
Association and Interaction Effect of BHMT Gene Polymorphisms and Maternal Dietary Habits with Ventricular Septal Defect in Offspring

 ,
,
Abstract
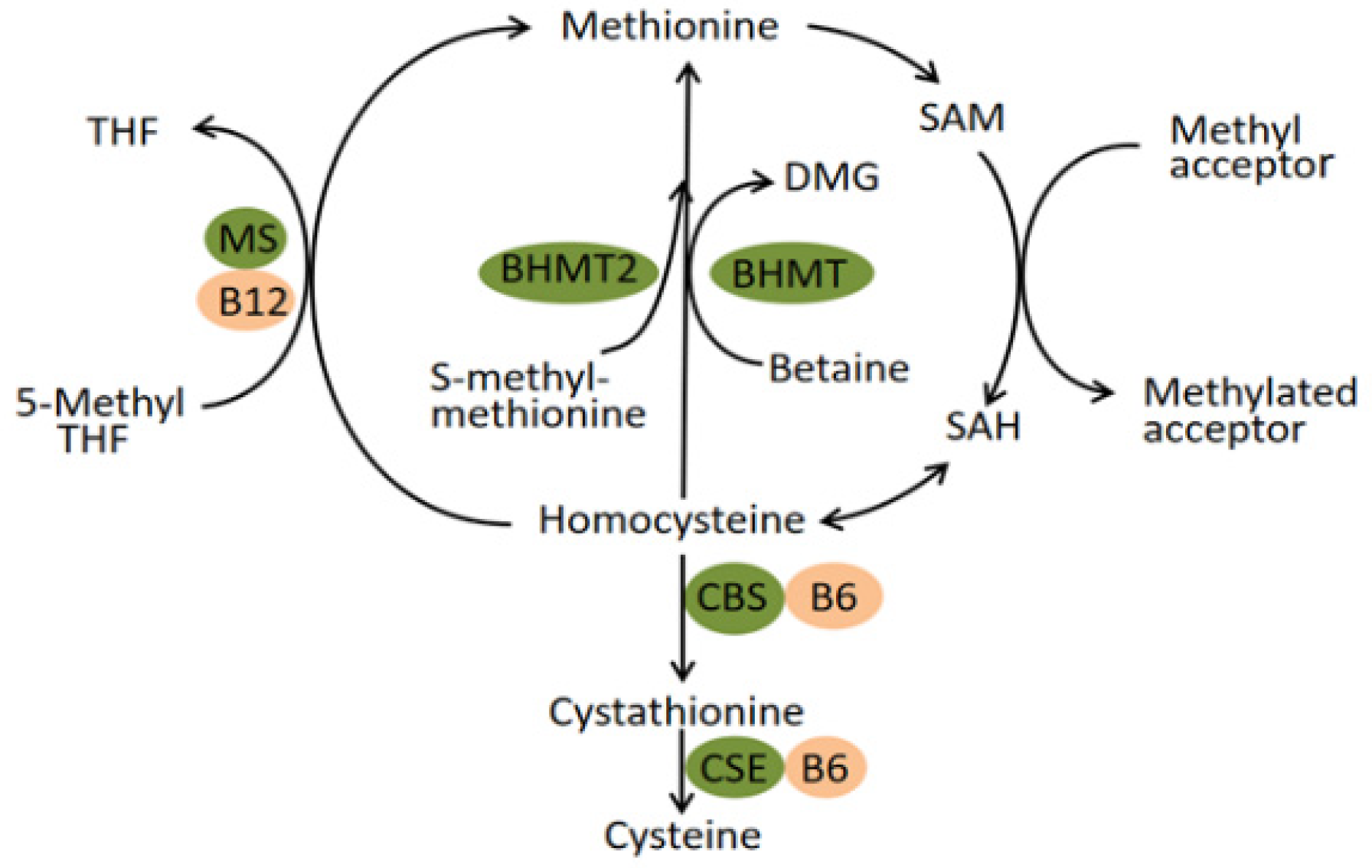
1. Introduction
2. Materials and Methods
2.1. Design and Participants
2.2. Information Collection
2.3. Sample Collection and Genotyping
2.4. Statistical Analysis
3. Results
3.1. Comparison of Maternal Baseline Characteristics
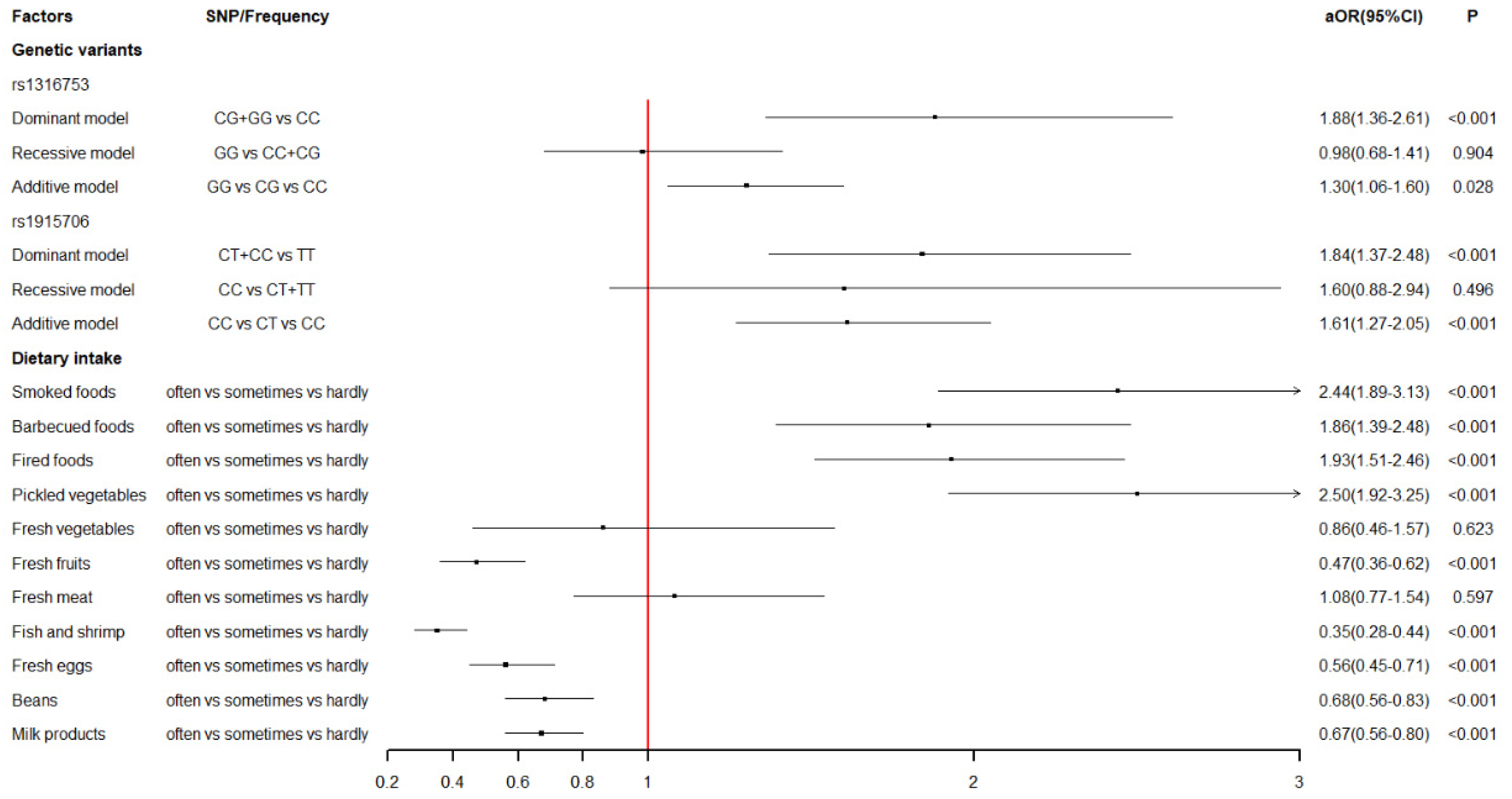
3.2. Maternal Dietary Habits and the Risk of VSD in Offspring
3.3. Maternal BHMT Gene Polymorphisms and the Risk of VSD in Offspring
3.4. Interaction of the Polymorphisms of BHMT Gene and Maternal Dietary Habits on the Risk of VSD in Offspring
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, Y.; Chen, S.; Zühlke, L.; Black, G.C.; Choy, M.K.; Li, N.; Keavney, B.D. Global birth prevalence of congenital heart defects 1970-2017: Updated systematic review and meta-analysis of 260 studies. Int. J. Epidemiol. 2019, 48, 455–463. [Google Scholar]
- Mitchell, S.C.; Korones, S.B.; Berendes, H.W. Congenital heart disease in 56, 109 births. Incidence and natural history. Circulation 1971, 43, 323–332. [Google Scholar]
- Van Der Linde, D.; Konings, E.E.; Slager, M.A.; Witsenburg, M.; Helbing, W.A.; Takkenberg, J.J.; Roos-Hesselink, J.W. Birth prevalence of congenital heart disease worldwide: A systematic review and meta-analysis. J. Am. Coll. Cardiol. 2011, 58, 2241–2247. [Google Scholar] [CrossRef]
- Penny, D.J.; Vick, G.W., 3rd. Ventricular septal defect. Lancet 2011, 377, 1103–1112. [Google Scholar] [CrossRef]
- Gelb, B.D.; Chung, W.K. Complex genetics and the etiology of human congenital heart disease. Cold Spring Harb. Perspect. Med. 2014, 4, a013953. [Google Scholar] [CrossRef]
- Zhang, T.N.; Wu, Q.J.; Liu, Y.S.; Lv, J.L.; Sun, H.; Chang, Q.; Liu, C.F.; Zhao, Y.H. Environmental Risk Factors and Congenital Heart Disease: An Umbrella Review of 165 Systematic Reviews and Meta-Analyses with More than 120 Million Participants. Front. Cardiovasc Med. 2021, 8, 640729. [Google Scholar] [CrossRef]
- Morton, S.U.; Quiat, D.; Seidman, J.G.; Seidman, C.E. Genomic frontiers in congenital heart disease. Nat. Rev. Cardiol. 2022, 19, 26–42. [Google Scholar]
- Boyd, R.; McMullen, H.; Beqaj, H.; Kalfa, D. Environmental Exposures and Congenital Heart Disease. Pediatrics 2022, 149, e2021052151. [Google Scholar]
- van der Bom, T.; Zomer, A.C.; Zwinderman, A.H.; Meijboom, F.J.; Bouma, B.J.; Mulder, B.J. The changing epidemiology of congenital heart disease. Nat. Rev. Cardiol. 2011, 8, 50–60. [Google Scholar] [CrossRef]
- Cao, J.; Wu, Q.; Huang, Y.; Wang, L.; Su, Z.; Ye, H. The role of DNA methylation in syndromic and non-syndromic congenital heart disease. Clin. Epigenetics. 2021, 13, 93. [Google Scholar] [CrossRef]
- Wilson, R.D.; O’Connor, D.L. Maternal folic acid and multivitamin supplementation: International clinical evidence with considerations for the prevention of folate-sensitive birth defects. Prev. Med. Rep. 2021, 24, 101617. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Cheng, Y.; Zeng, L.; Dang, S.; Yan, H. Maternal dietary diversity during pregnancy and congenital heart defects: A case-control study. Eur. J. Clin. Nutr. 2021, 75, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Kang, Y.; Chang, Q.; Zhang, B.; Liu, X.; Zeng, L.; Yan, H.; Dang, S. Maternal Zinc, Copper, and Selenium Intakes during Pregnancy and Congenital Heart Defects. Nutrients 2022, 14, 1055. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Guo, L.; Zhao, D.; Qu, P.; Dang, S.; Yan, H. Maternal B-vitamin intake and B-vitamin supplementation during pregnancy in relation to neonatal congenital heart defects: A case-control study with propensity score matching. Eur. J. Clin. Nutr. 2021, 75, 782–791. [Google Scholar] [CrossRef]
- Koster, M.P.; van Duijn, L.; Krul-Poel, Y.H.; Laven, J.S.; Helbing, W.A.; Simsek, S.; Steegers-Theunissen, R.P. A compromised maternal vitamin D status is associated with congenital heart defects in offspring. Early Hum. Dev. 2018, 117, 50–56. [Google Scholar] [CrossRef]
- Sunden, S.L.; Renduchintala, M.S.; Park, E.I.; Miklasz, S.D.; Garrow, T.A. Betaine-homocysteine methyltransferase expression in porcine and human tissues and chromosomal localization of the human gene. Arch. Biochem. Biophys. 1997, 345, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Park, E.I.; Garrow, T.A. Interaction between dietary methionine and methyl donor intake on rat liver betaine-homocysteine methyltransferase gene expression and organization of the human gene. J. Biol. Chem. 1999, 274, 7816–7824. [Google Scholar] [CrossRef] [PubMed]
- Teng, Y.W.; Cerdena, I.; Zeisel, S.H. Homocysteinemia in mice with genetic betaine homocysteine S-methyltransferase deficiency is independent of dietary folate intake. J. Nutr. 2012, 142, 1964–1967. [Google Scholar] [CrossRef]
- Szegedi, S.S.; Castro, C.C.; Koutmos, M.; Garrow, T.A. Betaine-homocysteine S-methyltransferase-2 is an S-methylmethionine-homocysteine methyltransferase. J. Biol. Chem. 2008, 283, 8939–8945. [Google Scholar] [CrossRef]
- Song, J.H.; Lee, H.R.; Shim, S.M. Determination of S-methyl-L-methionine (SMM) from Brassicaceae Family Vegetables and Characterization of the Intestinal Transport of SMM by Caco-2 Cells. J. Food Sci. 2017, 82, 36–43. [Google Scholar] [CrossRef]
- Pérez-Miguelsanz, J.; Vallecillo, N.; Garrido, F.; Reytor, E.; Pérez-Sala, D.; Pajares, M.A. Betaine homocysteine S-methyltransferase emerges as a new player of the nuclear methionine cycle. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 1165–1182. [Google Scholar] [CrossRef] [PubMed]
- Feng, Q.; Kalari, K.; Fridley, B.L.; Jenkins, G.; Ji, Y.; Abo, R.; Hebbring, S.; Zhang, J.; Nye, M.D.; Leeder, J.S.; et al. Betaine-homocysteine methyltransferase: Human liver genotype-phenotype correlation. Mol. Genet. Metab. 2011, 102, 126–133. [Google Scholar] [CrossRef]
- Blanco, R.; Colombo, A.; Pardo, R.; Suazo, J. Maternal biomarkers of methylation status and non-syndromic orofacial cleft risk: A meta-analysis. Int. J. Oral Maxillofac. Surg. 2016, 45, 1323–1332. [Google Scholar] [CrossRef]
- Kucha, W.; Seifu, D.; Tirsit, A.; Yigeremu, M.; Abebe, M.; Hailu, D.; Tsehay, D.; Genet, S. Folate, Vitamin B12, and Homocysteine Levels in Women With Neural Tube Defect-Affected Pregnancy in Addis Ababa, Ethiopia. Front. Nutr. 2022, 9, 873900. [Google Scholar] [CrossRef]
- Dilli, D.; Doğan, N.N.; Örün, U.A.; Koç, M.; Zenciroğlu, A.; Karademir, S.; Akduman, H. Maternal neonatal micronutrient levels in newborns with, C.H.D. Cardiol. Young 2018, 28, 523–529. [Google Scholar] [CrossRef]
- Nelen, W.L. Hyperhomocysteinaemia and human reproduction. Clin. Chem. Lab. Med. 2001, 39, 758–763. [Google Scholar] [CrossRef]
- Shaw, G.M.; Lu, W.; Zhu, H.; Yang, W.; Briggs, F.; Carmichael, S.L.; Barcellos, L.F.; Lammer, E.J.; Finnell, R.H. 118 SNPs of folate-related genes and risks of spina bifida and conotruncal heart defects. BMC Med. Genet. 2009, 10, 49. [Google Scholar] [CrossRef]
- Hobbs, C.A.; Cleves, M.A.; MacLeod, S.L.; Erickson, S.W.; Tang, X.; Li, J.; Li, M.; Nick, T.; Malik, S.; National Birth Defects Prevention Study. Conotruncal heart defects and common variants in maternal and fetal genes in folate, homocysteine, and transsulfuration pathways. Birth Defects Res. A Clin. Mol. Teratol. 2014, 100, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Nembhard, W.N.; Tang, X.; Hu, Z.; MacLeod, S.; Stowe, Z.; Webber, D. Maternal and infant genetic variants, maternal periconceptional use of selective serotonin reuptake inhibitors, and risk of congenital heart defects in offspring: Population based study. Bmj 2017, 356, j832. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Tang, X.; Cleves, M.A.; Nick, T.G.; Li, M.; MacLeod, S.L.; Erickson, S.W.; Li, J.; Shaw, G.M.; Mosley, B.S.; National Birth Defects Prevention Study. Obstructive heart defects associated with candidate genes, maternal obesity, and folic acid supplementation. Am. J. Med. Genet. A 2015, 167, 1231–1242. [Google Scholar] [CrossRef]
- Ganu, R.S.; Ishida, Y.; Koutmos, M.; Kolokotronis, S.O.; Roca, A.L.; Garrow, T.A.; Schook, L.B. Evolutionary Analyses and Natural Selection of Betaine-Homocysteine S-Methyltransferase (BHMT) and BHMT2 Genes. PLoS ONE 2015, 10, e0134084. [Google Scholar] [CrossRef] [PubMed]
- Wallace, T.C.; Blusztajn, J.K.; Caudill, M.A.; Klatt, K.C.; Zeisel, S.H. Choline: The Neurocognitive Essential Nutrient of Interest to Obstetricians and Gynecologists. J. Diet Suppl. 2020, 17, 733–752. [Google Scholar] [CrossRef] [PubMed]
- Vailati-Riboni, M.; Crookenden, M.; Kay, J.K.; Meier, S.; Mitchell, M.D.; Heiser, A.; Roche, J.R.; Loor, J.J. Hepatic one-carbon metabolism enzyme activities and intermediate metabolites are altered by prepartum body condition score and plane of nutrition in grazing Holstein dairy cows. J. Dairy Sci. 2020, 103, 2662–2676. [Google Scholar] [CrossRef] [PubMed]
- Jacometo, C.B.; Zhou, Z.; Luchini, D.; Corrêa, M.N.; Loor, J.J. Maternal supplementation with rumen-protected methionine increases prepartal plasma methionine concentration and alters hepatic mRNA abundance of 1-carbon, methionine, and transsulfuration pathways in neonatal Holstein calves. J. Dairy Sci. 2017, 100, 3209–3219. [Google Scholar] [CrossRef] [PubMed]
- Coleman, D.N.; Vailati-Riboni, M.; Elolimy, A.A.; Cardoso, F.C.; Rodriguez-Zas, S.L.; Miura, M.; Pan, Y.X.; Loor, J.J. Hepatic betaine-homocysteine methyltransferase and methionine synthase activity and intermediates of the methionine cycle are altered by choline supply during negative energy balance in Holstein cows. J. Dairy Sci. 2019, 102, 8305–8318. [Google Scholar] [CrossRef] [PubMed]
- Petersen, J.M.; Parker, S.E.; Crider, K.S.; Tinker, S.C.; Mitchell, A.A.; Werler, M.M. One-Carbon Cofactor Intake and Risk of Neural Tube Defects Among Women Who Meet Folic Acid Recommendations: A Multicenter Case-Control Study. Am. J. Epidemiol. 2019, 188, 1136–1143. [Google Scholar] [CrossRef] [PubMed]
- Stingone, J.A.; Luben, T.J.; Carmichael, S.L.; Aylsworth, A.S.; Botto, L.D.; Correa, A.; Gilboa, S.M.; Langlois, P.H.; Nembhard, W.N.; Richmond-Bryant, J.; et al. Maternal Exposure to Nitrogen Dioxide, Intake of Methyl Nutrients, and Congenital Heart Defects in Offspring. Am. J. Epidemiol. 2017, 186, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Bedale, W.; Sindelar, J.J.; Milkowski, A.L. Dietary nitrate and nitrite: Benefits, risks, and evolving perceptions. Meat Sci. 2016, 120, 85–92. [Google Scholar] [CrossRef]
- Romanowicz, J.; Guerrelli, D.; Dhari, Z.; Mulvany, C.; Reilly, M.; Swift, L.; Vasandani, N.; Ramadan, M.; Leatherbury, L.; Ishibashi, N.; et al. Chronic perinatal hypoxia delays cardiac maturation in a mouse model for cyanotic congenital heart disease. Am. J. Physiol. Heart Circ. Physiol. 2021, 320, H1873–H1886. [Google Scholar] [CrossRef]
- Friedman, M. Chemistry, biochemistry, and safety of acrylamide. A review. J. Agric. Food Chem. 2003, 51, 4504–4526. [Google Scholar] [CrossRef] [PubMed]
- Tareke, E.; Rydberg, P.; Karlsson, P.; Eriksson, S.; Törnqvist, M. Analysis of acrylamide, a carcinogen formed in heated foodstuffs. J. Agric. Food Chem. 2002, 50, 4998–5006. [Google Scholar] [CrossRef] [PubMed]
- Bušová, M.; Bencko, V.; Veszelits Laktičová, K.; Holcátová, I.; Vargová, M. Risk of exposure to acrylamide. Cent. Eur. J. Public Health 2020, 28, S43–S46. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Li, N.; Liu, Z.; Qiu, J.; Deng, Y.; Li, X.; Chen, M.; Yu, J.; Zhu, J.; Yu, P.; et al. Risk of congenital heart diseases associated with NAT2 genetic polymorphisms and maternal polycyclic aromatic hydrocarbons exposure. Prenat. Diagn. 2019, 39, 968–975. [Google Scholar] [CrossRef]
- Sarmah, S.; Marrs, J.A. Zebrafish as a Vertebrate Model System to Evaluate Effects of Environmental Toxicants on Cardiac Development and Function. Int. J. Mol. Sci. 2016, 17, 2123. [Google Scholar] [CrossRef]
- Huang, L.; Wang, C.; Zhang, Y.; Li, J.; Zhong, Y.; Zhou, Y.; Chen, Y.; Zuo, Z. Benzo[a]pyrene exposure influences the cardiac development and the expression of cardiovascular relative genes in zebrafish (Danio rerio) embryos. Chemosphere 2012, 87, 369–375. [Google Scholar] [CrossRef]
- Hobbs, C.A.; Cleves, M.A.; Karim, M.A.; Zhao, W.; MacLeod, S.L. Maternal folate-related gene environment interactions and congenital heart defects. Obstet Gynecol. 2010, 116 Pt 1, 16–22. [Google Scholar] [CrossRef]
- Cao, L.; Wang, Y.; Zhang, R.; Dong, L.; Cui, H.; Fang, Y.; Zhao, L.; Shi, O.; Cai, C. Association of neural tube defects with gene polymorphisms in one-carbon metabolic pathway. Childs Nerv. Syst. 2018, 34, 277–284. [Google Scholar] [CrossRef]
- Imani, M.M.; Lopez-Jornet, P.; López, E.P.; Ghanbari, F.; Sadeghi, M. Association of Betaine-Homocysteine S-Methyl Transferase (rs3797546 and rs3733890) polymorphisms with non-syndromic cleft lip/palate: A meta-analysis. Int. Orthod. 2019, 17, 643–651. [Google Scholar] [CrossRef]
- Kalisch-Smith, J.I.; Ved, N.; Sparrow, D.B. Environmental Risk Factors for Congenital Heart Disease. Cold Spring Harb. Perspect. Biol. 2020, 12, a037234. [Google Scholar] [CrossRef]
- Malik, R.A.; Lone, M.R.; Ahmed, A.; Koul, K.A.; Malla, R.R. Maternal hyperhomocysteinemia and congenital heart defects: A prospective case control study in Indian population. Indian Heart J. 2017, 69, 17–19. [Google Scholar] [CrossRef]
- Mei, X.; Qi, D.; Zhang, T.; Zhao, Y.; Jin, L.; Hou, J.; Wang, J.; Lin, Y.; Xue, Y.; Zhu, P.; et al. Inhibiting MARSs reduces hyperhomocysteinemia-associated neural tube and congenital heart defects. EMBO Mol. Med. 2020, 12, e9469. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Pandey, G. Biofortification of pulses and legumes to enhance nutrition. Heliyon 2020, 6, e03682. [Google Scholar] [CrossRef] [PubMed]
- Wiedeman, A.M.; Barr, S.I.; Green, T.J.; Xu, Z.; Innis, S.M.; Kitts, D.D. Dietary Choline Intake: Current State of Knowledge Across the Life Cycle. Nutrients 2018, 10, 1513. [Google Scholar] [CrossRef]
- Millian, N.S.; Garrow, T.A. Human betaine-homocysteine methyltransferase is a zinc metalloenzyme. Arch. Biochem. Biophys. 1998, 356, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Cuny, H.; Rapadas, M.; Gereis, J.; Martin, E.M.; Kirk, R.B.; Shi, H.; Dunwoodie, S.L. NAD deficiency due to environmental factors or gene-environment interactions causes congenital malformations and miscarriage in mice. Proc. Natl. Acad. Sci. USA 2020, 117, 3738–3747. [Google Scholar] [CrossRef]
- Gibson, G.; Berger, K. Dietary modification, penetrance, and the origins of congenital malformation. Proc. Natl. Acad. Sci. USA 2020, 117, 5097–5099. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Baseline Characteristics | Control Group | Case Group | χ2 | p |
|---|---|---|---|---|
| (n = 740) | (n = 426) | |||
| Child-bearing age (years) | 0.912 | 0.340 | ||
| <35 | 635(85.8%) | 374(87.8%) | ||
| ≥35 | 105(14.2%) | 52(12.2%) | ||
| Pre-pregnancy BMI a | ||||
| <18.5 | 192(25.9%) | 77(18.1%) | 11.810 | 0.008 |
| 18.5–23.9 | 406(54.9%) | 274(64.3%) | ||
| 24–26.9 | 91(12.3%) | 47(11.0%) | ||
| ≥27 | 51(6.9%) | 28(6.6%) | ||
| Education level | 187.573 | <0.001 | ||
| Less than primary or primary | 9(1.2%) | 43(10.1%) | ||
| Junior high school | 144(19.5%) | 195(45.8%) | ||
| High school or Technical secondary school | 246(33.2%) | 123(28.9%) | ||
| College or above | 341(46.1%) | 65(15.3%) | ||
| Consanguineous marriages | 13.989 | <0.001 | ||
| No | 737(99.6%) | 413(96.9%) | ||
| Yes | 3(0.4%) | 13(3.1%) | ||
| Gestational diabetes mellitus | 34.302 | <0.001 | ||
| No | 717(96.9%) | 376(88.3%) | ||
| Yes | 23(3.1%) | 50(11.7%) | ||
| Gestational hypertension | 23.594 | <0.001 | ||
| No | 723(97.7%) | 390(91.5%) | ||
| Yes | 17(2.3%) | 36(8.5%) | ||
| Abnormal pregnancy history pregnancy | 9.363 | 0.002 | ||
| No | 411(55.5%) | 197(46.2%) | ||
| Yes | 329(44.5%) | 229(53.8%) | ||
| Congenital malformations in family members | 19.837 | <0.001 | ||
| No | 733(99.1%) | 404(94.8%) | ||
| Yes | 7(0.9%) | 22(5.2%) | ||
| Exposure to environmental pollutants | 43.687 | <0.001 | ||
| No | 687(92.8%) | 340(79.8%) | ||
| Yes | 53(7.2%) | 86(20.2%) | ||
| Antibiotic use in early pregnancy | 7.234 | 0.007 | ||
| No | 729(98.5%) | 409(96.0%) | ||
| Yes | 11(1.5%) | 17(4.0%) | ||
| Tobacco exposure in early pregnancy | 78.692 | <0.001 | ||
| No | 602(81.4%) | 244(57.3%) | ||
| Yes | 138(18.6%) | 182(42.7%) | ||
| Alcohol exposure in early pregnancy | 9.461 | 0.002 | ||
| No | 712(96.2%) | 392(92.0%) | ||
| Yes | 28(3.8%) | 34(8.0%) | ||
| Periconceptional folate use | 7.026 | 0.008 | ||
| Yes | 687(92.8%) | 376(88.3%) | ||
| No | 53(7.2%) | 50(11.7%) |
| Maternal Dietary Habits | Control Group | Case Group | Univariate Logistic Regression | Multivariable Logistic Regression a Regression | ||
|---|---|---|---|---|---|---|
| (n = 740) | (n = 426) | Cor (95%CI) | p | aOR (95%CI) | p | |
| Smoked foods | 1.81(1.48-2.21) | <0.001 | 2.44(1.89–3.13) | <0.001 | ||
| Hardly b | 407(55.0%) | 172(40.4%) | 1 | 1 | ||
| Sometimes c | 310(41.9%) | 213(50.0%) | 1.63(1.27–2.09) | <0.001 | 2.14(1.57–2.91) | <0.001 |
| Often d | 23(3.1%) | 41(9.6%) | 4.22(2.46–7.24) | <0.001 | 7.98(4.16–15.32) | <0.001 |
| Barbecued foods | 1.94(1.53–2.47) | <0.001 | 1.86(1.39–2.48) | <0.001 | ||
| Hardly | 558(75.4%) | 260(61.0%) | 1 | 1 | ||
| Sometimes | 177(23.9%) | 153(35.9%) | 1.86(1.43–2.41) | <0.001 | 1.89(1.37–2.60) | <0.001 |
| Often | 5(0.7%) | 13(3.1%) | 5.58(1.97–15.82) | 0.001 | 3.01(0.90–10.07) | 0.073 |
| Fried foods | 1.55(1.27–1.89) | <0.001 | 1.93(1.51–2.46) | <0.001 | ||
| Hardly | 458(61.9%) | 214(50.2%) | 1 | 1 | ||
| Sometimes | 253(34.2%) | 177(41.5%) | 1.50(1.16–1.92) | 0.002 | 2.15(1.57–2.94) | <0.001 |
| Often | 29(3.9%) | 35(8.2%) | 2.58(1.54–4.34) | <0.001 | 3.02(1.62–5.60) | <0.001 |
| Pickled vegetables | 1.87(1.51–2.32) | <0.001 | 2.50(1.92–3.25) | <0.001 | ||
| Hardly | 448(60.5%) | 184(43.2%) | 1 | 1 | ||
| Sometimes | 274(37.0%) | 220(51.6%) | 1.96(1.53–2.50) | <0.001 | 2.58(1.90–3.52) | <0.001 |
| Often | 18(2.4%) | 22(5.2%) | 2.98(1.56–5.68) | 0.001 | 5.53(2.58–11.82) | <0.001 |
| Fresh vegetables | 0.89(0.52–1.52) | 0.664 | 0.86(0.46–1.57) | 0.615 | ||
| Hardly | 3(0.4%) | 3(0.7%) | 1 | 1 | ||
| Sometimes | 21(2.8%) | 12(2.8%) | 0.57(0.10–3.29) | 0.531 | 0.17(0.02–1.11) | 0.064 |
| Often | 716(96.8%) | 411(96.5%) | 0.57(0.12–2.86) | 0.498 | 0.24(0.04–1.27) | 0.093 |
| Fresh fruits | 0.37(0.30–0.47) | <0.001 | 0.47(0.36–0.62) | <0.001 | ||
| Hardly | 14(1.9%) | 81(19.0%) | 1 | 1 | ||
| Sometimes | 41(5.5%) | 16(3.8%) | 0.07(0.03–0.15) | <0.001 | 0.06(0.03–0.16) | <0.001 |
| Often | 685(92.6%) | 329(77.2%) | 0.08(0.05–0.15) | <0.001 | 0.12(0.06–0.24) | <0.001 |
| Fresh meat | 0.81(0.61–1.08) | 0.155 | 1.08(0.77–1.54) | 0.644 | ||
| Hardly | 21(2.8%) | 12(2.8%) | 1 | 1 | ||
| Sometimes | 38(5.1%) | 37(8.7%) | 1.70(0.74–3.95) | 0.214 | 1.41(0.52–3.84) | 0.498 |
| Often | 681(92.0%) | 377(88.5%) | 0.97(0.47–1.99) | 0.931 | 1.34(0.58–3.08) | 0.493 |
| Fish and shrimp | 0.27(0.22–0.33) | <0.001 | 0.35(0.28–0.44) | <0.001 | ||
| Hardly | 29(3.9%) | 91(21.4%) | 1 | 1 | ||
| Sometimes | 207(28.0%) | 210(49.3%) | 0.32(0.20–0.51) | <0.001 | 0.33(0.20–0.56) | <0.001 |
| Often | 504(68.1%) | 125(29.3%) | 0.08(0.05–0.12) | <0.001 | 0.12(0.07–0.20) | <0.001 |
| Fresh eggs | 0.40(0.33–0.49) | <0.001 | 0.56(0.45–0.71) | <0.001 | ||
| Hardly | 36(4.9%) | 58(13.6%) | 1 | 1 | ||
| Sometimes | 86(11.6%) | 127(29.8%) | 0.92(0.56–1.51) | 0.732 | 0.76(0.42–1.37) | 0.355 |
| Often | 618(83.5%) | 241(56.6%) | 0.24(0.16–0.38) | <0.001 | 0.37(0.21–0.63) | <0.001 |
| Beans | 0.52(0.44–0.61) | <0.001 | 0.68(0.56–0.83) | <0.001 | ||
| Hardly | 107(14.5%) | 107(25.1%) | 1 | 1 | ||
| Sometimes | 216(29.2%) | 192(45.1%) | 0.89(0.64–1.24) | 0.486 | 1.13(0.76–1.69) | 0.544 |
| Often | 417(56.4%) | 127(29.8%) | 0.30(0.22–0.42) | <0.001 | 0.52(0.35–0.79) | 0.002 |
| Milk products | 0.51(0.44–0.59) | <0.001 | 0.67(0.56–0.80) | <0.001 | ||
| Hardly | 143(19.3%) | 173(40.6%) | 1 | 1 | ||
| Sometimes | 150(20.3%) | 109(25.6%) | 0.60(0.43–0.84) | 0.003 | 0.88(0.59–1.31) | 0.533 |
| Often | 447(60.4%) | 144(33.8%) | 0.27(0.20–0.36) | <0.001 | 0.46(0.32–0.65) | <0.001 |
| SNPs | Location | Major Allele | Minor Allele | MAF | Group | Genotype Frequencies a | χ2 | p | ||
|---|---|---|---|---|---|---|---|---|---|---|
| AA | AB | BB | ||||||||
| rs3733890 | Chr5: 79126136 | G | A | 0.3250 | control | 333(45.0%) | 333(45.0%) | 74(10.0%) | 0.4865 | 0.4855 |
| case | 162(38.0%) | 216(50.7%) | 48(11.3%) | |||||||
| rs1316753 | Chr5: 79235514 | C | G | 0.4338 | control | 248(33.5%) | 342(46.2%) | 150(20.3%) | 2.5913 | 0.1075 |
| case | 95(22.3%) | 247(58.0%) | 84(19.7%) | |||||||
| rs567754 | Chr5: 79120593 | C | T | 0.4628 | control | 203(27.4%) | 389(52.6%) | 148(20.0%) | 2.4204 | 0.1198 |
| case | 132(31.0%) | 227(53.3%) | 67(15.7%) | |||||||
| rs1915706 | Chr5: 79140388 | T | C | 0.2257 | control | 442(59.7%) | 262(35.4%) | 36(4.9%) | 0.1261 | 0.7225 |
| case | 223(52.3%) | 176(41.3%) | 27(6.3%) | |||||||
| SNPs | Univariate Logistic Reregression | Multivariate Logistic Regression a | |||
|---|---|---|---|---|---|
| cOR (95%CI) | p | aOR (95%CI) | p | FDR_P | |
| rs3733890 | |||||
| GG | 1 | 1 | |||
| GA | 1.33(1.03–1.72) | 0.026 | 1.28(0.94–1.73) | 0.118 | 0.189 |
| AA | 1.33(0.89–2.01) | 0.168 | 1.03(0.61–1.74) | 0.918 | 0.918 |
| Dominant model b | 1.33(1.04–1.70) | 0.021 | 1.23(0.92–1.65) | 0.163 | 0.217 |
| Recessive model c | 1.14(0.78–1.68) | 0.496 | 0.90(0.55–1.48) | 0.681 | 0.904 |
| Additive model d | 1.21(1.01–1.45) | 0.038 | 1.11(0.88–1.39) | 0.373 | 0.373 |
| rs1316753 | |||||
| CC | 1 | 1 | |||
| CG | 1.88(1.41–2.51) | <0.001 | 2.01(1.43–2.83) | <0.001 | <0.001 |
| GG | 1.46(1.02–2.09) | 0.037 | 1.55(1.00–2.40) | 0.048 | 0.096 |
| Dominant model | 1.76(1.34–2.31) | <0.001 | 1.88(1.36–2.61) | <0.001 | <0.001 |
| Recessive model | 0.97(0.72–1.30) | 0.821 | 0.98(0.68–1.41) | 0.904 | 0.904 |
| Additive model | 1.25(1.05–1.48) | 0.012 | 1.30(1.06–1.60) | 0.014 | 0.028 |
| rs567754 | |||||
| CC | 1 | 1 | |||
| CT | 0.90(0.68–1.18) | 0.438 | 0.90(0.65–1.26) | 0.555 | 0.634 |
| TT | 0.70(0.48–1.00) | 0.050 | 0.78(0.51–1.19) | 0.249 | 0.332 |
| Dominant model | 0.84(0.65–1.09) | 0.197 | 0.87(0.64–1.20) | 0.393 | 0.393 |
| Recessive model | 0.75(0.54–1.02) | 0.071 | 0.83(0.57–1.20) | 0.323 | 0.646 |
| Additive model | 0.84(0.71–1.01) | 0.058 | 0.88(0.72–1.09) | 0.255 | 0.340 |
| rs1915706 | |||||
| TT | 1 | 1 | |||
| CT | 1.33(1.04–1.71) | 0.025 | 1.81(1.33–2.46) | <0.001 | <0.001 |
| CC | 1.49(0.88–2.51) | 0.138 | 2.05(1.10–3.82) | 0.023 | 0.061 |
| Dominant model | 1.35(1.06–1.72) | 0.014 | 1.84(1.37–2.48) | <0.001 | <0.001 |
| Recessive model | 1.32(0.79–2.21) | 0.285 | 1.60(0.88–2.94) | 0.124 | 0.496 |
| Additive model | 1.28(1.05–1.56) | 0.015 | 1.61(1.27–2.05) | <0.001 | <0.001 |
| Dietary Habits a | Interaction with rs1316753 b | Interaction with rs1915706 b | ||||
|---|---|---|---|---|---|---|
| aOR (95%CI) c | p | FDR_P | aOR (95%CI) c | p | FDR_P | |
| Smoked foods | 0.52 (0.26–1.01) | 0.055 | 0.165 | 0.62 (0.34–1.14) | 0.122 | 0.305 |
| Barbecued foods | 1.24 (0.62–2.48) | 0.548 | 0.616 | 1.33 (0.71–2.49) | 0.377 | 0.610 |
| Fried foods | 1.40 (0.72–2.71) | 0.316 | 0.406 | 1.19 (0.66–2.15) | 0.570 | 0.634 |
| Pickled vegetables | 0.48 (0.24–0.95) | 0.034 | 0.165 | 0.66 (0.36–1.19) | 0.170 | 0.340 |
| Fresh fruits | 0.30 (0.05–1.68) | 0.168 | 0.360 | 0.68 (0.18–2.58) | 0.571 | 0.634 |
| Fish and shrimp | 0.85 (0.29–2.53) | 0.776 | 0.776 | 0.66 (0.24–1.84) | 0.427 | 0.610 |
| Fresh eggs | 2.37 (0.64–8.83) | 0.200 | 0.360 | 0.39 (0.13–1.18) | 0.095 | 0.305 |
| Beans | 0.40 (0.17–0.95) | 0.038 | 0.165 | 0.33 (0.15–0.73) | 0.006 | 0.035 |
| Milk products | 0.66 (0.32–1.38) | 0.273 | 0.406 | 1.14 (0.60–2.19) | 0.687 | 0.687 |
| rs1915706 a | Maternal Beans Intake b | Cases | Controls | cOR(95%CI) | aOR(95%CI) c |
|---|---|---|---|---|---|
| - | - | 175 (41.1%) | 356 (48.1%) | 1 | 1 |
| - | + | 48 (11.3%) | 86 (11.6%) | 1.14 (0.76–1.69) | 0.88 (0.54–1.42) |
| + | - | 144 (33.8%) | 277 (37.4%) | 1.06 (0.81–1.39) | 1.52 (1.09–2.11) |
| + | + | 59 (13.8%) | 21 (2.8%) | 5.72 (3.36–9.71) | 4.00 (2.17–7.40) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, M.; Wang, T.; Huang, P.; Zhang, S.; Song, X.; Sun, M.; Liu, Y.; Wei, J.; Shu, J.; Zhong, T.; et al. Association and Interaction Effect of BHMT Gene Polymorphisms and Maternal Dietary Habits with Ventricular Septal Defect in Offspring. Nutrients 2022, 14, 3094. https://doi.org/10.3390/nu14153094
Luo M, Wang T, Huang P, Zhang S, Song X, Sun M, Liu Y, Wei J, Shu J, Zhong T, et al. Association and Interaction Effect of BHMT Gene Polymorphisms and Maternal Dietary Habits with Ventricular Septal Defect in Offspring. Nutrients. 2022; 14(15):3094. https://doi.org/10.3390/nu14153094
Chicago/Turabian StyleLuo, Manjun, Tingting Wang, Peng Huang, Senmao Zhang, Xinli Song, Mengting Sun, Yiping Liu, Jianhui Wei, Jing Shu, Taowei Zhong, and et al. 2022. "Association and Interaction Effect of BHMT Gene Polymorphisms and Maternal Dietary Habits with Ventricular Septal Defect in Offspring" Nutrients 14, no. 15: 3094. https://doi.org/10.3390/nu14153094
APA StyleLuo, M., Wang, T., Huang, P., Zhang, S., Song, X., Sun, M., Liu, Y., Wei, J., Shu, J., Zhong, T., Chen, Q., Zhu, P., & Qin, J. (2022). Association and Interaction Effect of BHMT Gene Polymorphisms and Maternal Dietary Habits with Ventricular Septal Defect in Offspring. Nutrients, 14(15), 3094. https://doi.org/10.3390/nu14153094