
The Microalgae Phaeodactylum tricornutum Is Well Suited as a Food with Positive Effects on the Intestinal Microbiota and the Generation of SCFA: Results from a Pre-Clinical Study

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Phaeodactylum Tricornutum Culture and Experimental Diets
2.2. Mouse Feeding Experiment
2.3. Histological Analyses
2.4. Measurement of Liver Fat Content
2.5. Fatty Acid Analyses in Mouse Tissues and Chow
2.6. Real-Time Quantitative Reverse Transcription PCR
2.7. Fecal Energy Loss and Fecal Energy Ratio
2.8. SCFA Analysis
2.9. Gut Microbiota Analysis
2.10. Statistical Analyses
3. Results
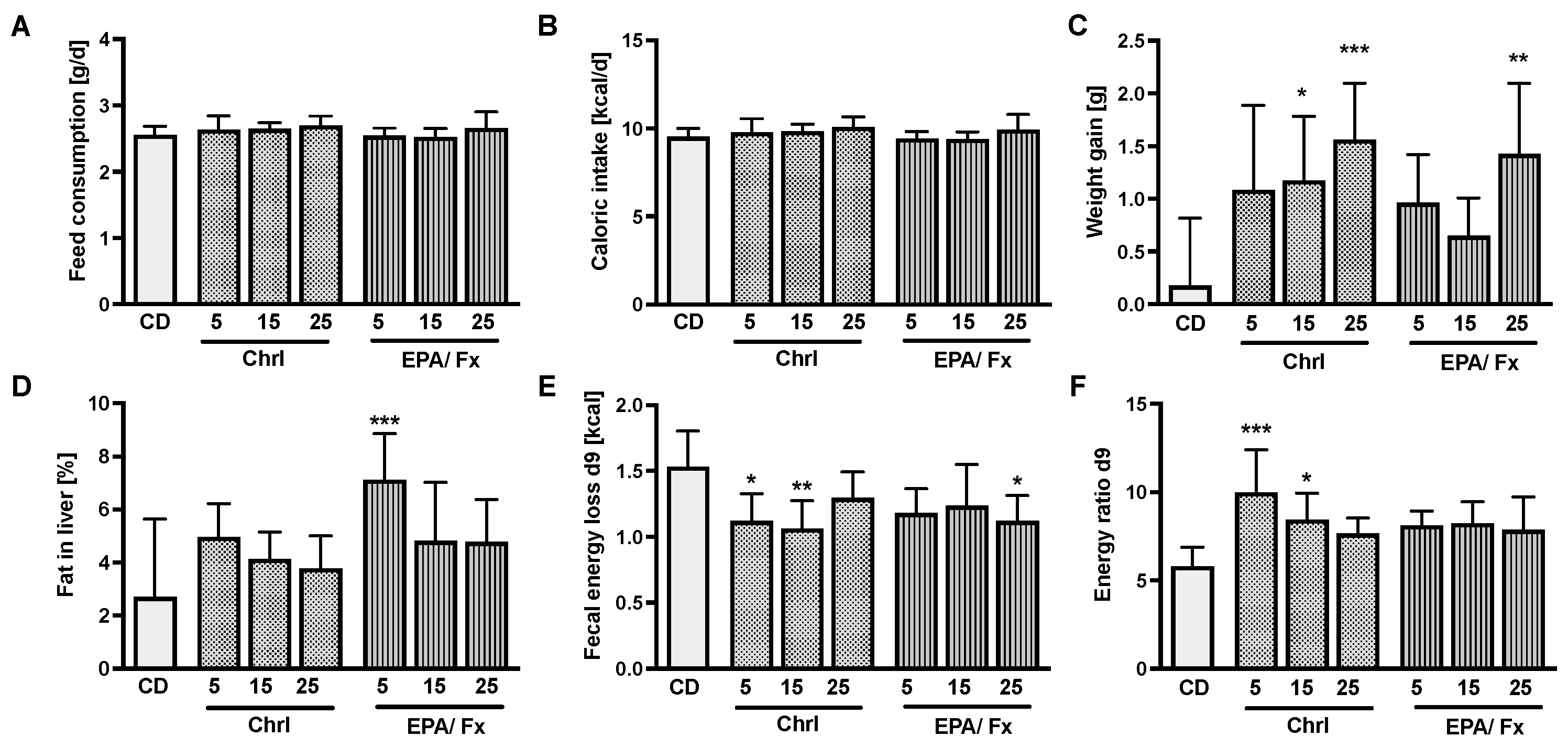
3.1. Microalgae Diet Acceptance and Effects on Body Weight (Bw) and Energy Uptake
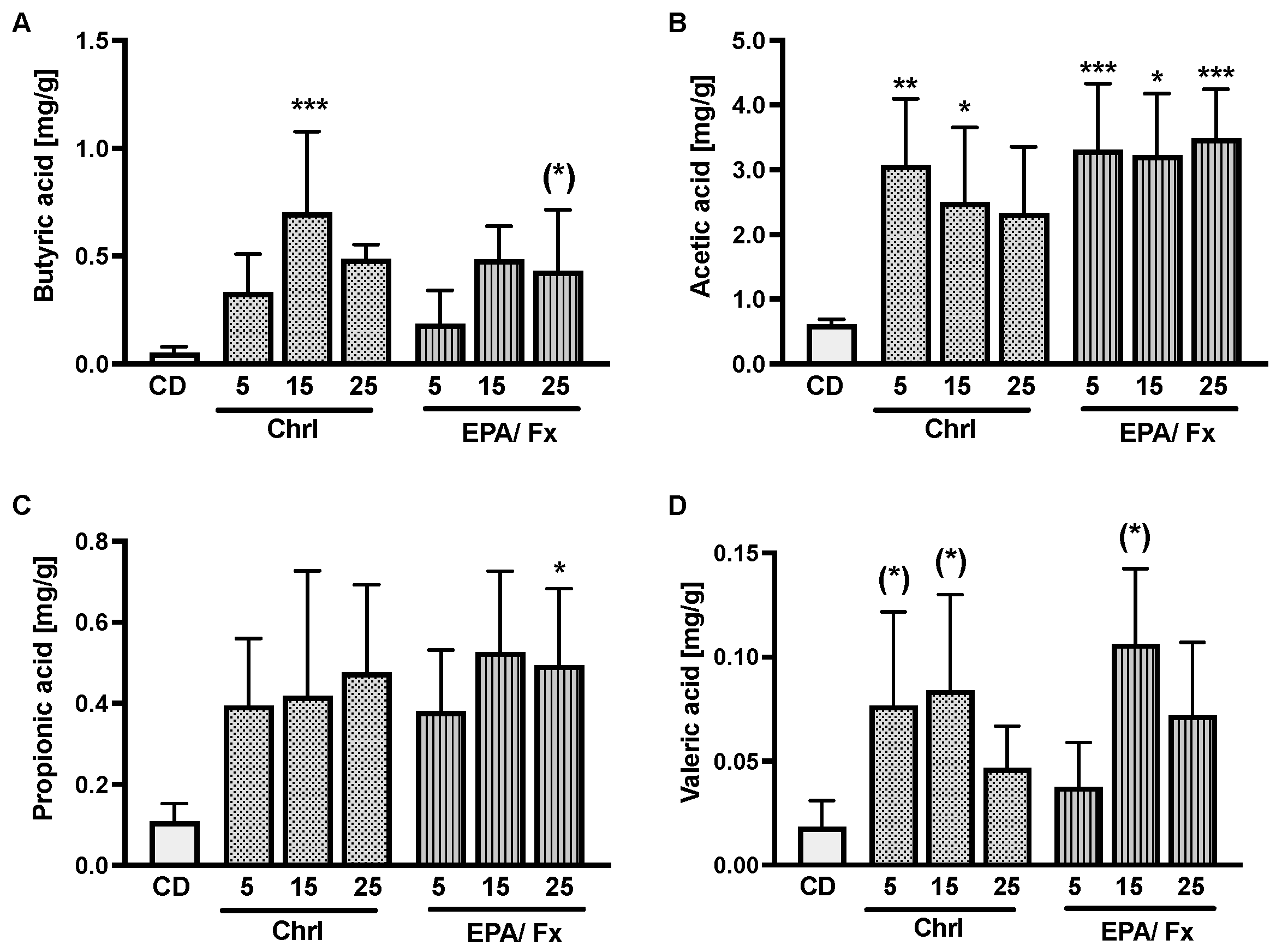
3.2. Short-Chain Fatty Acids, Markers of Intestinal Permeability, and Inflammation
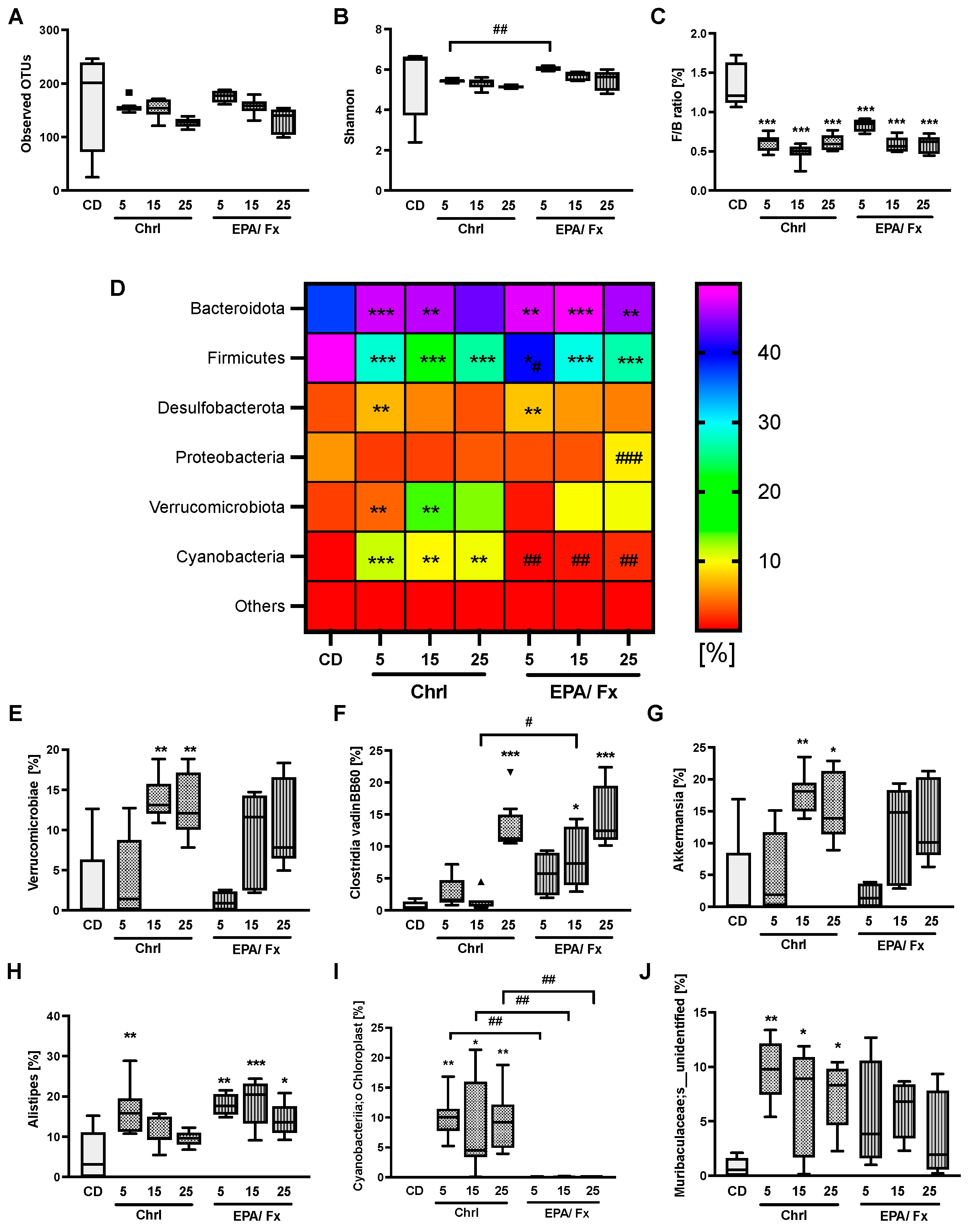
3.3. Microbiome Analysis in Feces
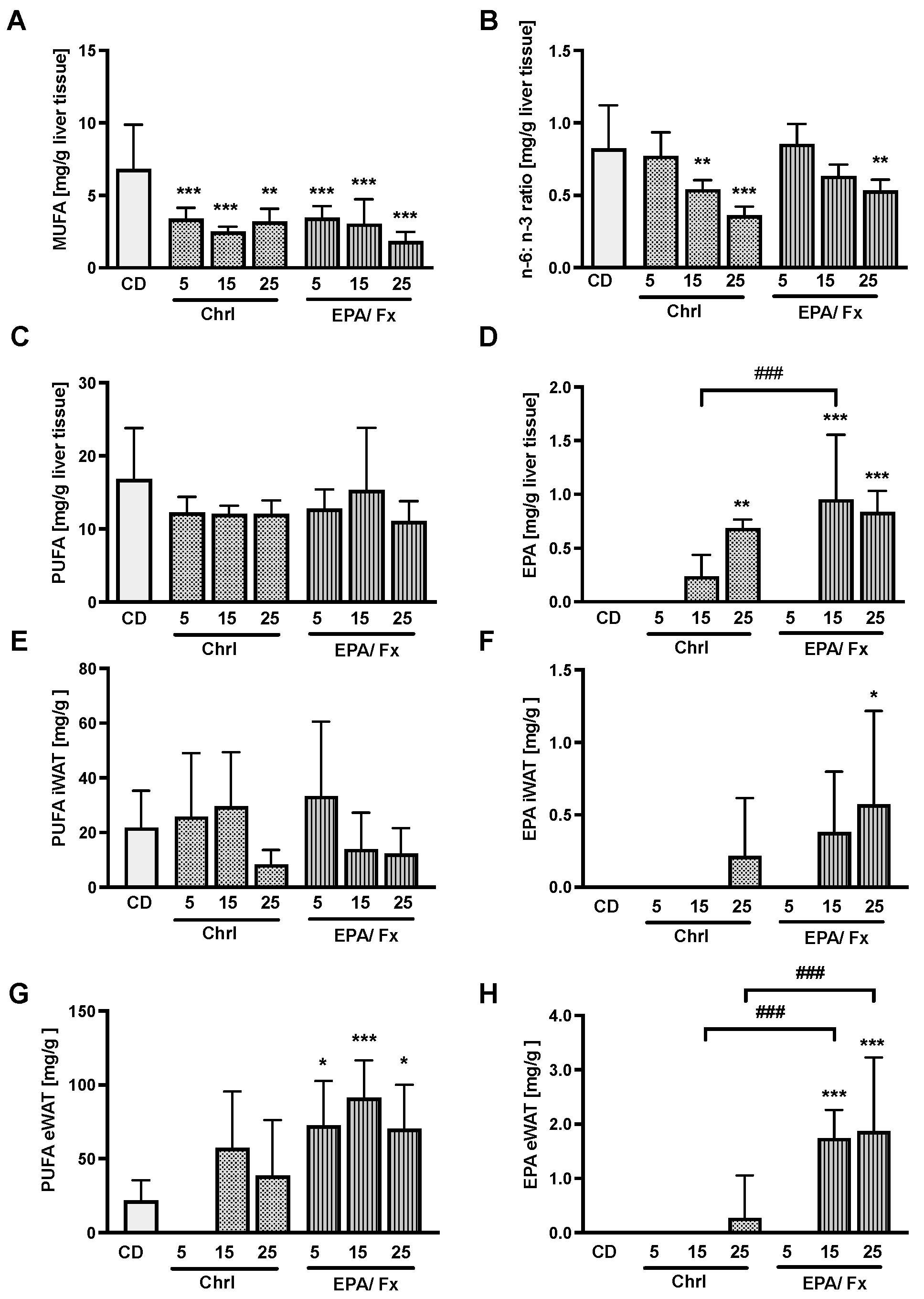
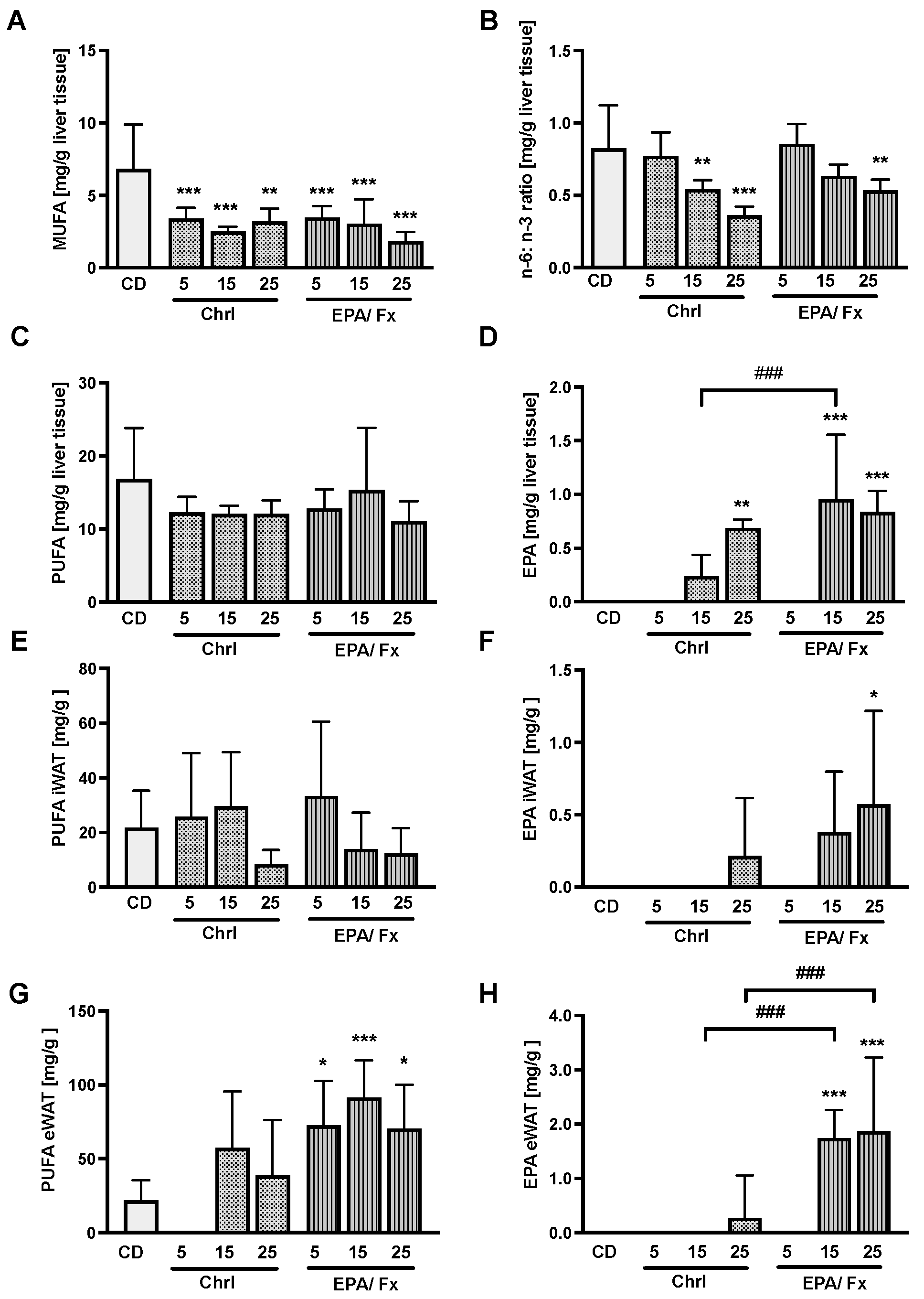
3.4. Fatty Acids Measured in the Liver and the White Adipose Tissue (WAT)
3.5. Correlations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Grosso, G.; Mateo, A.; Rangelov, N.; Buzeti, T.; Birt, C.; on behalf of the Food and Nutrition Section of the European Public Health Association. Nutrition in the Context of the Sustainable Development Goals. Eur. J. Public Health 2020, 30, i19–i23. [Google Scholar] [CrossRef]
- Barkia, I.; Saari, N.; Manning, S.R. Microalgae for High-Value Products Towards Human Health and Nutrition. Mar. Drugs 2019, 17, 304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, R.; Wei, D.; Xie, J. Diatoms as Cell Factories for High-Value Products: Chrysolaminarin, Eicosapentaenoic Acid, and Fucoxanthin. Crit. Rev. Biotechnol. 2020, 40, 993–1009. [Google Scholar] [CrossRef]
- Bertrand, M. Carotenoid Biosynthesis in Diatoms. Photosynth. Res. 2010, 106, 89–102. [Google Scholar] [CrossRef]
- Mikami, K.; Hosokawa, M. Biosynthetic Pathway and Health Benefits of Fucoxanthin, an Algae-Specific Xanthophyll in Brown Seaweeds. Int. J. Mol. Sci. 2013, 14, 13763–13781. [Google Scholar] [CrossRef] [Green Version]
- Gille, A.; Neumann, U.; Louis, S.; Bischoff, S.C.; Briviba, K. Microalgae as a Potential Source of Carotenoids: Comparative Results of an in Vitro Digestion Method and a Feeding Experiment with C57BL/6J Mice. J. Funct. Foods 2018, 49, 285–294. [Google Scholar] [CrossRef]
- Stiefvatter, L.; Lehnert, K.; Frick, K.; Montoya-Arroyo, A.; Frank, J.; Vetter, W.; Schmid-Staiger, U.; Bischoff, S.C. Oral Bioavailability of Omega-3 Fatty Acids and Carotenoids from the Microalgae Phaeodactylum Tricornutum in Healthy Young Adults. Mar. Drugs 2021, 19, 700. [Google Scholar] [CrossRef]
- Bae, M.; Kim, M.-B.; Park, Y.-K.; Lee, J.-Y. Health Benefits of Fucoxanthin in the Prevention of Chronic Diseases. Biochim. Biophys. Acta BBA Mol. Cell Biol. Lipids 2020, 1865, 158618. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, S.M.; Cha, K.H.; Mok, I.-K.; Koo, S.Y.; Pan, C.-H.; Lee, J.K. Evaluation of the Anti-Obesity Effect of the Microalga Phaeodactylum Tricornutum. Appl. Biol. Chem. 2016, 59, 283–290. [Google Scholar] [CrossRef]
- Peng, J.; Yuan, J.-P.; Wu, C.-F.; Wang, J.-H. Fucoxanthin, a Marine Carotenoid Present in Brown Seaweeds and Diatoms: Metabolism and Bioactivities Relevant to Human Health. Mar. Drugs 2011, 9, 1806–1828. [Google Scholar] [CrossRef]
- Gao, B.; Chen, A.; Zhang, W.; Li, A.; Zhang, C. Co-Production of Lipids, Eicosapentaenoic Acid, Fucoxanthin, and Chrysolaminarin by Phaeodactylum Tricornutum Cultured in a Flat-Plate Photobioreactor under Varying Nitrogen Conditions. J. Ocean Univ. China 2017, 16, 916–924. [Google Scholar] [CrossRef]
- Becker, W. 18 Microalgae in Human and Animal Nutrition. In Handbook of Microalgal Culture: Biotechnology and Applied Phycology; Wiley Online Library: Hoboken, NJ, USA, 2004; Volume 312. [Google Scholar]
- Burdge, G.C. Metabolism of α-Linolenic Acid in Humans. Prostaglandins Leukot. Essent. Fatty Acids 2006, 75, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Bresson, J.L.; Fairweather-Tait, S.; Flynn, A.; Golly, I.; Korhonen, H.; Lagiou, P.; Løvik, M.; Marchelli, R.; Martin, A.; Moseley, B.; et al. Scientific Opinion on Dietary Reference Values for Fats, Including Saturated Fatty Acids, Polyunsaturated Fatty Acids, Monounsaturated Fatty Acids, Trans Fatty Acids, and Cholesterol. EFSA J. 2010, 8, 1461. [Google Scholar] [CrossRef] [Green Version]
- Serhan, C.N. Novel Pro-Resolving Lipid Mediators in Inflammation Are Leads for Resolution Physiology. Nature 2014, 510, 92–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coll, M.; Libralato, S.; Tudela, S.; Palomera, I.; Pranovi, F. Ecosystem Overfishing in the Ocean. PLoS ONE 2008, 3, e3881. [Google Scholar] [CrossRef] [Green Version]
- European Commission EUR-Lex—32017R2470—Durchführungsverordnung (EU) 2017/2470 (2017). Available online: https://eur-lex.europa.eu/eli/reg_impl/2017/2470/oj/deu (accessed on 8 December 2021).
- Xia, S.; Gao, B.; Fu, J.; Xiong, J.; Zhang, C. Production of Fucoxanthin, Chrysolaminarin, and Eicosapentaenoic Acid by Odontella Aurita under Different Nitrogen Supply Regimes. J. Biosci. Bioeng. 2018, 126, 723–729. [Google Scholar] [CrossRef]
- Neumann, U.; Derwenskus, F.; Gille, A.; Louis, S.; Schmid-Staiger, U.; Briviba, K.; Bischoff, S. Bioavailability and Safety of Nutrients from the Microalgae Chlorella Vulgaris, Nannochloropsis Oceanica and Phaeodactylum Tricornutum in C57BL/6 Mice. Nutrients 2018, 10, 965. [Google Scholar] [CrossRef] [Green Version]
- Caballero, M.A.; Jallet, D.; Shi, L.; Rithner, C.; Zhang, Y.; Peers, G. Quantification of Chrysolaminarin from the Model Diatom Phaeodactylum Tricornutum. Algal Res. 2016, 20, 180–188. [Google Scholar] [CrossRef] [Green Version]
- Beattie, A.; Hirst, E.L.; Percival, E. Studies on the Metabolism of the Chrysophyceae. Comparative Structural Investigations on Leucosin (Chrysolaminarin) Separated from Diatoms and Laminarin from the Brown Algae. Biochem. J. 1961, 79, 531–537. [Google Scholar] [CrossRef]
- Yin, G.; Li, W.; Lin, Q.; Lin, X.; Lin, J.; Zhu, Q.; Jiang, H.; Huang, Z. Dietary Administration of Laminarin Improves the Growth Performance and Immune Responses in Epinephelus Coioides. Fish Shellfish Immunol. 2014, 41, 402–406. [Google Scholar] [CrossRef]
- Ciecierska, A.; Drywień, M.E.; Hamulka, J.; Sadkowski, T. Nutraceutical Functions of Beta-Glucans in Human Nutrition. Rocz. Państw. Zakładu Hig. 2019, 70, 315–324. [Google Scholar] [CrossRef]
- Zhu, F.; Du, B.; Xu, B. A Critical Review on Production and Industrial Applications of Beta-Glucans. Food Hydrocoll. 2016, 52, 275–288. [Google Scholar] [CrossRef]
- Jayachandran, M.; Chen, J.; Chung, S.S.M.; Xu, B. A Critical Review on the Impacts of β-Glucans on Gut Microbiota and Human Health. J. Nutr. Biochem. 2018, 61, 101–110. [Google Scholar] [CrossRef]
- Kadam, S.U.; Tiwari, B.K.; O’Donnell, C.P. Extraction, Structure and Biofunctional Activities of Laminarin from Brown Algae. Int. J. Food Sci. Technol. 2015, 50, 24–31. [Google Scholar] [CrossRef]
- Kusaikin, M.I.; Ermakova, S.P.; Shevchenko, N.M.; Isakov, V.V.; Gorshkov, A.G.; Vereshchagin, A.L.; Grachev, M.A.; Zvyagintseva, T.N. Structural Characteristics and Antitumor Activity of a New Chrysolaminaran from the Diatom Alga Synedra Acus. Chem. Nat. Compd. 2010, 46, 1–4. [Google Scholar] [CrossRef]
- Xia, S.; Gao, B.; Li, A.; Xiong, J.; Ao, Z.; Zhang, C. Preliminary Characterization, Antioxidant Properties and Production of Chrysolaminarin from Marine Diatom Odontella Aurita. Mar. Drugs 2014, 12, 4883–4897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carballo, C.; Chronopoulou, E.G.; Letsiou, S.; Maya, C.; Labrou, N.E.; Infante, C.; Power, D.M.; Manchado, M. Antioxidant Capacity and Immunomodulatory Effects of a Chrysolaminarin-Enriched Extract in Senegalese Sole. Fish Shellfish Immunol. 2018, 82, 1–8. [Google Scholar] [CrossRef]
- Reis, B.; Gonçalves, A.T.; Santos, P.; Sardinha, M.; Conceição, L.E.C.; Serradeiro, R.; Pérez, J.; Calduch, J.; Schmid, U.; Frick, K.; et al. Immune Status and Hepatic Antioxidant Capacity of Gilthead Seabream Sparus Aurata Juveniles Fed Yeast and Microalga Derived Β-Glucans. Mar. Drugs 2021, 21, 653. [Google Scholar] [CrossRef]
- Vijay, A.; Astbury, S.; Le Roy, C.; Spector, T.D.; Valdes, A.M. The Prebiotic Effects of Omega-3 Fatty Acid Supplementation: A Six-Week Randomised Intervention Trial. Gut Microbes 2021, 13, 1863133. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Zhao, H.; Liu, Z.; Sun, X.; Zhang, D.; Wang, S.; Xu, Y.; Zhang, G.; Wang, D. Modulation of Gut Microbiota by Fucoxanthin During Alleviation of Obesity in High-Fat Diet-Fed Mice. J. Agric. Food Chem. 2020, 68, 5118–5128. [Google Scholar] [CrossRef]
- Atanasov, J.; Schlörmann, W.; Trautvetter, U.; Glei, M. The Effects of β-Glucans on Intestinal Health. Ernahrungs Umsch. 2020, 67, 52–59. [Google Scholar] [CrossRef]
- Derwenskus, F.; Metz, F.; Gille, A.; Schmid-Staiger, U.; Briviba, K.; Schließmann, U.; Hirth, T. Pressurized Extraction of Unsaturated Fatty Acids and Carotenoids from Wet Chlorella Vulgaris and Phaeodactylum Tricornutum Biomass Using Subcritical Liquids. GCB Bioenergy 2019, 11, 335–344. [Google Scholar] [CrossRef]
- Gille, A.; Stojnic, B.; Derwenskus, F.; Trautmann, A.; Schmid-Staiger, U.; Posten, C.; Briviba, K.; Palou, A.; Bonet, M.L.; Ribot, J. A Lipophilic Fucoxanthin-Rich Phaeodactylum Tricornutum Extract Ameliorates Effects of Diet-Induced Obesity in C57BL/6J Mice. Nutrients 2019, 11, 796. [Google Scholar] [CrossRef] [Green Version]
- Neumann, U.; Louis, S.; Gille, A.; Derwenskus, F.; Schmid-Staiger, U.; Briviba, K.; Bischoff, S.C. Anti-Inflammatory Effects of Phaeodactylum Tricornutum Extracts on Human Blood Mononuclear Cells and Murine Macrophages. J. Appl. Phycol. 2018, 30, 2837–2846. [Google Scholar] [CrossRef]
- Zimmermann, J.; De Fazio, L.; Kaden-Volynets, V.; Hitzmann, B.; Bischoff, S.C. Consumption of Yeast-Fermented Wheat and Rye Breads Increases Colitis and Mortality in a Mouse Model of Colitis. Dig. Dis. Sci. 2022. [Google Scholar] [CrossRef] [PubMed]
- Drew, J.E.; Reichardt, N.; Williams, L.M.; Mayer, C.-D.; Walker, A.W.; Farquharson, A.J.; Kastora, S.; Farquharson, F.; Milligan, G.; Morrison, D.J.; et al. Dietary Fibers Inhibit Obesity in Mice, but Host Responses in the Cecum and Liver Appear Unrelated to Fiber-Specific Changes in Cecal Bacterial Taxonomic Composition. Sci. Rep. 2018, 8, 15566. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Chen, D.; Yu, B.; Luo, Y.; Zheng, P.; Mao, X.; Yu, J.; He, J. Effect of Different Dietary Non-Starch Fiber Fractions on Growth Performance, Nutrient Digestibility, and Intestinal Development in Weaned Pigs. Nutrition 2018, 51–52, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Hwang, P.-A.; Phan, N.N.; Lu, W.-J.; Ngoc Hieu, B.T.; Lin, Y.-C. Low-Molecular-Weight Fucoidan and High-Stability Fucoxanthin from Brown Seaweed Exert Prebiotics and Anti-Inflammatory Activities in Caco-2 Cells. Food Nutr. Res. 2016, 60, 32033. [Google Scholar] [CrossRef] [Green Version]
- Xiao, K.; Liu, C.; Qin, Q.; Zhang, Y.; Wang, X.; Zhang, J.; Odle, J.; Lin, X.; Hu, C.-A.A.; Liu, Y. EPA and DHA Attenuate Deoxynivalenol-Induced Intestinal Porcine Epithelial Cell Injury and Protect Barrier Function Integrity by Inhibiting Necroptosis Signaling Pathway. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2020, 34, 2483–2496. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, Q.; Wang, M.; Zhao, S.; Xu, G.; Li, J. N-3 Polyunsaturated Fatty Acids Prevent Disruption of Epithelial Barrier Function Induced by Proinflammatory Cytokines. Mol. Immunol. 2008, 45, 1356–1365. [Google Scholar] [CrossRef]
- EFSA. Scientific Opinion on the Substantiation of Health Claims Related to Undaria Pinnatifida (Harvey) Suringar and Maintenance or Achievement of a Normal Body Weight (ID 2345) Pursuant to Article 13(1) of Regulation (EC) No 1924/2006. EFSA J. 2009, 7, 1302. [Google Scholar] [CrossRef]
- EFSA. Scientific Opinion on the Safety of ‘Yeast Beta-Glucans’ as a Novel Food Ingredient. EFSA J. 2011, 9, 2137. [Google Scholar] [CrossRef]
- Babíček, K.; Čechová, I.; Simon, R.R.; Harwood, M.; Cox, D.J. Toxicological Assessment of a Particulate Yeast (1,3/1,6)-β-d-Glucan in Rats. Food Chem. Toxicol. 2007, 45, 1719–1730. [Google Scholar] [CrossRef] [PubMed]
- Commission Implementing Regulation (EU) 2020/1820 of 2 December 2020 Authorising the Placing on the Market of Dried Euglena Gracilis as a Novel Food. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/HTML/?uri=CELEX:32020R1820 (accessed on 21 April 2022).
- Hazards (BIOHAZ), E.P. on B.; Koutsoumanis, K.; Allende, A.; Alvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Hilbert, F.; et al. Update of the List of QPS-Recommended Biological Agents Intentionally Added to Food or Feed as Notified to EFSA 10: Suitability of Taxonomic Units Notified to EFSA until March 2019. EFSA J. 2019, 17, e05753. [Google Scholar] [CrossRef] [Green Version]
- Lage, S.; Ström, L.; Godhe, A.; Rydberg, S. Kinetics of β-N-Methylamino-L-Alanine (BMAA) and 2, 4-Diaminobutyric Acid (DAB) Production by Diatoms: The Effect of Nitrogen. Eur. J. Phycol. 2019, 54, 115–125. [Google Scholar] [CrossRef]
- Réveillon, D.; Séchet, V.; Hess, P.; Amzil, Z. Production of BMAA and DAB by Diatoms (Phaeodactylum Tricornutum, Chaetoceros Sp., Chaetoceros Calcitrans and, Thalassiosira Pseudonana) and Bacteria Isolated from a Diatom Culture. Harmful Algae 2016, 58, 45–50. [Google Scholar] [CrossRef]
- Van Onselen, R.; Downing, T.G. β-N-Methylamino-L-Alanine Inhibits Human Catalase Activity: Possible Implications for Neurodegenerative Disease Development. Int. J. Toxicol. 2019, 38, 129–134. [Google Scholar] [CrossRef]
- Salomonsson, M.L.; Fredriksson, E.; Alfjorden, A.; Hedeland, M.; Bondesson, U. Seafood Sold in Sweden Contains BMAA: A Study of Free and Total Concentrations with UHPLC–MS/MS and Dansyl Chloride Derivatization. Toxicol. Rep. 2015, 2, 1473–1481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lance, E.; Arnich, N.; Maignien, T.; Biré, R. Occurrence of β-N-Methylamino-l-Alanine (BMAA) and Isomers in Aquatic Environments and Aquatic Food Sources for Humans. Toxins 2018, 10, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sivaprakasam, S.; Prasad, P.D.; Singh, N. Benefits of Short-Chain Fatty Acids and Their Receptors in Inflammation and Carcinogenesis. Pharmacol. Ther. 2016, 164, 144–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bischoff, S.C. “Gut Health”: A New Objective in Medicine? BMC Med. 2011, 9, 24. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Nie, Q.; Xie, M.; Yao, H.; Zhang, K.; Yin, J.; Nie, S. Protective Effects of β-Glucan Isolated from Highland Barley on Ethanol-Induced Gastric Damage in Rats and Its Benefits to Mice Gut Conditions. Food Res. Int. 2019, 122, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Belobrajdic, D.P.; Jobling, S.A.; Morell, M.K.; Taketa, S.; Bird, A.R. Wholegrain Barley β-Glucan Fermentation Does Not Improve Glucose Tolerance in Rats Fed a High-Fat Diet. Nutr. Res. 2015, 35, 162–168. [Google Scholar] [CrossRef]
- Qu, X.; Nazarenko, Y.; Yang, W.; Nie, Y.; Zhang, Y.; Li, B. Effect of Oat β-Glucan on the Rheological Characteristics and Microstructure of Set-Type Yogurt. Molecules 2021, 26, 4752. [Google Scholar] [CrossRef]
- Rattigan, R.; Sweeney, T.; Maher, S.; Thornton, K.; Rajauria, G.; O’Doherty, J. Laminarin-Rich Extract Improves Growth Performance, Small Intestinal Morphology, Gene Expression of Nutrient Transporters and the Large Intestinal Microbial Composition of Piglets during the Critical Post-Weaning Period. Br. J. Nutr. 2019, 123, 255–263. [Google Scholar] [CrossRef]
- Rattigan, R.; O’Doherty, J.V.; Vigors, S.; Ryan, M.T.; Sebastiano, R.S.; Callanan, J.J.; Thornton, K.; Rajauria, G.; Margassery, L.M.; Dobson, A.D.W.; et al. The Effects of the Marine-Derived Polysaccharides Laminarin and Chitosan on Aspects of Colonic Health in Pigs Challenged with Dextran Sodium Sulphate. Mar. Drugs 2020, 18, 262. [Google Scholar] [CrossRef]
- Watson, H.; Mitra, S.; Croden, F.C.; Taylor, M.; Wood, H.M.; Perry, S.L.; Spencer, J.A.; Quirke, P.; Toogood, G.J.; Lawton, C.L.; et al. A Randomised Trial of the Effect of Omega-3 Polyunsaturated Fatty Acid Supplements on the Human Intestinal Microbiota. Gut 2018, 67, 1974–1983. [Google Scholar] [CrossRef]
- Zhu, L.; Sha, L.; Li, K.; Wang, Z.; Wang, T.; Li, Y.; Liu, P.; Dong, X.; Dong, Y.; Zhang, X.; et al. Dietary Flaxseed Oil Rich in Omega-3 Suppresses Severity of Type 2 Diabetes Mellitus via Anti-Inflammation and Modulating Gut Microbiota in Rats. Lipids Health Dis. 2020, 19, 20. [Google Scholar] [CrossRef] [Green Version]
- Costantini, L.; Molinari, R.; Farinon, B.; Merendino, N. Impact of Omega-3 Fatty Acids on the Gut Microbiota. Int. J. Mol. Sci. 2017, 18, 2645. [Google Scholar] [CrossRef] [Green Version]
- Smith, P.M.; Howitt, M.R.; Panikov, N.; Michaud, M.; Gallini, C.A.; Bohlooly, Y.M.; Glickman, J.N.; Garrett, W.S. The Microbial Metabolites, Short-Chain Fatty Acids, Regulate Colonic Treg Cell Homeostasis. Science 2013, 341, 569–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menni, C.; Zierer, J.; Pallister, T.; Jackson, M.A.; Long, T.; Mohney, R.P.; Steves, C.J.; Spector, T.D.; Valdes, A.M. Omega-3 Fatty Acids Correlate with Gut Microbiome Diversity and Production of N-Carbamylglutamate in Middle Aged and Elderly Women. Sci. Rep. 2017, 7, 11079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Ames, N.P.; Tun, H.M.; Tosh, S.M.; Jones, P.J.; Khafipour, E. High Molecular Weight Barley β-Glucan Alters Gut Microbiota Toward Reduced Cardiovascular Disease Risk. Front. Microbiol. 2016, 7, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, Y.; Zhu, L.; Li, Y.; Jiang, S.; Sun, Q.; Xie, E.; Chen, H.; Zhao, Z.; Qiao, W.; Xu, J.; et al. Structure of a Laminarin-Type β-(1→3)-Glucan from Brown Algae Sargassum Henslowianum and Its Potential on Regulating Gut Microbiota. Carbohydr. Polym. 2021, 255, 117389. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, S.G.; Kim, J.; Guevarra, R.B.; Lee, J.-H.; Kim, E.; Kim, S.; Unno, T. Laminarin Favorably Modulates Gut Microbiota in Mice Fed a High-Fat Diet. Food Funct. 2016, 7, 4193–4201. [Google Scholar] [CrossRef]
- Taylor, H.B.; Gudi, R.; Brown, R.; Vasu, C. Dynamics of Structural and Functional Changes in Gut Microbiota during Treatment with a Microalgal β-Glucan, Paramylon and the Impact on Gut Inflammation. Nutrients 2020, 12, 2193. [Google Scholar] [CrossRef]
- Aparicio, E.; Martín-Grau, C.; Bedmar, C.; Serrat Orus, N.; Basora, J.; Arija, V.; The ECLIPSES Study Group. Maternal Factors Associated with Levels of Fatty Acids, Specifically n-3 PUFA during Pregnancy: ECLIPSES Study. Nutrients 2021, 13, 317. [Google Scholar] [CrossRef]
- Caesar, R.; Tremaroli, V.; Kovatcheva-Datchary, P.; Cani, P.D.; Bäckhed, F. Crosstalk between Gut Microbiota and Dietary Lipids Aggravates WAT Inflammation through TLR Signaling. Cell Metab. 2015, 22, 658–668. [Google Scholar] [CrossRef] [Green Version]
- Cani, P.D.; Van Hul, M. Novel Opportunities for Next-Generation Probiotics Targeting Metabolic Syndrome. Curr. Opin. Biotechnol. 2015, 32, 21–27. [Google Scholar] [CrossRef]
- Sato, N. Are Cyanobacteria an Ancestor of Chloroplasts or Just One of the Gene Donors for Plants and Algae? Genes 2021, 12, 823. [Google Scholar] [CrossRef]
- Louis, P.; Flint, H.J. Formation of Propionate and Butyrate by the Human Colonic Microbiota. Environ. Microbiol. 2017, 19, 29–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, F.C.; Wasmund, K.; Cobankovic, I.; Jehmlich, N.; Herbold, C.W.; Lee, K.S.; Sziranyi, B.; Vesely, C.; Decker, T.; Stocker, R.; et al. Rational Design of a Microbial Consortium of Mucosal Sugar Utilizers Reduces Clostridiodes Difficile Colonization. Nat. Commun. 2020, 11, 5104. [Google Scholar] [CrossRef] [PubMed]
- Richards, P.; Fothergill, J.; Bernardeau, M.; Wigley, P. Development of the Caecal Microbiota in Three Broiler Breeds. Front. Vet. Sci. 2019, 6, 201. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Suppl | ME | SFA | UFA | PUFA | MUFA | EPA | n-3:n-6 | Fx | β-Carotin | Chrl |
|---|---|---|---|---|---|---|---|---|---|---|---|
| [%] | [MJ/kg] | [g/kg] | [g/kg] | [g/kg] | [g/kg] | [g/kg] | [g/kg] | [g/kg] | [g/kg] | ||
| Control diet | 15.6 | 9.33 | 11.18 | 3.60 | 7.58 | 0.00 | 0.08 | 0 | 0 | 0 | |
| PT_Chrl | 5 | 15.6 | 16.98 | 30.83 | 51.45 | 21.59 | 0.00 | 0.10 | 0.14 | 0.02 | 10.92 |
| 15 | 15.6 | 19.24 | 43.19 | 45.57 | 24.68 | 5.11 | 0.31 | 0.42 | 0.05 | 32.77 | |
| 25 | 15.6 | 19.11 | 49.58 | 27.50 | 24.41 | 8.32 | 0.90 | 0.71 | 0.09 | 54.62 | |
| PT_EPA/Fx | 5 | 15.6 | 20.49 | 33.81 | 68.14 | 24.534 | 3.32 | 0.16 | 0.57 | 0.07 | 0.66 |
| 15 | 15.6 | 22.44 | 52.96 | 72.56 | 26.68 | 11.14 | 0.30 | 1.71 | 0.22 | 1.98 | |
| 25 | 15.6 | 21.23 | 53.57 | 60.74 | 24.01 | 15.17 | 0.47 | 2.85 | 0.37 | 3.3 |
| Primer | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| Occludin | ACTCCTCCAATGGACAAGTG | CCCCACCTGTCGTGTAGTCT |
| ZO1 | CCACCTCTGTCCAGCTCTTC | CACCGGAGTGATGGTTTTCT |
| TNFα | ACCACCATCAAGGACTCA | AGGTCTGAAGGTAGGAAG |
| IL-1β | ACGGATTCCATGGTGAAGTC | GAGTGTGGATCCCAAGCAAT |
| IL-6 | AGTCACAGAAGGAGTGGCTA | CTGACCACAGTGAGGAATGT |
| Treatment | Suppl [%] | ZO1 | Occludin | TNF-α | IL-6 | IL-1β |
|---|---|---|---|---|---|---|
| CD | 29.94 ± 15 | 39.17 ± 27 | 10.06 ± 8.6 | 49.75 ± 82 | 5.850 ± 2.8 | |
| Chrl | 5 | 59.92 ± 30 | 46.56 ± 24 | 69.87 ± 28 ** | 148.8 ± 127 | 25.23 ± 15 |
| 15 | 25.67 ± 21 | 28.50 ± 18 | 22.58 ± 17 | 113.2 ± 135 | 14.18 ± 14 | |
| 25 | 25.31 ± 26 | 48.57 ± 40 | 118.5 ± 94 ** | 8.526 ± 8 | 29.60 ± 49 | |
| EPA/Fx | 5 | 24.15 ± 22 | 39.07 ± 26 | 32.88 ± 26 | 244.8± 344 | 18.73 ± 14 |
| 15 | 16.90 ± 8 | 27.18± 15 | 27.57± 18 | 93.87 ± 129 | 7.355 ± 5.3 | |
| 25 | 96.19 ± 161 | 75.82 ± 105 | 62.58± 40 * | 152.3 ± 99 | 32.19 ± 48 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stiefvatter, L.; Neumann, U.; Rings, A.; Frick, K.; Schmid-Staiger, U.; Bischoff, S.C. The Microalgae Phaeodactylum tricornutum Is Well Suited as a Food with Positive Effects on the Intestinal Microbiota and the Generation of SCFA: Results from a Pre-Clinical Study. Nutrients 2022, 14, 2504. https://doi.org/10.3390/nu14122504
Stiefvatter L, Neumann U, Rings A, Frick K, Schmid-Staiger U, Bischoff SC. The Microalgae Phaeodactylum tricornutum Is Well Suited as a Food with Positive Effects on the Intestinal Microbiota and the Generation of SCFA: Results from a Pre-Clinical Study. Nutrients. 2022; 14(12):2504. https://doi.org/10.3390/nu14122504
Chicago/Turabian StyleStiefvatter, Lena, Ulrike Neumann, Andreas Rings, Konstantin Frick, Ulrike Schmid-Staiger, and Stephan C. Bischoff. 2022. "The Microalgae Phaeodactylum tricornutum Is Well Suited as a Food with Positive Effects on the Intestinal Microbiota and the Generation of SCFA: Results from a Pre-Clinical Study" Nutrients 14, no. 12: 2504. https://doi.org/10.3390/nu14122504
APA StyleStiefvatter, L., Neumann, U., Rings, A., Frick, K., Schmid-Staiger, U., & Bischoff, S. C. (2022). The Microalgae Phaeodactylum tricornutum Is Well Suited as a Food with Positive Effects on the Intestinal Microbiota and the Generation of SCFA: Results from a Pre-Clinical Study. Nutrients, 14(12), 2504. https://doi.org/10.3390/nu14122504