
Maternal Vegetable and Fruit Consumption during Pregnancy and Its Effects on Infant Gut Microbiome

 , , , , , ,
, , , , , ,  ,
, 
Abstract
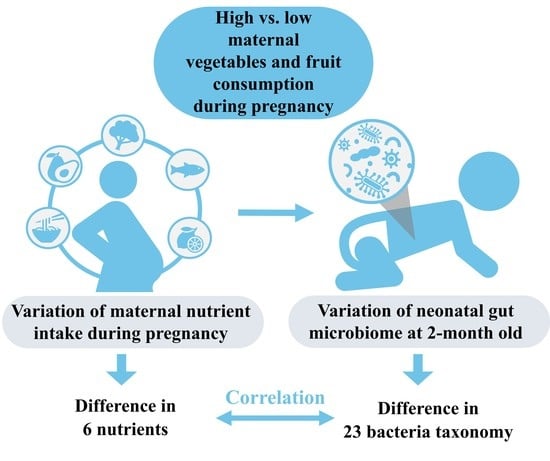
1. Introduction
2. Materials and Methods
2.1. Cohort Establishment and Data Collection
2.2. Maternal Dietary Assessment
2.3. Sample Collection and DNA Extraction
2.4. Targeted 16S rRNA Gene Sequencing
2.5. Confounding Factors
2.6. Data Analysis
3. Results
3.1. Demographic Characteristics
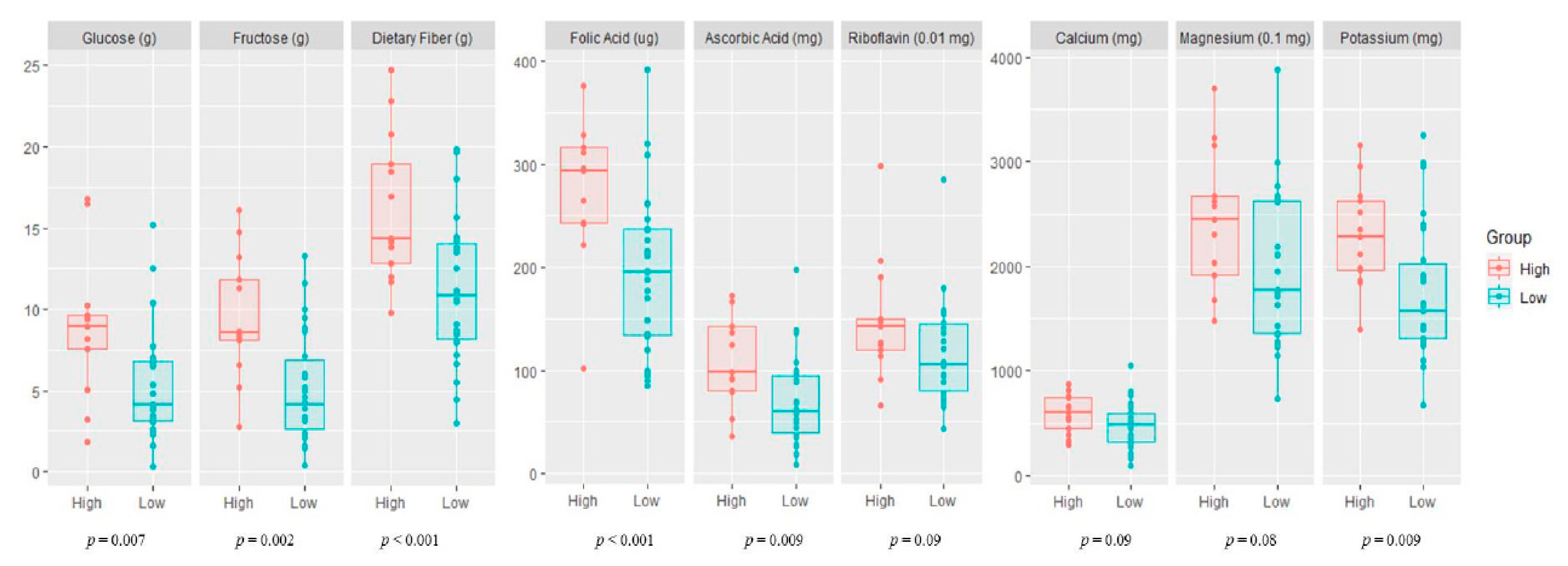
3.2. Variation of Maternal Nutrient Intake
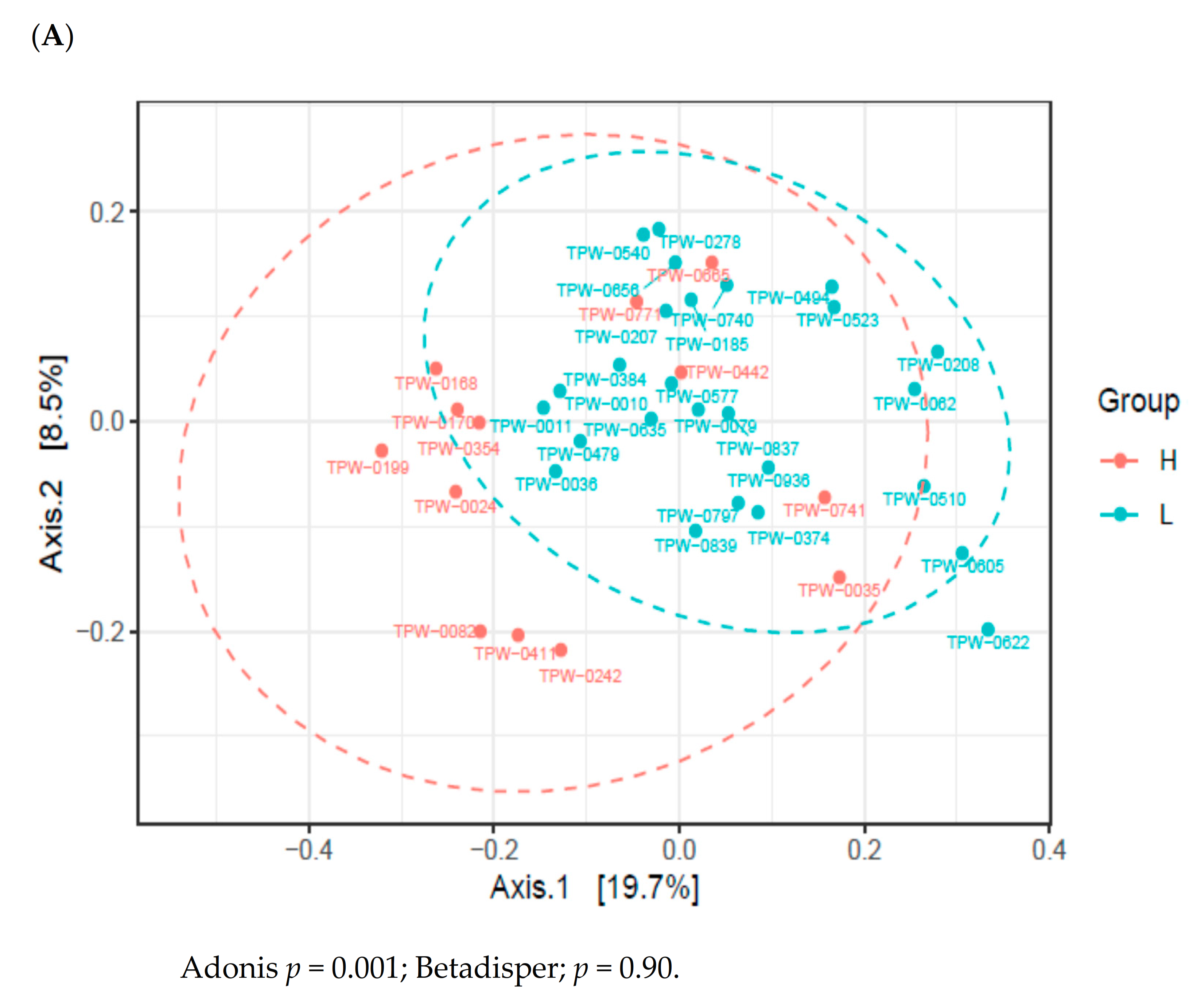
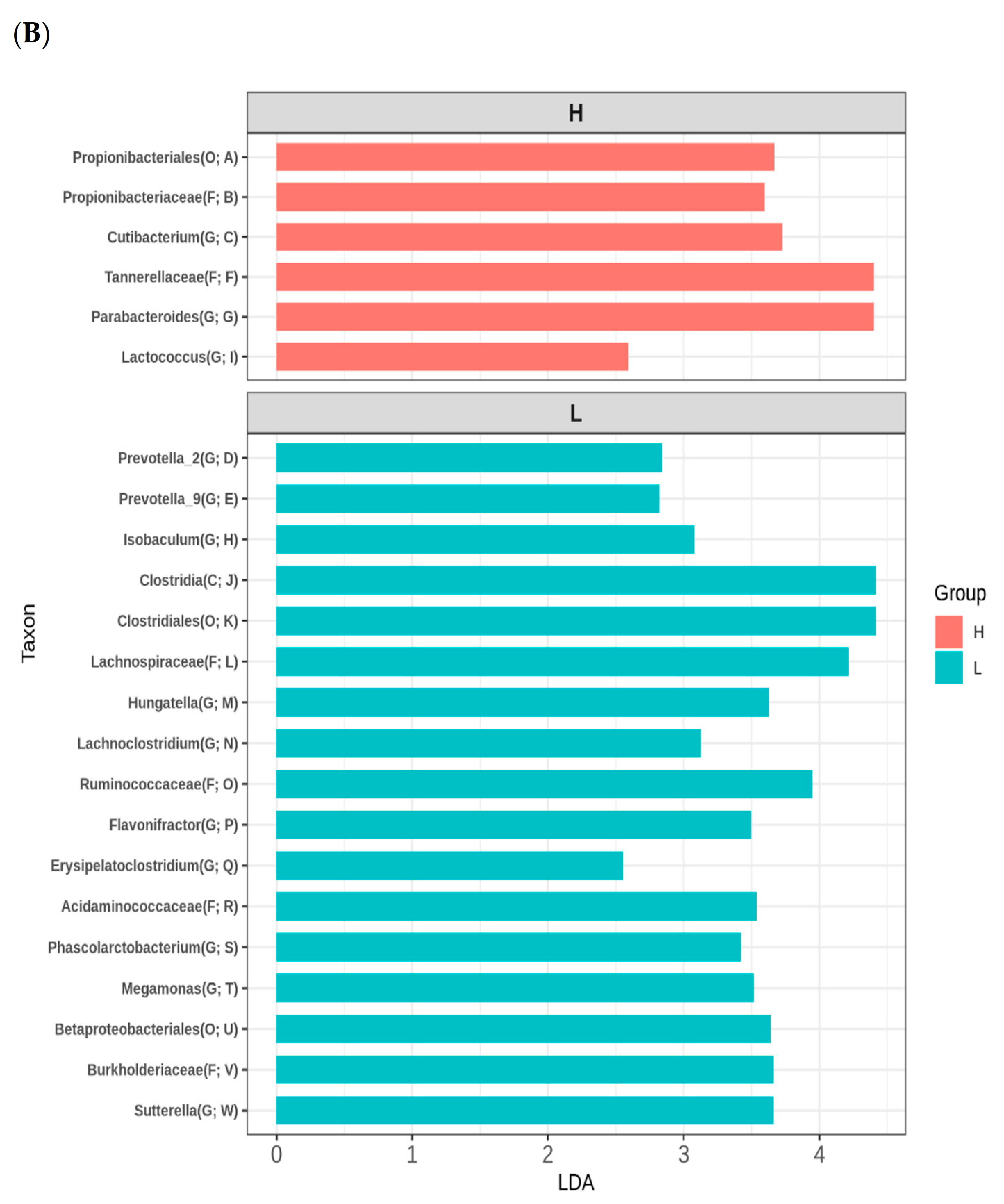
3.3. Variation of Infant Gut Microbiome
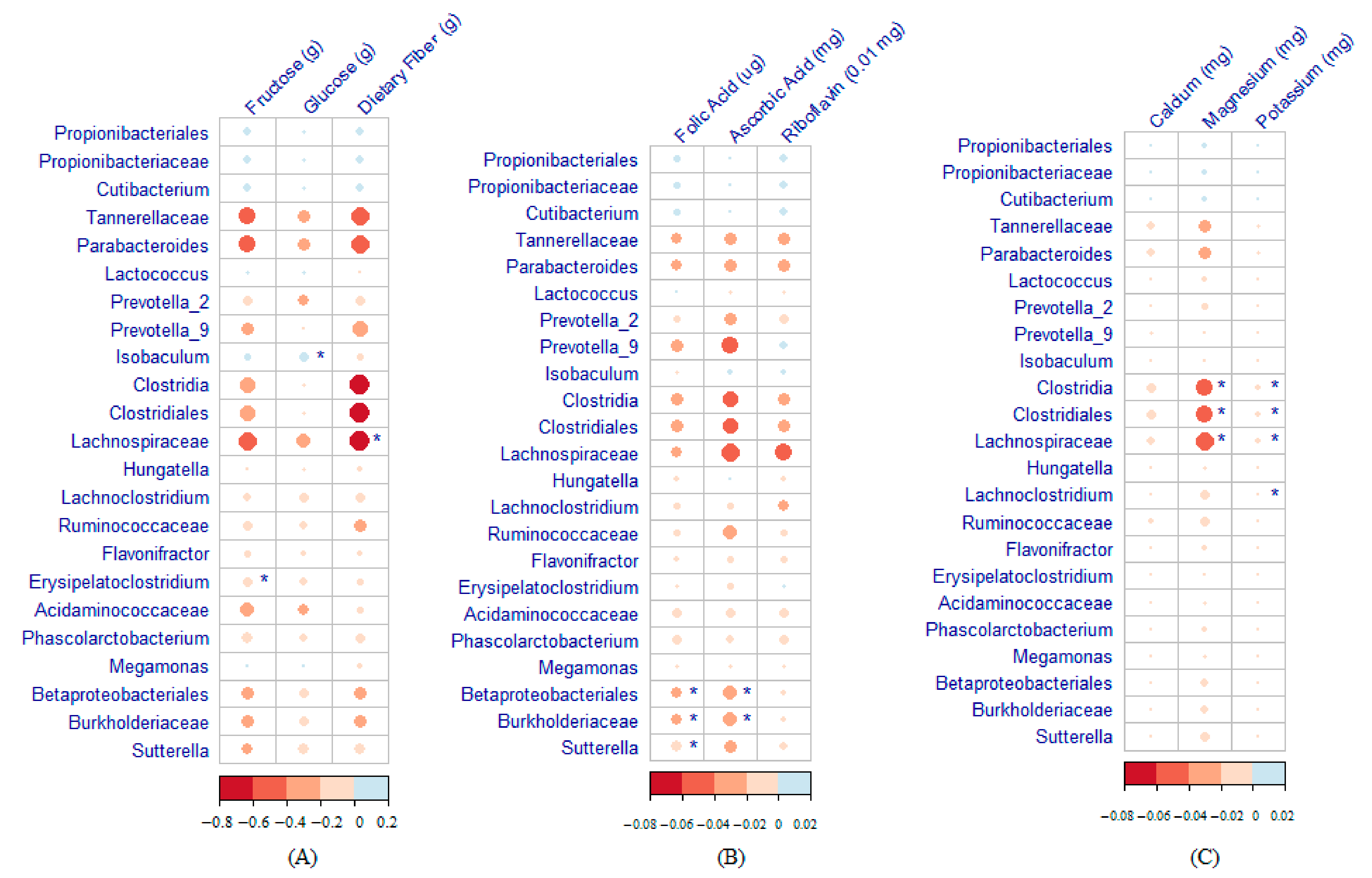
3.4. Maternal Nutrient Intake and Infant Gut Microbiome
4. Discussion
4.1. Maternal Fruit and Vegetable Consumption Affects Infant Microbiome
4.2. Maternal Difference in Six Nutrients and 23 Bacterial Taxa
4.3. Strengths and Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Barker, D.J. The fetal and infant origins of adult disease. BMJ 1990, 301, 1111. [Google Scholar] [CrossRef] [PubMed]
- Netting, M.J.; Middleton, P.F.; Makrides, M. Does maternal diet during pregnancy and lactation affect outcomes in offspring? A systematic review of food-based approaches. Nutrients 2014, 30, 1225–1241. [Google Scholar] [CrossRef]
- Martin, C.L.; Siega-Riz, A.M.; Sotres-Alvarez, D.; Robinson, W.R.; Daniels, J.L.; Perrin, E.M.; Stuebe, A.M. Maternal Dietary Patterns during Pregnancy Are Associated with Child Growth in the First 3 Years of Life. J. Nutr. 2016, 146, 2281–2288. [Google Scholar] [CrossRef]
- Chu, D.M.; Meyer, K.M.; Prince, A.L.; Aagaard, K.M. Impact of maternal nutrition in pregnancy and lactation on offspring gut microbial composition and function. Gut Microbes 2016, 7, 459–470. [Google Scholar] [CrossRef]
- Ma, J.; Prince, A.L.; Bader, D.; Hu, M.; Ganu, R.; Baquero, K.; Blundell, P.; Harris, R.A.; Frias, A.E.; Grove, K.L.; et al. High-fat maternal diet during pregnancy persistently alters the offspring microbiome in a primate model. Nat. Commun. 2014, 5, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Chu, D.M.; Antony, K.M.; Ma, J.; Prince, A.L.; Showalter, L.; Moller, M.; Aagaard, K.M. The early infant gut microbiome varies in association with a maternal high-fat diet. Genome Med. 2016, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Myles, I.A.; Fontecilla, N.M.; Janelsins, B.M.; Vithayathil, P.J.; Segre, J.A.; Datta, S.K. Parental Dietary Fat Intake Alters Offspring Microbiome and Immunity. J. Immunol. 2013, 191, 3200–3209. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, H.; Zhang, L.; Yang, Y.; Lin, Y.; Zhuo, Y.; Fang, Z.; Che, L.; Feng, B.; Xu, S.; et al. Maternal Dietary Fiber Composition during Gestation Induces Changes in Offspring Antioxidative Capacity, Inflammatory Response, and Gut Microbiota in a Sow Model. Int. J. Mol. Sci. 2019, 21, 31. [Google Scholar] [CrossRef] [PubMed]
- Lundgren, S.N.; Madan, J.C.; Emond, J.A.; Morrison, H.G.; Christensen, B.C.; Karagas, M.R.; Hoen, A.G. Maternal diet during pregnancy is related with the infant stool microbiome in a delivery mode-dependent manner. Microbiome 2018, 6, 1–11. [Google Scholar] [CrossRef]
- Yang, Q.; Liang, Q.; Balakrishnan, B.; Belobrajdic, D.P.; Feng, Q.-J.; Zhang, W. Role of Dietary Nutrients in the Modulation of Gut Microbiota: A Narrative Review. Nutrients 2020, 12, 381. [Google Scholar] [CrossRef]
- Thorburn, A.N.; McKenzie, C.I.; Shen, S.; Stanley, D.; Macia, L.; Mason, L.J.; Roberts, L.K.; Wong, C.H.Y.; Shim, R.; Robert, R.; et al. Evidence that asthma is a developmental origin disease influenced by maternal diet and bacterial metabolites. Nat. Commun. 2015, 6, 7320. [Google Scholar] [CrossRef]
- Li, L.; Krause, L.; Somerset, S. Associations between micronutrient intakes and gut microbiota in a group of adults with cystic fibrosis. Clin. Nutr. 2017, 36, 1097–1104. [Google Scholar] [CrossRef]
- Astbury, S.; Song, A.; Zhou, M.; Nielsen, B.; Hoedl, A.; Willing, B.P.; Symonds, M.E.; Bell, R.C. High Fructose Intake During Pregnancy in Rats Influences the Maternal Microbiome and Gut Development in the Offspring. Front. Genet. 2018, 9, 203. [Google Scholar] [CrossRef] [PubMed]
- Dunn, A.B.; Jordan, S.; Baker, B.J.; Carlson, N.S. The Maternal Infant Microbiome: Considerations for Labor and Birth. MCN Am. J. Matern. Child Nurs. 2017, 42, 318–325. [Google Scholar] [CrossRef]
- Wang, S.; Ryan, C.A.; Boyaval, P.; Dempsey, E.M.; Ross, R.P.; Stanton, C. Maternal Vertical Transmission Affecting Early-life Microbiota Development. Trends Microbiol. 2020, 28, 28–45. [Google Scholar] [CrossRef]
- Wang, J.-S.; Hsieh, R.-H.; Tung, Y.-T.; Chen, Y.-H.; Yang, C.; Chen, Y.C. Evaluation of a Technological Image-Based Dietary Assessment Tool for Children during Pubertal Growth: A Pilot Study. Nutrients 2019, 11, 2527. [Google Scholar] [CrossRef]
- Carruthers, L.V.; Moses, A.; Adriko, M.; Faust, C.L.; Tukahebwa, E.M.; Hall, L.J.; Ranford-Cartwright, L.C.; Lamberton, P.H. The impact of storage conditions on human stool 16S rRNA microbiome composition and diversity. PeerJ 2019, 7, e8133. [Google Scholar] [CrossRef]
- Yang, Y.-C.S.H.; Chang, H.-W.; Lin, I.H.; Chien, L.-N.; Wu, M.-J.; Liu, Y.-R.; Chu, P.G.; Xie, G.; Dong, F.; Jia, W.; et al. Long-term Proton Pump Inhibitor Administration Caused Physiological and Microbiota Changes in Rats. Sci. Rep. 2020, 10, 866. [Google Scholar] [CrossRef]
- Callahan, B.J.; Sankaran, K.; Fukuyama, J.A.; McMurdie, P.J.; Holmes, S.P. Bioconductor Workflow for Microbiome Data Analysis: From raw reads to community analyses. F1000Research 2016, 5, 1492. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Schliep, K.P. phangorn: Phylogenetic analysis in R. Bioinformatics 2011, 27, 592–593. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Bittinger, K.; Charlson, E.S.; Hoffmann, C.; Lewis, J.; Wu, G.D.; Collman, R.G.; Bushman, F.D.; Li, H. Associating microbiome composition with environmental covariates using generalized UniFrac distances. Bioinformatics 2012, 28, 2106–2113. [Google Scholar] [CrossRef] [PubMed]
- Asnicar, F.; Weingart, G.; Tickle, T.L.; Huttenhower, C.; Segata, N. Compact graphical representation of phylogenetic data and metadata with GraPhlAn. PeerJ 2015, 3, e1029. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Nakajima, A.; Kaga, N.; Nakanishi, Y.; Ohno, H.; Miyamoto, J.; Kimura, I.; Hori, S.; Sasaki, T.; Hiramatsu, K.; Okumura, K.; et al. Maternal High Fiber Diet during Pregnancy and Lactation Influences Regulatory T Cell Differentiation in Offspring in Mice. J. Immunol. 2017, 199, 3516–3524. [Google Scholar] [CrossRef]
- Maslowski, K.M.; Mackay, C.R. Diet, gut microbiota and immune responses. Nat. Immunol. 2010, 12, 5–9. [Google Scholar] [CrossRef]
- Trompette, A.; Gollwitzer, E.S.; Yadava, K.; Sichelstiel, A.K.; Sprenger, N.; Ngom-Bru, C.; Blanchard, C.; Junt, T.; Nicod, L.P.; Harris, N.L.; et al. Gut microbiota metabolism of dietary fiber influences allergic airway disease and hematopoiesis. Nat. Med. 2014, 20, 159–166. [Google Scholar] [CrossRef]
- Wood, H.G. Metabolic Cycles in the Fermentation by Propionic Acid Bacteria. Curr. Top. Cell. Regul. 1981, 18, 255–287. [Google Scholar] [CrossRef]
- Clarke, J.M.; Topping, D.L.; Christophersen, C.T.; Bird, A.R.; Lange, K.; Saunders, I.; Cobiac, L. Butyrate esterified to starch is released in the human gastrointestinal tract. Am. J. Clin. Nutr. 2011, 94, 1276–1283. [Google Scholar] [CrossRef]
- Sánchez, E.; Donat, E.; Ribes-Koninckx, C.; Calabuig, M.; Sanz, Y. Intestinal Bacteroides species associated with coeliac disease. J. Clin. Pathol. 2010, 63, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Liao, M.; Zhou, N.; Bao, L.; Ma, K.; Zheng, Z.; Wang, Y.; Liu, C.; Wang, W.; Wang, J.; et al. Parabacteroides distasonis Alleviates Obesity and Metabolic Dysfunctions via Production of Succinate and Secondary Bile Acids. Cell Rep. 2019, 26, 222–235. [Google Scholar] [CrossRef] [PubMed]
- Kverka, M.; Zakostelska, Z.; Klimesova, K.; Sokol, D.; Hudcovic, T.; Hrncir, T.; Rossmann, P.; Mrazek, J.; Kopecny, J.; Verdu, E.F.; et al. Oral administration of Parabacteroides distasonis antigens attenuates experimental murine colitis through modulation of immunity and microbiota composition. Clin. Exp. Immunol. 2010, 163, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Xiang, Y.; Lou, F.; Tu, P.; Zhang, X.; Hu, X.; Lyu, W.; Xiao, Y. Microbial Community and Short-Chain Fatty Acid Mapping in the Intestinal Tract of Quail. Animals 2020, 10, 1006. [Google Scholar] [CrossRef]
- Beck, B.R.; Kim, D.; Jeon, J.; Lee, S.-M.; Kim, H.K.; Kim, O.-J.; Lee, J.I.; Suh, B.S.; Do, H.K.; Lee, K.H.; et al. The effects of combined dietary probiotics Lactococcus lactis BFE920 and Lactobacillus plantarum FGL0001 on innate immunity and disease resistance in olive flounder (Paralichthys olivaceus). Fish Shellfish Immunol. 2015, 42, 177–183. [Google Scholar] [CrossRef]
- Kim, D.; Beck, B.R.; Heo, S.-B.; Kim, J.; Kim, H.D.; Lee, S.-M.; Kim, Y.; Oh, S.Y.; Lee, K.; Do, H.; et al. Lactococcus lactis BFE920 activates the innate immune system of olive flounder (Paralichthys olivaceus), resulting in protection against Streptococcus iniae infection and enhancing feed efficiency and weight gain in large-scale field studies. Fish Shellfish Immunol. 2013, 35, 1585–1590. [Google Scholar] [CrossRef]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef]
- Arpaia, N.; Campbell, C.; Fan, X.; Dikiy, S.; van der Veeken, J.; deRoos, P.; Liu, H.; Cross, J.R.; Pfeffer, K.; Coffer, P.J.; et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature 2013, 504, 451–455. [Google Scholar] [CrossRef]
- Smith, P.M.; Howitt, M.R.; Panikov, N.; Michaud, M.; Gallini, C.A.; Bohlooly-Y, M.; Glickman, J.N.; Garrett, W.S. The Microbial Metabolites, Short-Chain Fatty Acids, Regulate Colonic Treg Cell Homeostasis. Science 2013, 341, 569–573. [Google Scholar] [CrossRef]
- Gonzalez-Granda, A.; Beisner, J.; Basrai, M.; Damms-Machado, A.; Bischoff, S. Fructose-induced intestinal microbiota shift following two types of short-term high fructose dietary phases: A microbiota analysis of a crossover intervention study in healthy women. Nutrition 2019, 44, 24. [Google Scholar] [CrossRef]
- Do, M.H.; Lee, E.; Oh, M.-J.; Kim, Y.; Park, H.-Y. High-Glucose or -Fructose Diet Cause Changes of the Gut Microbiota and Metabolic Disorders in Mice without Body Weight Change. Nutrition 2018, 10, 761. [Google Scholar] [CrossRef]
- Gurwara, S.; Ajami, N.J.; Jang, A.; Hessel, F.C.; Chen, L.; Plew, S.; Wang, Z.; Graham, D.Y.; Hair, C.; White, D.L.; et al. Dietary Nutrients Involved in One-Carbon Metabolism and Colonic Mucosa-Associated Gut Microbiome in Individuals with an Endoscopically Normal Colon. Nutrition 2019, 11, 613. [Google Scholar] [CrossRef]
- Padilha, M.; Danneskiold-Samsøe, N.B.; Brejnrod, A.; Hoffmann, C.; Cabral, V.P.; Iaucci, J.D.M.; Sales, C.H.; Fisberg, R.M.; Cortez, R.V.; Brix, S.; et al. The Human Milk Microbiota is Modulated by Maternal Diet. Microorganisms 2019, 7, 502. [Google Scholar] [CrossRef]
- Hoppu, U.; Rinne, M.; Salo-Väänänen, P.; Lampi, A.-M.; Piironen, V.; Isolauri, E.; Salo-Väävnänen, P. Vitamin C in breast milk may reduce the risk of atopy in the infant. Eur. J. Clin. Nutr. 2004, 59, 123–128. [Google Scholar] [CrossRef]
- Shen, X.; Wang, M.; Zhang, X.; He, M.; Li, M.; Cheng, G.; Wan, C.; He, F. Dynamic construction of gut microbiota may influence allergic diseases of infants in Southwest China. BMC Microbiol. 2019, 19, 1–13. [Google Scholar] [CrossRef]
- Chua, H.-H.; Chou, H.-C.; Tung, Y.-L.; Chiang, B.-L.; Liao, C.-C.; Liu, H.-H.; Ni, Y.-H. Intestinal Dysbiosis Featuring Abundance of Ruminococcus gnavus Associates with Allergic Diseases in Infants. Gastroenterology 2018, 154, 154–167. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | High Consumption * (n = 13) | Low Consumption (n = 26) | Comparison | |||
|---|---|---|---|---|---|---|
| N or Mean | % or (SD) | N or Mean | % or (SD) | Statistics | p | |
| Maternal age at baseline | 34.2 | (2.6) | 33.5 | (4.5) | 0.64 | 0.53 |
| <30 | 0 | 0.0% | 5 | 19.2% | 2.87 | 0.32 |
| 30–35 | 7 | 53.8% | 11 | 42.3% | ||
| ≥35 | 6 | 46.2% | 10 | 38.5% | ||
| Maternal education level | ||||||
| Senior high school or below | 0 | 0.0% | 1 | 3.8% | 0.67 | 0.82 |
| College | 8 | 61.5% | 17 | 65.4% | ||
| Graduate school and higher | 5 | 38.5% | 8 | 30.8% | ||
| Family income | ||||||
| <60,000 | 5 | 38.4% | 10 | 38.4% | 2.14 | 0.32 |
| 60,000–100,000 | 5 | 38.4% | 5 | 19.2% | ||
| >100,000 | 3 | 23.1% | 11 | 42.3% | ||
| Maternal history of diseases | ||||||
| Cardiovascular disease | 1 | 7.6% | 0 | 0.0% | 0.12 | 0.72 |
| Gestational diabetes mellitus | 0 | 0.0% | 1 | 3.8% | 0.00 | 1.00 |
| Hyperthyroidism | 2 | 15.3% | 0 | 0.0% | 1.64 | 0.20 |
| Hypothyroidism | 1 | 7.6% | 0 | 0.0% | 0.12 | 0.72 |
| Timing of dietary assessment | ||||||
| <13 weeks | 0 | 0.0% | 8 | 30.8% | 5.12 | 0.08 |
| 13–28 weeks | 8 | 61.5% | 12 | 46.2% | ||
| >28 weeks | 5 | 38.5% | 6 | 23.1% | ||
| Gestational age | 38.7 | (1.3) | 38.1 | (1.2) | 1.20 | 0.24 |
| <37 weeks | 1 | 8.3% | 2 | 8.3% | 0.00 | 1.00 |
| Excess gestational weight gain | 2 | 15.4% | 4 | 15.4% | 0.00 | 1.00 |
| Normal spontaneous delivery | 9 | 69.2% | 17 | 65.4% | 0.00 | 1.00 |
| Antepartum antibiotics | 1 | 7.7% | 4 | 15.4% | 0.03 | 0.64 |
| Group B streptococcus positive | 2 | 15.4% | 5 | 19.2% | 0.00 | 1.00 |
| Neonatal sex (male) | 7 | 58.3% | 17 | 65.4% | 0.04 | 0.72 |
| Breastfeeding (yes) | 5 | 38.5% | 10 | 38.5% | 0.00 | 1.00 |
| Dietary intake | ||||||
| Calories (kcal) | 1679.3 | (354.4) | 1541.1 | (378.9) | 204.0 | 0.31 |
| Vegetables (cups †) | 2.1 | (0.7) | 1.8 | (1.5) | 228.0 | 0.08 |
| Fruit (cups †) | 1.9 | (0.5) | 0.6 | (0.6) | 317.0 | <0.001 |
| Dairy (cups †) | 0.7 | (0.5) | 0.6 | (0.8) | 202.5 | 0.32 |
| Grain (1/4 cups †) | 8.4 | (2.1) | 8.2 | (2.4) | 175.0 | 0.87 |
| Meat (1/6 cups †) | 5.9 | (2.4) | 6.0 | (3.3) | 185.0 | 0.65 |
| Fat (tbsp †) | 5.5 | (1.8) | 5.9 | (2.3) | 150.0 | 0.59 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, H.-Y.; Tung, Y.-T.; Yang, Y.-C.S.H.; Hsu, J.B.; Lee, C.-Y.; Chang, T.-H.; Su, E.C.-Y.; Hsieh, R.-H.; Chen, Y.-C. Maternal Vegetable and Fruit Consumption during Pregnancy and Its Effects on Infant Gut Microbiome. Nutrients 2021, 13, 1559. https://doi.org/10.3390/nu13051559
Fan H-Y, Tung Y-T, Yang Y-CSH, Hsu JB, Lee C-Y, Chang T-H, Su EC-Y, Hsieh R-H, Chen Y-C. Maternal Vegetable and Fruit Consumption during Pregnancy and Its Effects on Infant Gut Microbiome. Nutrients. 2021; 13(5):1559. https://doi.org/10.3390/nu13051559
Chicago/Turabian StyleFan, Hsien-Yu, Yu-Tang Tung, Yu-Chen S. H. Yang, Justin BoKai Hsu, Cheng-Yang Lee, Tzu-Hao Chang, Emily Chia-Yu Su, Rong-Hong Hsieh, and Yang-Ching Chen. 2021. "Maternal Vegetable and Fruit Consumption during Pregnancy and Its Effects on Infant Gut Microbiome" Nutrients 13, no. 5: 1559. https://doi.org/10.3390/nu13051559
APA StyleFan, H.-Y., Tung, Y.-T., Yang, Y.-C. S. H., Hsu, J. B., Lee, C.-Y., Chang, T.-H., Su, E. C.-Y., Hsieh, R.-H., & Chen, Y.-C. (2021). Maternal Vegetable and Fruit Consumption during Pregnancy and Its Effects on Infant Gut Microbiome. Nutrients, 13(5), 1559. https://doi.org/10.3390/nu13051559