
Association of Gut Microbiota Enterotypes with Blood Trace Elements in Women with Infertility

Abstract
:1. Introduction
2. Materials and Methods
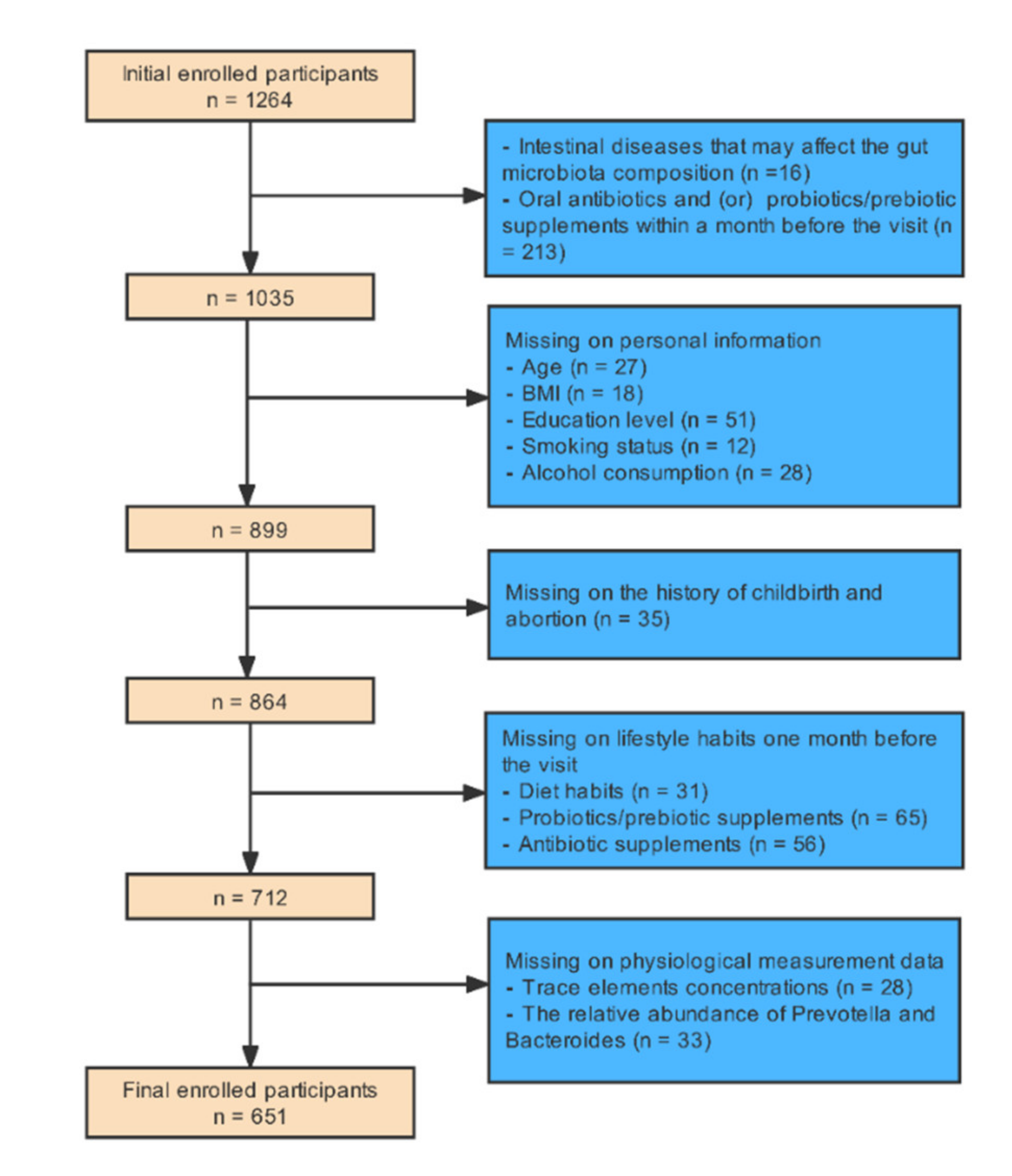
2.1. Study Design and Study Population
2.2. Sample Measurement
2.3. Statistical Analysis
3. Results
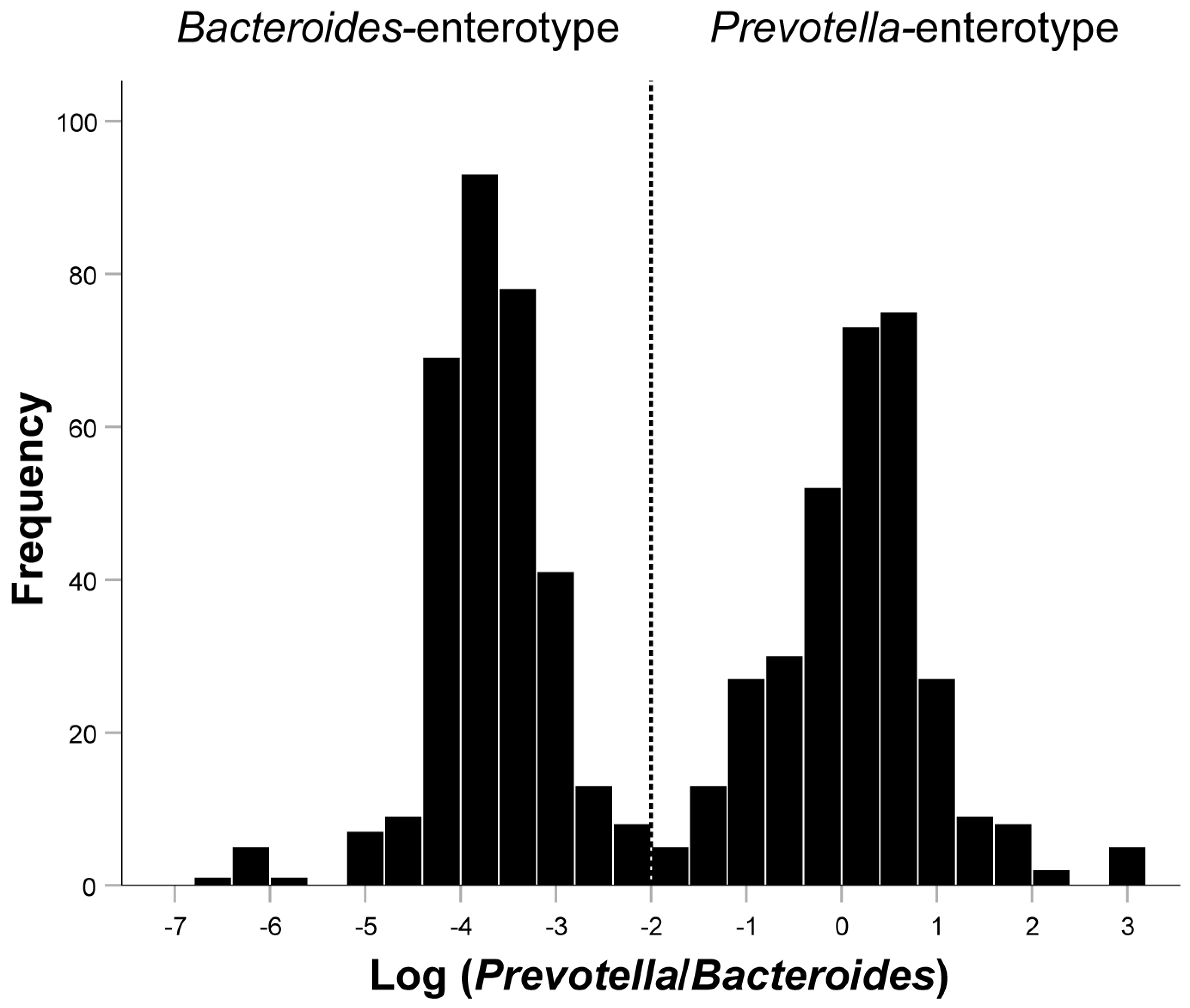
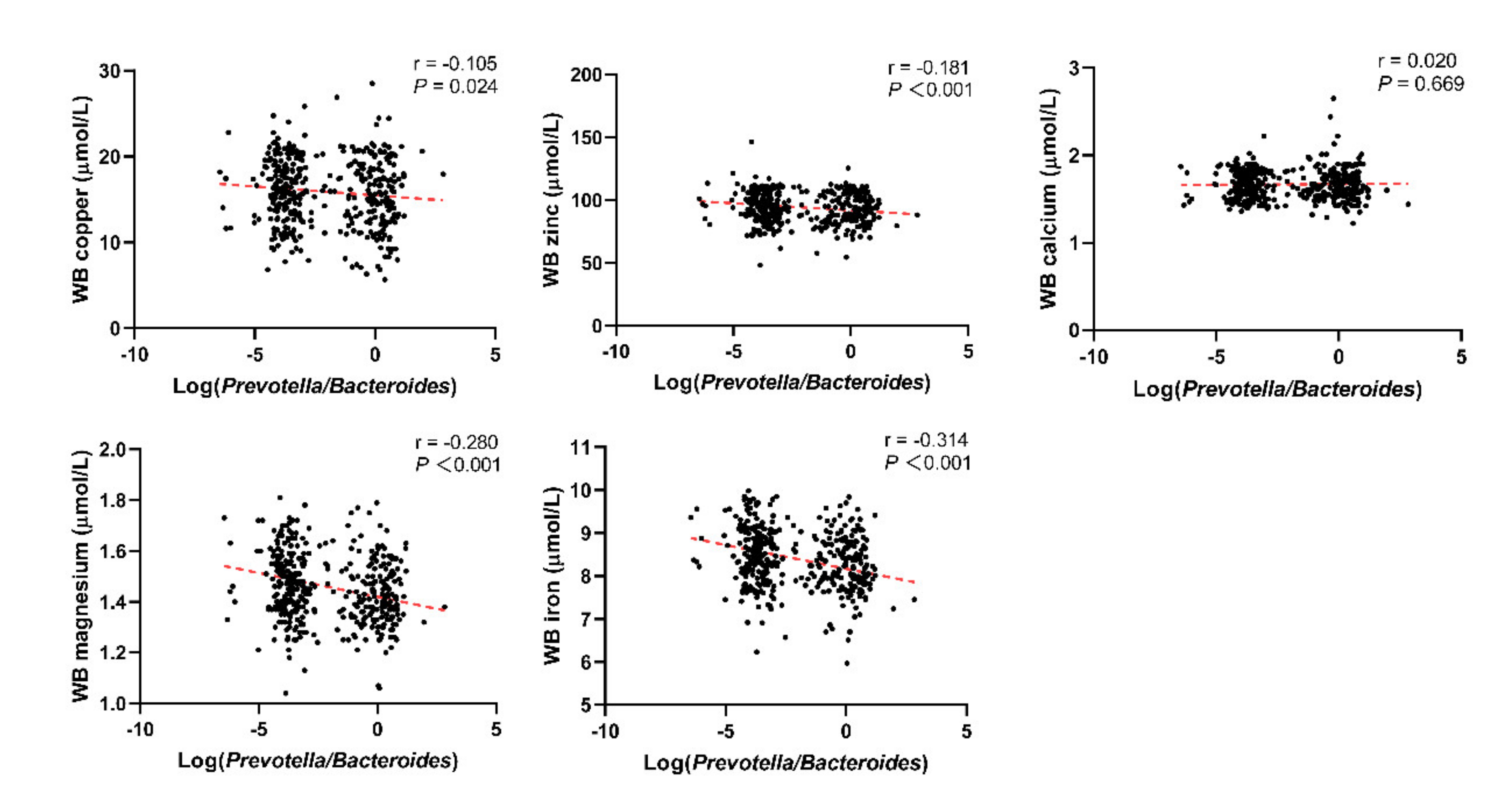
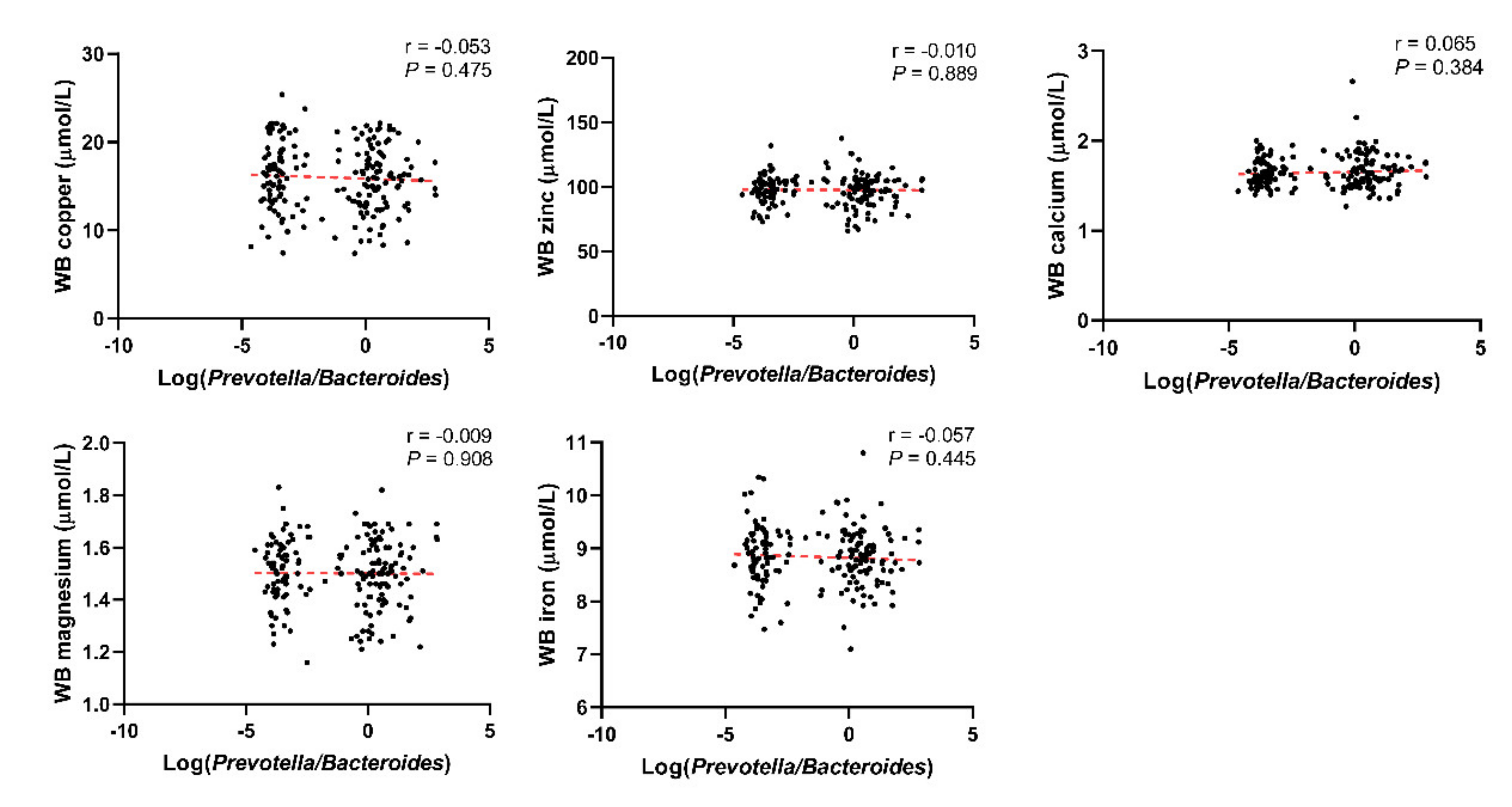
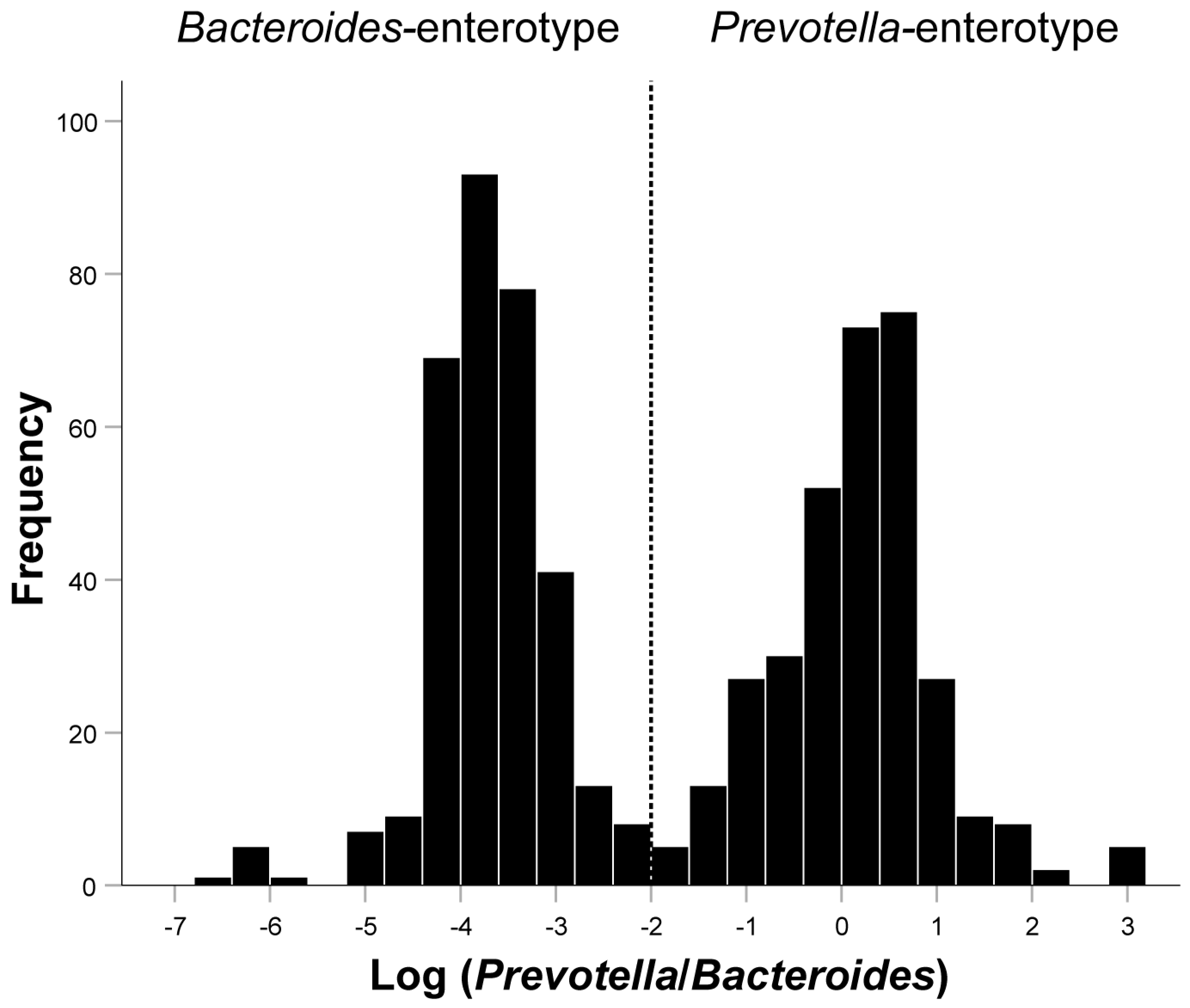
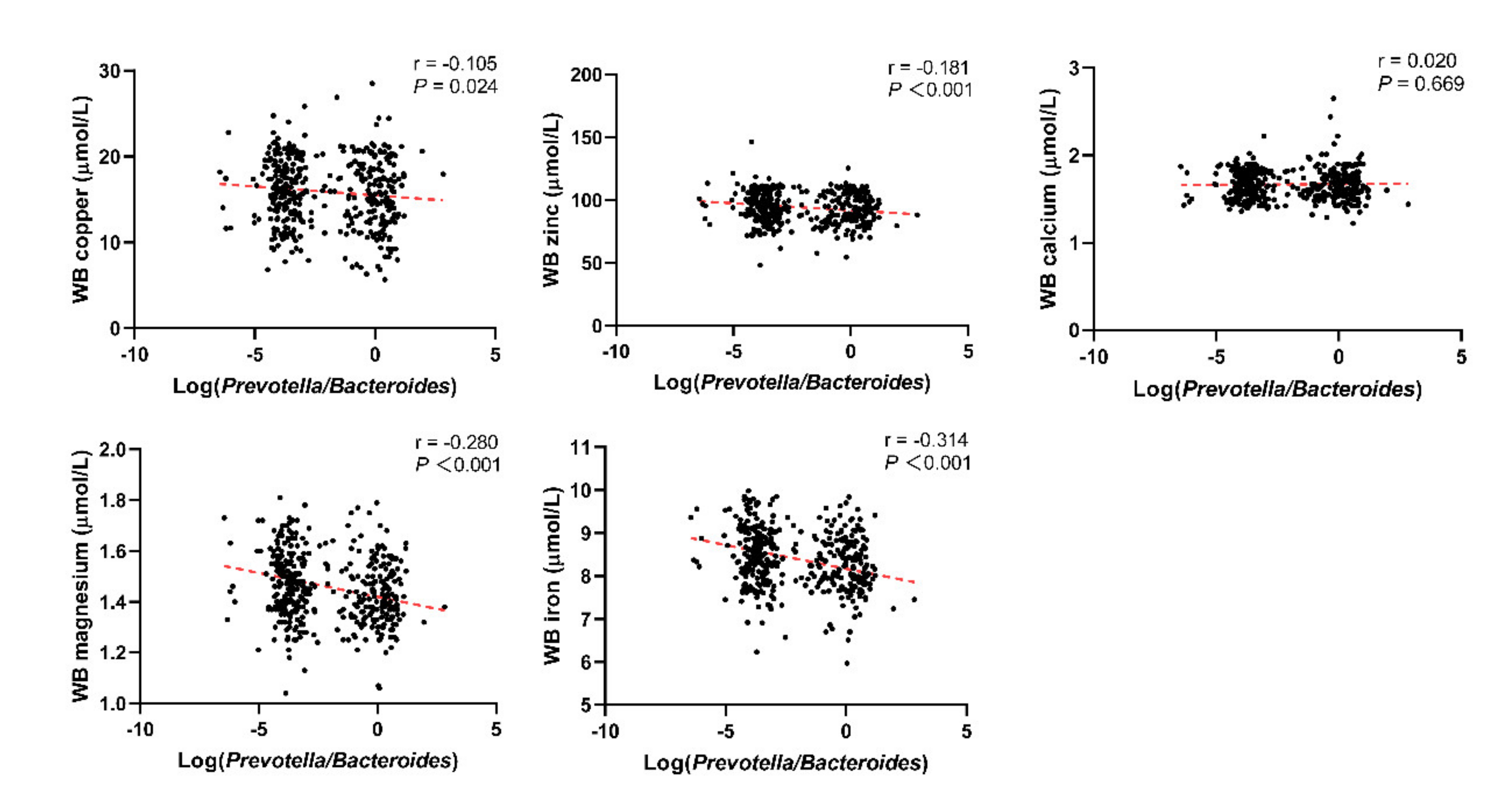
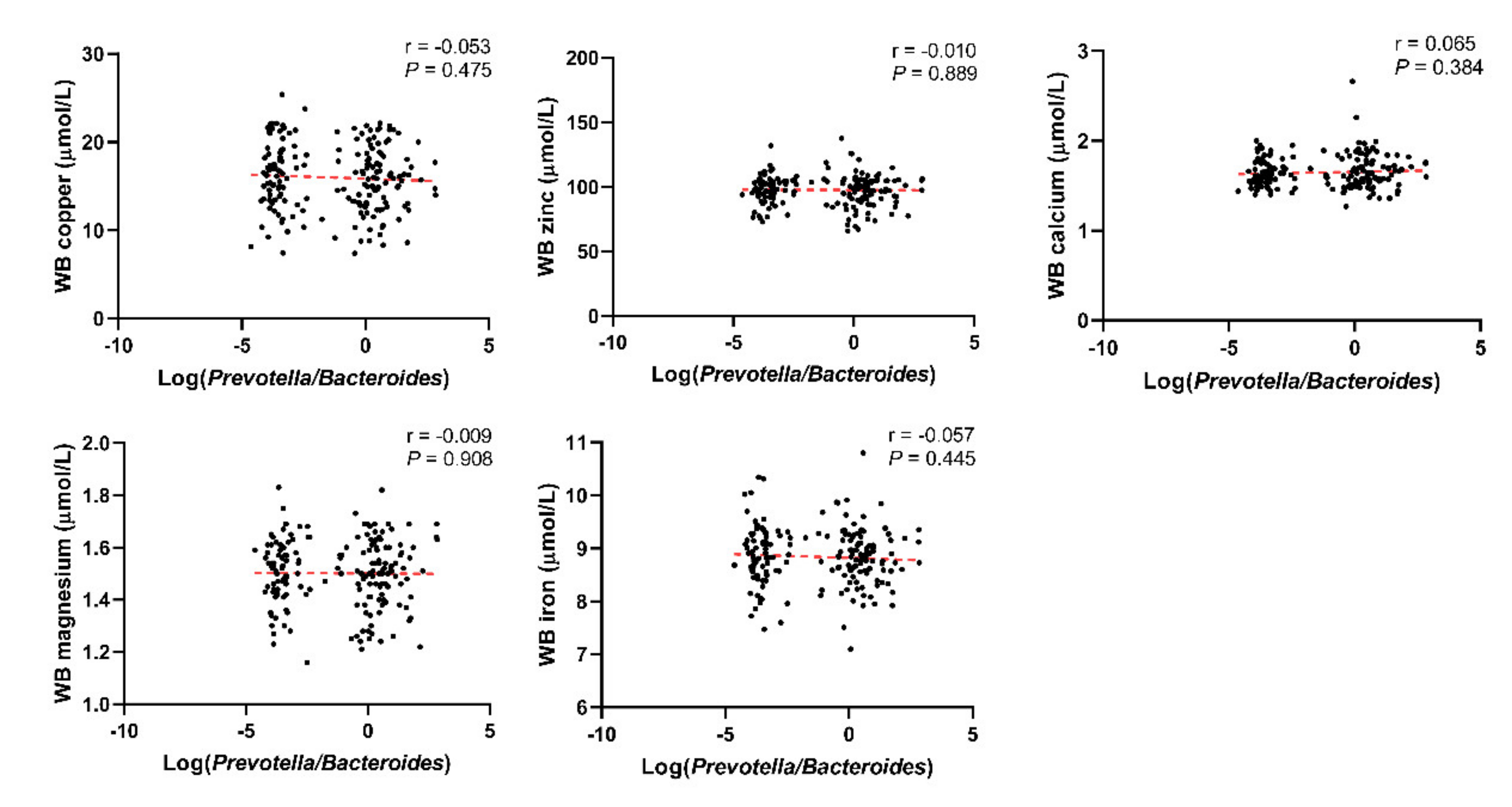
3.1. Basic Characteristics of the Included Population and the Relationship between the Log(P/B), Enterotypes, and Whole Blood Trace Elements
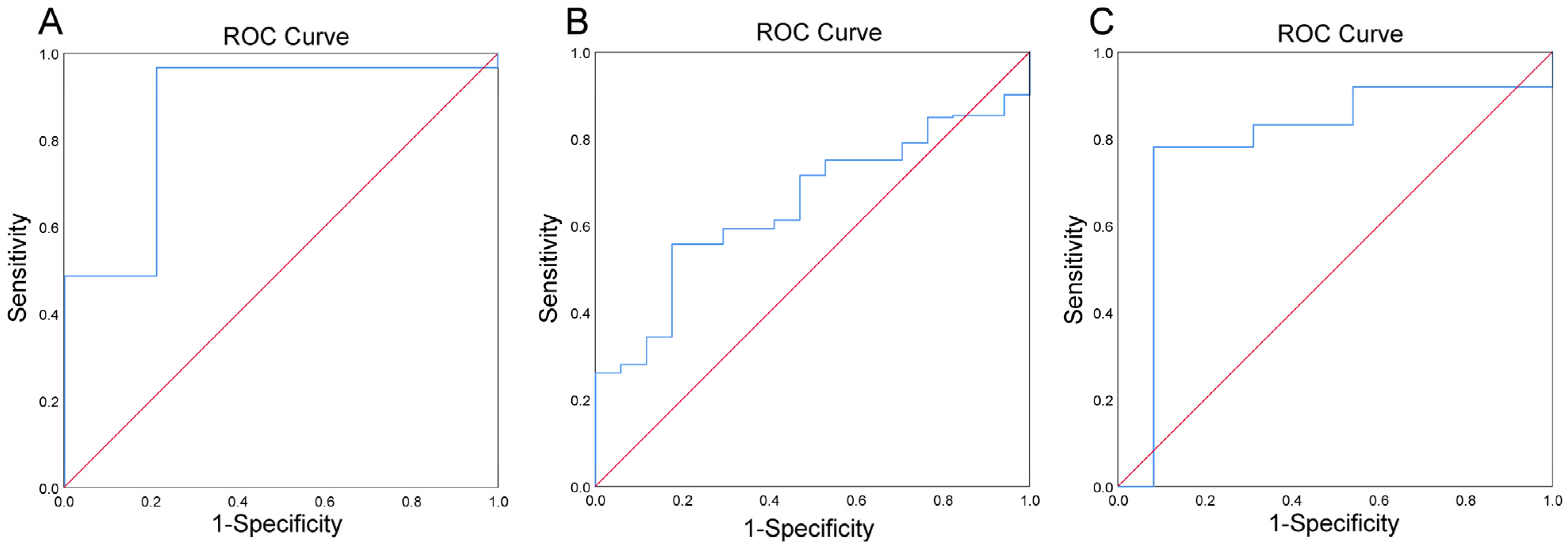
3.2. Prevotella-Enterotype Had an Acceptable Predictive Power to Low Levels of Whole Blood Iron in Obese Population
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Practice Committee of the American Society for Reproductive Medicine. Diagnostic evaluation of the infertile female: A committee opinion. Fertil. Steril. 2015, 103, e44–e50. [Google Scholar] [CrossRef]
- Quigley, E.M. Gut bacteria in health and disease. Gastroenterol. Hepatol. 2013, 9, 560–569. [Google Scholar]
- Gill, S.R.; Pop, M.; DeBoy, R.T.; Eckburg, P.B.; Turnbaugh, P.J.; Samuel, B.S.; Gordon, J.I.; Relman, D.A.; Fraser-Liggett, C.M.; Nelson, K.E. Metagenomic Analysis of the Human Distal Gut Microbiome. Science 2006, 312, 1355–1359. [Google Scholar] [CrossRef] [Green Version]
- Vrieze, A.; Van Nood, E.; Holleman, F.; Salojärvi, J.; Kootte, R.S.; Bartelsman, J.F.; Dallinga-Thie, G.M.; Ackermans, M.T.; Serlie, M.J.; Oozeer, R.; et al. Transfer of Intestinal Microbiota from Lean Donors Increases Insulin Sensitivity in Individuals with Metabolic Syndrome. Gastroenterology 2012, 143, 913–916. [Google Scholar] [CrossRef]
- Qi, X.; Yun, C.; Pang, Y.; Qiao, J. The impact of the gut microbiota on the reproductive and metabolic endocrine system. Gut Microbes 2021, 13, 1894070. [Google Scholar] [CrossRef]
- Kontic-Vucinic, O.; Sulovic, N.; Radunovic, N. Micronutrients in women’s reproductive health: II. Minerals and trace elements. Int. J. Fertil. Women′s Med. 2006, 51, 116–124. [Google Scholar]
- Lewicka, I.; Kocyłowski, R.; Grzesiak, M.; Gaj, Z.; Oszukowski, P.; Suliburska, J. Selected trace elements concentrations in pregnancy and their possible role—Literature review. Ginekol. Polska 2017, 88, 509–514. [Google Scholar] [CrossRef]
- Garner, T.B.; Hester, J.M.; Carothers, A.; Diaz, F.J. Role of zinc in female reproduction. Biol. Reprod. 2021, 104, 976–994. [Google Scholar] [CrossRef]
- Tonai, S.; Kawabata, A.; Nakanishi, T.; Lee, J.Y.; Okamoto, A.; Shimada, M.; Yamashita, Y. Iron deficiency induces female infertile in order to failure of follicular development in mice. J. Reprod. Dev. 2020, 66, 475–483. [Google Scholar] [CrossRef]
- Dring, J.C.; Forma, A.; Chilimoniuk, Z.; Dobosz, M.; Teresiński, G.; Buszewicz, G.; Flieger, J.; Cywka, T.; Januszewski, J.; Baj, J. Essentiality of Trace Elements in Pregnancy, Fertility, and Gynecologic Cancers—A State-of-the-Art Review. Nutrients 2021, 14, 185. [Google Scholar] [CrossRef]
- Keen, C.L.; Uriu-Hare, J.Y.; Hawk, S.N.; A Jankowski, M.; Daston, G.P.; Kwik-Uribe, C.L.; Rucker, R.B. Effect of copper deficiency on prenatal development and pregnancy outcome. Am. J. Clin. Nutr. 1998, 67, 1003S–1011S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takaya, J. Calcium-Deficiency during Pregnancy Affects Insulin Resistance in Offspring. Int. J. Mol. Sci. 2021, 22, 7008. [Google Scholar] [CrossRef]
- Rosner, J.Y.; Gupta, M.; McGill, M.; Xue, X.; Chatterjee, P.; Yoshida-Hay, M.; Robeson, W.; Metz, C. Magnesium deficiency during pregnancy in mice impairs placental size and function. Placenta 2016, 39, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Georgieff, M.K. Iron deficiency in pregnancy. Am. J. Obstet. Gynecol. 2020, 223, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Pajarillo, E.A.B.; Lee, E.; Kang, D.-K. Trace metals and animal health: Interplay of the gut microbiota with iron, manganese, zinc, and copper. Anim. Nutr. 2021, 7, 750–761. [Google Scholar] [CrossRef]
- Yatsunenko, T.; Rey, F.E.; Manary, M.J.; Trehan, I.; Dominguez-Bello, M.G.; Contreras, M.; Magris, M.; Hidalgo, G.; Baldassano, R.N.; Anokhin, A.P.; et al. Human gut microbiome viewed across age and geography. Nature 2012, 486, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.M.; et al. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.-Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roager, H.M.; Licht, T.R.; Poulsen, S.K.; Larsen, T.M.; Bahl, M.I. Microbial Enterotypes, Inferred by the Prevotella-to-Bacteroides Ratio, Remained Stable during a 6-Month Randomized Controlled Diet Intervention with the New Nordic Diet. Appl. Environ. Microbiol. 2014, 80, 1142–1149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hjorth, M.F.; Roager, H.M.; Larsen, T.M.; Poulsen, S.K.; Licht, T.R.; Bahl, M.I.; Zohar, Y.; Astrup, A. Pre-treatment microbial Prevotella-to-Bacteroides ratio, determines body fat loss success during a 6-month randomized controlled diet intervention. Int. J. Obes. 2018, 42, 580–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hjorth, M.F.; Christensen, L.; Kjølbæk, L.; Larsen, L.H.; Roager, H.M.; Kiilerich, P.; Kristiansen, K.; Astrup, A. Pretreatment Prevotella-to-Bacteroides ratio and markers of glucose metabolism as prognostic markers for dietary weight loss maintenance. Eur. J. Clin. Nutr. 2019, 74, 338–347. [Google Scholar] [CrossRef]
- Christensen, L.; Roager, H.M.; Astrup, A.; Hjorth, M. Microbial enterotypes in personalized nutrition and obesity management. Am. J. Clin. Nutr. 2018, 108, 645–651. [Google Scholar] [CrossRef]
- Costea, P.I.; Hildebrand, F.; Arumugam, M.; Bäckhed, F.; Blaser, M.J.; Bushman, F.D.; De Vos, W.M.; Ehrlich, S.D.; Fraser, C.M.; Hattori, M.; et al. Enterotypes in the landscape of gut microbial community composition. Nat. Microbiol. 2017, 3, 8–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieira-Silva, S.; Falony, G.; Darzi, Y.; Lima-Mendez, G.; Yunta, R.G.; Okuda, S.; Vandeputte, D.; Valles-Colomer, M.; Hildebrand, F.; Chaffron, S.; et al. Species–function relationships shape ecological properties of the human gut microbiome. Nat. Microbiol. 2016, 1, 16088. [Google Scholar] [CrossRef] [PubMed]
- Eriksen, A.K.; Brunius, C.; Mazidi, M.; Hellström, P.M.; Risérus, U.; Iversen, K.N.; Fristedt, R.; Sun, L.; Huang, Y.; Nørskov, N.P.; et al. Effects of whole-grain wheat, rye, and lignan supplementation on cardiometabolic risk factors in men with metabolic syndrome: A randomized crossover trial. Am. J. Clin. Nutr. 2020, 111, 864–876. [Google Scholar] [CrossRef] [PubMed]
- Kovatcheva-Datchary, P.; Nilsson, A.; Akrami, R.; Lee, Y.S.; De Vadder, F.; Arora, T.; Hallen, A.; Martens, E.; Björck, I.; Bäckhed, F. Dietary Fiber-Induced Improvement in Glucose Metabolism Is Associated with Increased Abundance of Prevotella. Cell Metab. 2015, 22, 971–982. [Google Scholar] [CrossRef] [Green Version]
- Zegers-Hochschild, F.; Adamson, G.D.; Dyer, S.; Racowsky, C.; de Mouzon, J.; Sokol, R.; Rienzi, L.; Sunde, A.; Schmidt, L.; Cooke, I.D.; et al. The International Glossary on Infertility and Fertility Care, 2017. Fertil. Steril. 2017, 108, 393–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, B.-F. Predictive values of body mass index and waist circumference for risk factors of certain related diseases in Chinese adults-study on optimal cut-off points of body mass index and waist circumference in Chinese adults. Biomed. Environ. Sci. 2002, 15, 5–10. [Google Scholar]
- Robinson, N.J.; Winge, D.R. Copper Metallochaperones. Annu. Rev. Biochem. 2010, 79, 537–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, X.; Chen, X.; Yin, N.; Du, H.; Sun, G.; Wang, L.; Xu, Y.; Chen, Y.; Cui, Y. Estimation of the bioaccessibility and bioavailability of Fe, Mn, Cu, and Zn in Chinese vegetables using the in vitro digestion/Caco-2 cell model: The influence of gut microbiota. Food Funct. 2017, 8, 4592–4600. [Google Scholar] [CrossRef] [PubMed]
- Knezevic, J.; Starchl, C.; Berisha, A.T.; Amrein, K. Thyroid-Gut-Axis: How Does the Microbiota Influence Thyroid Function? Nutrients 2020, 12, 1769. [Google Scholar] [CrossRef]
- Bielik, V.; Kolisek, M. Bioaccessibility and Bioavailability of Minerals in Relation to a Healthy Gut Microbiome. Int. J. Mol. Sci. 2021, 22, 6803. [Google Scholar] [CrossRef] [PubMed]
- Whisner, C.M.; Martin, B.R.; Nakatsu, C.H.; McCabe, G.P.; McCabe, L.D.; Peacock, M.; Weaver, C.M. Soluble maize fibre affects short-term calcium absorption in adolescent boys and girls: A randomised controlled trial using dual stable isotopic tracers. Br. J. Nutr. 2014, 112, 446–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whisner, C.M.; Martin, B.R.; Nakatsu, C.H.; A Story, J.; Macdonald-Clarke, C.J.; McCabe, L.D.; McCabe, G.P.; Weaver, C.M. Soluble Corn Fiber Increases Calcium Absorption Associated with Shifts in the Gut Microbiome: A Randomized Dose-Response Trial in Free-Living Pubertal Females. J. Nutr. 2016, 146, 1298–1306. [Google Scholar] [CrossRef] [Green Version]
- Aljewicz, M.; Siemianowska, E.; Cichosz, G.; Tońska, E. The effect of probiotics (Lactobacillus rhamnosus HN001, Lactobacillus paracasei LPC-37, and Lactobacillus acidophilus NCFM) on the availability of minerals from Dutch-type cheese. J. Dairy Sci. 2014, 97, 4824–4831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergillos-Meca, T.; Cabrera-Vique, C.; Artacho, R.; Moreno-Montoro, M.; Navarro-Alarcón, M.; Olalla, M.; Giménez, R.; Seiquer, I.; Ruiz-López, M.D. Does Lactobacillus plantarum or ultrafiltration process improve Ca, Mg, Zn and P bioavailability from fermented goats’ milk? Food Chem. 2015, 187, 314–321. [Google Scholar] [CrossRef]
- Lane, D.J.R.; Bae, D.-H.; Merlot, A.M.; Sahni, S.; Richardson, D.R. Duodenal Cytochrome b (DCYTB) in Iron Metabolism: An Update on Function and Regulation. Nutrients 2015, 7, 2274–2296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luu, Y.-S.; Ramsay, J.A. Review: Microbial mechanisms of accessing insoluble Fe(III) as an energy source. World J. Microbiol. Biotechnol. 2003, 19, 215–225. [Google Scholar] [CrossRef]
- Das, N.K.; Schwartz, A.J.; Barthel, G.; Inohara, N.; Liu, Q.; Sankar, A.; Hill, D.R.; Ma, X.; Lamberg, O.; Schnizlein, M.K.; et al. Microbial Metabolite Signaling Is Required for Systemic Iron Homeostasis. Cell Metab. 2019, 31, 115–130. [Google Scholar] [CrossRef] [PubMed]
- Metwally, M.; Ong, K.; Ledger, W.; Li, T.C. Does high body mass index increase the risk of miscarriage after spontaneous and assisted conception? A meta-analysis of the evidence. Fertil. Steril. 2008, 90, 714–726. [Google Scholar] [CrossRef] [PubMed]
- Bellver, J.; Ayllón, Y.; Ferrando, M.; Melo, M.; Goyri, E.; Pellicer, A.; Remohí, J.; Meseguer, M. Female obesity impairs in vitro fertilization outcome without affecting embryo quality. Fertil. Steril. 2010, 93, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Luke, B.; Brown, M.B.; Stern, J.E.; Missmer, S.A.; Fujimoto, V.Y.; Leach, R.; A SART Writing Group. Female obesity adversely affects assisted reproductive technology (ART) pregnancy and live birth rates. Hum. Reprod. 2010, 26, 245–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Zheng, D.; Wu, H.; Li, R.; Xu, S.; Kang, Y.; Cao, Y.; Chen, X.; Zhu, Y.; Chen, Z.-J.; et al. Epidemiology of infertility in China: A population-based study. BJOG Int. J. Obstet. Gynaecol. 2017, 125, 432–441. [Google Scholar] [CrossRef] [Green Version]
- Grieger, J.A.; Grzeskowiak, L.E.; Wilson, R.L.; Bianco-Miotto, T.; Leemaqz, S.Y.; Jankovic-Karasoulos, T.; Perkins, A.V.; Norman, R.J.; Dekker, G.A.; Roberts, C.T. Maternal Selenium, Copper and Zinc Concentrations in Early Pregnancy, and the Association with Fertility. Nutrients 2019, 11, 1609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chavarro, J.E.; Rich-Edwards, J.W.; Rosner, B.A.; Willett, W.C. Iron Intake and Risk of Ovulatory Infertility. Obstet. Gynecol. 2006, 108, 1145–1152. [Google Scholar] [CrossRef] [PubMed]
- Janiszewska, J.; Ostrowska, J.; Szostak-Węgierek, D. Milk and Dairy Products and Their Impact on Carbohydrate Metabolism and Fertility—A Potential Role in the Diet of Women with Polycystic Ovary Syndrome. Nutrients 2020, 12, 3491. [Google Scholar] [CrossRef] [PubMed]
- Barceloux, D.G. Copper. J. Toxicol. Clin. Toxicol. 1999, 37, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Geissler, C.; Singh, M. Iron, meat and health. Nutrients 2011, 3, 283–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, H.; Kawamura, K.; Kawamura, T.; Odamaki, T.; Katsumata, N.; Xiao, J.-Z.; Suzuki, N.; Tanaka, M. Distinctive subpopulations of the intestinal microbiota are present in women with unexplained chronic anovulation. Reprod. Biomed. Online 2018, 38, 570–578. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Ming, Q.; Liang, J.; Zhang, Y.; Zhang, H.; Shen, T. Gut microbiota dysbiosis in polycystic ovary syndrome: Association with obesity—A preliminary report. Can. J. Physiol. Pharmacol. 2020, 98, 803–809. [Google Scholar] [CrossRef] [PubMed]
- Insenser, M.; Murri, M.; Del Campo, R.; Martínez-García, M.; Fernández-Durán, E.; Escobar-Morreale, H.F. Gut Microbiota and the Polycystic Ovary Syndrome: Influence of Sex, Sex Hormones, and Obesity. J. Clin. Endocrinol. Metab. 2018, 103, 2552–2562. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Yun, C.; Sun, L.; Xia, J.; Wu, Q.; Wang, L.; Wang, L.; Zhang, Y.; Liang, X.; Gonzalez, F.J.; et al. Gut microbiota–bile acid–interleukin-22 axis orchestrates polycystic ovary syndrome. Nat. Med. 2019, 25, 1225–1233. [Google Scholar] [CrossRef] [PubMed]
- Jobira, B.; Frank, D.N.; Pyle, L.; Silveira, L.J.; Kelsey, M.M.; Garcia-Reyes, Y.; E Robertson, C.; Ir, D.; Nadeau, K.J.; Cree-Green, M. Obese Adolescents with PCOS Have Altered Biodiversity and Relative Abundance in Gastrointestinal Microbiota. J. Clin. Endocrinol. Metab. 2020, 105, e2134–e2144. [Google Scholar] [CrossRef]
- Li, G.; Yin, P.; Chu, S.; Gao, W.; Cui, S.; Guo, S.; Xu, Y.; Yuan, E.; Zhu, T.; You, J.; et al. Correlation Analysis between GDM and Gut Microbial Composition in Late Pregnancy. J. Diabetes Res. 2021, 2021, 8892849. [Google Scholar] [CrossRef] [PubMed]
- Jaeggi, T.; Kortman, G.A.M.; Moretti, D.; Chassard, C.; Holding, P.; Dostal, A.; Boekhorst, J.; Timmerman, H.M.; Swinkels, D.W.; Tjalsma, H.; et al. Iron fortification adversely affects the gut microbiome, increases pathogen abundance and induces intestinal inflammation in Kenyan infants. Gut 2014, 64, 731–742. [Google Scholar] [CrossRef] [PubMed]
- Muleviciene, A.; D’Amico, F.; Turroni, S.; Candela, M.; Jankauskiene, A. Iron deficiency anemia-related gut microbiota dysbiosis in infants and young children: A pilot study. Acta Microbiol. Immunol. Hung. 2018, 65, 551–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dillon, S.M.; Lee, E.J.; Kotter, C.V.; Austin, G.L.; Gianella, S.; Siewe, B.; Smith, D.M.; Landay, A.L.; McManus, M.C.; Robertson, C.E.; et al. Gut dendritic cell activation links an altered colonic microbiome to mucosal and systemic T-cell activation in untreated HIV-1 infection. Mucosal Immunol. 2015, 9, 24–37. [Google Scholar] [CrossRef]
- Kanamori, Y.; Murakami, M.; Sugiyama, M.; Hashimoto, O.; Matsui, T.; Funaba, M. Hepcidin and IL-1β. Vitam. Horm. 2019, 110, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Visitchanakun, P.; Saisorn, W.; Wongphoom, J.; Chatthanathon, P.; Somboonna, N.; Svasti, S.; Fucharoen, S.; Leelahavanichkul, A. Gut leakage enhances sepsis susceptibility in iron-overloaded β-thalassemia mice through macrophage hyperinflammatory responses. Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 318, G966–G979. [Google Scholar] [CrossRef] [PubMed]
- Cuisiniere, T.; Calvé, A.; Fragoso, G.; Oliero, M.; Hajjar, R.; Gonzalez, E.; Santos, M.M. Oral iron supplementation after antibiotic exposure induces a deleterious recovery of the gut microbiota. BMC Microbiol. 2021, 21, 259. [Google Scholar] [CrossRef] [PubMed]
- Ganz, T. Macrophages and Iron Metabolism. Microbiol. Spectr. 2016, 4, 492–504. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Prescott, R.; Cherayil, B.J. The commensal bacterium Bacteroides fragilis down-regulates ferroportin expression and alters iron homeostasis in macrophages. J. Leukoc. Biol. 2019, 106, 1079–1088. [Google Scholar] [CrossRef] [PubMed]
- Saleh, F.; Behbehani, A.; Asfar, S.; Khan, I.; Ibrahim, G. Abnormal Blood Levels of Trace Elements and Metals, DNA Damage, and Breast Cancer in the State of Kuwait. Biol. Trace Element Res. 2010, 141, 96–109. [Google Scholar] [CrossRef]
- Verma, S.; Cherayil, B.J. Iron and inflammation—The gut reaction. Metallomics 2017, 9, 101–111. [Google Scholar] [CrossRef] [Green Version]
- Arezes, J.; Jung, G.; Gabayan, V.; Valore, E.; Ruchala, P.; Gulig, P.A.; Ganz, T.; Nemeth, E.; Bulut, Y. Hepcidin-Induced Hypoferremia Is a Critical Host Defense Mechanism against the Siderophilic Bacterium Vibrio vulnificus. Cell Host Microbe 2015, 17, 47–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Zhang, X.; Shen, Y.; Fang, X.; Wang, Y.; Wang, F. Obesity and iron deficiency: A quantitative meta-analysis. Obes. Rev. 2015, 16, 1081–1093. [Google Scholar] [CrossRef]
- González-Domínguez, Á.; Visiedo-García, F.M.; Domínguez-Riscart, J.; González-Domínguez, R.; Mateos, R.M.; Lechuga-Sancho, A.M. Iron Metabolism in Obesity and Metabolic Syndrome. Int. J. Mol. Sci. 2020, 21, 5529. [Google Scholar] [CrossRef] [PubMed]
- Precup, G.; Vodnar, D.-C. Gut Prevotella as a possible biomarker of diet and its eubiotic versus dysbiotic roles: A comprehensive literature review. Br. J. Nutr. 2019, 122, 131–140. [Google Scholar] [CrossRef]
- Nirmalkar, K.; Murugesan, S.; Pizano-Zárate, M.L.; Villalobos-Flores, L.E.; García-González, C.; Morales-Hernández, R.M.; Nuñez-Hernández, J.A.; Hernández-Quiroz, F.; Romero-Figueroa, M.D.S.; Hernández-Guerrero, C.; et al. Gut Microbiota and Endothelial Dysfunction Markers in Obese Mexican Children and Adolescents. Nutrients 2018, 10, 2009. [Google Scholar] [CrossRef] [Green Version]
- Meldrum, D.R. Introduction. Fertil. Steril. 2017, 107, 831–832. [Google Scholar] [CrossRef] [PubMed]
- Christensen, L.; Vuholm, S.; Roager, H.M.; Nielsen, D.S.; Krych, L.; Kristensen, M.; Astrup, A.; Hjorth, M. Prevotella Abundance Predicts Weight Loss Success in Healthy, Overweight Adults Consuming a Whole-Grain Diet Ad Libitum: A Post Hoc Analysis of a 6-Wk Randomized Controlled Trial. J. Nutr. 2019, 149, 2174–2181. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Infertile Group | Control Group | P Value |
|---|---|---|---|
| Baseline | |||
| N (%) | 469 (72.04) | 182 (27.96) | |
| Log(P/B) | −0.32 (−3.45, 0.53) | −2.86 (−3.76, 0.03) | <0.001 |
| Age (years) | 33.04 ± 4.29 | 33.32 ± 4.37 | 0.459 |
| BMI (kg/m2) | 23.68 ± 2.80 | 22.69 ± 4.09 | 0.001 |
| Current smoking, N (%) | 14 (2.99) | 3 (1.65) | 0.423 |
| Trace elements | |||
| Cu (μmol/L) | 15.89 ± 3.95 | 16.25 ± 3.75 | 0.083 |
| Zn (μmol/L) | 94.07 ± 12.41 | 97.59 ± 11.55 | 0.001 |
| Ca (μmol/L) | 1.67 ± 0.17 | 1.65 ± 0.18 | 0.206 |
| Mg (μmol/L) | 1.46 ± 0.14 | 1.50 ± 0.13 | <0.001 |
| Fe (μmol/L) | 8.39 ± 0.71 | 8.84 ± 0.56 | <0.001 |
| Education, N (%) | <0.001 | ||
| Middle school and below | 68 (10.45) | 13 (1.20) | |
| High school | 84 (12.90) | 17 (2.61) | |
| College | 238 (36.56) | 97 (14.90) | |
| Master and above | 79 (12.14) | 55 (9.24) | |
| Diagnosis, N (%) | |||
| RSA | 114 (24.31) | − | |
| PCOS | 81 (17.27) | − | |
| RIF | 43 (9.17) | − | |
| DOR | 17 (3.62) | − | |
| Tubal factor infertility/endometriosis | 128 (27.29) | − | |
| Unexplained infertility | 86 (18.34) | − | |
| Childbirth and abortion history | |||
| Previous pregnancies | 93 | 58 | |
| Previous deliveries | 74 | 20 | |
| Previous spontaneous abortions | 101 | 9 |
| Control Group (n = 182) | Infertile Group (n = 469) | |||||
|---|---|---|---|---|---|---|
| P-Enterotype | B-Enterotype | P Value | P-Enterotype | B-Enterotype | P Value | |
| N | 106 | 76 | 220 | 249 | ||
| Cu (μmol/L) | 15.70 ± 3.66 | 16.33 ± 3.87 | 0.262 | 15.48 ± 4.28 | 16.26 ± 3.59 | 0.033 |
| Zn (μmol/L) | 97.42 ± 12.44 | 97.83 ± 10.27 | 0.815 | 91.71 ± 11.99 | 96.16 ± 12.41 | <0.001 |
| Ca (μmol/L) | 1.66 ± 0.20 | 1.64 ± 0.15 | 0.441 | 1.67 ± 0.19 | 1.66 ± 0.16 | 0.549 |
| Mg (μmol/L) | 1.50 ± 0.13 | 1.51 ± 0.13 | 0.586 | 1.42 ± 0.12 | 1.49 ± 0.14 | <0.001 |
| Fe (μmol/L) | 8.82 ± 0.56 | 8.86 ± 0.57 | 0.591 | 8.17 ± 0.64 | 8.58 ± 0.71 | <0.001 |
| Trace Element | Status | Concentration (µmol/L) | P-Enterotype (N) | B-Enterotype (N) | χ2 | P Value |
|---|---|---|---|---|---|---|
| Cu | LL | <9.25 | 15 | 8 | 3.300 | 0.187 |
| NL | 9.25–21.21 | 193 | 228 | |||
| HL | >21.21 | 12 | 13 | |||
| Zn | LL | <74.66 | 11 | 12 | 5.420 | 0.063 |
| NL | 74.66-111.17 | 204 | 220 | |||
| HL | >111.17 | 5 | 17 | |||
| Ca | LL | <1.42 | 6 | 9 | 3.168 | 0.186 |
| NL | 1.42–1.90 | 202 | 234 | |||
| HL | >1.90 | 12 | 6 | |||
| Mg | LL | <1.25 | 6 | 6 | 4.048 | 0.140 |
| NL | 1.25-1.70 | 209 | 228 | |||
| HL | >1.70 | 5 | 15 | |||
| Fe | LL | <7.37 | 14 | 9 | 13.309 | 0.001 |
| NL | 7.37–9.60 | 203 | 220 | |||
| HL | >9.60 | 3 | 20 |
| Characteristic | Whole Blood Iron Status | P Value a | P Value b | P Value | ||
|---|---|---|---|---|---|---|
| LL | NL | HL | ||||
| N (%) | 23 | 423 | 23 | |||
| Log(P/B) | −0.57 (−3.15, 0.36) | −2.65 (−3.77, 0.03) | −3.71 (−4.05, −3.06) | 0.013 | 0.006 | 0.002 |
| Age (years) | 33.00 (30.00, 34.00) | 33.00 (31.00, 36.00) | 32.00 (30.00, 36.00) | 0.669 | 0.606 | 0.998 |
| BMI (kg/m2) | 23.59 (20.57, 24.80) | 23.80(22.00, 26.50) | 23.20 (21.17, 26.38) | 0.043 | 0.482 | 0.045 |
| Diet in the last month (N) | 0.134 | 0.325 | 0.184 | |||
| A vegetarian diet | 2 | 10 | 0 | |||
| A meat-based diet | 1 | 17 | 2 | |||
| A meat and vegetarian diet | 20 | 396 | 21 | |||
| Drinking in the last month (N) | 0.945 | 0.751 | 0.953 | |||
| None | 21 | 386 | 22 | |||
| ≤3 times a month | 2 | 35 | 1 | |||
| >3 times a month | 0 | 2 | 0 | |||
| AUC | SE | Cutoff | 95% CI | Sensitivity % | Specificity % | P Value | |
|---|---|---|---|---|---|---|---|
| Lean/normal | 0.594 | 0.054 | −0.64 | 0.487–0.700 | 84.2 | 61.3 | 0.172 |
| Overweight | 0.599 | 0.073 | −3.23 | 0.454–0.743 | 51.1 | 96.4 | 0.633 |
| Obesity | 0.894 | 0.087 | −0.27 | 0.724–0.993 | 98.8 | 78.8 | 0.042 |
| AUC | SE | Cutoff | 95% CI | Sensitivity % | Specificity % | P Value | |
|---|---|---|---|---|---|---|---|
| Lean/normal | 0.648 | 0.052 | −2.76 | 0.546–0.750 | 55.7 | 82.4 | 0.041 |
| Overweight | 0.863 | 0.040 | −3.99 | 0.786–0.941 | 78.1 | 92.7 | 0.013 |
| Obesity | 0.818 | 0.088 | −3.70 | 0.645–0.992 | 72.7 | 78.3 | 0.136 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, X.; Zuo, N.; Guan, W.; Fu, L.; Jiang, S.; Jiao, J.; Wang, X. Association of Gut Microbiota Enterotypes with Blood Trace Elements in Women with Infertility. Nutrients 2022, 14, 3195. https://doi.org/10.3390/nu14153195
Yao X, Zuo N, Guan W, Fu L, Jiang S, Jiao J, Wang X. Association of Gut Microbiota Enterotypes with Blood Trace Elements in Women with Infertility. Nutrients. 2022; 14(15):3195. https://doi.org/10.3390/nu14153195
Chicago/Turabian StyleYao, Xinrui, Na Zuo, Wenzheng Guan, Lingjie Fu, Shuyi Jiang, Jiao Jiao, and Xiuxia Wang. 2022. "Association of Gut Microbiota Enterotypes with Blood Trace Elements in Women with Infertility" Nutrients 14, no. 15: 3195. https://doi.org/10.3390/nu14153195
APA StyleYao, X., Zuo, N., Guan, W., Fu, L., Jiang, S., Jiao, J., & Wang, X. (2022). Association of Gut Microbiota Enterotypes with Blood Trace Elements in Women with Infertility. Nutrients, 14(15), 3195. https://doi.org/10.3390/nu14153195