
Safety of Dietary Guanidinoacetic Acid: A Villain of a Good Guy?

{kind=link}
{kind=link}
Abstract
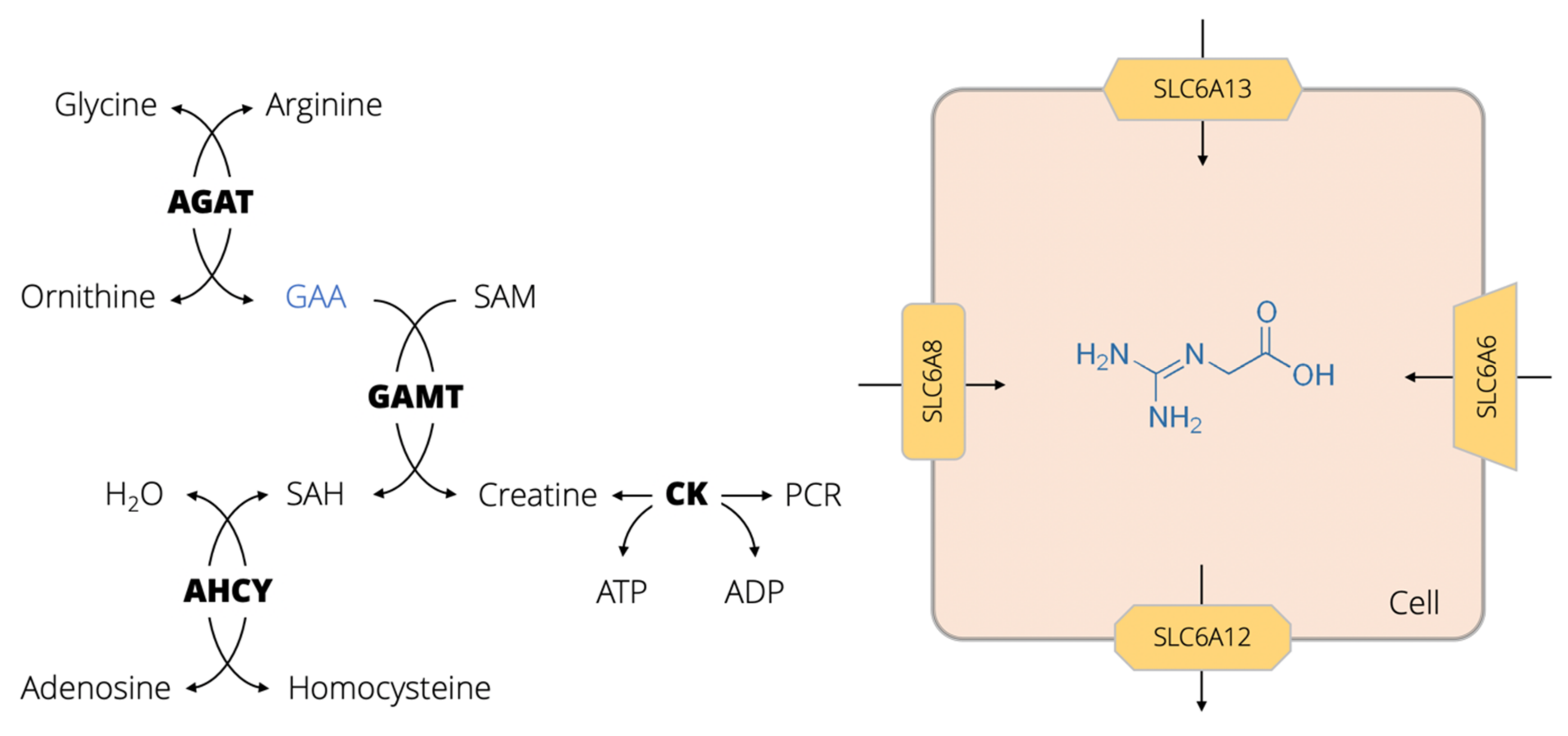
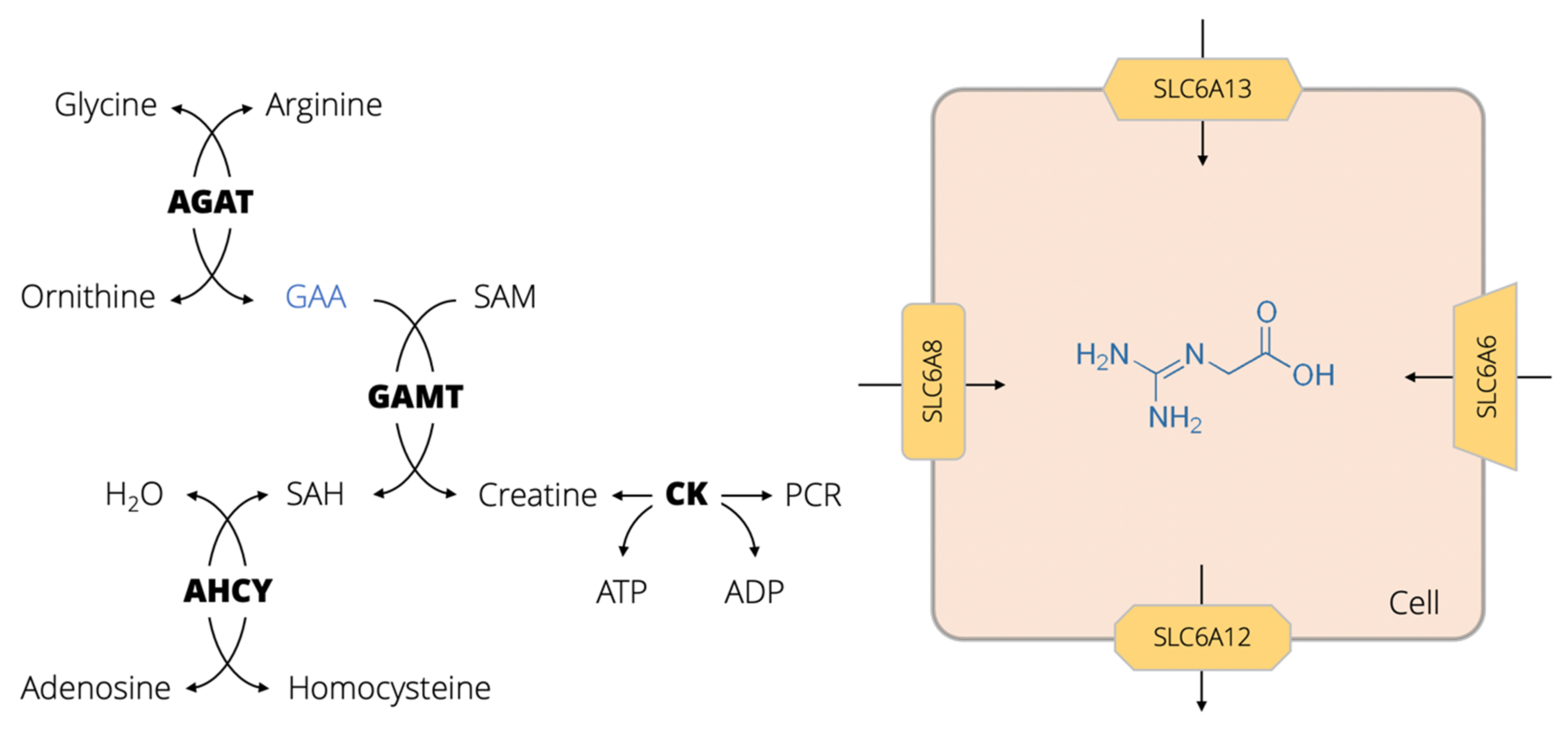
:1. GAA Physiology, Biomolecular Interactions and Pathways
2. GAA as a Dietary Agent in Human Nutrition
3. Dietary GAA Safety and Toxicity
3.1. Methyl Group Depletion
3.2. Hyperhomocysteinemia
3.3. Neurotoxicity
3.4. Other Adverse Effects
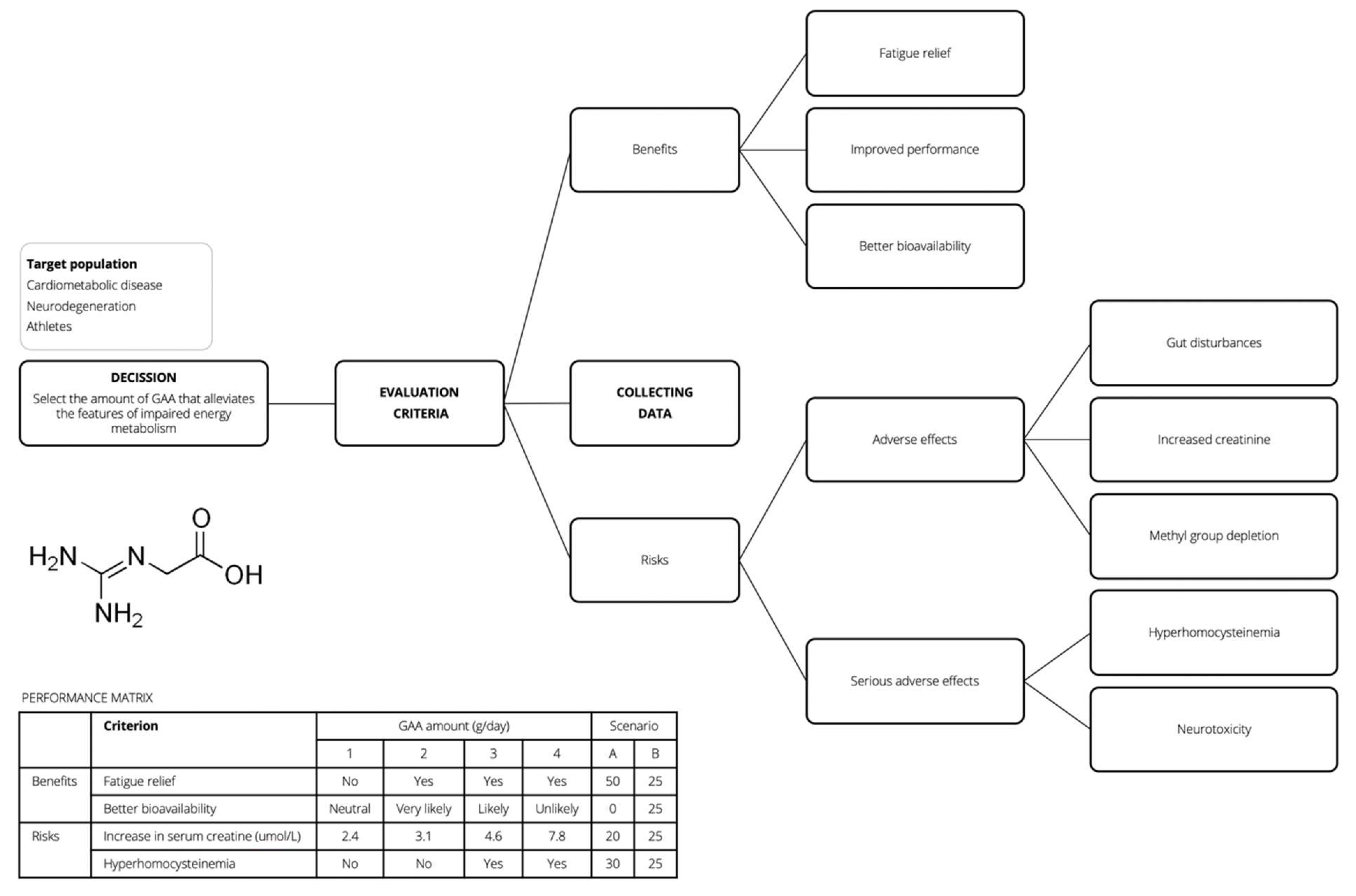
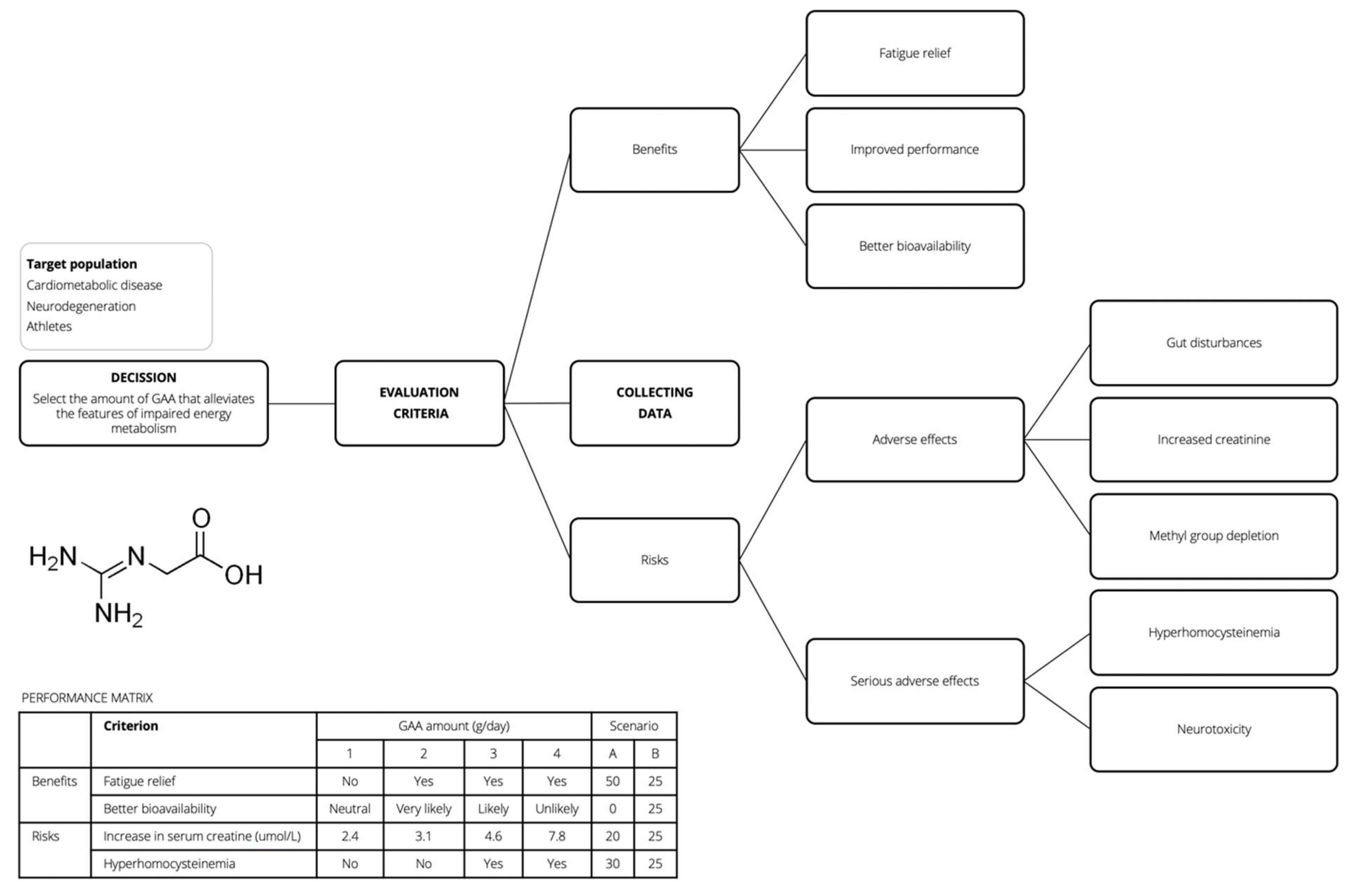
4. Blueprint for GAA Risk-Benefit Assessment
5. Conclusions
Funding
Conflicts of Interest
References
- Edison, E.E.; Brosnan, M.E.; Meyer, C.; Brosnan, J.T. Creatine synthesis: Production of guanidinoacetate by the rat and human kidney in vivo. Am. J. Physiol. Renal Physiol. 2007, 293, F1799–F1804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, S.A.; Gajera, C.R.; Wawro, A.M.; Corces, M.R.; Montine, T.J. GATM and GAMT synthesize creatine locally throughout the mammalian body and within oligodendrocytes of the brain. Brain Res. 2021, 1770, 147627. [Google Scholar] [CrossRef]
- Ostojic, S.M. Creatine synthesis in the skeletal muscle: The times they are a-changin’. Am. J. Physiol. Endocrinol. Metab. 2021, 320, E390–E391. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.B. Creatine: Biosynthesis, Regulation, and Function. Adv. Enzymol. Relat. Areas Mol. Biol. 1979, 50, 177–242. [Google Scholar] [CrossRef] [PubMed]
- Wallimann, T.; Tokarska-Schlattner, M.; Schlattner, U. The creatine kinase system and pleiotropic effects of creatine. Amino Acids 2011, 40, 1271–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tachikawa, M.; Kasai, Y.; Yokoyama, R.; Fujinawa, J.; Ganapathy, V.; Terasaki, T.; Hosoya, K.-I. The blood-brain barrier transport and cerebral distribution of guanidinoacetate in rats: Involvement of creatine and taurine transporters. J. Neurochem. 2009, 111, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Tachikawa, M.; Yashiki, A.; Akanuma, S.-I.; Matsukawa, H.; Ide, S.; Minami, M.; Hosoya, K.-I. Astrocytic γ-aminobutyric acid (GABA) transporters mediate guanidinoacetate transport in rat brain. Neurochem. Int. 2018, 113, 1–7. [Google Scholar] [CrossRef]
- Ostojic, S.M. Tackling guanidinoacetic acid for advanced cellular bioenergetics. Nutrition 2017, 34, 55–57. [Google Scholar] [CrossRef]
- Jomura, R.; Tanno, Y.; Akanuma, S.-I.; Kubo, Y.; Tachikawa, M.; Hosoya, K.-I. Monocarboxylate transporter 12 as a guanidinoacetate efflux transporter in renal proximal tubular epithelial cells. Biochim. Biophys. Acta BBA Biomembr. 2020, 1862, 183434. [Google Scholar] [CrossRef]
- Curt, M.J.-C.; Cheillan, D.; Briand, G.; Salomons, G.S.; Mention-Mulliez, K.; Dobbelaere, D.; Cuisset, J.-M.; Lion-François, L.; Portes, V.D.; Chabli, A.; et al. Creatine and guanidinoacetate reference values in a French population. Mol. Genet. Metab. 2013, 110, 263–267. [Google Scholar] [CrossRef]
- Ostojic, S.M.; Ratgeber, L.; Olah, A.; Betlehem, J.; Acs, P. Guanidinoacetic acid deficiency: A new entity in clinical medicine? Int. J. Med. Sci. 2020, 17, 2544–2550. [Google Scholar] [CrossRef]
- Ostojic, S.M. Advanced physiological roles of guanidinoacetic acid. Eur. J. Nutr. 2015, 54, 1211–1215. [Google Scholar] [CrossRef]
- European Food Safety Authority. Panel on Additives and Products or Substances used in Animal Feed (FEEDAP). Safety and efficacy of guanidinoacetic acid for chickens for fattening, breeder hens and roosters, and pigs. EFSA J. 2016, 14, 4394. [Google Scholar] [CrossRef] [Green Version]
- Ostojic, S.M.; Stajer, V.; Ratgeber, L.; Betlehem, J.; Acs, P. Guanidinoacetic Acid Consumption via Regular Diet in Adults. Ann. Nutr. Metab. 2021, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Ostojic, S.M. Human gut microbiota as a source of guanidinoacetic acid. Med. Hypotheses 2020, 142, 109745. [Google Scholar] [CrossRef] [PubMed]
- Borsook, H.; Borsook, M.E. The biochemical basis of betaine-glycocyamine therapy. Ann. West. Med. Surg. 1951, 5, 825–829. [Google Scholar]
- Graybiel, A.; Patterson, C.A. Use of betaine and glycocyamine in the treatment of patients with heart disease: Preliminary report. Ann. West. Med. Surg. 1951, 5, 863–875. [Google Scholar] [PubMed]
- Van Zandt, V.; Borsook, H. New biochemical approach to the treatment of congestive heart failure. Ann. West. Med. Surg. 1951, 5, 856–862. [Google Scholar]
- Higgins, A.R.; Harper, H.A.; Kline, E.F.; Merrill, R.S.; Jones, R.E.; Smith, T.W.D.; Kimmel, J.R. Effects of creatine precursors in arthritis; clinical and metabolic study of glycocyamine and betaine. Calif. Med. 1952, 77, 14–18. [Google Scholar] [PubMed]
- Borsook, M.E.; Billig, H.K.; Golseth, J.G. Betaine and glycocyamine in the treatment of disability resulting from acute anterior poliomyelitis. Ann. West. Med. Surg. 1952, 6, 423–427. [Google Scholar]
- Fallis, B.D.; Lam, R.L. Betaine and glycocyamine therapy for the chronic residuals of poliomyelitis. J. Am. Med. Assoc. 1952, 150, 851–853. [Google Scholar] [CrossRef] [PubMed]
- Watkins, A.L. Betaine and Glycocyamine in Treatment of Poliomyelitis. N. Engl. J. Med. 1953, 248, 621–623. [Google Scholar] [CrossRef] [PubMed]
- Basom, W.C.; Breck, L.W.; Leonard, M.H. The effect of betaine and glycocyamine in the management of chronic anterior polio-myelitis. Int. Rec. Med. Gen. Pract. Clin. 1955, 168, 70–71. [Google Scholar]
- Benassi, P. Effects of guanidinoacetic acid on the metabolism of creatine and creatinine in myopathic muscular dystrophy. Boll. Soc. Ital. Biol. Sper. 1954, 30, 365–368. [Google Scholar]
- Dixon, H.H.; Dickel, H.A.; Shanklin, J.G.; Peterson, R.D.; West, E.S. Therapy in anxiety states and anxiety complicated by depression. West. J. Surg. Obstet. Gynecol. 1954, 62, 338–341. [Google Scholar]
- Mune, N. Betasyamine in therapy of coronary arteriosclerosis. Ugeskr. Laeger 1954, 116, 1505. [Google Scholar]
- Billig, H.E., Jr.; Morehouse, L.E. Performance and metabolic alterations during betaine glycocyamine feeding in myasthenia gravis. Arch. Phys. Med. Rehabil. 1955, 36, 233–236. [Google Scholar]
- Liveksedge, L. Glycocyamine and betaine in motor-neurone disease. Lancet 1956, 268, 1136–1138. [Google Scholar] [CrossRef]
- Aldes, J.H. Glycocyamine betaine as an adjunct in the treatment of neuromuscular disease patients. J. Ark. Med. Soc. 1957, 54, 186–194. [Google Scholar]
- Tsubakihara, Y.; Suzuki, A.; Hayashi, T.; Shoji, T.; Togawa, M.; Okada, N. The effect of guanidinoacetic acid supplementation in patients with chronic renal failure. In Guanidino Compounds in Biology and Medicine; Mori, A., Ishida, M., Clark, J.F., Eds.; Blackwell Science: Japan, Tokyo, 1999; Volume 5, pp. 139–144. [Google Scholar]
- Ostojic, S.M.; Niess, B.; Stojanovic, M.; Obrenovic, M. Creatine Metabolism and Safety Profiles after Six-Week Oral Guanidinoacetic Acid Administration in Healthy Humans. Int. J. Med. Sci. 2013, 10, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Ostojic, S.M.; Niess, B.; Stojanovic, M.; Obrenovic, M. Co-administration of methyl donors along with guanidinoacetic acid reduces the incidence of hyperhomocysteinaemia compared with guanidinoacetic acid administration alone. Br. J. Nutr. 2013, 110, 865–870. [Google Scholar] [CrossRef] [Green Version]
- Semeredi, S.; Stajer, V.; Ostojic, J.; Vranes, M.; Ostojic, S.M. Guanidinoacetic acid with creatine compared with creatine alone for tissue creatine content, hyperhomocysteinemia, and exercise performance: A randomized, double-blind superiority trial. Nutrition 2019, 57, 162–166. [Google Scholar] [CrossRef] [Green Version]
- Ostojic, S.M.; Stojanovic, M.; Drid, P.; Hoffman, J. Dose–response effects of oral guanidinoacetic acid on serum creatine, homocysteine and B vitamins levels. Eur. J. Nutr. 2014, 53, 1637–1643. [Google Scholar] [CrossRef] [PubMed]
- Ostojic, S.M.; Vojvodic-Ostojic, A. Single-dose oral guanidinoacetic acid exhibits dose-dependent pharmacokinetics in healthy volunteers. Nutr. Res. 2015, 35, 198–205. [Google Scholar] [CrossRef]
- Ostojic, S.M.; Stojanovic, M. Guanidinoacetic acid loading affects plasma γ-aminobutyric acid in healthy men. Eur. J. Nutr. 2015, 54, 855–858. [Google Scholar] [CrossRef]
- Ostojic, S.; Stojanovic, M.D.; Hoffman, J. Six-Week Oral Guanidinoacetic Acid Administration Improves Muscular Performance in Healthy Volunteers. J. Investig. Med. 2015, 63, 942–946. [Google Scholar] [CrossRef] [PubMed]
- Ostojic, S.M.; Stojanovic, M.D.; Olcina, G. Oxidant-Antioxidant Capacity of Dietary Guanidinoacetic Acid. Ann. Nutr. Metab. 2015, 67, 243–246. [Google Scholar] [CrossRef]
- Ostojic, S.; Drid, P.; Ostojic, J. Guanidinoacetic acid increases skeletal muscle creatine stores in healthy men. Nutrition 2016, 32, 723–724. [Google Scholar] [CrossRef]
- Ostojic, S.M.; Ostojic, J.; Drid, P.; Vranes, M.; Jovanov, P. Dietary guanidinoacetic acid increases brain creatine levels in healthy men. Nutrition 2017, 33, 149–156. [Google Scholar] [CrossRef]
- Ostojic, S.M.; Mojsin, M.; Drid, P.; Vranes, M. Does Dietary Provision of Guanidinoacetic Acid Induce Global DNA Hypomethylation in Healthy Men and Women? Lifestyle Genom. 2018, 11, 16–18. [Google Scholar] [CrossRef]
- Ostojic, S.M.; Stojanovic, M.; Drid, P.; Hoffman, J.R.; Sekulic, D.; Zenic, N. Supplementation with Guanidinoacetic Acid in Women with Chronic Fatigue Syndrome. Nutrients 2016, 8, 72. [Google Scholar] [CrossRef]
- Seper, V.; Korovljev, D.; Todorovic, N.; Stajer, V.; Ostojic, J.; Nesic, N.; Ostojic, S.M. Guanidinoacetate-Creatine Supplementation Improves Functional Performance and Muscle and Brain Bioenergetics in the Elderly: A Pilot Study. Ann. Nutr. Metab. 2021, 77, 244–247. [Google Scholar] [CrossRef]
- Ostojic, S.M. Benefits and drawbacks of guanidinoacetic acid as a possible treatment to replenish cerebral creatine in AGAT deficiency. Nutr. Neurosci. 2019, 22, 302–305. [Google Scholar] [CrossRef] [PubMed]
- Ostojic, S.M.; Premusz, V.; Nagy, D.; Acs, P. Guanidinoacetic acid as a novel food for skeletal muscle health. J. Funct. Foods 2020, 73, 104129. [Google Scholar] [CrossRef]
- Ostojic, S.M.; Ostojic, J.; Drid, P.; Vranes, M. Guanidinoacetic acid versus creatine for improved brain and muscle creatine levels: A superiority pilot trial in healthy men. Appl. Physiol. Nutr. Metab. 2016, 41, 1005–1007. [Google Scholar] [CrossRef] [PubMed]
- Ostojic, S.M. Short-term GAA loading: Responders versus nonresponders analysis. Food Sci. Nutr. 2020, 8, 4446–4448. [Google Scholar] [CrossRef]
- Obeid, R. The Metabolic Burden of Methyl Donor Deficiency with Focus on the Betaine Homocysteine Methyltransferase Pathway. Nutrients 2013, 5, 3481–3495. [Google Scholar] [CrossRef] [Green Version]
- Ganguly, P.; Alam, S.F. Role of homocysteine in the development of cardiovascular disease. Nutr. J. 2015, 14, 6. [Google Scholar] [CrossRef] [Green Version]
- Ostojic, S.M.; Trivic, T.; Drid, P.; Stajer, V.; Vranes, M. Effects of Guanidinoacetic Acid Loading on Biomarkers of Cardiometabolic Risk and Inflammation. Ann. Nutr. Metab. 2018, 72, 18–20. [Google Scholar] [CrossRef]
- Ostojic, S.M.; Todorovic, N.; Stajer, V. Effect of Creatine and Guanidinoacetate Supplementation on Plasma Homocysteine in Metabolically Healthy Men and Women. Ann. Nutr. Metab. 2021, 77, 307–308. [Google Scholar] [CrossRef]
- Zugno, A.I.; Stefanello, F.M.; Streck, E.L.; Calcagnotto, T.; Wannmacher, C.M.; Wajner, M.; Wyse, A.T. Inhibition of Na+, K+ -ATPase activity in rat striatum by guanidinoacetate. Int. J. Dev. Neurosci. 2003, 21, 183–189. [Google Scholar] [CrossRef]
- Zugno, A.I.; Oliveira, D.L.; Scherer, E.B.S.; Wajner, M.; Wofchuk, S.; Wyse, A.T.S. Guanidinoacetate Inhibits Glutamate Uptake in Rat Striatum of Rats at Different Ages. Neurochem. Res. 2007, 32, 959–964. [Google Scholar] [CrossRef]
- Zugno, A.I.; Stefanello, F.M.; Scherer, E.B.S.; Mattos, C.; Pederzolli, C.D.; Andrade, V.M.; Wannmacher, C.M.D.; Wajner, M.; Dutra-Filho, C.S.; Wyse, A.T.S. Guanidinoacetate Decreases Antioxidant Defenses and Total Protein Sulfhydryl Content in Striatum of Rats. Neurochem. Res. 2008, 33, 1804–1810. [Google Scholar] [CrossRef] [PubMed]
- Hanna-El-Daher, L.; Béard, E.; Henry, H.; Tenenbaum, L.; Braissant, O. Mild guanidinoacetate increase under partial guanidinoacetate methyltransferase deficiency strongly affects brain cell development. Neurobiol. Dis. 2015, 79, 14–27. [Google Scholar] [CrossRef] [Green Version]
- Neuab, A.; Neuhoffa, H.; Trubec, G.; Fehra, S.; Ullrichb, K.; Roeper, J.; Isbrandta, D. Activation of GABAA Receptors by Guanidinoacetate: A Novel Pathophysiological Mechanism. Neurobiol. Dis. 2002, 11, 298–307. [Google Scholar] [CrossRef] [Green Version]
- Stromberger, C.; Bodamer, O.A.; Stöckler-Ipsiroglu, S. Clinical characteristics and diagnostic clues in inborn errors of creatine metabolism. J. Inherit. Metab. Dis. 2003, 26, 299–308. [Google Scholar] [CrossRef]
- Almeida, L.S.; Verhoeven, N.M.; Roos, B.; Valongo, C.; Cardoso, M.L.; Vilarinho, L.; Salomons, G.S.; Jakobs, C. Creatine and guanidinoacetate: Diagnostic markers for inborn errors in creatine biosynthesis and transport. Mol. Genet. Metab. 2004, 82, 214–219. [Google Scholar] [CrossRef]
- Ostojic, S.M.; Ostojic, J. Dietary guanidinoacetic acid does not accumulate in the brain of healthy men. Eur. J. Nutr. 2018, 57, 3003–3005. [Google Scholar] [CrossRef] [PubMed]
- Kharbanda, K.K.; Todero, S.L.; Thomes, P.G.; Orlicky, D.J.; Osna, N.A.; French, S.W.; Tuma, D.J. Increased methylation demand exacerbates ethanol-induced liver injury. Exp. Mol. Pathol. 2014, 97, 49–56. [Google Scholar] [CrossRef]
- Osna, N.A.; Feng, D.; Ganesan, M.; Maillacheruvu, P.F.; Orlicky, D.J.; French, S.W.; Tuma, D.J.; Kharbanda, K.K. Prolonged feeding with guanidinoacetate, a methyl group consumer, exacerbates ethanol-induced liver injury. World J. Gastroenterol. 2016, 22, 8497–8508. [Google Scholar] [CrossRef]
- Schepers, E.; Glorieux, G.; Dou, L.; Cerini, C.; Gayrard, N.; Louvet, L.; Maugard, C.; Preus, P.; Rodriguez-Ortiz, M.; Argiles, A.; et al. Guanidino Compounds as Cause of Cardiovascular Damage in Chronic Kidney Disease: An in vitro Evaluation. Blood Purif. 2010, 30, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Mori, A.; Kohno, M.; Masumizu, T.; Noda, Y.; Packer, L. Guanidino compounds generate reactive oxygen species. IUBMB Life 1996, 40, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Arai, B.; Koshino, C. Effects of guanidinoacetic acid, γ-guanidinobutyric acid and γ-guanidinobutyrylmethylester on the mammalian cerebral cortex. Jpn. J. Physiol. 1961, 11, 403–409. [Google Scholar] [CrossRef] [Green Version]
- Van Der Voet, H.; De Mul, A.; Van Klaveren, J.D. A probabilistic model for simultaneous exposure to multiple compounds from food and its use for risk–benefit assessment. Food Chem. Toxicol. 2007, 45, 1496–1506. [Google Scholar] [CrossRef]
- Ruzante, J.M.; Grieger, K.; Woodward, K.; Lambertini, E.; Kowalcyk, B. The use of multi-criteria decision analysis in food safety risk-benefit assessment. Food Protect Trends 2017, 37, 132–139. [Google Scholar]
- Stratil, J.M.; Baltussen, R.; Scheel, I.; Nacken, A.; Rehfuess, E.A. Development of the WHO-INTEGRATE evidence-to-decision framework: An overview of systematic reviews of decision criteria for health decision-making. Cost Eff. Resour. Alloc. 2020, 18, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Schug, S.; Pogatzki-Zahn, E.; Phillips, L.D.; Essex, M.N.; Xia, F.; Reader, A.J.; Pawinski, R. Multi-Criteria Decision Analysis to Develop an Efficacy-Safety Profile of Parenteral Analgesics Used in the Treatment of Postoperative Pain. J. Pain Res. 2020, 13, 1969–1977. [Google Scholar] [CrossRef]
- Durbach, I.N.; Stewart, T.J. Modeling uncertainty in multi-criteria decision analysis. Eur. J. Oper. Res. 2012, 223, 1–14. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ostojic, S.M. Safety of Dietary Guanidinoacetic Acid: A Villain of a Good Guy? Nutrients 2022, 14, 75. https://doi.org/10.3390/nu14010075
Ostojic SM. Safety of Dietary Guanidinoacetic Acid: A Villain of a Good Guy? Nutrients. 2022; 14(1):75. https://doi.org/10.3390/nu14010075
Chicago/Turabian StyleOstojic, Sergej M. 2022. "Safety of Dietary Guanidinoacetic Acid: A Villain of a Good Guy?" Nutrients 14, no. 1: 75. https://doi.org/10.3390/nu14010075
APA StyleOstojic, S. M. (2022). Safety of Dietary Guanidinoacetic Acid: A Villain of a Good Guy? Nutrients, 14(1), 75. https://doi.org/10.3390/nu14010075