
Gene Set Enrichment Analysis Reveals That Fucoidan Induces Type I IFN Pathways in BMDC

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
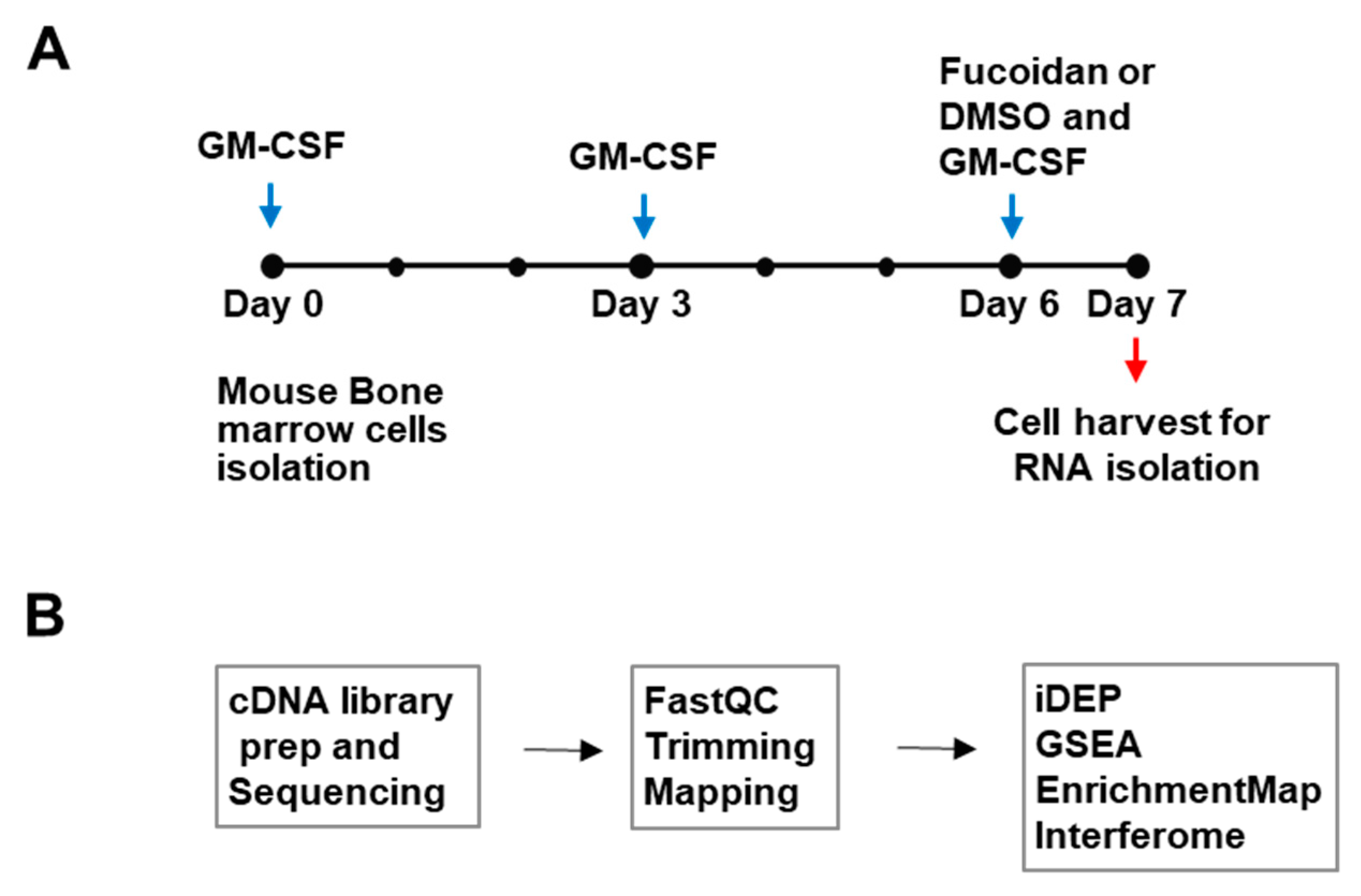
2.1. Bone Marrow-Derived Dendritic Cell (BMDC) Culture
2.2. Cell Staining and Flow Cytometry
2.3. RNA Sequencing
2.4. Bioinformatic Analyses
2.4.1. Analysis of Differentially Expressed Genes in IDEP
2.4.2. GSEA
2.4.3. Functional Enrichment Visualization and Annotation of GSEA Results
2.4.4. Interferome Analysis
2.5. Statistical Analysis
3. Results
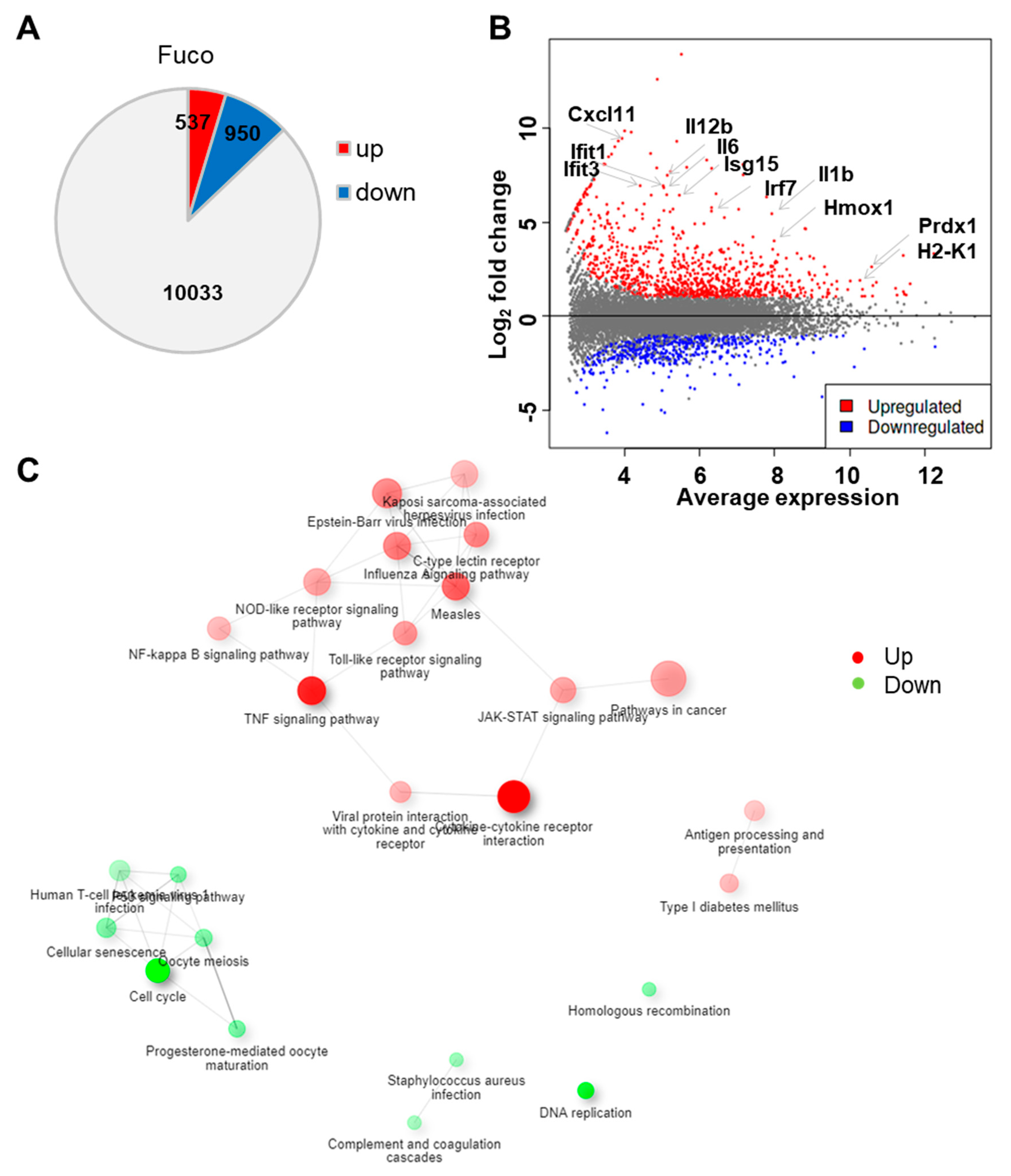
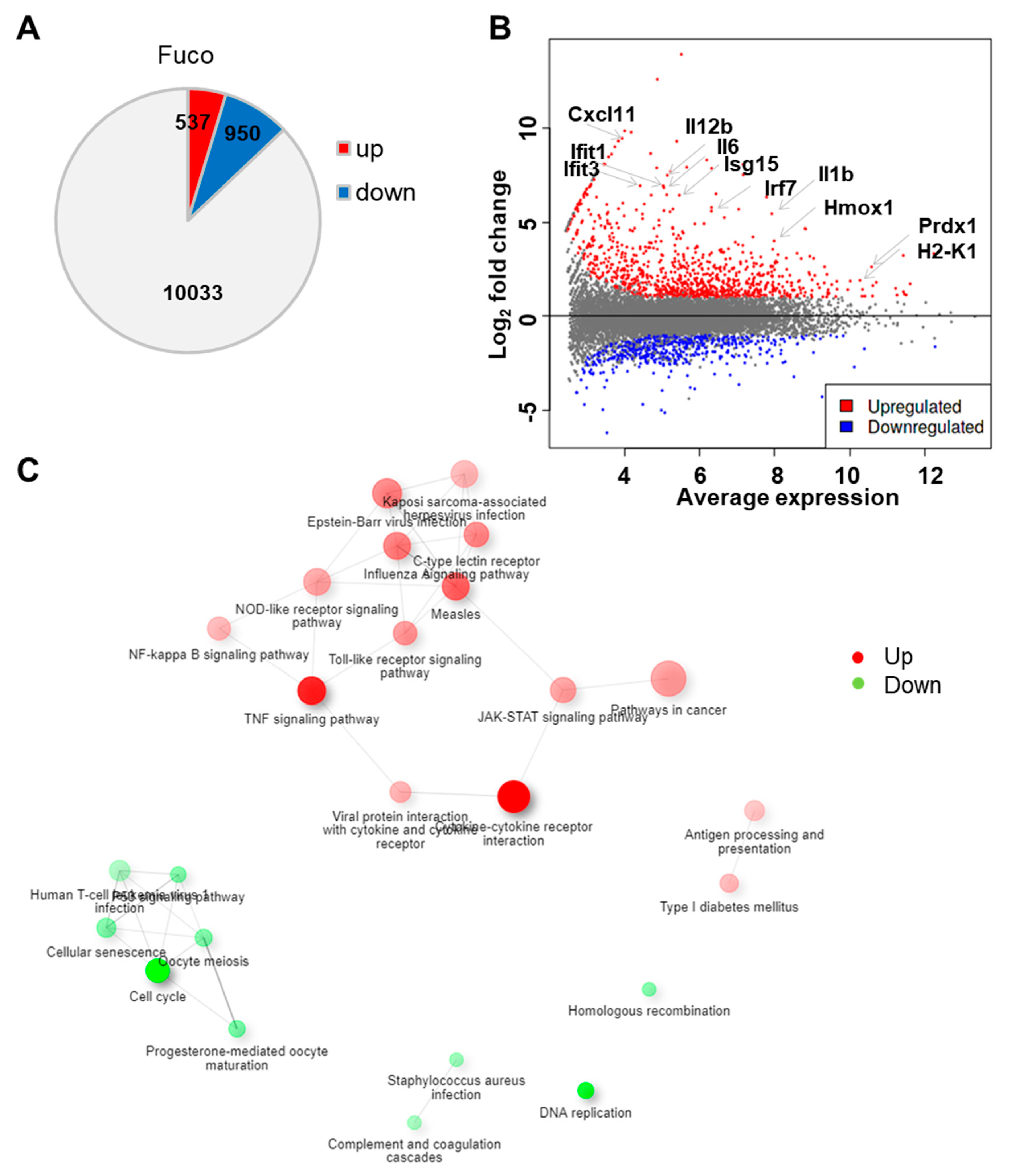
3.1. Fucoidan Treatment Upregulates Gene Sets Associated with Immune Responses to Exogenous Stimuli
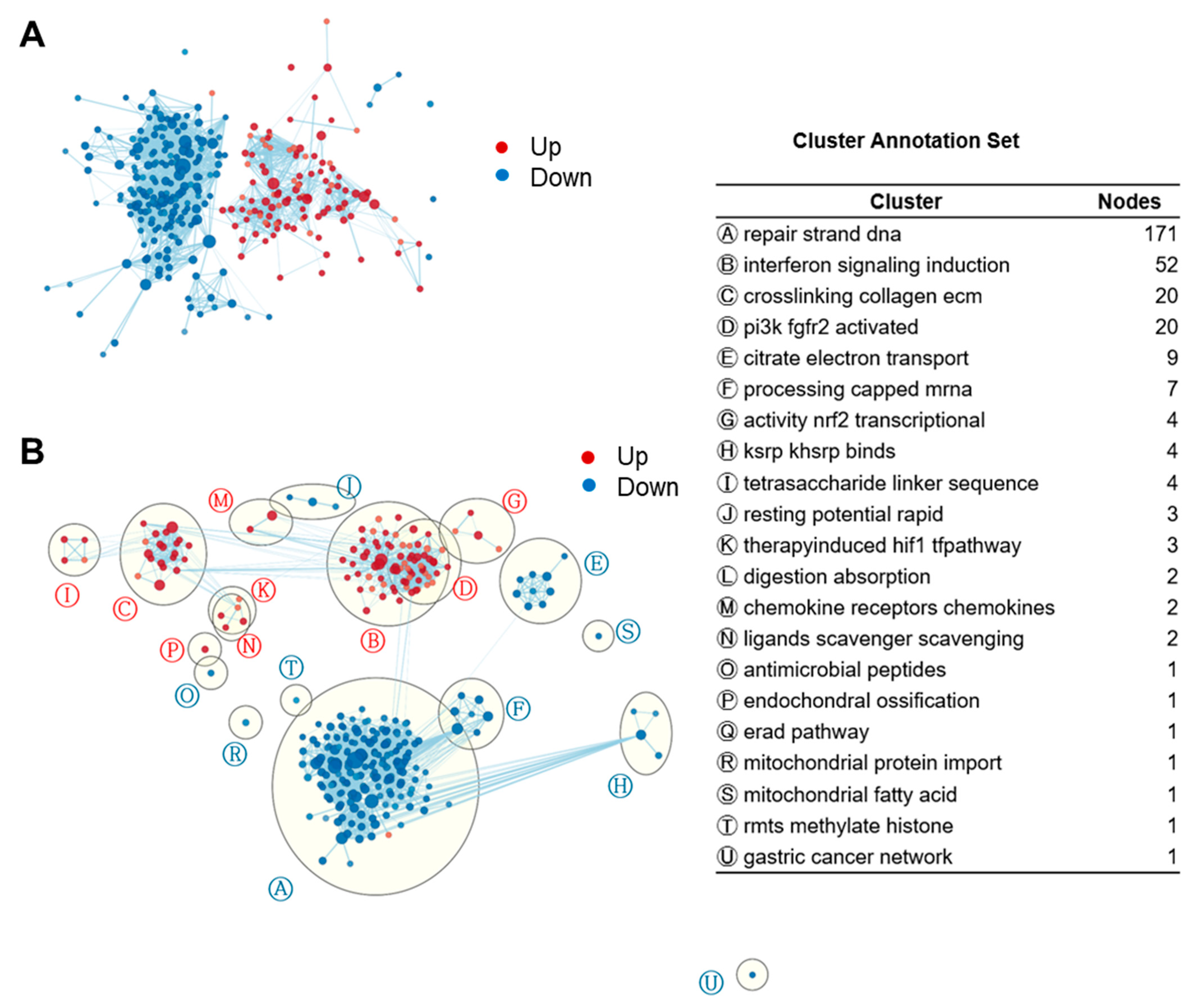
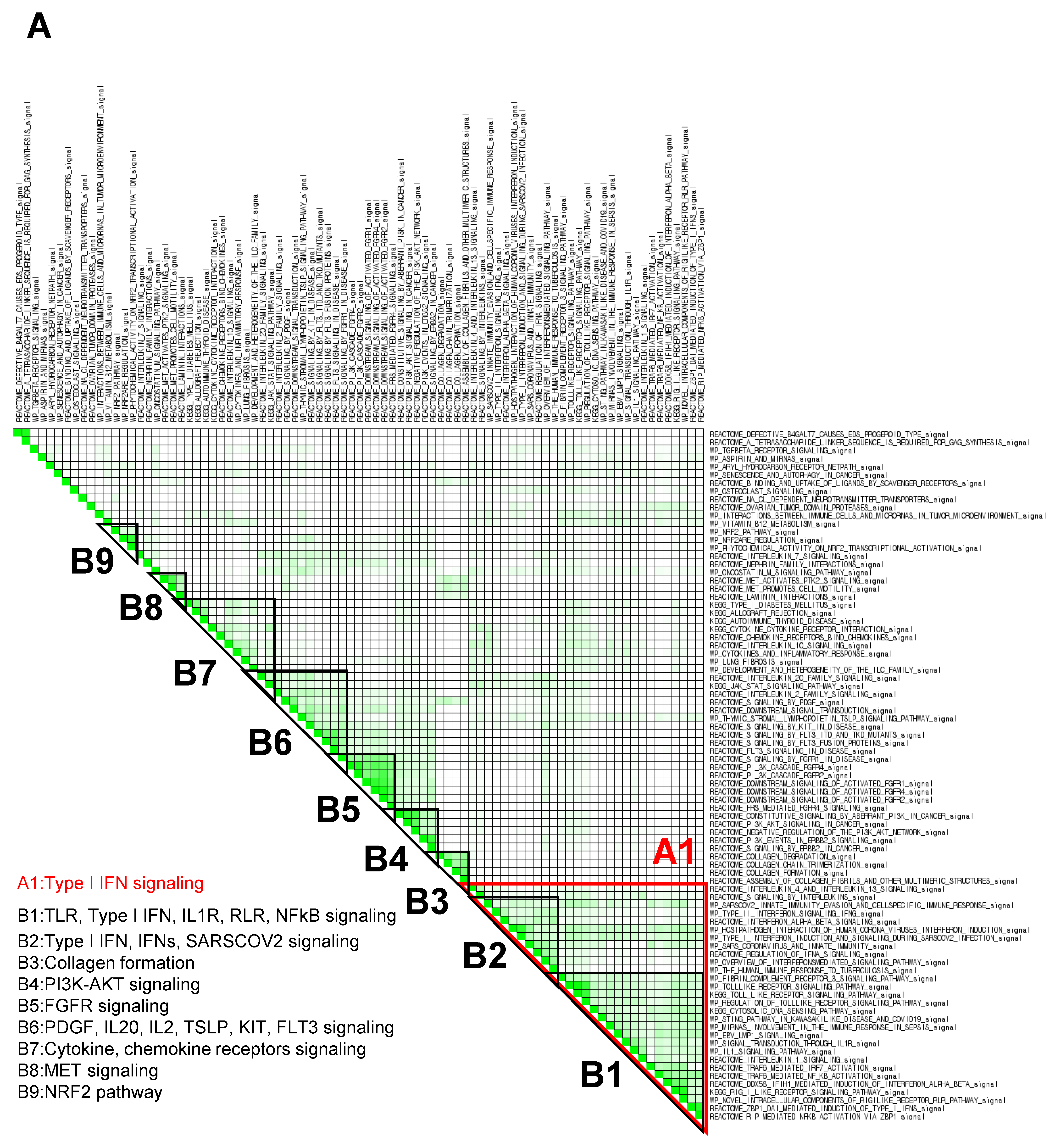
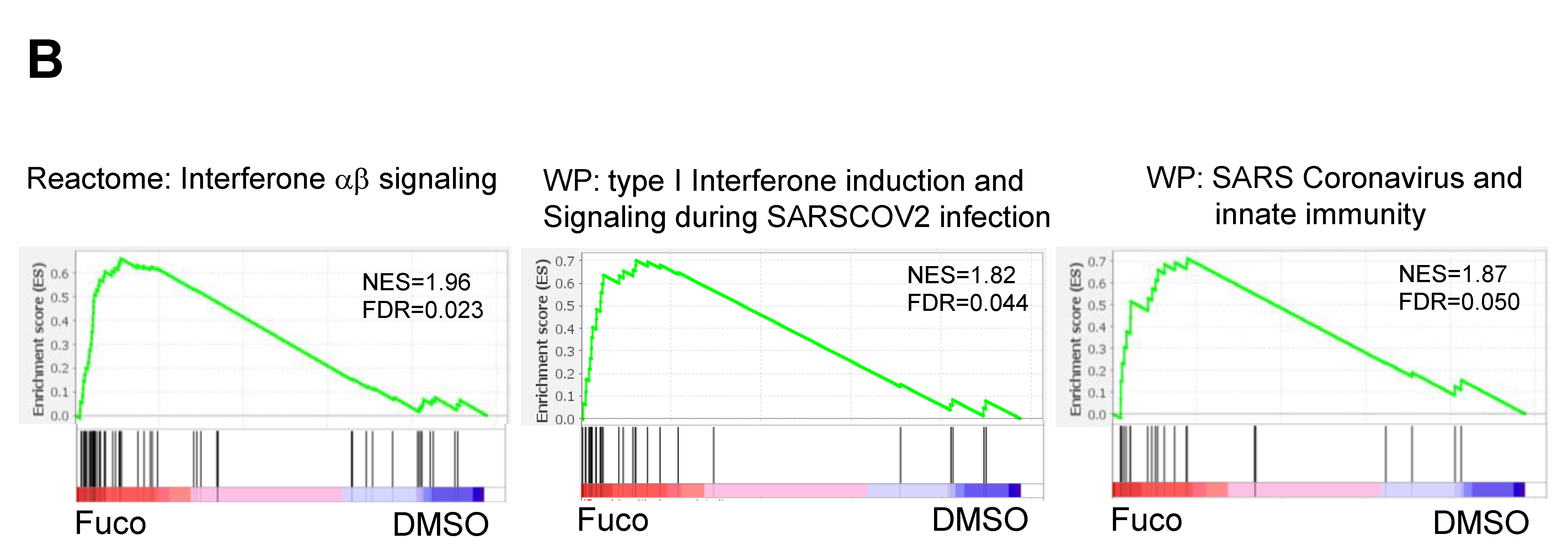
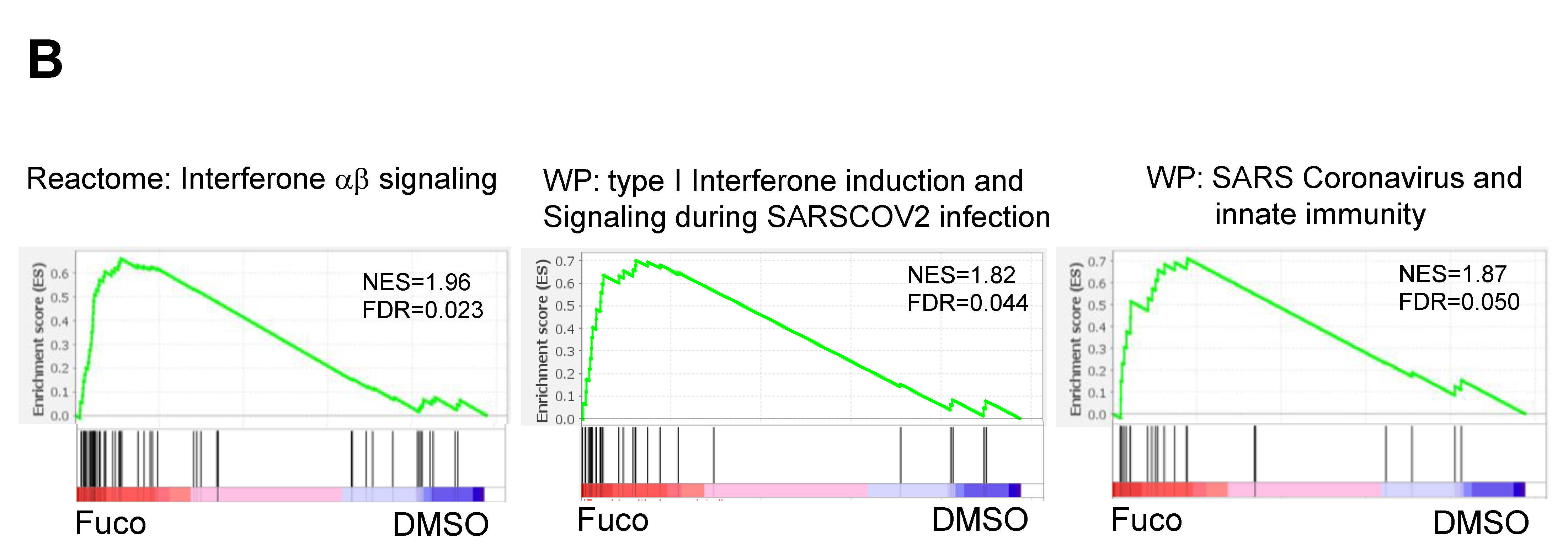
3.2. Gene Set Enrichment Analysis (GSEA) Showed That Fucoidan Strongly Induces Interferon Signaling Pathways in BMDCs
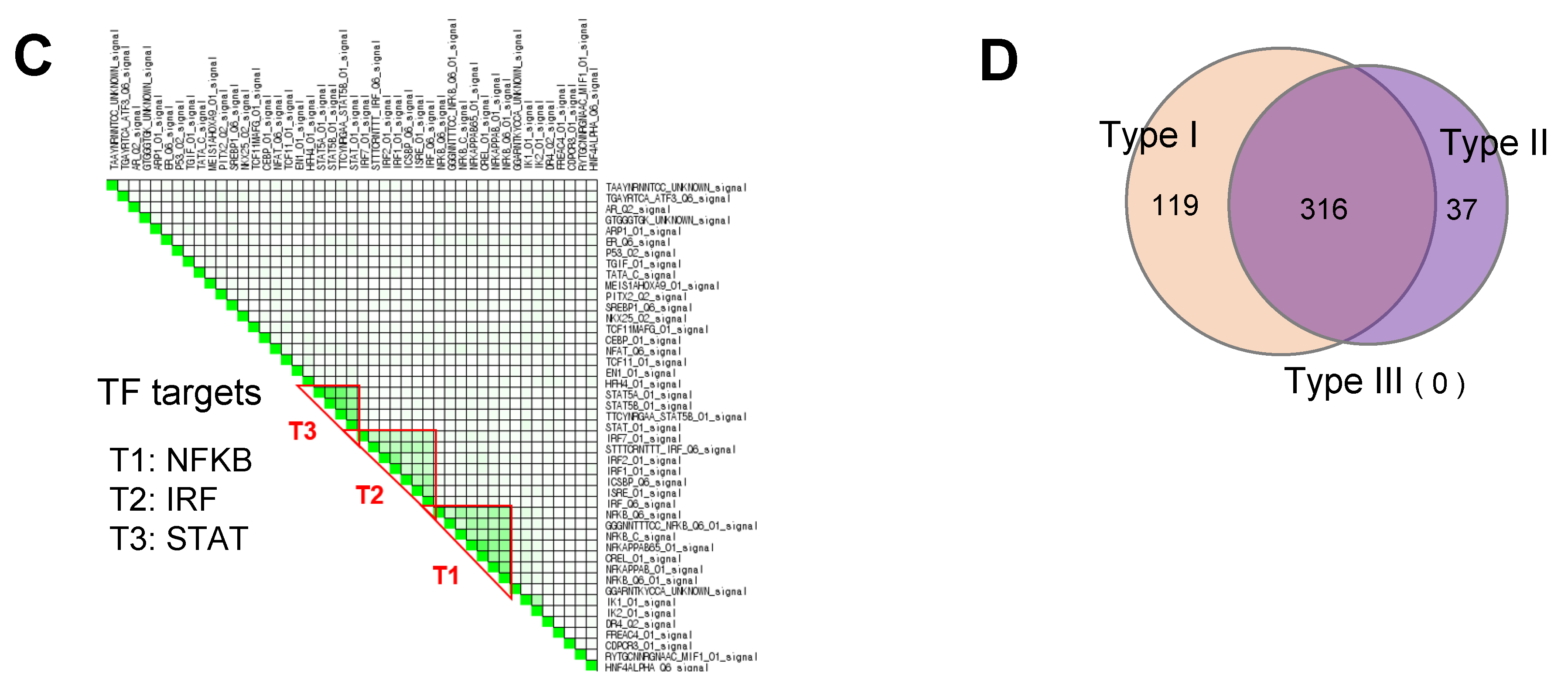
3.3. GSEA Leading-Edge Analysis Identified a Cluster of Type I IFN Signaling-Centered Host Defense Mechanisms
3.4. Fucoidan Stimulation Promotes BMDC Maturation and Activation In Vitro
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ponce, N.M.A.; Stortz, C.A. A Comprehensive and Comparative Analysis of the Fucoidan Compositional Data Across the Phaeophyceae. Front. Plant Sci. 2020, 11, 556312. [Google Scholar] [CrossRef] [PubMed]
- Luthuli, S.; Wu, S.; Cheng, Y.; Zheng, X.; Wu, M.; Tong, H. Therapeutic Effects of Fucoidan: A Review on Recent Studies. Mar. Drugs 2019, 17, 487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Xing, M.; Cao, Q.; Ji, A.; Liang, H.; Song, S. Biological Activities of Fucoidan and the Factors Mediating Its Therapeutic Effects: A Review of Recent Studies. Mar. Drugs 2019, 17, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apostolova, E.; Lukova, P.; Baldzhieva, A.; Katsarov, P.; Nikolova, M.; Iliev, I.; Peychev, L.; Trica, B.; Oancea, F.; Delattre, C.; et al. Immunomodulatory and Anti-Inflammatory Effects of Fucoidan: A Review. Polymers 2020, 12, 2338. [Google Scholar] [CrossRef] [PubMed]
- Sanjeewa, K.K.A.; Herath, K.; Yang, H.W.; Choi, C.S.; Jeon, Y.J. Anti-Inflammatory Mechanisms of Fucoidans to Treat Inflammatory Diseases: A Review. Mar. Drugs 2021, 19, 678. [Google Scholar] [CrossRef]
- Lazear, H.M.; Schoggins, J.W.; Diamond, M.S. Shared and Distinct Functions of Type I and Type III Interferons. Immunity 2019, 50, 907–923. [Google Scholar] [CrossRef]
- Mandal, P.; Mateu, C.G.; Chattopadhyay, K.; Pujol, C.A.; Damonte, E.B.; Ray, B. Structural features and antiviral activity of sulphated fucans from the brown seaweed Cystoseira indica. Antivir. Chem. Chemother. 2007, 18, 153–162. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, K.; Nakano, T.; Hashimoto, M.; Kanekiyo, K.; Hayashi, T. Defensive effects of a fucoidan from brown alga Undaria pinnatifida against herpes simplex virus infection. Int. Immunopharmacol. 2008, 8, 109–116. [Google Scholar] [CrossRef]
- Rabanal, M.; Ponce, N.M.; Navarro, D.A.; Gomez, R.M.; Stortz, C.A. The system of fucoidans from the brown seaweed Dictyota dichotoma: Chemical analysis and antiviral activity. Carbohydr. Polym. 2014, 101, 804–811. [Google Scholar] [CrossRef]
- Krylova, N.V.; Ermakova, S.P.; Lavrov, V.F.; Leneva, I.A.; Kompanets, G.G.; Iunikhina, O.V.; Nosik, M.N.; Ebralidze, L.K.; Falynskova, I.N.; Silchenko, A.S.; et al. The Comparative Analysis of Antiviral Activity of Native and Modified Fucoidans from Brown Algae Fucus evanescens In Vitro and In Vivo. Mar. Drugs 2020, 18, 224. [Google Scholar] [CrossRef] [Green Version]
- Prokofjeva, M.M.; Imbs, T.I.; Shevchenko, N.M.; Spirin, P.V.; Horn, S.; Fehse, B.; Zvyagintseva, T.N.; Prassolov, V.S. Fucoidans as Potential Inhibitors of HIV-1. Mar. Drugs 2013, 11, 3000–3014. [Google Scholar] [CrossRef] [PubMed]
- Thuy, T.T.; Ly, B.M.; Van, T.T.; Quang, N.V.; Tu, H.C.; Zheng, Y.; Seguin-Devaux, C.; Mi, B.; Ai, U. Anti-HIV activity of fucoidans from three brown seaweed species. Carbohydr. Polym. 2015, 115, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Dinesh, S.; Menon, T.; Hanna, L.E.; Suresh, V.; Sathuvan, M.; Manikannan, M. In Vitro anti-HIV-1 activity of fucoidan from Sargassum swartzii. Int. J. Biol. Macromol. 2016, 82, 83–88. [Google Scholar] [CrossRef]
- Araya, N.; Takahashi, K.; Sato, T.; Nakamura, T.; Sawa, C.; Hasegawa, D.; Ando, H.; Aratani, S.; Yagishita, N.; Fujii, R.; et al. Fucoidan therapy decreases the proviral load in patients with human T-lymphotropic virus type-1-associated neurological disease. Antivir. Ther. 2011, 16, 89–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makarenkova, I.D.; Deriabin, P.G.; L’Vov, D.K.; Zviagintseva, T.N.; Besednova, N.N. Antiviral activity of sulfated polysaccharide from the brown algae Laminaria japonica against avian influenza A (H5N1) virus infection in the cultured cells. Vopr. Virusol. 2010, 55, 41–45. [Google Scholar] [PubMed]
- Borst, E.M.; Standker, L.; Wagner, K.; Schulz, T.F.; Forssmann, W.G.; Messerle, M. A peptide inhibitor of cytomegalovirus infection from human hemofiltrate. Antimicrob. Agents Chemother. 2013, 57, 4751–4760. [Google Scholar] [CrossRef] [Green Version]
- Elizondo-Gonzalez, R.; Cruz-Suarez, L.E.; Ricque-Marie, D.; Mendoza-Gamboa, E.; Rodriguez-Padilla, C.; Trejo-Avila, L.M. In vitro characterization of the antiviral activity of fucoidan from Cladosiphon okamuranus against Newcastle Disease Virus. Virol. J. 2012, 9, 307. [Google Scholar] [CrossRef] [Green Version]
- Aguilar-Briseño, J.A.; Cruz-Suarez, L.E.; Sassi, J.F.; Ricque-Marie, D.; Zapata-Benavides, P.; Mendoza-Gamboa, E.; Rodríguez-Padilla, C.; Trejo-Avila, L.M. Sulphated polysaccharides from Ulva clathrata and Cladosiphon okamuranus seaweeds both inhibit viral attachment/entry and cell-cell fusion, in NDV infection. Mar. Drugs 2015, 13, 697–712. [Google Scholar] [CrossRef]
- Hayashi, K.; Lee, J.B.; Nakano, T.; Hayashi, T. Anti-influenza A virus characteristics of a fucoidan from sporophyll of Undaria pinnatifida in mice with normal and compromised immunity. Microbes Infect. 2013, 15, 302–309. [Google Scholar] [CrossRef]
- Wang, W.; Wu, J.; Zhang, X.; Hao, C.; Zhao, X.; Jiao, G.; Shan, X.; Tai, W.; Yu, G. Inhibition of Influenza A Virus Infection by Fucoidan Targeting Viral Neuraminidase and Cellular EGFR Pathway. Sci. Rep. 2017, 7, 40760. [Google Scholar] [CrossRef]
- Sun, T.; Zhang, X.; Miao, Y.; Zhou, Y.; Shi, J.; Yan, M.; Chen, A. Studies on antiviral and immuno-regulation activity of low molecular weight fucoidan from Laminaria japonica. J. Ocean. Univ. China 2018, 17, 705–711. [Google Scholar] [CrossRef]
- Kuznetsova, T.A.; Ivanushko, L.A.; Persiyanova, E.V.; Shutikova, A.L.; Ermakova, S.P.; Khotimchenko, M.Y.; Besednova, N.N. Evaluation of adjuvant effects of fucoidane from brown seaweed Fucus evanescens and its structural analogues for the strengthening vaccines effectiveness. Biomed. Khim. 2017, 63, 553–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Li, J.; Tang, Y.; Lin, L.; Xie, Z.; Zhou, J.; Zhang, L.; Zhang, X.; Zhao, X.; Chen, Z.; et al. Fucoidan from Fucus vesiculosus suppresses hepatitis B virus replication by enhancing extracellular signal-regulated Kinase activation. Virol. J. 2017, 14, 178. [Google Scholar] [CrossRef] [Green Version]
- Kwon, P.S.; Oh, H.; Kwon, S.J.; Jin, W.; Zhang, F.; Fraser, K.; Hong, J.J.; Linhardt, R.J.; Dordick, J.S. Sulfated polysaccharides effectively inhibit SARS-CoV-2 in vitro. Cell Discov. 2020, 6, 50. [Google Scholar] [CrossRef]
- Song, S.; Peng, H.; Wang, Q.; Liu, Z.; Dong, X.; Wen, C.; Ai, C.; Zhang, Y.; Wang, Z.; Zhu, B. Inhibitory activities of marine sulfated polysaccharides against SARS-CoV-2. Food Funct. 2020, 11, 7415–7420. [Google Scholar] [CrossRef]
- Ray, B.; Ali, I.; Jana, S.; Mukherjee, S.; Pal, S.; Ray, S.; Schütz, M.; Marschall, M. Antiviral Strategies Using Natural Source-Derived Sulfated Polysaccharides in the Light of the COVID-19 Pandemic and Major Human Pathogenic Viruses. Viruses 2021, 14, 35. [Google Scholar] [CrossRef]
- Yim, S.K.; Kim, K.; Kim, I.H.; Chun, S.H.; Oh, T.H.; Kim, J.U.; Kim, J.W.; Jung, W.H.; Moon, H.S.; Ku, B.S.; et al. Inhibition of SARS-CoV-2 Virus Entry by the Crude Polysaccharides of Seaweeds and Abalone Viscera In Vitro. Mar. Drugs 2021, 19, 219. [Google Scholar] [CrossRef]
- Morokutti-Kurz, M.; Froba, M.; Graf, P.; Grosse, M.; Grassauer, A.; Auth, J.; Schubert, U.; Prieschl-Grassauer, E. Iota-carrageenan neutralizes SARS-CoV-2 and inhibits viral replication in vitro. PLoS ONE 2021, 16, e0237480. [Google Scholar] [CrossRef] [PubMed]
- McNab, F.; Mayer-Barber, K.; Sher, A.; Wack, A.; O’Garra, A. Type I interferons in infectious disease. Nat. Rev. Immunol. 2015, 15, 87–103. [Google Scholar] [CrossRef]
- Crouse, J.; Kalinke, U.; Oxenius, A. Regulation of antiviral T cell responses by type I interferons. Nat. Rev. Immunol. 2015, 15, 231–242. [Google Scholar] [CrossRef]
- Lei, X.; Dong, X.; Ma, R.; Wang, W.; Xiao, X.; Tian, Z.; Wang, C.; Wang, Y.; Li, L.; Ren, L.; et al. Activation and evasion of type I interferon responses by SARS-CoV-2. Nat. Commun. 2020, 11, 3810. [Google Scholar] [CrossRef] [PubMed]
- Park, A.; Iwasaki, A. Type I and Type III Interferons—Induction, Signaling, Evasion, and Application to Combat COVID-19. Cell Host. Microbe 2020, 27, 870–878. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.; Cao, Z.; Xie, X.; Zhang, X.; Chen, J.Y.; Wang, H.; Menachery, V.D.; Rajsbaum, R.; Shi, P.Y. Evasion of Type I Interferon by SARS-CoV-2. Cell Rep. 2020, 33, 108234. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, O.; Akira, S. Innate immunity to virus infection. Immunol. Rev. 2009, 227, 75–86. [Google Scholar] [CrossRef]
- Iwasaki, A. A virological view of innate immune recognition. Annu. Rev. Microbiol. 2012, 66, 177–196. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.Z.; Wang, S.Y.; Zheng, Z.Q.; Yi, H.; Li, W.W.; Xu, Z.S.; Wang, Y.Y. SARS-CoV-2 membrane glycoprotein M antagonizes the MAVS-mediated innate antiviral response. Cell Mol. Immunol. 2021, 18, 613–620. [Google Scholar] [CrossRef]
- Kim, Y.M.; Shin, E.C. Type I and III interferon responses in SARS-CoV-2 infection. Exp. Mol. Med. 2021, 53, 750–760. [Google Scholar] [CrossRef]
- Blanco-Melo, D.; Nilsson-Payant, B.E.; Liu, W.C.; Uhl, S.; Hoagland, D.; Moller, R.; Jordan, T.X.; Oishi, K.; Panis, M.; Sachs, D.; et al. Imbalanced Host Response to SARS-CoV-2 Drives Development of COVID-19. Cell 2020, 181, 1036–1045.e1039. [Google Scholar] [CrossRef]
- Hadjadj, J.; Yatim, N.; Barnabei, L.; Corneau, A.; Boussier, J.; Smith, N.; Pere, H.; Charbit, B.; Bondet, V.; Chenevier-Gobeaux, C.; et al. Impaired type I interferon activity and inflammatory responses in severe COVID-19 patients. Science 2020, 369, 718–724. [Google Scholar] [CrossRef]
- Soto, J.A.; Galvez, N.M.S.; Andrade, C.A.; Pacheco, G.A.; Bohmwald, K.; Berrios, R.V.; Bueno, S.M.; Kalergis, A.M. The Role of Dendritic Cells During Infections Caused by Highly Prevalent Viruses. Front. Immunol. 2020, 11, 1513. [Google Scholar] [CrossRef]
- Marongiu, L.; Valache, M.; Facchini, F.A.; Granucci, F. How dendritic cells sense and respond to viral infections. Clin. Sci. 2021, 135, 2217–2242. [Google Scholar] [CrossRef] [PubMed]
- Swiecki, M.; Colonna, M. The multifaceted biology of plasmacytoid dendritic cells. Nat. Rev. Immunol. 2015, 15, 471–485. [Google Scholar] [CrossRef] [PubMed]
- Arunachalam, P.S.; Wimmers, F.; Mok, C.K.P.; Perera, R.; Scott, M.; Hagan, T.; Sigal, N.; Feng, Y.; Bristow, L.; Tak-Yin Tsang, O.; et al. Systems biological assessment of immunity to mild versus severe COVID-19 infection in humans. Science 2020, 369, 1210–1220. [Google Scholar] [CrossRef] [PubMed]
- Laing, A.G.; Lorenc, A.; Del Molino Del Barrio, I.; Das, A.; Fish, M.; Monin, L.; Munoz-Ruiz, M.; McKenzie, D.R.; Hayday, T.S.; Francos-Quijorna, I.; et al. Author Correction: A dynamic COVID-19 immune signature includes associations with poor prognosis. Nat. Med. 2020, 26, 1951. [Google Scholar] [CrossRef] [PubMed]
- Peruzzi, B.; Bencini, S.; Capone, M.; Mazzoni, A.; Maggi, L.; Salvati, L.; Vanni, A.; Orazzini, C.; Nozzoli, C.; Morettini, A.; et al. Quantitative and qualitative alterations of circulating myeloid cells and plasmacytoid DC in SARS-CoV-2 infection. Immunology 2020, 161, 345–353. [Google Scholar] [CrossRef]
- Liu, C.; Martins, A.J.; Lau, W.W.; Rachmaninoff, N.; Chen, J.; Imberti, L.; Mostaghimi, D.; Fink, D.L.; Burbelo, P.D.; Dobbs, K.; et al. Time-resolved systems immunology reveals a late juncture linked to fatal COVID-19. Cell 2021, 184, 1836–1857.e1822. [Google Scholar] [CrossRef] [PubMed]
- Saichi, M.; Ladjemi, M.Z.; Korniotis, S.; Rousseau, C.; Ait Hamou, Z.; Massenet-Regad, L.; Amblard, E.; Noel, F.; Marie, Y.; Bouteiller, D.; et al. Single-cell RNA sequencing of blood antigen-presenting cells in severe COVID-19 reveals multi-process defects in antiviral immunity. Nat. Cell Biol. 2021, 23, 538–551. [Google Scholar] [CrossRef]
- Hilligan, K.L.; Ronchese, F. Antigen presentation by dendritic cells and their instruction of CD4+ T helper cell responses. Cell Mol. Immunol. 2020, 17, 587–599. [Google Scholar] [CrossRef]
- Cabeza-Cabrerizo, M.; Cardoso, A.; Minutti, C.M.; Pereira da Costa, M.; Reis e Sousa, C. Dendritic Cells Revisited. Annu. Rev. Immunol. 2021, 39, 131–166. [Google Scholar] [CrossRef]
- Parackova, Z.; Zentsova, I.; Bloomfield, M.; Vrabcova, P.; Smetanova, J.; Klocperk, A.; Meseznikov, G.; Casas Mendez, L.F.; Vymazal, T.; Sediva, A. Disharmonic Inflammatory Signatures in COVID-19: Augmented Neutrophils’ but Impaired Monocytes’ and Dendritic Cells’ Responsiveness. Cells 2020, 9, 2206. [Google Scholar] [CrossRef]
- Marongiu, L.; Protti, G.; Facchini, F.A.; Valache, M.; Mingozzi, F.; Ranzani, V.; Putignano, A.R.; Salviati, L.; Bevilacqua, V.; Curti, S.; et al. Maturation signatures of conventional dendritic cell subtypes in COVID-19 suggest direct viral sensing. Eur. J. Immunol. 2022, 52, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Kim, Y.; Lee, H.; Kim, J.; Han, B.K.; Go, E.; Kwon, S.; Kang, J.G.; You, S.; Kwon, J. Carrot Pomace Polysaccharide (CPP) Improves Influenza Vaccine Efficacy in Immunosuppressed Mice via Dendritic Cell Activation. Nutrients 2020, 12, 2740. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, A.C.; Bourbon, A.I.; Cerqueira, M.A.; Maricato, E.; Nunes, C.; Coimbra, M.A.; Vicente, A.A. Chitosan/fucoidan multilayer nanocapsules as a vehicle for controlled release of bioactive compounds. Carbohydr. Polym. 2015, 115, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.M.; Liu, P.Y.; Chen, Y.A.; Tseng, H.Y.; Shen, P.C.; Hwang, P.A.; Hsu, H.L. Oligo-Fucoidan prevents IL-6 and CCL2 production and cooperates with p53 to suppress ATM signaling and tumor progression. Sci. Rep. 2017, 7, 11864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, X. iDEP Web Application for RNA-Seq Data Analysis. Methods Mol. Biol. 2021, 2284, 417–443. [Google Scholar]
- Ge, S.X.; Son, E.W.; Yao, R. iDEP: An integrated web application for differential expression and pathway analysis of RNA-Seq data. BMC Bioinform. 2018, 19, 534. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.M.; McFadden, G. Modulation of NF-kappaB signalling by microbial pathogens. Nat. Rev. Microbiol. 2011, 9, 291–306. [Google Scholar] [CrossRef]
- Gonzalez-Navajas, J.M.; Lee, J.; David, M.; Raz, E. Immunomodulatory functions of type I interferons. Nat. Rev. Immunol. 2012, 12, 125–135. [Google Scholar] [CrossRef] [Green Version]
- Mogensen, T.H. IRF and STAT Transcription Factors—From Basic Biology to Roles in Infection, Protective Immunity, and Primary Immunodeficiencies. Front. Immunol. 2018, 9, 3047. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]
- Reimand, J.; Isserlin, R.; Voisin, V.; Kucera, M.; Tannus-Lopes, C.; Rostamianfar, A.; Wadi, L.; Meyer, M.; Wong, J.; Xu, C.; et al. Pathway enrichment analysis and visualization of omics data using g:Profiler, GSEA, Cytoscape and EnrichmentMap. Nat. Protoc. 2019, 14, 482–517. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Tan, X.; Zhang, Y.; Li, F.; Luo, P.; Liu, H. Molecular Targets and Related Biologic Activities of Fucoidan: A Review. Mar. Drugs 2020, 18, 376. [Google Scholar] [CrossRef] [PubMed]
- Rusinova, I.; Forster, S.; Yu, S.; Kannan, A.; Masse, M.; Cumming, H.; Chapman, R.; Hertzog, P.J. Interferome v2.0: An updated database of annotated interferon-regulated genes. Nucleic Acids Res. 2013, 41, D1040–D1046. [Google Scholar] [CrossRef] [PubMed]
- Baranek, T.; Manh, T.P.; Alexandre, Y.; Maqbool, M.A.; Cabeza, J.Z.; Tomasello, E.; Crozat, K.; Bessou, G.; Zucchini, N.; Robbins, S.H.; et al. Differential responses of immune cells to type I interferon contribute to host resistance to viral infection. Cell Host. Microbe 2012, 12, 571–584. [Google Scholar] [CrossRef] [Green Version]
- Stanifer, M.L.; Pervolaraki, K.; Boulant, S. Differential Regulation of Type I and Type III Interferon Signaling. Int. J. Mol. Sci. 2019, 20, 1445. [Google Scholar] [CrossRef] [Green Version]
- O’Garra, A. Cytokines induce the development of functionally heterogeneous T helper cell subsets. Immunity 1998, 8, 275–283. [Google Scholar] [CrossRef] [Green Version]
- Ferlazzo, G.; Pack, M.; Thomas, D.; Paludan, C.; Schmid, D.; Strowig, T.; Bougras, G.; Muller, W.A.; Moretta, L.; Munz, C. Distinct roles of IL-12 and IL-15 in human natural killer cell activation by dendritic cells from secondary lymphoid organs. Proc. Natl. Acad. Sci. USA 2004, 101, 16606–16611. [Google Scholar] [CrossRef] [Green Version]
- Mehta, P.; McAuley, D.F.; Brown, M.; Sanchez, E.; Tattersall, R.S.; Manson, J.J. COVID-19: Consider cytokine storm syndromes and immunosuppression. Lancet 2020, 395, 1033–1034. [Google Scholar] [CrossRef]
- Olagnier, D.; Farahani, E.; Thyrsted, J.; Blay-Cadanet, J.; Herengt, A.; Idorn, M.; Hait, A.; Hernaez, B.; Knudsen, A.; Iversen, M.B.; et al. SARS-CoV2-mediated suppression of NRF2-signaling reveals potent antiviral and anti-inflammatory activity of 4-octyl-itaconate and dimethyl fumarate. Nat. Commun. 2020, 11, 4938. [Google Scholar] [CrossRef]
- Cuadrado, A.; Pajares, M.; Benito, C.; Jiménez-Villegas, J.; Escoll, M.; Fernández-Ginés, R.; Garcia Yagüe, A.J.; Lastra, D.; Manda, G.; Rojo, A.I.; et al. Can Activation of NRF2 Be a Strategy against COVID-19? Trends Pharmacol. Sci. 2020, 41, 598–610. [Google Scholar] [CrossRef]
- Bousquet, J.; Cristol, J.P.; Czarlewski, W.; Anto, J.M.; Martineau, A.; Haahtela, T.; Fonseca, S.C.; Iaccarino, G.; Blain, H.; Fiocchi, A.; et al. Nrf2-interacting nutrients and COVID-19: Time for research to develop adaptation strategies. Clin. Transl. Allergy 2020, 10, 58. [Google Scholar] [CrossRef] [PubMed]
- Kelley, J.L.; Ozment, T.R.; Li, C.; Schweitzer, J.B.; Williams, D.L. Scavenger receptor-A (CD204): A two-edged sword in health and disease. Crit. Rev. Immunol. 2014, 34, 241–261. [Google Scholar] [CrossRef] [PubMed]
- Makarenkova, I.D.; Logunov, D.Y.; Tukhvatulin, A.I.; Semenova, I.B.; Besednova, N.N.; Zvyagintseva, T.N. Interactions between sulfated polysaccharides from sea brown algae and Toll-like receptors on HEK293 eukaryotic cells in vitro. Bull. Exp. Biol. Med. 2012, 154, 241–244. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Direction | Pathways | nGenes | adj.Pval |
|---|---|---|---|
| Down | Cell cycle | 24 | 8.90 × 10−9 |
| Regulated | DNA replication | 11 | 4.40 × 10−6 |
| Progesterone-mediated oocyte maturation | 12 | 1.70 × 10−3 | |
| Homologous recombination | 8 | 2.10 × 10−3 | |
| P53 signaling pathway | 11 | 2.10 × 10−3 | |
| Cellular senescence | 17 | 2.10 × 10−3 | |
| Oocyte meiosis | 13 | 2.10 × 10−3 | |
| Staphylococcus aureus infection | 8 | 4.30 × 10−3 | |
| Complement and coagulation cascades | 8 | 6.50 × 10−3 | |
| Human T-cell leukemia virus 1 infection | 19 | 8.60 × 10−3 | |
| Up | Cytokine–cytokine receptor interaction | 54 | 4.80 × 10−20 |
| Regulated | TNF signaling pathway | 39 | 5.70 × 10−16 |
| Measles | 36 | 5.00 × 10−12 | |
| Influenza A | 36 | 1.10 × 10−10 | |
| Epstein–Barr virus infection | 42 | 2.30 × 10−10 | |
| C-type lectin receptor signaling pathway | 29 | 4.30 × 10−10 | |
| Toll-like receptor signaling pathway | 26 | 1.90 × 10−9 | |
| JAK-STAT signaling pathway | 31 | 6.70 × 10−9 | |
| Pathways in cancer | 67 | 6.70 × 10−9 | |
| NOD-like receptor signaling pathway | 35 | 1.20 × 10−8 | |
| Viral protein interaction with cytokine and cytokine receptor | 20 | 3.90 × 10−8 | |
| NF-kappa B signaling pathway | 25 | 7.70 × 10−8 | |
| Type I diabetes mellitus | 15 | 7.70 × 10−8 | |
| Kaposi sarcoma-associated herpesvirus infection | 35 | 9.60 × 10−8 | |
| Antigen processing and presentation | 18 | 3.30 × 10−7 |
| Pathways | Genes |
|---|---|
| Irf7 target gene | Isg15 B2m Cd80 H2-K1 H2-M3 H2-Q4 Irf8 Irgm1 Cxcl10 Ifit1 Ifit2 Ifit3 Igtp Ikbkg Irf1 Irf4 Irf9 Tlr7 Gbp4 Mt2 Pml Lgals3bp Trim30a Ccl5 Spp1 Stat1 Tank Tap1 Tnf Traf6 Cmpk2 Oasl1 Ddx60 Oasl2 Usp18 Parp12 Oas3 Xaf1 Parp14 Rsad2 Zbp1 Iigp1 Rtp4 Bst2 Ifi35 Ifih1 Uba7 Dhx58 Ifi44 |
| Nfkb1 target gene | Acp5 Birc3 Cxcr5 Casp1 Cd14 Cebpb Cflar Socs3 Csf1 Cxcl1 Cfb Hif1a Cxcl10 Ikbkg Il12a Il12b Il15 Il1a Il1b Il1rn Il2ra Il6 Kdr Gadd45b Nfkb1 Nfkb2 Nfkbia Nfkbib Nfkbie ENSMUSG00000023947 Nos2 Sqstm1 Eif2ak2 Ptgs2 Ripk2 Rel Ccl3 Ccl4 Ccl5 Cxcl2 Sdc4 Tapbp Tnf Tnfaip3 Traf1 Traf6 Vegfa Akt3 Malt1 Map3k8 Tnfsf15 Ebi3 Map3k14 Zbp1 Nfkbiz Il23a |
| Stat1 target gene | Isg15 Ahr Apbb2 Rhoc Atf3 B2m Bmpr2 Cacna1d Casp1 Casp4 Ccnd2 Socs3 Socs1 Crem Fosl1 H2-K1 H2-M3 H2-Q4 Irf8 Irgm1 Cxcl10 Ifit1 Ifit2 Ifit3 Il18bp Igtp Il2ra Il6 Irf1 Irf4 Irf9 Jak3 Lta Hook2 Man1a Mt1 Mt2 Nos2 Osm Enpp2 Pik3r1 Pml Lgals3bp Eif2ak2 Dusp1 Rab10 Trim30a Rras Ccl2 Sphk1 Trim21 Stat1 Stat3 Stat5a Tap1 Socs2 Tnf Tnfrsf8 Traf6 Cmpk2 Ubc Vdr Vegfa Yes1 Ikzf4 Gbp5 Oasl1 Ppp2r3a Oasl2 Usp18 Parp12 Vasn Mllt6 Oas3 Etv3 Dock6 Pik3r5 Txndc17 Map3k14 Irf7 Parp14 Gbp4 Mtor Rsad2 Zbp1 Iigp1 Cd274 Asph Herc6 Cpeb4 Rtp4 Ifi35 Ifih1 Ppme1 Uba7 Dtx2 Ddit4 Ccdc6 Usp6nl Ifi44 |
| Irf9 target gene | Isg15 B2m H2-K1 H2-M3 H2-Q4 Irf8 Irgm1 Ifit1 Ifit3 Igtp Irf1 Irf4 Mt2 Pml Stat1 Stat2 Stat3 Stat4 Stat5a Oasl1 Oasl2 Usp18 Oas3 Irf7 Parp14 Rsad2 Rtp4 Ifi35 |
| Irf5 target gene | B2m H2-K1 H2-M3 H2-Q4 Irf8 Il12a Il12b Il1b Il6 Irf1 Irf4 Irf9 Tlr7 Mt2 Pml Ccl3 Ccl4 Ccl5 Stat1 Tnf Traf6 Ubc Oasl1 Oas3 Irf7 |
| Rela target gene | Spred2 Ahr Birc3 Casp1 Cd14 Cebpb Socs3 Socs1 Csf3 Igf2bp1 F3 Cxcl1 Hif1a Ier3 Cxcl10 Ikbkg Il12a Il12b Il1a Il1b Il2ra Il6 Maff Gadd45b Nfkb1 Nfkb2 Nfkbia Nfkbib Nfkbie ENSMUSG00000023947 Nos2 Nr4a2 Sqstm1 Ptgs2 Ripk2 Rel Ccl2 Ccl3 Ccl4 Ccl5 Cxcl2 Sema4c Socs2 Tnf Tnfaip3 Tnfrsf1b Tnfrsf9 Traf1 Traf6 Ubc Vegfa Akt3 Malt1 Mllt6 Tnfsf15 Ebi3 Map3k14 Zbp1 Rffl Nfkbiz Il23a |
| Irf1 target gene | Isg15 Spred2 Agrn B2m Cdkn2b F3 Fmn1 H2-K1 H2-M3 H2-Q4 Irf8 Irgm1 Cxcl10 Igtp Il12a Il12b Il15ra Il6 Irf4 Irf9 Mt2 Osm Pik3r1 Pml Lgals3bp Eif2ak2 PSME2b Stat1 Stat3 Stat4 Dtx3l Ncoa7 Tap1 Tapbp Tgif1 Oasl1 Ets2 Usp18 Oas3 Tmtc2 Ccbe1 Flrt2 C1ra Irf7 Pcdh7 Parp14 Gbp4 Rybp Chst11 Cd274 Cxcl16 Grina Rtp4 Tmem140 Bst2 Ifi35 Znfx1 |
| Irf8 target gene | B2m H2-K1 H2-M3 H2-Q4 Il12a Il12b Irf1 Irf4 Irf9 Mt2 Pml Ccl5 Stat1 Traf6 Ubc Oasl1 Oas3 Irf7 |
| Relb target gene | Ahr Bcl3 Cdkn1a Daxx Cxcl1 Ikbkg Nfkb1 Nfkb2 Nfkbia Nfkbib Nfkbie ENSMUSG00000023947 Cxcl2 Tnf Cd40 Map3k14 |
| Irf6 target gene | B2m H2-K1 H2-M3 H2-Q4 Irf8 Irf1 Irf4 Irf9 Mt2 Pml Stat1 Oasl1 Oas3 Irf7 |
| Pml target gene | B2m Daxx H2-K1 H2-M3 H2-Q4 Irf8 Irf1 Irf4 Irf9 Mt2 Pml Skil Stat1 Ubc Oasl1 Oas3 Irf7 Mtor |
| Nfkb2 target gene | Birc3 Bcl3 Casp1 Ikbkg Il1b Nfkb1 Nfkbia Nfkbib Nfkbie ENSMUSG00000023947 Nos2 Ptgs2 Tnf Traf1 Traf6 Map3k14 Zbp1 Nfkbiz |
| Irf2 target gene | Isg15 Spred2 Agrn Fabp4 B2m Cdkn2b F3 Fmn1 H2-K1 H2-M3 H2-Q4 Irf8 Cxcl10 Il15ra Irf1 Irf4 Irf9 Mt2 Osm Pik3r1 Pml Lgals3bp PSME2b Stat1 Ncoa7 Tap1 Tapbp Suco Oasl1 Usp18 Oas3 Tmtc2 Rabgap1l Ccbe1 Irf7 Pcdh7 Rybp Chst11 Cxcl16 Grina Rtp4 Bst2 Ifi35 Ogfr Mvp Znfx1 |
| Tnfaip3 target gene | Ikbkg Nfkb1 Nfkbia Nfkbie ENSMUSG00000023947 Ripk2 Tnf Tnfaip3 Traf1 Traf6 Tnip3 Tax1bp1 Tnip1 Nfkbiz |
| Ikbkg target gene | Erc1 Birc3 Hspa1b Ikbkg Nfkb1 Nfkb2 Nfkbia Nfkbib Nfkbie ENSMUSG00000023947 Tank Tnfaip3 Cd40 Traf1 Traf6 Ubc Akt3 Malt1 Map3k8 Rnf31 Map3k14 Irf7 Zbp1 Nfkbiz |
| NAME | SIZE | NES | NOM p-Value |
|---|---|---|---|
| REACTOME_INTERFERON_ALPHA_BETA_SIGNALING | 58 | 2.028 | 0.000 |
| WP_CYTOKINES_AND_INFLAMMATORY_RESPONSE | 24 | 1.933 | 0.000 |
| WP_SARSCOV2_INNATE_IMMUNITY_EVASION_AND_CELLSPECIFIC_IMMUNE_RESPONSE | 62 | 1.925 | 0.000 |
| WP_TYPE_I_INTERFERON_INDUCTION_AND_SIGNALING_DURING_SARSCOV2_INFECTION | 28 | 1.838 | 0.000 |
| WP_OVERVIEW_OF_INTERFERONSMEDIATED_SIGNALING_PATHWAY | 30 | 1.829 | 0.000 |
| WP_SARS_CORONAVIRUS_AND_INNATE_IMMUNITY | 27 | 1.825 | 0.000 |
| WP_TYPE_II_INTERFERON_SIGNALING_IFNG | 31 | 1.821 | 0.000 |
| WP_EBV_LMP1_SIGNALING | 23 | 1.816 | 0.005 |
| WP_HOSTPATHOGEN_INTERACTION_OF_HUMAN_CORONA_VIRUSES_INTERFERON_INDUCTION | 32 | 1.805 | 0.003 |
| KEGG_RIG_I_LIKE_RECEPTOR_SIGNALING_PATHWAY | 63 | 1.795 | 0.000 |
| REACTOME_DDX58_IFIH1_MEDIATED_INDUCTION_OF_INTERFERON_ALPHA_BETA | 72 | 1.767 | 0.000 |
| WP_TOLLLIKE_RECEPTOR_SIGNALING_PATHWAY | 99 | 1.766 | 0.000 |
| KEGG_TOLL_LIKE_RECEPTOR_SIGNALING_PATHWAY | 97 | 1.763 | 0.000 |
| REACTOME_TRAF6_MEDIATED_NF_KB_ACTIVATION | 23 | 1.756 | 0.005 |
| REACTOME_REGULATION_OF_IFNA_SIGNALING | 22 | 1.755 | 0.000 |
| REACTOME_TRAF6_MEDIATED_IRF7_ACTIVATION | 25 | 1.737 | 0.005 |
| REACTOME_INTERLEUKIN_4_AND_INTERLEUKIN_13_SIGNALING | 101 | 1.724 | 0.000 |
| REACTOME_SIGNALING_BY_INTERLEUKINS | 422 | 1.720 | 0.000 |
| WP_MIRNAS_INVOLVEMENT_IN_THE_IMMUNE_RESPONSE_IN_SEPSIS | 53 | 1.713 | 0.000 |
| KEGG_CYTOSOLIC_DNA_SENSING_PATHWAY | 49 | 1.660 | 0.003 |
| WP_REGULATION_OF_TOLLLIKE_RECEPTOR_SIGNALING_PATHWAY | 137 | 1.655 | 0.000 |
| REACTOME_ZBP1_DAI_MEDIATED_INDUCTION_OF_TYPE_I_IFNS | 20 | 1.637 | 0.019 |
| REACTOME_INTERLEUKIN_1_SIGNALING | 99 | 1.618 | 0.000 |
| WP_NOVEL_INTRACELLULAR_COMPONENTS_OF_RIGILIKE_RECEPTOR_RLR_PATHWAY | 57 | 1.608 | 0.000 |
| WP_STING_PATHWAY_IN_KAWASAKILIKE_DISEASE_AND_COVID19 | 18 | 1.606 | 0.010 |
| WP_THE_HUMAN_IMMUNE_RESPONSE_TO_TUBERCULOSIS | 22 | 1.605 | 0.016 |
| WP_SIGNAL_TRANSDUCTION_THROUGH_IL1R | 34 | 1.603 | 0.011 |
| WP_IL1_SIGNALING_PATHWAY | 56 | 1.595 | 0.000 |
| REACTOME_RIP_MEDIATED_NFKB_ACTIVATION_VIA_ZBP1 | 17 | 1.565 | 0.026 |
| Number of Gene Sets | Gene |
|---|---|
| 21 | Nfkb1 |
| 19 | Ikbkb, Ikbkg, Nfkbia |
| 18 | Ifnb1, Traf6 |
| 15 | Tbk1 |
| 14 | Stat1 |
| 12 | Ikbke, Tnf, Mapk8 |
| 11 | Il1b, Ifnar2, Ifnar1, Nfkb1b, Irf7 |
| 10 | Stat2, Cxcl10, Il6 |
| 9 | Nfkb2, Ddx58 |
| 8 | Jun, Ticami |
| 7 | Il12b, Pi3kr1, Tollip, Ifih1, Tlr7, Tlr3, Socs1, Irf9, Ccl5, Ccl4 |
| 6 | Trim25, Il12a, Nkiras1, Il1a, Map3k1, Map2k1, Irak2, Traf2, Pik3ca, Ccl3 |
| 5 | Jak2, Jak1, Tank, Saa1, Map3k8, Socs3, Isg15, Cxcl9, Irak3, Ccl2 |
| 4 | App, Icam1, Dhx58, Il1r1, Cd14, Ifna4, Il10, Peli1, Tlr4, Psmb8, Cd86, Cd80, Irf1, Nos2, Il1rap, Sqstm1, Ly96, Cxcl11, Pik3cb, Hsp90aa1, Irf4, Traf3 |
| 3 | Il1rn, Ifit2, Akt3, Atg12, Pik3r5, Eif2ak2, Cyld, Rel, Cd40, Syk, Ifngr2, Tlr1, Tlr6, Spp1, Psmb9, Zbp1, Ube2v1, Ubc, Map3k14, Casp8, Oas2, Oas3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, S.; Jeon, S.A.; Heo, B.Y.; Kang, J.-G.; Jung, Y.; Duong, P.T.T.; Song, I.-C.; Kim, J.-H.; Kim, S.-Y.; Kwon, J. Gene Set Enrichment Analysis Reveals That Fucoidan Induces Type I IFN Pathways in BMDC. Nutrients 2022, 14, 2242. https://doi.org/10.3390/nu14112242
Choi S, Jeon SA, Heo BY, Kang J-G, Jung Y, Duong PTT, Song I-C, Kim J-H, Kim S-Y, Kwon J. Gene Set Enrichment Analysis Reveals That Fucoidan Induces Type I IFN Pathways in BMDC. Nutrients. 2022; 14(11):2242. https://doi.org/10.3390/nu14112242
Chicago/Turabian StyleChoi, Suyoung, Sol A Jeon, Bu Yeon Heo, Ju-Gyeong Kang, Yunju Jung, Pham Thi Thuy Duong, Ik-Chan Song, Jeong-Hwan Kim, Seon-Young Kim, and Jaeyul Kwon. 2022. "Gene Set Enrichment Analysis Reveals That Fucoidan Induces Type I IFN Pathways in BMDC" Nutrients 14, no. 11: 2242. https://doi.org/10.3390/nu14112242
APA StyleChoi, S., Jeon, S. A., Heo, B. Y., Kang, J.-G., Jung, Y., Duong, P. T. T., Song, I.-C., Kim, J.-H., Kim, S.-Y., & Kwon, J. (2022). Gene Set Enrichment Analysis Reveals That Fucoidan Induces Type I IFN Pathways in BMDC. Nutrients, 14(11), 2242. https://doi.org/10.3390/nu14112242