
Phenolic Release during In Vitro Digestion of Cold and Hot Extruded Noodles Supplemented with Starch and Phenolic Extracts

 ,
, 

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Chemical Extraction
2.3. Preparation of Extruded Noodles
2.4. X-ray Diffraction (XRD) Analysis
2.5. Simulated In Vitro Digestion Analysis
2.6. Morphology
2.7. Content of Unbound, Bound, Total Phenolics
2.8. Profiles of Unbound, Bound, Total Phenolics
2.9. Bioaccessibility Index
2.10. Antioxidant Activity
2.11. Statistical Analysis
3. Results and Discussion
3.1. Content and Profiles of Phenolics of Raw Starch and Phenolic Extract
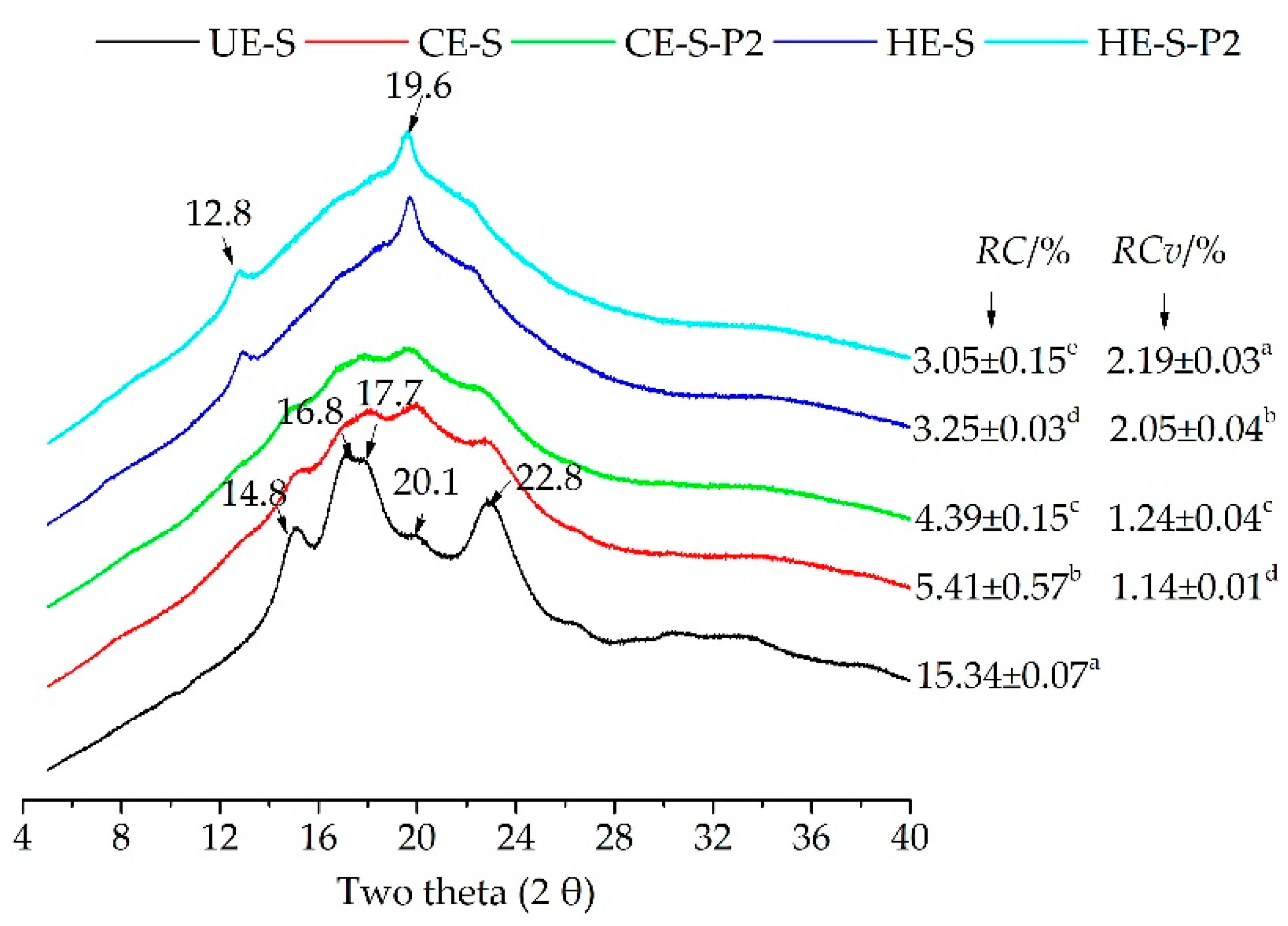
3.2. Characteristics of Starch-Phenolic Complexes Induced by Extrusion
3.3. Simulated In Vitro Digestion of Extruded Noodles
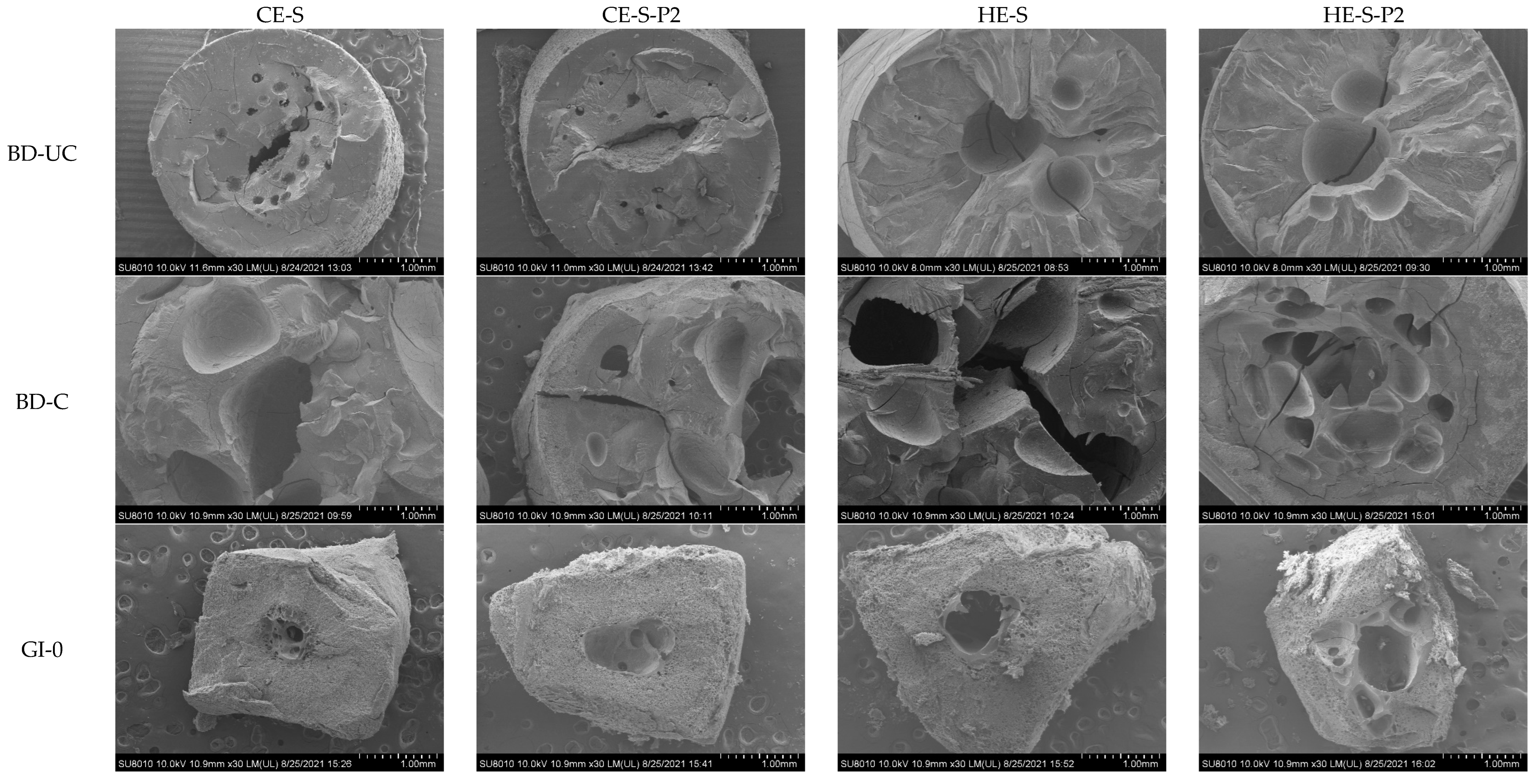
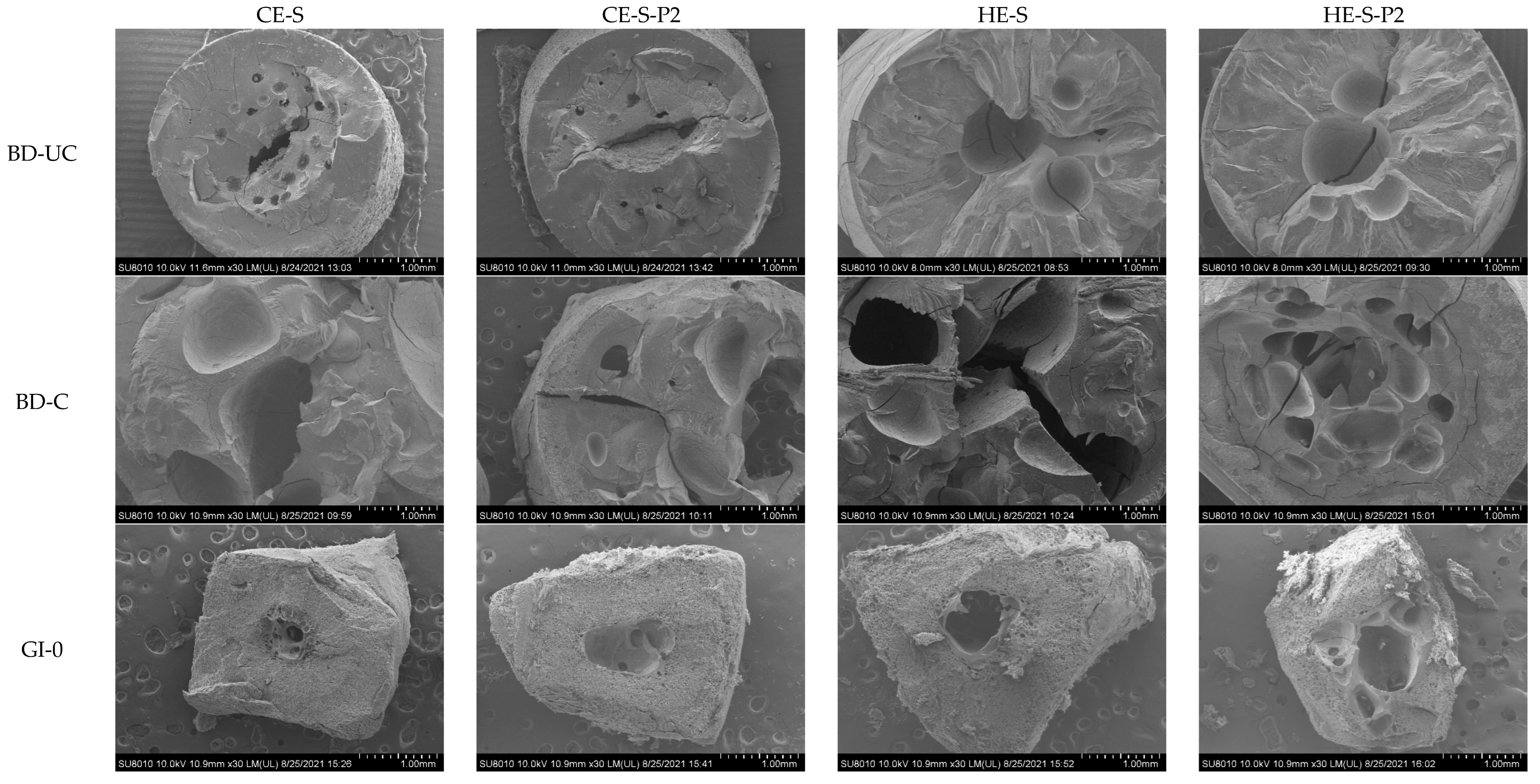
3.4. Morphology of Extruded Noodles and Residues during Digestion
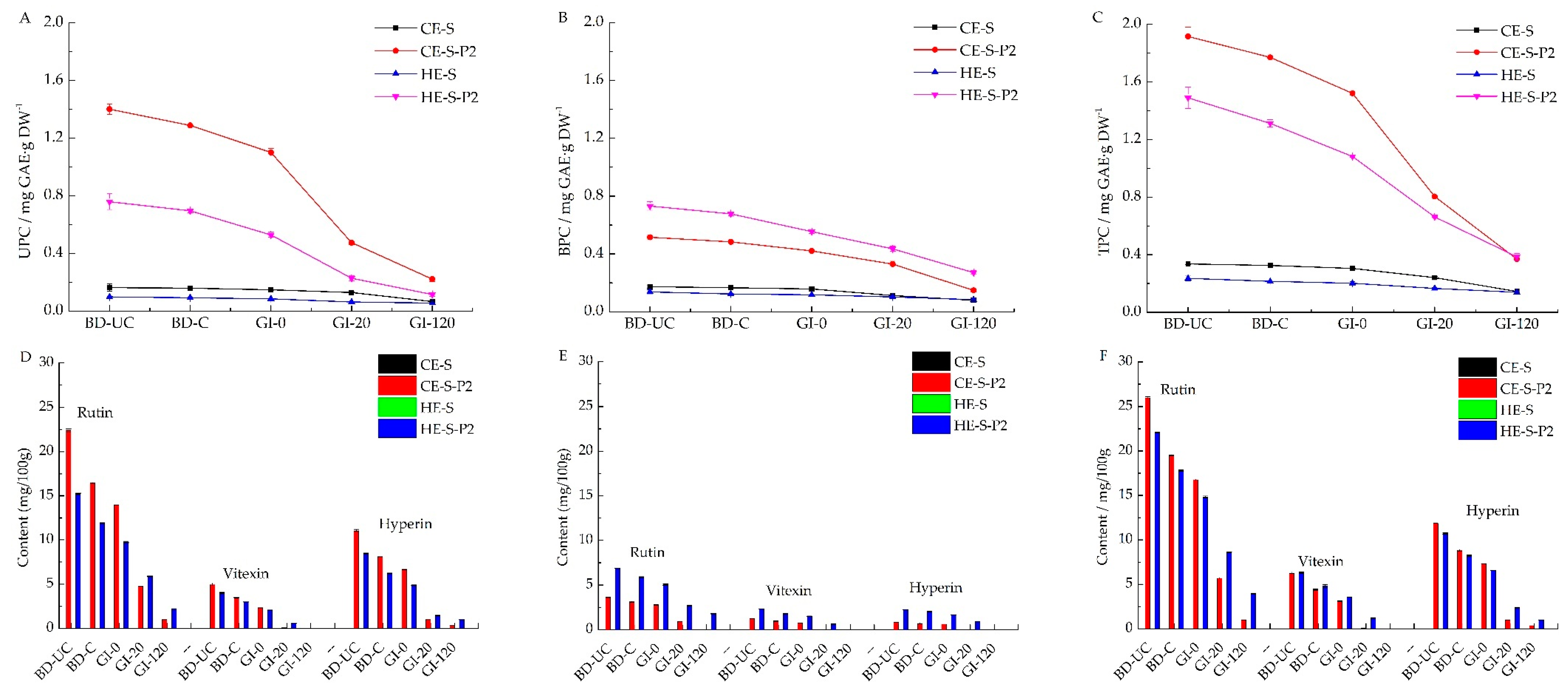
3.5. Phenolic Contents of Extruded Noodles and Residues during Digestion
3.6. Phenolic Profiles of Extruded Noodles and Residues during Digestion
3.7. Bioaccessibility Index of Main Phenolic Profiles
3.8. Antioxidant Activity of Extruded Noodles and Residues during Digestion
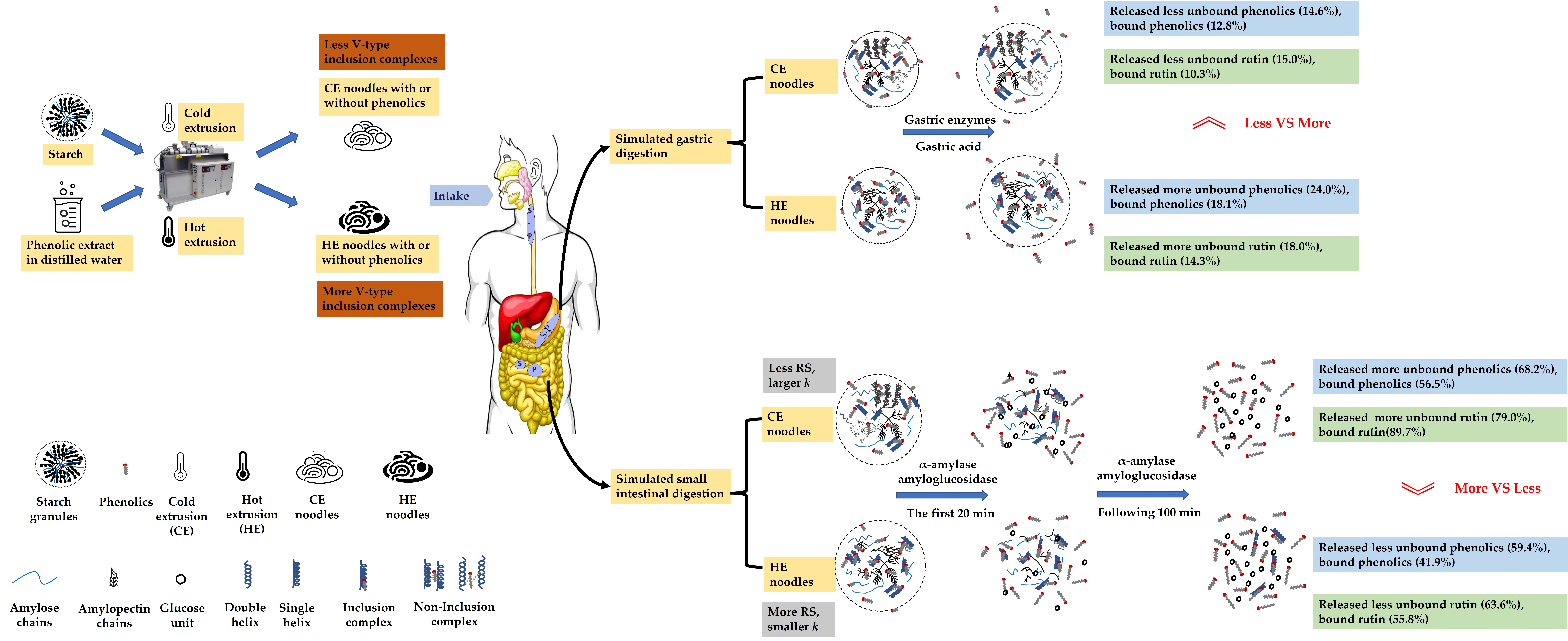
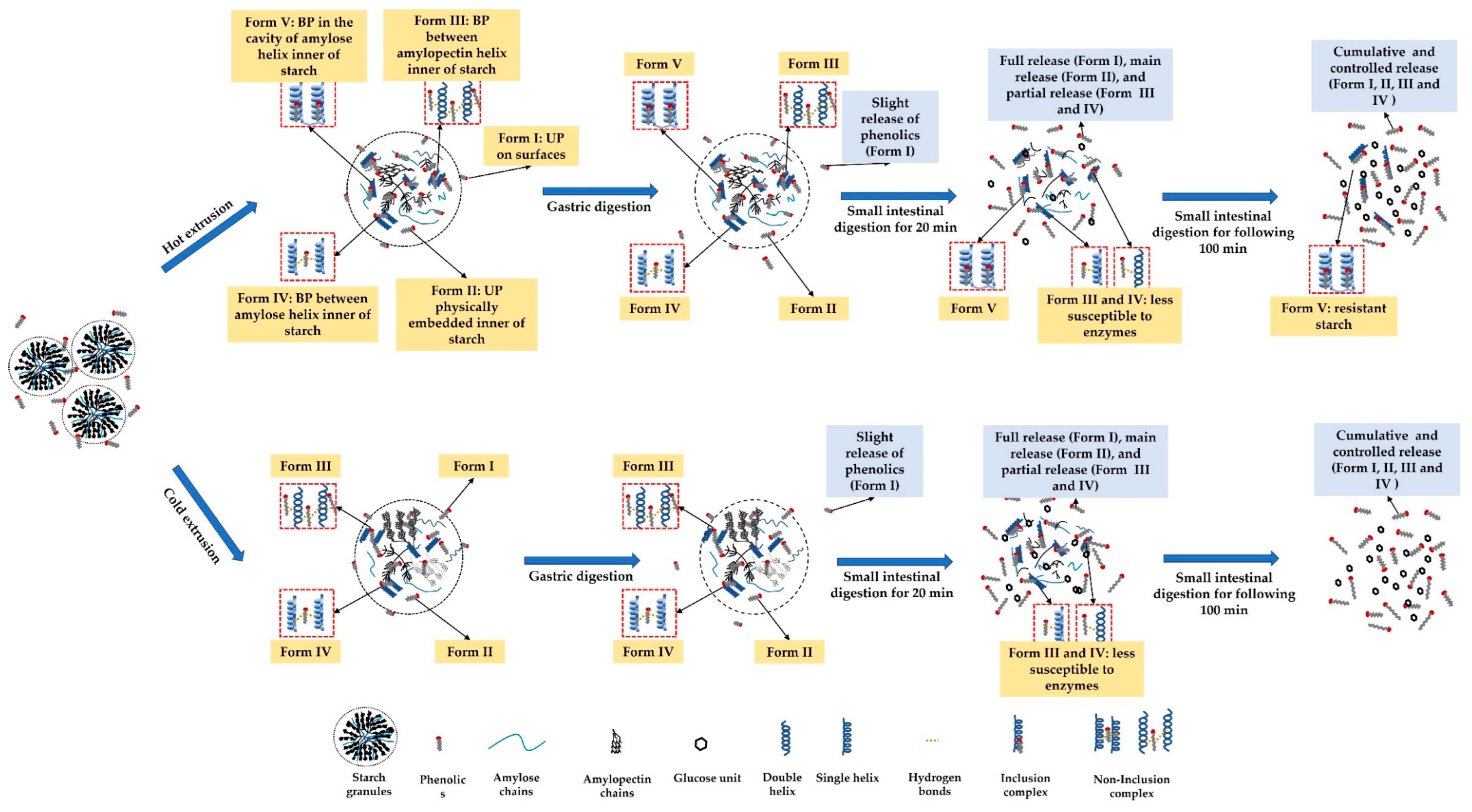
3.9. Possible Mechanism of Unbound and Bound Phenolic Release
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arfaoui, L. Dietary Plant Polyphenols: Effects of Food Processing on Their Content and Bioavailability. Molecules 2021, 26, 2959. [Google Scholar] [CrossRef] [PubMed]
- Le, N.L. Functional compounds in dragon fruit peels and their potential health benefits: A review. Int. J. Food Sci. Technol. 2021, 57, 2571–2580. [Google Scholar] [CrossRef]
- Mu, J.; Wu, G.; Chen, Z.; Klosek, A.; Brennan, M.A.; Hui, X.; Brennan, C.S. The effects of blackcurrant and strawberry powder on the physicochemical and in vitro glycaemic response of starches derived from sweet potato (Ipomoea batatas) and potato (Solanum tuberosum). Int. J. Food Sci. Technol. 2021, 56, 6062–6072. [Google Scholar] [CrossRef]
- Feng, S.; Yi, J.; Li, X.; Wu, X.; Zhao, Y.; Ma, Y.; Bi, J. Systematic Review of Phenolic Compounds in Apple Fruits: Compositions, Distribution, Absorption, Metabolism, and Processing Stability. J. Agric. Food Chem. 2021, 69, 7–27. [Google Scholar] [CrossRef]
- Chi, C.; Li, X.; Zhang, Y.; Chen, L.; Xie, F.; Li, L.; Bai, G. Modulating the in vitro digestibility and predicted glycemic index of rice starch gels by complexation with gallic acid. Food Hydrocoll. 2019, 89, 821–828. [Google Scholar] [CrossRef]
- Cohen, R.; Schwartz, B.; Peri, I.; Shimoni, E. Improving bioavailability and stability of genistein by complexation with high-amylose corn starch. J. Agric. Food Chem. 2011, 59, 7932–7938. [Google Scholar] [CrossRef]
- Zhu, S.; Liu, B.; Wang, F.; Huang, D.; Zhong, F.; Li, Y. Characterization and in vitro digestion properties of cassava starch and epigallocatechin-3-gallate (EGCG) blend. LWT 2021, 137, 110398. [Google Scholar] [CrossRef]
- Gutierrez, A.S.A.; Guo, J.; Feng, J.; Tan, L.; Kong, L. Inhibition of starch digestion by gallic acid and alkyl gallates. Food Hydrocoll. 2020, 102, 105603. [Google Scholar] [CrossRef]
- Shi, L.; Hopfer, H.; Ziegler, G.R.; Kong, L. Starch-menthol inclusion complex: Structure and release kinetics. Food Hydrocoll. 2019, 97, 105183. [Google Scholar] [CrossRef]
- Ferruzzi, M.G.; Hamaker, B.R.; Bordenave, N. Phenolic compounds are less degraded in presence of starch than in presence of proteins through processing in model porridges. Food Chem. 2020, 309, 125769. [Google Scholar] [CrossRef]
- Wang, R.; Li, M.; Wu, G.; Hui, X.; Tu, J.; Brennan, M.A.; Guo, B.; Brennan, C.S. Inhibition of phenolics on the in vitro digestion of noodles from the view of phenolics release. Int. J. Food Sci. Technol. 2022, 57, 1208–1217. [Google Scholar] [CrossRef]
- Li, M.; Hasjim, J.; Xie, F.; Halley, P.J.; Gilbert, R.G. Shear degradation of molecular, crystalline, and granular structures of starch during extrusion. Starch-Stärke 2014, 66, 595–605. [Google Scholar] [CrossRef]
- Wang, R.; Li, M.; Chen, S.; Hui, Y.; Tang, A.; Wei, Y. Effects of flour dynamic viscosity on the quality properties of buckwheat noodles. Carbohydr. Polym. 2019, 207, 815–823. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Yu, C.; Fu, M.; Wu, D.; Gao, C.; Feng, X.; Cheng, W.; Shen, X.; Tang, X. Extruded whole buckwheat noodles: Effects of processing variables on the degree of starch gelatinization, changes of nutritional components, cooking characteristics and in vitro starch digestibility. Food Funct. 2019, 10, 6362–6373. [Google Scholar] [CrossRef]
- Takahama, U.; Hirota, S. Interactions of flavonoids with alpha-amylase and starch slowing down its digestion. Food Funct. 2018, 9, 677–687. [Google Scholar] [CrossRef]
- Amoako, D.B.; Awika, J.M. Polyphenol interaction with food carbohydrates and consequences on availability of dietary glucose. Curr. Opin. Food Sci. 2016, 8, 14–18. [Google Scholar] [CrossRef]
- Sun, L.; Miao, M. Dietary polyphenols modulate starch digestion and glycaemic level: A review. Crit. Rev. Food Sci. Nutr. 2020, 60, 541–555. [Google Scholar] [CrossRef]
- Sun, L.; Wang, Y.; Miao, M. Inhibition of α-amylase by polyphenolic compounds: Substrate digestion, binding interactions and nutritional intervention. Trends Food Sci. Technol. 2020, 104, 190–207. [Google Scholar] [CrossRef]
- Giuberti, G.; Rocchetti, G.; Lucini, L. Interactions between phenolic compounds, amylolytic enzymes and starch: An updated overview. Curr. Opin. Food Sci. 2020, 31, 102–113. [Google Scholar] [CrossRef]
- Xu, T.; Li, X.; Ji, S.; Zhong, Y.; Simal-Gandara, J.; Capanoglu, E.; Xiao, J.; Lu, B. Starch modification with phenolics: Methods, physicochemical property alteration, and mechanisms of glycaemic control. Trends Food Sci. Technol. 2021, 111, 12–26. [Google Scholar] [CrossRef]
- Cho, M.; Choi, W.-S.; You, S. Steady and Dynamic Shear Rheology of Fucoidan-Buckwheat Starch Mixtures. Starch-Stärke 2009, 61, 282–290. [Google Scholar] [CrossRef]
- AACC. Crude Protein—Improved Kjeldahl Method. In AACC International Approved Methods; AACC: St. Paul, MN, USA, 2002; Volume 46-10. [Google Scholar]
- AACC. Crude Fat in Flour, Bread, and Baked Cereal Products Not Containing Fruit. In AACC International Approved Methods; AACC: St. Paul, MN, USA, 2002; Volume 30-10. [Google Scholar]
- AACC. Ash—Basic Method. In AACC International Approved Methods; AACC: St. Paul, MN, USA, 2002; Volume 08-01. [Google Scholar]
- Zhang, W.; Zhu, Y.; Liu, Q.; Bao, J.; Liu, Q. Identification and quantification of polyphenols in hull, bran and endosperm of common buckwheat (Fagopyrum esculentum) seeds. J. Funct. Foods 2017, 38, 363–369. [Google Scholar] [CrossRef]
- Machado, A.P.D.F.; Sumere, B.R.; Mekaru, C.; Martinez, J.; Bezerra, R.M.N.; Rostagno, M.A. Extraction of polyphenols and antioxidants from pomegranate peel using ultrasound: Influence of temperature, frequency and operation mode. Int. J. Food Sci. Technol. 2019, 54, 2792–2801. [Google Scholar] [CrossRef]
- Wen, C.; Zhang, J.; Zhang, H.; Dzah, C.S.; Zandile, M.; Duan, Y.; Ma, H.; Luo, X. Advances in ultrasound assisted extraction of bioactive compounds from cash crops—A review. Ultrason. Sonochem. 2018, 48, 538–549. [Google Scholar] [CrossRef]
- Liu, F.; He, C.; Wang, L.; Wang, M. Effect of milling method on the chemical composition and antioxidant capacity of Tartary buckwheat flour. Int. J. Food Sci. Technol. 2018, 53, 2457–2464. [Google Scholar] [CrossRef]
- Aguirre, J.F.; Osella, C.A.; Carrara, C.R.; Sánchez, H.D.; Buera, M.d.P. Effect of storage temperature on starch retrogradation of bread staling. Starch-Stärke 2011, 63, 587–593. [Google Scholar] [CrossRef]
- Zeng, X.; Zheng, B.; Xiao, G.; Chen, L. Synergistic effect of extrusion and polyphenol molecular interaction on the short/long-term retrogradation properties of chestnut starch. Carbohydr. Polym. 2022, 276, 118731. [Google Scholar] [CrossRef]
- Tian, Y.; Li, M.; Liu, X.; Jane, J.L.; Guo, B.; Dhital, S. Storage temperature and time affect the enzyme resistance starch and glycemic response of cooked noodles. Food Chem. 2021, 344, 128702. [Google Scholar] [CrossRef]
- Gao, S.; Liu, H.; Sun, L.; Cao, J.; Yang, J.; Lu, M.; Wang, M. Rheological, thermal and in vitro digestibility properties on complex of plasma modified Tartary buckwheat starches with quercetin. Food Hydrocoll. 2021, 110, 106209. [Google Scholar] [CrossRef]
- Goni, I.; GarciaAlonso, A.; SauraCalixto, F. A starch hydrolysis procedure to estimate glycemic index. Nutr. Res. 1997, 17, 427–437. [Google Scholar] [CrossRef]
- Singleton, V.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1964, 16, 144–158. [Google Scholar]
- Park, B.I.; Kim, J.; Lee, K.; Lim, T.; Hwang, K.T. Flavonoids in common and tartary buckwheat hull extracts and antioxidant activity of the extracts against lipids in mayonnaise. J. Food Sci. Technol. 2019, 56, 2712–2720. [Google Scholar] [CrossRef] [PubMed]
- Chait, Y.A.; Gunenc, A.; Bendali, F.; Hosseinian, F. Simulated gastrointestinal digestion and in vitro colonic fermentation of carob polyphenols: Bioaccessibility and bioactivity. LWT 2020, 117, 108623. [Google Scholar] [CrossRef]
- Wang, L.; Wang, L.; Li, Z.; Gao, Y.; Cui, S.W.; Wang, T.; Qiu, J. Diverse effects of rutin and quercetin on the pasting, rheological and structural properties of Tartary buckwheat starch. Food Chem. 2021, 335, 127556. [Google Scholar] [CrossRef]
- Liu, B.; Zhong, F.; Yokoyama, W.; Huang, D.; Zhu, S.; Li, Y. Interactions in starch co-gelatinized with phenolic compound systems: Effect of complexity of phenolic compounds and amylose content of starch. Carbohydr. Polym. 2020, 247, 116667. [Google Scholar] [CrossRef]
- Amoako, D.B.; Awika, J.M. Polymeric tannins significantly alter properties and in vitro digestibility of partially gelatinized intact starch granule. Food Chem. 2016, 208, 10–17. [Google Scholar] [CrossRef]
- Chou, S.; Li, B.; Tan, H.; Zhang, W.; Zang, Z.; Cui, H.; Wang, H.; Zhang, S.; Meng, X. The effect of pH on the chemical and structural interactions between apple polyphenol and starch derived from rice and maize. Food Sci. Nutr. 2020, 8, 5026–5035. [Google Scholar] [CrossRef]
- Amoako, D.B.; Awika, J.M. Resistant starch formation through intrahelical V-complexes between polymeric proanthocyanidins and amylose. Food Chem. 2019, 285, 326–333. [Google Scholar] [CrossRef]
- Zhao, B.; Sun, S.; Lin, H.; Chen, L.; Qin, S.; Wu, W.; Zheng, B.; Guo, Z. Physicochemical properties and digestion of the lotus seed starch-green tea polyphenol complex under ultrasound-microwave synergistic interaction. Ultrason. Sonochem. 2019, 52, 50–61. [Google Scholar] [CrossRef]
- Yan, X.; Wu, Z.Z.; Li, M.Y.; Yin, F.; Ren, K.X.; Tao, H. The combined effects of extrusion and heat-moisture treatment on the physicochemical properties and digestibility of corn starch. Int. J. Biol. Macromol. 2019, 134, 1108–1112. [Google Scholar] [CrossRef]
- Tao, H.; Zhu, X.F.; Nan, B.X.; Jiang, R.Z.; Wang, H.L. Effect of extruded starches on the structure, farinograph characteristics and baking behavior of wheat dough. Food Chem. 2021, 348, 129017. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Singh, N.; Singh, B. Effect of extrusion on morphology, structural, functional properties and in vitro digestibility of corn, field pea and kidney bean starches. Starch-Stärke 2015, 67, 721–728. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, Y.; Zhang, R.; Zhong, Y.; Luo, Y.; Xu, S.; Liu, J.; Xue, J.; Guo, D. Effects of extrusion treatment on physicochemical properties and in vitro digestion of pregelatinized high amylose maize flour. J. Cereal Sci. 2016, 68, 108–115. [Google Scholar] [CrossRef]
- Chang, R.; Jin, Z.; Lu, H.; Qiu, L.; Sun, C.; Tian, Y. Type III Resistant Starch Prepared from Debranched Starch: Structural Changes under Simulated Saliva, Gastric, and Intestinal Conditions and the Impact on Short-Chain Fatty Acid Production. J. Agric. Food Chem. 2021, 69, 2595–2602. [Google Scholar] [CrossRef] [PubMed]
- Xing, J.-j.; Li, D.; Wang, L.-J.; Adhikari, B. Temperature thresholds and time-temperature dependence of gelatinization for heat-moisture treated corn starch. J. Food Eng. 2018, 217, 43–49. [Google Scholar] [CrossRef]
- Jiang, X.; Wang, J.; Ou, Y.; Zheng, B. Effect of chlorogenic acid on the structural properties and digestibility of lotus seed starch during microwave gelatinization. Int. J. Biol. Macromol. 2021, 191, 474–482. [Google Scholar] [CrossRef]
- Liu, R.; Xu, C.; Cong, X.; Wu, T.; Song, Y.; Zhang, M. Effects of oligomeric procyanidins on the retrogradation properties of maize starch with different amylose/amylopectin ratios. Food Chem. 2017, 221, 2010–2017. [Google Scholar] [CrossRef]
- Wang, M.; Shen, Q.; Hu, L.; Hu, Y.; Ye, X.; Liu, D.; Chen, J. Physicochemical properties, structure and in vitro digestibility on complex of starch with lotus (Nelumbo nucifera Gaertn.) leaf flavonoids. Food Hydrocoll. 2018, 81, 191–199. [Google Scholar] [CrossRef]
- Kale, S.J.; Jha, S.K.; Jha, G.K.; Sinha, J.P.; Lal, S.B. Soaking Induced Changes in Chemical Composition, Glycemic Index and Starch Characteristics of Basmati Rice. Rice Sci. 2015, 22, 227–236. [Google Scholar] [CrossRef]
- Henderson, A.M.; Rudin, A. Effects of water on starch-g-polystyrene and starch-g-poly(methyl acrylate) extrudates. J. Appl. Polym. Sci. 1982, 27, 4115–4135. [Google Scholar] [CrossRef]
- Wang, R.; Li, M.; Wei, Y.; Guo, B.; Brennan, M.; Brennan, C.S. Quality Differences between Fresh and Dried Buckwheat Noodles Associated with Water Status and Inner Structure. Foods 2021, 10, 187. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.; Oztop, M.H.; Singh, R.P.; McCarthy, M.J. Physical changes in white and brown rice during simulated gastric digestion. J. Food Sci. 2011, 76, E450–E457. [Google Scholar] [CrossRef] [PubMed]
- Bordoloi, A.; Singh, J.; Kaur, L. In vitro digestibility of starch in cooked potatoes as affected by guar gum: Microstructural and rheological characteristics. Food Chem. 2012, 133, 1206–1213. [Google Scholar] [CrossRef]
- Nadia, J.; Bronlund, J.; Singh, R.P.; Singh, H.; Bornhorst, G.M. Structural breakdown of starch-based foods during gastric digestion and its link to glycemic response: In vivo and in vitro considerations. Compr. Rev. Food Sci. Food Saf. 2021, 20, 2660–2698. [Google Scholar] [CrossRef]
- Rocchetti, G.; Giuberti, G.; Gallo, A.; Bernardi, J.; Marocco, A.; Lucini, L. Effect of dietary polyphenols on the in vitro starch digestibility of pigmented maize varieties under cooking conditions. Food Res. Int. 2018, 108, 183–191. [Google Scholar] [CrossRef]
- Tuersuntuoheti, T.; Wang, Z.; Duan, M.; Asimi, S.; Ren, X.; Wang, Z.; Zheng, Y.; Wu, Y.; Liang, S.; Zhang, M. Noodle processing, storage time and cooking affect the antioxidant activities and phenolic compounds content of Qingke barley noodles. Int. J. Food Sci. Technol. 2020, 55, 2730–2739. [Google Scholar] [CrossRef]
- Adarkwah-Yiadom, M.; Duodu, K.G. Effect of extrusion cooking and simulated in vitro gastrointestinal digestion on condensed tannins and radical scavenging activity of type II and type III whole grain sorghum. Int. J. Food Sci. Technol. 2017, 52, 2282–2294. [Google Scholar] [CrossRef]
- Rocchetti, G.; Giuberti, G.; Busconi, M.; Marocco, A.; Trevisan, M.; Lucini, L. Pigmented sorghum polyphenols as potential inhibitors of starch digestibility: An in vitro study combining starch digestion and untargeted metabolomics. Food Chem. 2020, 312, 126077. [Google Scholar] [CrossRef]
- Hernández, H.A.R.; Gutiérrez, T.J.; Bello-Pérez, L.A. Can starch-polyphenol V-type complexes be considered as resistant starch? Food Hydrocoll. 2022, 124, 107226. [Google Scholar] [CrossRef]
- Yu, M.; Zhu, S.; Zhong, F.; Zhang, S.; Du, C.; Li, Y. Insight into the multi-scale structure changes and mechanism of corn starch modulated by different structural phenolic acids during retrogradation. Food Hydrocoll. 2022, 128, 107581. [Google Scholar] [CrossRef]
- Beta, T.; Corke, H. Effect of Ferulic Acid and Catechin on Sorghum and Maize Starch Pasting Properties. Cereal Chem. J. 2004, 81, 418–422. [Google Scholar] [CrossRef]
- Asensio, J.L.; Ardá, A.; Cañada, F.J.; Jiménez-Barbero, J. Carbohydrate–Aromatic Interactions. Acc. Chem. Res. 2013, 46, 946–954. [Google Scholar] [CrossRef]
- Li, L.; Lietz, G.; Seal, C.J. Phenolic, apparent antioxidant and nutritional composition of quinoa (Chenopodium quinoa Willd.) seeds. Int. J. Food Sci. Technol. 2021, 56, 3245–3254. [Google Scholar] [CrossRef]
- Liu, W.-C.; Halley, P.J.; Gilbert, R.G. Mechanism of Degradation of Starch, a Highly Branched Polymer, during Extrusion. Macromolecules 2010, 43, 2855–2864. [Google Scholar] [CrossRef]
- El-Khalek, E.A.; Janssens, G.P.J. Effect of extrusion processing on starch gelatinisation and performance in poultry. Worlds Poult. Sci. J. 2019, 66, 53–64. [Google Scholar] [CrossRef]
- Nadia, J.; Bronlund, J.E.; Singh, H.; Singh, R.P.; Bornhorst, G.M. Contribution of the proximal and distal gastric phases to the breakdown of cooked starch-rich solid foods during static in vitro gastric digestion. Food Res. Int. 2022, 157, 111270. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | RDS (%) | SDS (%) | RS (%) | k (min−1) | C∞ (%) |
|---|---|---|---|---|---|
| UE-S | 58.77 ± 2.10 a | 20.95 ± 0.53 e | 20.28 ± 1.58 e | 0.0401 ± 0.0015 a | 98.34 ± 0.47 a |
| CE-S | 29.74 ± 0.78 c | 43.24 ± 1.71 a | 27.02 ± 0.94 d | 0.0272 ± 0.0011 b | 91.80 ± 1.54 b |
| CE-S-P2 | 24.12 ± 0.88 e | 39.60 ± 0.49 b | 36.28 ± 1.37 b | 0.0222 ± 0.0005 d | 82.38 ± 1.43 c |
| HE-S | 34.75 ± 0.85 b | 37.08 ± 0.69 c | 28.17 ± 0.15 c | 0.0237 ± 0.0006 c | 90.28 ± 0.55 b |
| HE-S-P2 | 26.62 ± 1.24 d | 33.22 ± 0.99 d | 40.15 ± 0.25 a | 0.0175 ± 0.0003 e | 81.09 ± 0.25 c |
| Noodles | Profiles | C0 (mg/100 g) | Cg (mg/100 g) | Ci (mg/100 g) | BIg (%) | BIi (%) |
|---|---|---|---|---|---|---|
| CE-S | Rutin, vitexin, hyperin | n.d. | n.d. | n.d. | n.d. | n.d. |
| CE-S-P2 | Rutin | 19.55 ± 0.10 a | 1.77 ± 0.01 a | 13.00 ± 0.06 b | 9.05 | 66.50 |
| vitexin | 4.39 ± 0.08 f | 0.71 ± 0.01 e | 1.42 ± 0.01 c | 16.17 | 32.35 | |
| hyperin | 8.83 ± 0.11 c | 1.21 ± 0.00 c | 2.87 ± 0.01 e | 13.70 | 32.50 | |
| HE-S | Rutin, vitexin, hyperin | n.d. | n.d. | n.d. | n.d. | n.d. |
| HE-S-P2 | Rutin | 17.81 ± 0.10 b | 1.39 ± 0.02 b | 11.72 ± 0.07 a | 7.80 | 65.81 |
| vitexin | 4.79 ± 0.15 e | 0.52 ± 0.02 f | 1.32 ± 0.04 d | 10.86 | 27.56 | |
| hyperin | 8.21 ± 0.10 d | 1.07 ± 0.01 d | 2.86 ± 0.01 e | 13.03 | 34.84 |
| Noodles | Unbound Phenolics | Bound Phenolics | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| BD-UC | BD-C | GI-0 | GI-20 | GI-120 | BD-UC | BD-C | GI-0 | GI-20 | GI-120 | |
| CE-S | 1.588 ± 0.078 aC | 1.535 ± 0.238 abC | 1.312 ± 0.022 bC | 1.171 ± 0.046 cC | 0.356 ± 0.016 dC | 1.625 ± 0.107 aC | 1.571 ± 0.053 aC | 1.489 ± 0.033 aC | 0.922 ± 0.035 bC | 0.522 ± 0.023 cD |
| CE-S-P2 | 19.088 ± 0.822 aA | 16.609 ± 0.850 bA | 13.708 ± 1.666 cA | 5.474 ± 0.098 dA | 2.884 ± 0.126 eA | 6.665 ± 0.209 aB | 5.775 ± 0.138 bB | 5.035 ± 0.224 cB | 3.686 ± 0.362 dB | 1.335 ± 0.021 eB |
| HE-S | 0.959 ± 0.036 aD | 0.912 ± 0.013 aD | 0.759 ± 0.032 bD | 0.416 ± 0.030 cD | 0.225 ± 0.030 dD | 1.235 ± 0.210 aD | 1.166 ± 0.065 aD | 1.002 ± 0.032 bD | 0.909 ± 0.012 cC | 0.669 ± 0.044 dC |
| HE-S-P2 | 9.581 ± 0.037 aB | 7.882 ± 0.543 bB | 7.413 ± 0.779 cB | 2.405 ± 0.254 dB | 1.070 ± 0.046 eB | 8.222 ± 0.613 aA | 7.452 ± 0.448 bA | 6.515 ± 0.201 cA | 4.673 ± 0.387 dA | 3.002 ± 0.552 eA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, R.; Li, M.; Brennan, M.A.; Kulasiri, D.; Guo, B.; Brennan, C.S. Phenolic Release during In Vitro Digestion of Cold and Hot Extruded Noodles Supplemented with Starch and Phenolic Extracts. Nutrients 2022, 14, 3864. https://doi.org/10.3390/nu14183864
Wang R, Li M, Brennan MA, Kulasiri D, Guo B, Brennan CS. Phenolic Release during In Vitro Digestion of Cold and Hot Extruded Noodles Supplemented with Starch and Phenolic Extracts. Nutrients. 2022; 14(18):3864. https://doi.org/10.3390/nu14183864
Chicago/Turabian StyleWang, Ruibin, Ming Li, Margaret Anne Brennan, Don Kulasiri, Boli Guo, and Charles Stephen Brennan. 2022. "Phenolic Release during In Vitro Digestion of Cold and Hot Extruded Noodles Supplemented with Starch and Phenolic Extracts" Nutrients 14, no. 18: 3864. https://doi.org/10.3390/nu14183864
APA StyleWang, R., Li, M., Brennan, M. A., Kulasiri, D., Guo, B., & Brennan, C. S. (2022). Phenolic Release during In Vitro Digestion of Cold and Hot Extruded Noodles Supplemented with Starch and Phenolic Extracts. Nutrients, 14(18), 3864. https://doi.org/10.3390/nu14183864