458. [Google Scholar] [CrossRef]
- Duncan, B.B.; Chambless, L.E.; Schmidt, M.I.; Folsom, A.R.; Szklo, M.; Crouse, J.R.; Carpenter, M.A. Association of the waist-to-hip ratio is different with wine than with beer or hard liquor consumption. Atherosclerosis Risk in Communities Study Investigators. Am. J. Epidemiol. 1995, 142, 1034–1038. [Google Scholar] [CrossRef] [PubMed]
- Lapidus, L.; Bengtsson, C.; Hällström, T.; Björntorp, P. Obesity, adipose tissue distribution and health in women--results from a population study in Gothenburg, Sweden. Appetite 1989, 13, 25–35. [Google Scholar] [CrossRef]
- Slattery, M.L.; McDonald, A.; Bild, D.E.; Caan, B.J.; Hilner, J.E.; Jacobs, D.R.; Liu, K. Associations of body fat and its distribution with dietary intake, physical activity, alcohol, and smoking in blacks and whites. Am. J. Clin. Nutr. 1992, 55, 943–949. [Google Scholar] [CrossRef]
- Snopek, L.; Mlcek, J.; Sochorova, L.; Baron, M.; Hlavacova, I.; Jurikova, T.; Kizek, R.; Sedlackova, E.; Sochor, J. Contribution of Red Wine Consumption to Human Health Protection. Molecules 2018, 23, 1684. [Google Scholar] [CrossRef]
- Golan, R.; Gepner, Y.; Shai, I. Wine and Health-New Evidence. Eur. J. Clin. Nutr. 2019, 72, 55–59. [Google Scholar] [CrossRef]
- Thomson, C.A.; Wertheim, B.C.; Hingle, M.; Wang, L.; Neuhouser, M.L.; Gong, Z.; Garcia, L.; Stefanick, M.L.; Manson, J.E. Alcohol consumption and body weight change in postmenopausal women: Results from the Women’s Health Initiative. Int. J. Obes. 2012, 36, 1158–1164. [Google Scholar] [CrossRef]
- Inan-Eroglu, E.; Powell, L.; Hamer, M.; O’Donovan, G.; Duncan, M.J.; Stamatakis, E. Is There a Link between Different Types of Alcoholic Drinks and Obesity? An Analysis of 280,183 UK Biobank Participants. Int. J. Environ. Res. Public Health 2020, 17, 5178. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.P.; Singh, R.; Verma, S.S.; Rai, V.; Kaschula, C.H.; Maiti, P.; Gupta, S.C. Health benefits of resveratrol: Evidence from clinical studies. Med. Res. Rev. 2019, 39, 1851–1891. [Google Scholar] [CrossRef] [PubMed]
- Wahab, A.; Gao, K.; Jia, C.; Zhang, F.; Tian, G.; Murtaza, G.; Chen, J. Significance of Resveratrol in Clinical Management of Chronic Diseases. Molecules 2017, 22, 1329. [Google Scholar] [CrossRef] [PubMed]
- Malik, V.S.; Schulze, M.B.; Hu, F.B. Intake of sugar-sweetened beverages and weight gain: A systematic review. Am. J. Clin. Nutr. 2006, 84, 274–288. [Google Scholar] [CrossRef] [PubMed]
- Dang, L.C.; Samanez-Larkin, G.R.; Smith, C.T.; Castrellon, J.J.; Perkins, S.F.; Cowan, R.L.; Claassen, D.O.; Zald, D.H. FTO affects food cravings and interacts with age to influence age-related decline in food cravings. Physiol. Behav. 2018, 192, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Hwang, L.; Lin, C.; Gharahkhani, P.; Cuellar-Partida, G.; Ong, J.; An, J.; Gordon, S.D.; Zhu, G.; MacGregor, S.; Lawlor, D.A.; et al. New insight into human sweet taste: A genome-wide association study of the perception and intake of sweet substances. Am. J. Clin. Nutr. 2019, 109, 1724–1737. [Google Scholar] [CrossRef]
- Kong, X.; Hong, J.; Chen, Y.; Chen, L.; Zhao, Z.; Li, Q.; Ge, J.; Chen, G.; Guo, X.; Lu, J.; et al. Association of Genetic Variants with Isolated Fasting Hyperglycaemia and Isolated Postprandial Hyperglycaemia in a Han Chinese Population. PLoS ONE 2013, 8, e71399. [Google Scholar] [CrossRef]
- Shimaoka, I.; Kamide, K.; Ohishi, M.; Katsuya, T.; Akasaka, H.; Saitoh, S.; Sugimoto, K.; Oguro, R.; Congrains, A.; Fujisawa, T.; et al. Association of gene polymorphism of the fat-mass and obesity-associated gene with insulin resistance in Japanese. Hypertens. Res. 2010, 33, 214–218. [Google Scholar] [CrossRef]
- Chauhan, G.; Tabassum, R.; Mahajan, A.; Dwivedi, O.P.; Mahendran, Y.; Kaur, I.; Nigam, S.; Dubey, H.; Varma, B.; Madhu, S.V.; et al. Common variants of FTO and the risk of obesity and type 2 diabetes in Indians. J. Hum. Genet. 2011, 56, 720–726. [Google Scholar] [CrossRef]
- Grzeszczak, W.; Molsa, M.; Tłuczykont, M.; Markowicz, A.; Swoboda, R.; Biedak, M.; Kałuża, A.; Sirek, S.; Strojek, K. The age of developing diabetes and FTO polymorphisms (rs9939609, rs1421085, and rs9930506). Endokrynol. Pol. 2017, 68, 402–406. [Google Scholar] [CrossRef]
- Lewin-Epstein, N.; Cohen, Y. Ethnic origin and identity in the Jewish population of Israel. J Ethn Migr Stud 2018, 45, 2118–2137. [Google Scholar] [CrossRef]
- Sentinelli, F.; Incani, M.; Coccia, F.; Capoccia, D.; Cambuli, V.M.; Romeo, S.; Cossu, E.; Cavallo, M.G.; Leonetti, F.; Baroni, M.G. Association of FTO Polymorphisms with Early Age of Obesity in Obese Italian Subjects. Exp. Diabetes Res. 2012, 2012, 872176. [Google Scholar] [CrossRef] [PubMed]
- Corella, D.; Carrasco, P.; Sorlí, J.V.; Coltell, O.; Ortega-Azorín, C.; Guillén, M.; González, J.I.; Sáiz, C.; Estruch, R.; Ordovas, J.M. Education modulates the association of the FTO rs9939609 polymorphism with body mass index and obesity risk in the Mediterranean population. Nutr. Metab. Cardiovasc. Dis. 2012, 22, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Zhu, Y.; Xu, F.; Ren, X.; Li, X.; Lai, M. FTO gene polymorphisms and obesity risk: A meta-analysis. BMC Med. 2011, 9, 71. [Google Scholar] [CrossRef]
- Maukonen, M.; Männistö, S.; Tolonen, H. A comparison of measured versus self-reported anthropometrics for assessing obesity in adults: A literature review. Scand. J. Public Health 2018, 46, 565–579. [Google Scholar] [CrossRef]


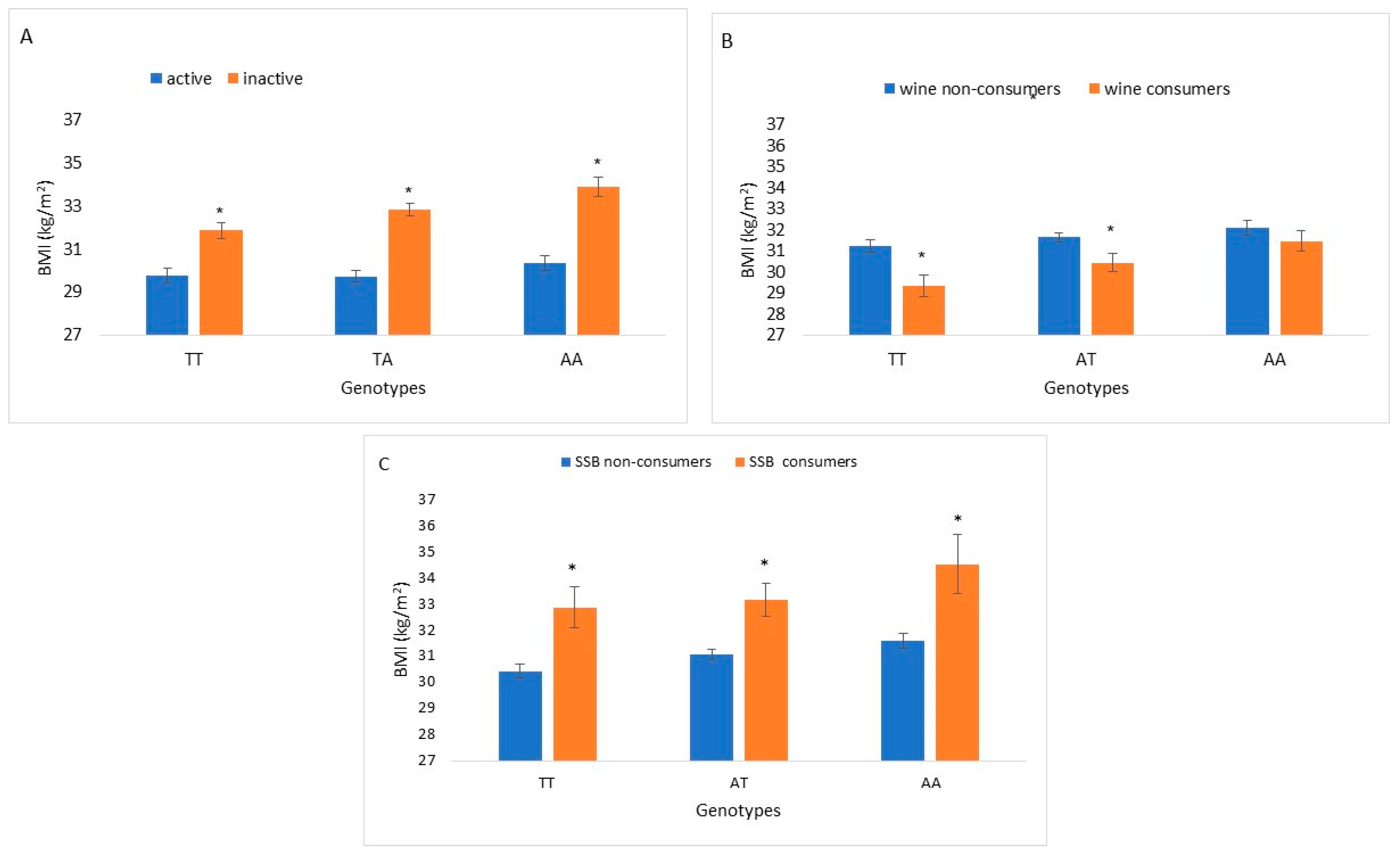{kind=link}
| All Population n = 1972 | Obese (BMI ≥ 30) n = 1098 | Non-Obese (BMI < 30) n = 874 | p-Value | |
|---|---|---|---|---|
| Gender (women, %) | 1377 (79%) | 721 (65.7%) | 650 (74.4%) | <0.001 |
| Age (mean ± SD) | 55.22 ± 14.36 | 54.97 ± 14.54 | 55.53 ± 14.14 | 0.6 |
| Weight (mean ± SD) | 87.32 ± 19.28 | 98.45 ± 17.25 | 74.34 ± 10.69 | <0.001 |
| Hight (mean ± SD) | 166.82 ±8.66 | 167.16 ± 8.99 | 166.38 ± 8.21 | 0.057 |
| BMI (mean ± SD) | 31.25 ± 5.82 | 35.11 ± 4.66 | 26.41 ± 2.65 | <0.001 |
| Physically active (n, %) * | 908 (46%) | 407 (37%) | 501 (57.3%) | <0.001 |
| Smoking (n, %) ** | 190 (9.6%) | 97 (8.8%) | 93 (10.6%) | 0.279 |
| SSB consumers (n, %) ** | 206 (10.4%) | 135 (6.8%) | 71 (3.6%) | 0.002 |
| Wine consumers (n, %) *** | 385 (21.4%) | 189 (18.8%) | 196 (24.7%) | <0.001 |
| Genotype Frequency (%) | p-Value OR ± 95% (CI) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| SNP | Allele | Overall Population (n = 1972) | Obese (n = 1098) | Non-Obese (n = 874) | Dominant Model | Recessive Model | Additive Model | Codominant Model | |
| rs9939609 | T > A | TT AT AA | 28.2% 46.9% 24.8% | 26% 47% 27% | 31% 46.8% 22.2% | 0.014 1.28 (1.05–1.56) | 0.014 1.3 (1.05–1.60) | 0.003 1.2 (1.07–1.36) | 0.003 1.46 (1.14–1.87) |
| rs1421085 | T > C | TT TC CC | 26.7% 47.8% 25.5% | 25% 47.4% 27.6% | 28.7% 48.4% 22.9% | 0.077 1.2 (0.98–1.47) | 0.016 1.29 (1.05–1.59) | 0.011 1.17 (1.04–1.33) | 0.01 1.38 (1.08–1.78) |
| rs8050136 | C > A | CC CA AA | 28.2% 46.6% 25.2% | 26% 46.4% 27.5% | 30.9% 46.8% 22.3% | 0.018 1.27 (1.04–1.55) | 0.008 1.33 (1.07–1.63) | 0.002 1.2 (1.07–1.37) | 0.002 1.47 (1.15–1.88) |
| rs8051591a | A > G | AA AG GG | 27.3% 47.7% 25% | 25.5% 47.1% 27.4% | 29.7% 48.4% 21.9% | 0.058 1.23 (0.99–1.53) | 0.011 1.34 (1.07–1.68) | 0.007 1.2 (1.05–1.38) | 0.007 1.45 (1.1–1.9) |
| rs3751812 a | G > T | GG GT TT | 28.8% 47.6% 23.6% | 27% 47.2% 25.8% | 31% 48.1% 20.9% | 0.074 1.21 (0.98–1.5) | 0.019 1.32 (1.05–1.66) | 0.012 1.19 (1.04–1.36) | 0.01 1.42 (1.09–1.86) |
| rs9935401 a | G > A | GG GA AA | 27.3% 47.9% 24.8% | 25.5% 47.2% 27.3% | 29.7% 48.7% 21.7% | 0.058 1.23 (0.99–1.53) | 0.009 1.35 (1.08–1.7) | 0.006 1.2 (1.06–1.38) | 0.005 1.47 (1.12–1.92) |
| rs11075989 a | C > T | CC CT TT | 27.2% 48% 24.8% | 25.2% 47.8% 27.1% | 29.8% 48.3% 21.9% | 0.033 1.27 (1.02–1.57) | 0.016 1.32 (1.05–1.66) | 0.006 1.2 (1.06–1.38) | 0.006 1.47 (1.12–1.92) |
| rs9923233 a | G > C | GG GC CC | 27.4% 47.7% 24.9% | 25.4% 47.4% 27.2% | 29.9% 48.1% 21.9% | 0.038 1.26 (1.01–1.56) | 0.014 1.33 (1.06–1.66) | 0.006 1.2 (1.06–1.38) | 0.006 1.46 (1.12–1.92) |
| rs9936385 a | T > C | TT TC CC | 27.4% 47.8% 24.8% | 25.4% 47.4% 27.2% | 29.9% 48.3% 21.8% | 0.038 1.26 (1.01–1.56) | 0.012 1.34 (1.07–1.68) | 0.005 1.2 (1.06–1.39) | 0.005 1.47 (1.13–1.93) |
| rs17817964 a | C > T | CC CT TT | 28.8% 47.3% 23.8% | 27.1% 46.8% 26.1% | 31% 48% 21% | 0.086 1.2 (0.97–1.49) | 0.016 1.33 (1.06–1.67) | 0.013 1.19 (1.04–1.36) | 0.011 1.42 (1.09–1.86) |
| rs8043757 a | A > T | AA AT TT | 27.3% 47.7% 24.9% | 25.4% 47.2% 27.4% | 29.8% 48.4% 21.8% | 0.045 1.25 (1–1.55) | 0.009 1.35 (1.08–1.7) | 0.005 1.2 (1.06–1.39) | 0.005 1.47 (1.13–1.93) |
| rs1121980 a | G > A | GG GA AA | 21.1% 48.5% 28.4% | 21.3% 48.2% 30.5% | 25.4% 48.8% 25.8% | 0.06 1.24 (0.99–1.56) | 0.037 1.26 (1.01–1.56) | 0.015 1.18 (1.03–1.35) | 0.016 1.4 (1.07–1.83) |
| rs17817449 a | T > G | TT TG GG | 27.6% 47.5% 24.9% | 25.7% 47% 27.3% | 30.1% 48.1% 21.8% | 0.5 1.24 (1–1.54) | 0.011 1.34 (1.07–1.68) | 0.006 1.2 (1.06–1.38) | 0.005 1.46 (1.12–1.92) |
| rs62033400 a | A > G | AA AG GG | 28.8% 47.6% 23.6% | 27% 47% 26% | 31% 48.4% 20.6% | 0.077 1.21 (0.98–1.5) | 0.011 1.35 (1.07–1.7) | 0.009 1.2 (1.05–1.37) | 0.007 1.45 (1.1–1.9) |
| rs7202116 a | A > G | AA AG GG | 27.3% 47.7% 25% | 25.3% 47.5% 27.2% | 29.9% 47.9% 22.2% | 0.033 1.26 (1.02–1.57) | 0.02 1.3 (1.04–1.64) | 0.007 1.2 (1.05–1.38) | 0.007 1.45 (1.1–1.9) |
| rs7193144 a | T > C | TT TC CC | 27.3% 47.8% 24.9% | 25.4% 47.3% 27.3% | 29.8% 48.4% 21.8% | 0.045 1.25 (1–1.55) | 0.011 1.34 (1.07–1.69) | 0.006 1.2 (1.05–1.38) | 0.005 1.47 (1.12–1.92) |
| rs11075990 a | A > G | AA AG GG | 27.2% 48% 24.8% | 25.2% 47.8% 27.1% | 29.8% 48.3% 21.9% | 0.033 1.27 (1.19–1.57) | 0.016 1.32 (1.53–1.66) | 0.006 1.2 (1.06–1.38) | 0.006 1.46 (1.12–1.92) |
| Rs6499640 a | G > A | GG AG AA | 17.3% 47.9% 34.8% | 19.2% 45.8% 35% | 14.8% 50.5% 34.6% | 0.022 0.74 (0.57–0.96) | 0.842 1.02 (0.83–1.25) | 0.269 0.93 (0.81–1.06) | 0.09 0.78 (0.58–1.05) |
| Rs13333228 b | T > C | TT TC CC | 5.5% 30.6% 64% | 4.9% 29% 66.1% | 6.2% 32.5% 61.3% | 0.217 1.3 (0.86–1.99) | 0.034 1.24 (1.2–1.52) | 0.028 1.2 (1.02–1.41) | 0.129 1.4 (0.9–2.14) |
| rs1558902 b | T > A | TT TA AA | 25.9% 48.5% 25.6% | 24.5% 47.7% 27.8% | 27.7% 49.5% 22.8% | 0.166 1.17 (0.94–1.46) | 0.019 1.3 (1.04–1.63) | 0.023 1.17 (1.02–1.34) | 0.022 1.37 (1.05–1.8) |
| rs9302652 b | C > T | CC CT TT | 9.8% 45.7% 44.5% | 9.4% 45.5% 45.1% | 10.3% 45.9% 43.8% | 0.502 1.12 (0.81–1.54) | 0.384 1.3 (0.99–1) | 0.477 1.06 (0.91–1.22) | 0.456 1.14 (0.81–1.6) |
| Total (n = 1972) | TT (n = 506) | TA (n = 843) | AA (n = 447) | p-Value | |
|---|---|---|---|---|---|
| Age | 55.22 ± 14.36 | 54.1 ± 14.5 | 55.5 ± 14 | 55.9 ± 14.8 | 0.1 |
| BMI | 31.25 ± 5.82 | 30.66 ± 5.74 | 31.3 ± 5.68 | 31.86 ± 5.74 | 0.004 |
| Hight | 166.82 ± 8.66 | 166.84 ± 8.33 | 166.6 ± 8.82 | 167.20 ± 8.76 | 0.455 |
| Weight | 87.32 ± 19.28 | 85.72 ± 19.23 | 87.18 ± 18.79 | 89.4 ± 20.11 | 0.008 |
| Obese (BMI > 30), n, (%) | 1098 (55.7%) | 286 (51.3%) | 516 (55.8%) | 296 (60.4%) | 0.013 |
| Physically active n, (%) | 908 (46%) | 255 (50.4%) | 413 (49%) | 240 (53.7%) | 0.38 |
| SSB consuming n, (%) | 206 (10.5%) | 70 (13.8%) | 88 (10.4%) | 48 (10.7%) | 0.16 |
| Wine consuming n, (%) | 385 (19.5%) | 109 (22.1%) | 175 (21.4%) | 101 (22.9%) | 0.98 |
| Physically Inactive | Physically Active | |||||
|---|---|---|---|---|---|---|
| β | Obesity (BMI > 30) OR ± CI | p-Value | β | Obesity (BMI > 30) OR ± CI | p-Value | |
| rs9939609 AA + TA vs. TT | 0.336 | 1.4 (1–1.93) | 0.04 | 0.1 | 1.1 (0.82–0.86) | 0.4 |
|
rs9939609 AA vs. TA + TT | 0.53 | 1.78 (1.23–2.57) | 0.002 | 0.13 | 1.14 (0.84–0.86) | 0.4 |
|
rs9939609 AA vs. TT | 0.174 | 1.99 (1.3–3.05) | 0.002 | 0.174 | 1.19 (0.83–1.7) | 0.34 |
| Physical Inactivity | Wine Consumption | |||||
|---|---|---|---|---|---|---|
| β | OR ± CI | p-Value | β | OR ± CI | p-Value | |
| rs9939609 TT | 0.826 | 2.29 (1.58–3.3) | <0.001 | −0.582 | 0.56 (0.36–0.87) | 0.009 |
| rs9939609 TA | 0.934 | 2.54 (1.91–3.39) | <0.001 | −0.554 | 0.58 (0.4–0.8) | <0.001 |
| rs9939609 AA + TA | 1.048 | 2.86 (2.25–3.6) | <0.001 | −0.386 | 0.68 (0.52–0.89) | <0.005 |
| rs9939609 AA | 1.326 | 3.77 (2.47–5.75) | <0.001 | −0.085 | 0.92 (0.58–1.45) | 0.715 |
| SNP n = 1746 | Obesity Risk (Interaction with SSB) OR ± CI | p-Value |
|---|---|---|
| rs9939609 | 2.23 ± (1.33–3.76) | 0.002 |
| rs8050136 | 2.23 ± (1.33–3.75) | 0.002 |
| rs1421085 | 2.3 ± (1.37–3.85) | 0.002 |
| rs1121980 | 1.9 ± (1.12–3.31) | 0.019 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chermon, D.; Birk, R. FTO Common Obesity SNPs Interact with Actionable Environmental Factors: Physical Activity, Sugar-Sweetened Beverages and Wine Consumption. Nutrients 2022, 14, 4202. https://doi.org/10.3390/nu14194202
Chermon D, Birk R. FTO Common Obesity SNPs Interact with Actionable Environmental Factors: Physical Activity, Sugar-Sweetened Beverages and Wine Consumption. Nutrients. 2022; 14(19):4202. https://doi.org/10.3390/nu14194202
Chicago/Turabian StyleChermon, Danyel, and Ruth Birk. 2022. "FTO Common Obesity SNPs Interact with Actionable Environmental Factors: Physical Activity, Sugar-Sweetened Beverages and Wine Consumption" Nutrients 14, no. 19: 4202. https://doi.org/10.3390/nu14194202
APA StyleChermon, D., & Birk, R. (2022). FTO Common Obesity SNPs Interact with Actionable Environmental Factors: Physical Activity, Sugar-Sweetened Beverages and Wine Consumption. Nutrients, 14(19), 4202. https://doi.org/10.3390/nu14194202