
Obesity Development and Signs of Metabolic Abnormalities in Young Göttingen Minipigs Consuming Energy Dense Diets Varying in Carbohydrate Quality





Abstract
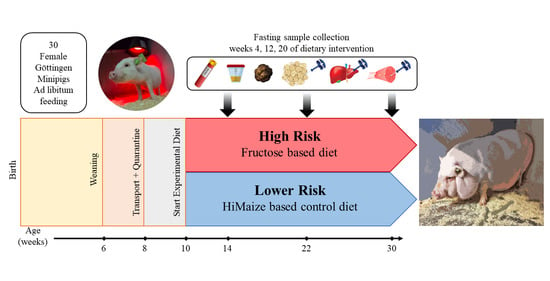
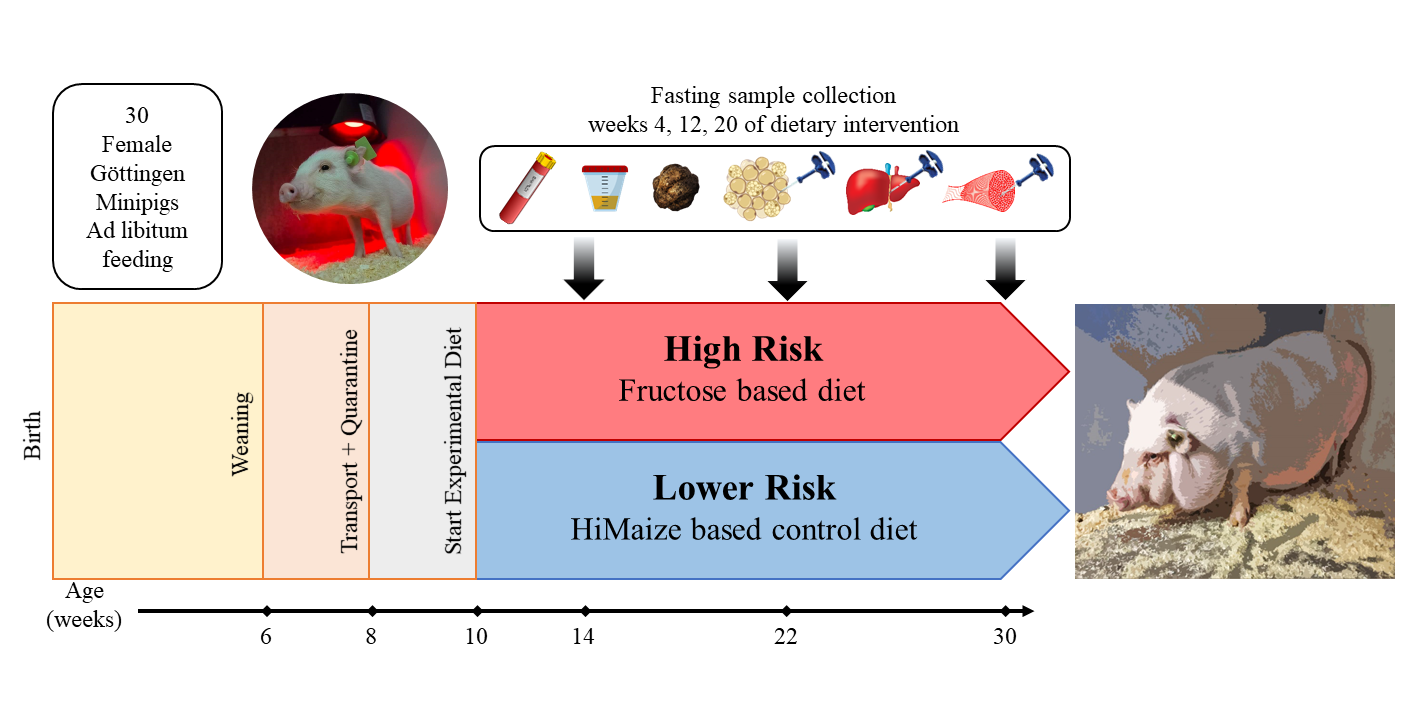
1. Introduction
2. Materials and Methods
2.1. Experimental Diets
2.2. Animals and Experimental Design
2.3. Sample Collection
2.4. Analytical Methods
2.5. Gene Expression Analysis of Liver, Muscle, and Subcutaneous Adipose Tissue (SAT) by Real-Time Reverse Transcriptase-Polymerase Chain Reaction (RT-PCR)
2.6. Calculations and Statistical Analyses
3. Results
3.1. Diet Composition
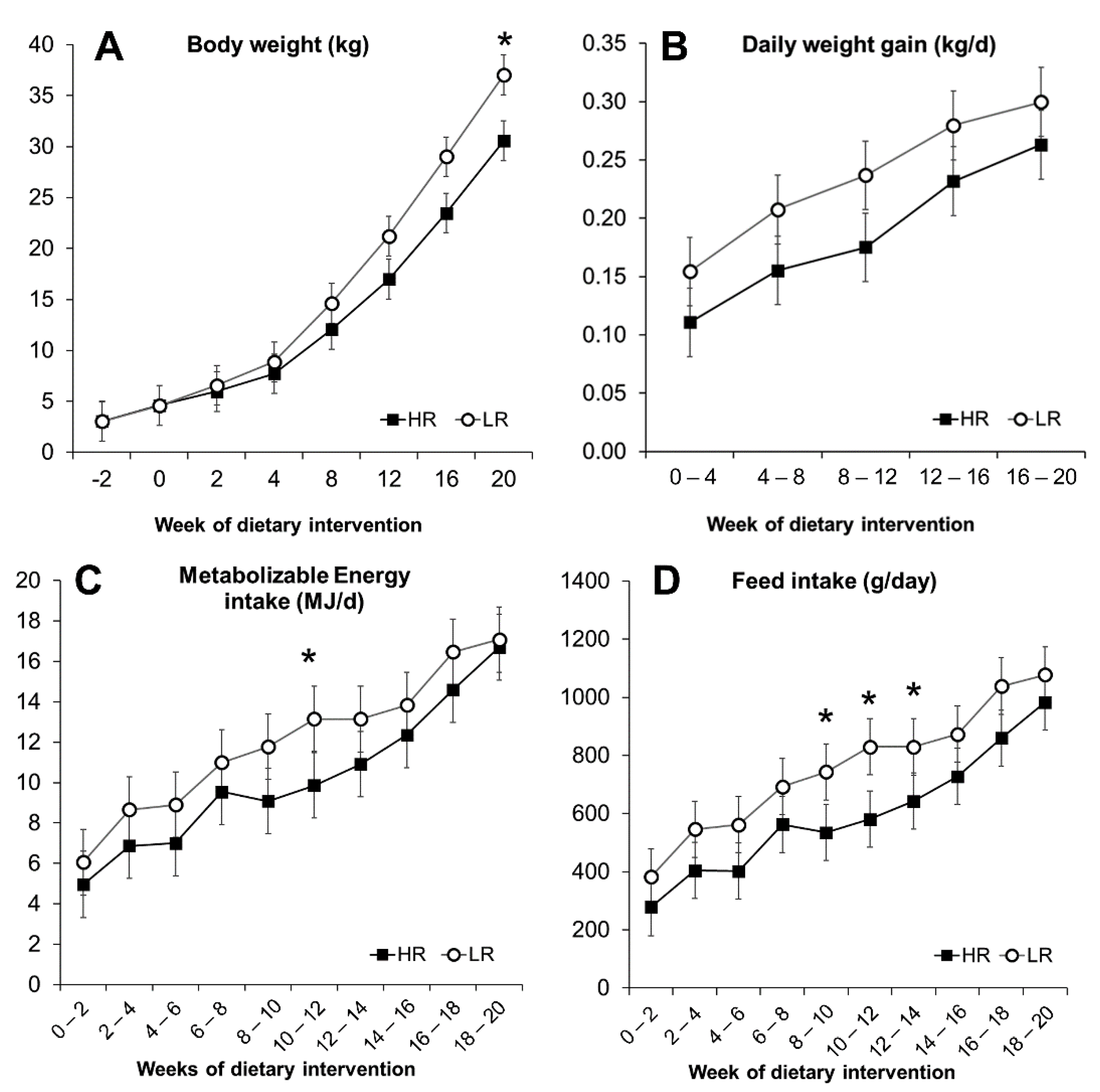
3.2. Nutrient and Energy Intake
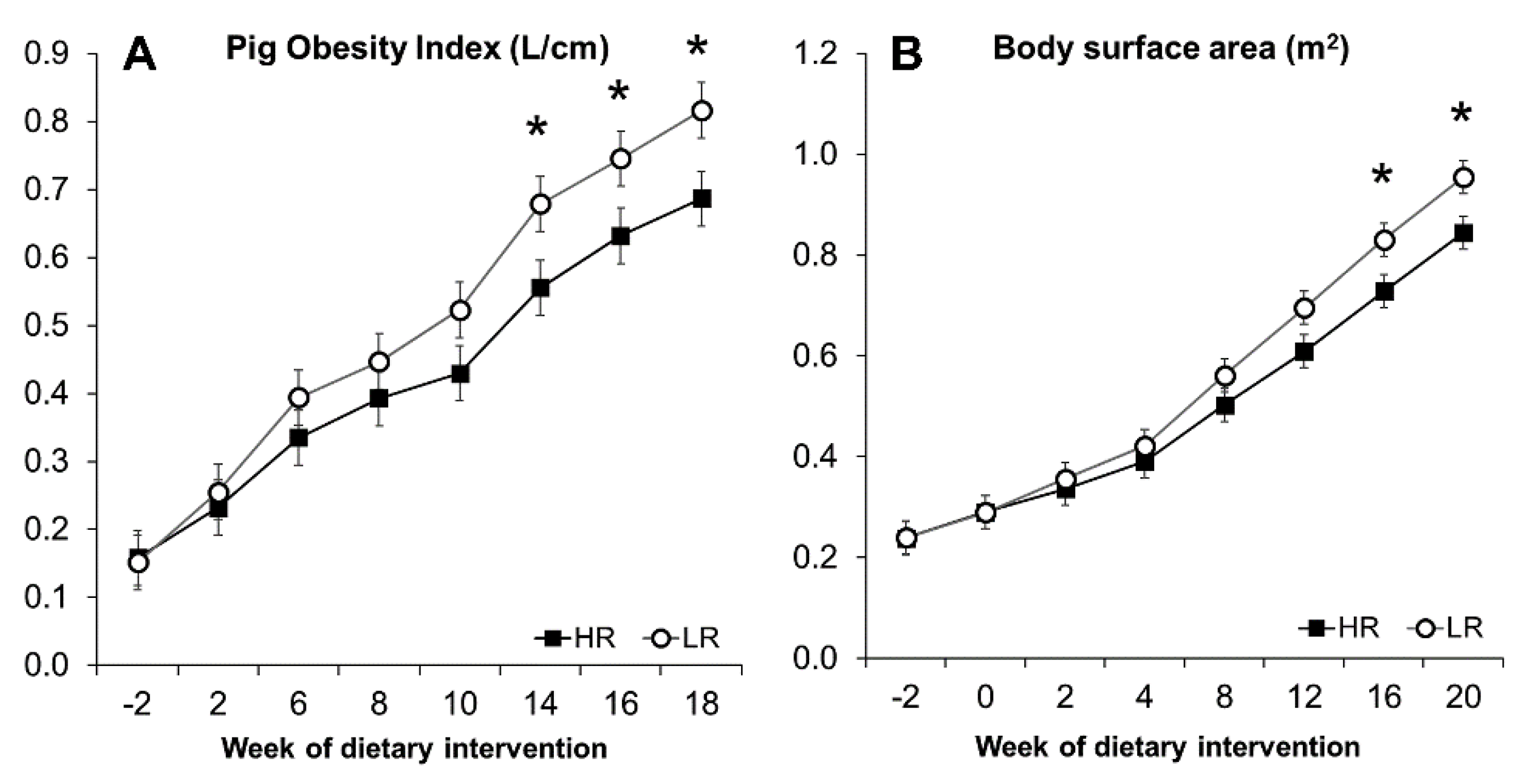
3.3. Obesity Development and Morphometric Measurements
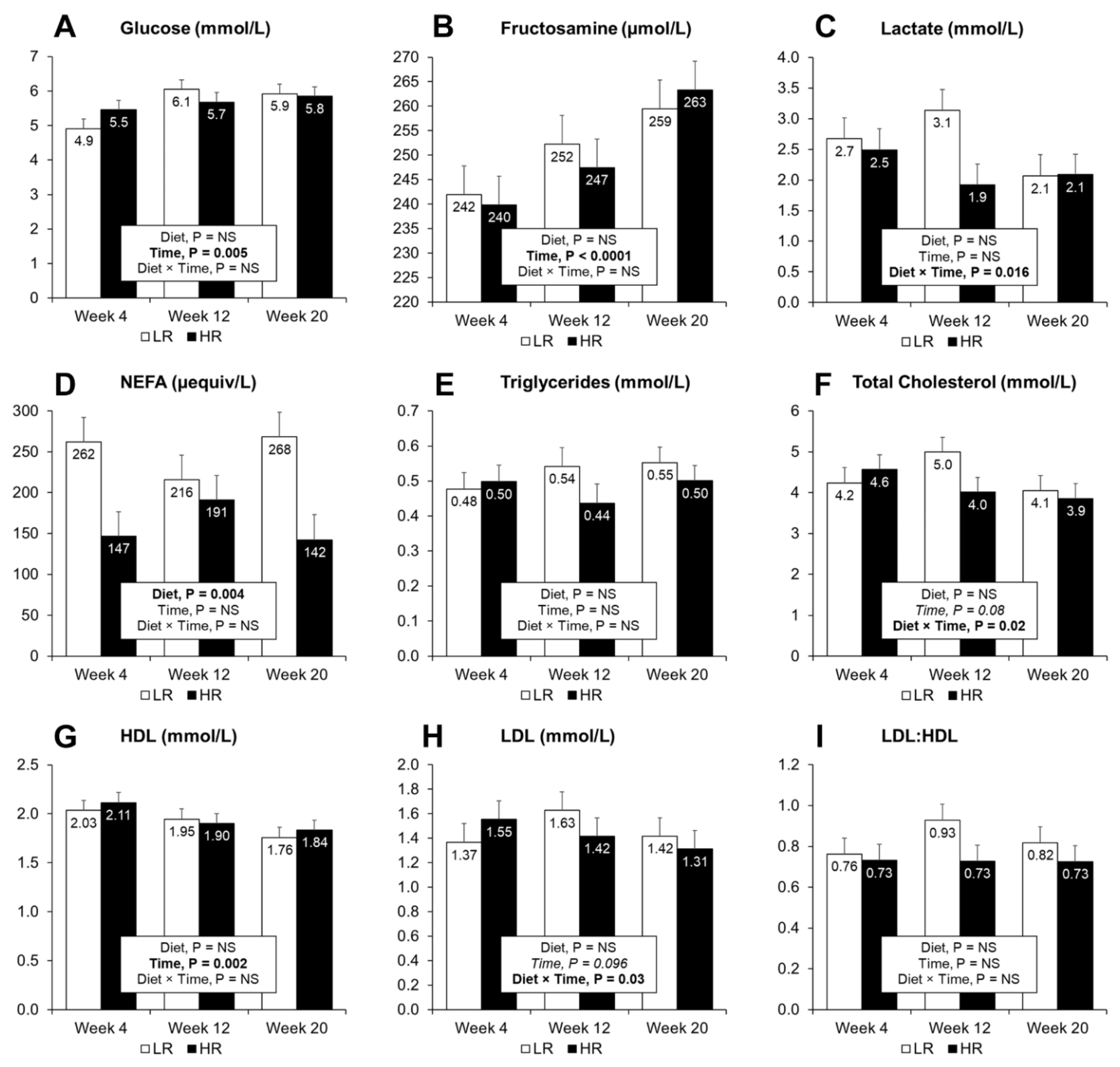
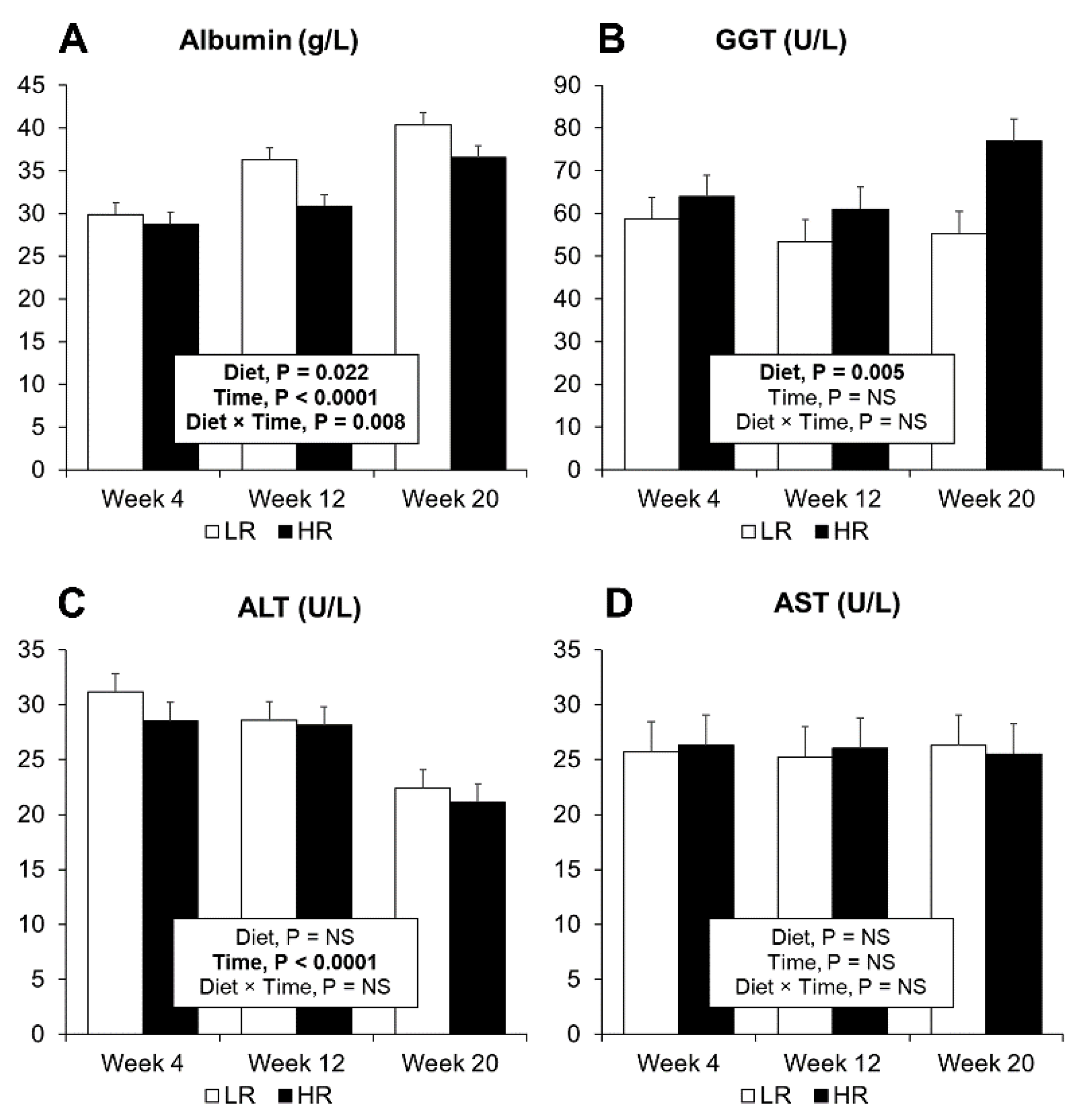
3.4. Plasma and Urine Biomarkers of MetS and Biomarkers of Inflammation
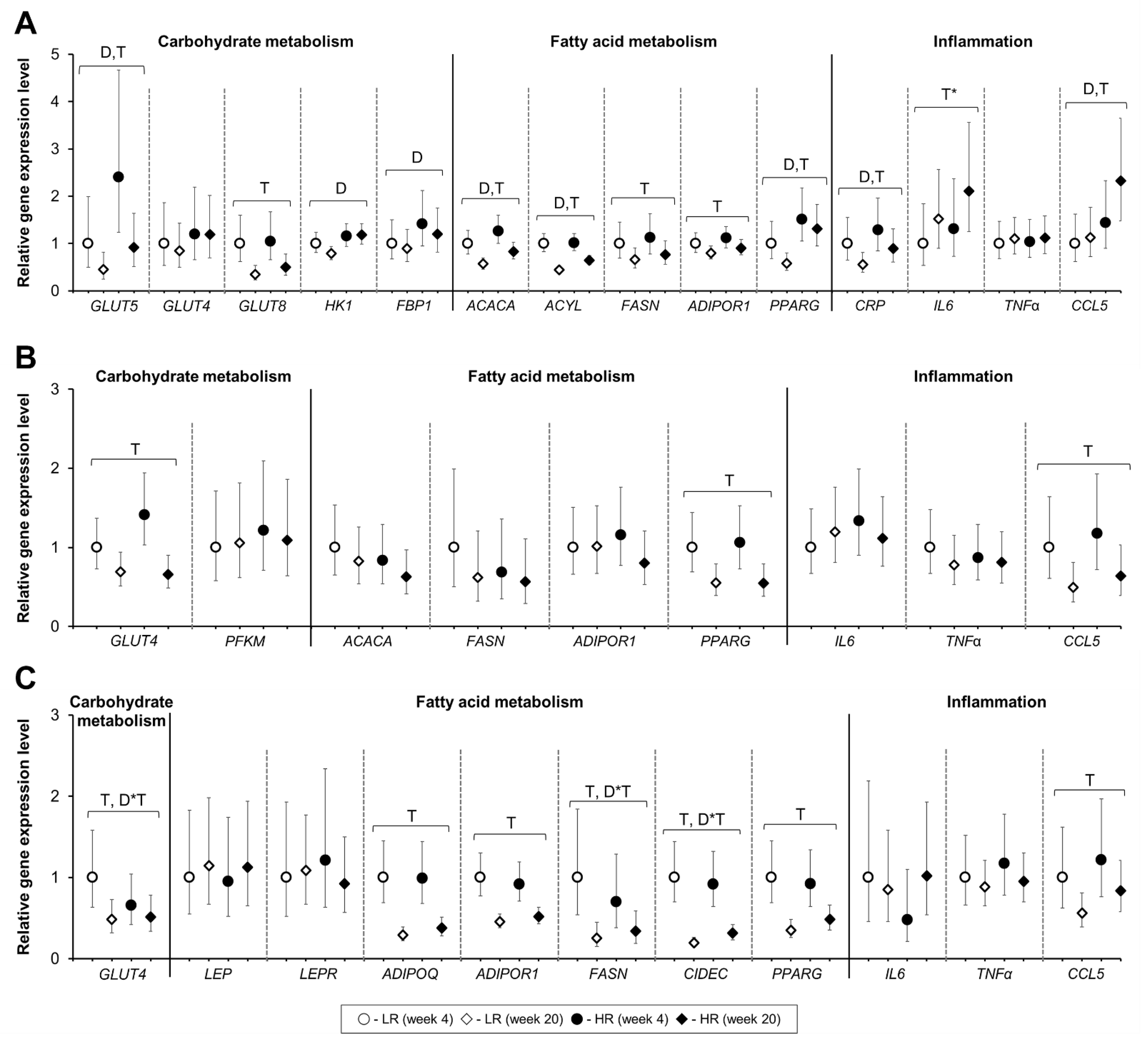
3.5. Tissue Gene Expression
4. Discussion
Study Limitations and Strengths
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alwahsh, S.M.; Gebhardt, R. Dietary fructose as a risk factor for non-alcoholic fatty liver disease (NAFLD). Arch. Toxicol. 2017, 91, 1545–1563. [Google Scholar] [CrossRef] [PubMed]
- Fryar, C.D.; Carroll, M.D.; Ogden, C.L. Prevalence of Overweight, Obesity, and Severe Obesity Among Children and Adolescents Aged 2–19 Years: United States, 1963–1965 Through 2015–2016. Available online: https://www.cdc.gov/nchs/data/hestat/obesity_child_15_16/obesity_child_15_16.htm (accessed on 3 March 2020).
- Huang, P.L. A comprehensive definition for metabolic syndrome. Dis. Model Mech. 2009, 2, 231–237. [Google Scholar] [CrossRef]
- Wong, S.K.; Chin, K.Y.; Suhaimi, F.H.; Fairus, A.; Ima-Nirwana, S. Animal models of metabolic syndrome: A review. Nutr. Metab. 2016, 13, 65. [Google Scholar] [CrossRef]
- Spurlock, M.E.; Gabler, N.K. The Development of Porcine Models of Obesity and the Metabolic Syndrome. J. Nutr. 2008, 138, 397–402. [Google Scholar] [CrossRef]
- Johansen, T.; Hansen, H.S.; Richelsen, B.; Malmlof, R. The obese Gottingen minipig as a model of the metabolic syndrome: Dietary effects on obesity, insulin sensitivity, and growth hormone profile. Comp. Med. 2001, 51, 150–155. [Google Scholar] [PubMed]
- Sébert, S.P.; Lecannu, G.; Kozlowski, F.; Siliart, B.; Bard, J.M.; Krempf, M.; Champ, M.M.J. Childhood obesity and insulin resistance in a Yucatan mini-piglet model: Putative roles of IGF-1 and muscle PPARs in adipose tissue activity and development. Int. J. Obes. 2005, 29, 324–333. [Google Scholar] [CrossRef]
- Renner, S.; Blutke, A.; Dobenecker, B.; Dhom, G.; Muller, T.D.; Finan, B.; Clemmensen, C.; Bernau, M.; Novak, I.; Rathkolb, B.; et al. Metabolic syndrome and extensive adipose tissue inflammation in morbidly obese Gottingen minipigs. Mol. Metab. 2018, 16, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Dawson, H.D.; Smith, A.D.; Chen, C.; Urban, J.F. An in-depth comparison of the porcine, murine and human inflammasomes; lessons from the porcine genome and transcriptome. Vet. Microbiol. 2017, 202, 2–15. [Google Scholar] [CrossRef]
- Christoffersen, B.; Golozoubova, V.; Pacini, G.; Svendsen, O.; Raun, K. The young göttingen minipig as a model of childhood and adolescent obesity: Influence of diet and gender. Obesity 2013, 21, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Christoffersen, B.O.; Grand, N.; Golozoubova, V.; Svendsen, O.; Raun, K. Gender-associated differences in metabolic syndrome-related parameters in Gottingen minipigs. Comp. Med. 2007, 57, 493–504. [Google Scholar]
- Stanhope, K.L.; Schwarz, J.M.; Havel, P.J. Adverse metabolic effects of dietary fructose: Results from the recent epidemiological, clinical, and mechanistic studies. Curr. Opin. Lipidol. 2013, 24, 198–206. [Google Scholar] [CrossRef]
- Havel, P.J. Dietary fructose: Implications for dysregulation of energy homeostasis and lipid/carbohydrate metabolism. Nutr. Rev. 2005, 63, 133–157. [Google Scholar] [CrossRef]
- Rutledge, A.C.; Adeli, K. Fructose and the metabolic syndrome: Pathophysiology and molecular mechanisms. Nutr. Rev. 2007, 65, S13–S23. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.Z.; Empie, M.W. Fructose metabolism in humans–What isotopic tracer studies tell us. Nutr. Metab. 2012, 9, 89. [Google Scholar] [CrossRef]
- Ter Horst, K.W.; Serlie, M.J. Fructose Consumption, Lipogenesis, and Non-Alcoholic Fatty Liver Disease. Nutrients 2017, 9, 981. [Google Scholar] [CrossRef]
- Nielsen, T.S.; Laerke, H.N.; Theil, P.K.; Sorensen, J.F.; Saarinen, M.; Forssten, S.; Knudsen, K.E. Diets high in resistant starch and arabinoxylan modulate digestion processes and SCFA pool size in the large intestine and faecal microbial composition in pigs. Br. J. Nutr. 2014, 112, 1837–1849. [Google Scholar] [CrossRef] [PubMed]
- Vangsøe, C.T.; Ingerslev, A.K.; Theil, P.K.; Hedemann, M.S.; Lærke, H.N.; Knudsen, K.E.B. In vitro starch digestion kinetics of diets varying in resistant starch and arabinoxylan compared with in vivo portal appearance of glucose in pigs. Food Res. Int. 2016, 88, 199–206. [Google Scholar] [CrossRef]
- Curtasu, M.V.; Tafintseva, V.; Bendiks, Z.A.; Marco, M.L.; Kohler, A.; Xu, Y.; Nørskov, N.P.; Nygaard Lærke, H.; Bach Knudsen, K.E.; Hedemann, M.S. Obesity-Related Metabolome and Gut Microbiota Profiles of Juvenile Göttingen Minipigs—Long-Term Intake of Fructose and Resistant Starch. Metabolites 2020, 10, 456. [Google Scholar] [CrossRef]
- Swindle, M.M.; Makin, A.; Herron, A.J.; Clubb, F.J.; Frazier, K.S. Swine as models in biomedical research and toxicology testing. Vet. Pathol. 2012, 49, 344–356. [Google Scholar] [CrossRef] [PubMed]
- Levy, J.C.; Matthews, D.R.; Hermans, M.P. Correct homeostasis model assessment (HOMA) evaluation uses the computer program. Diabetes Care 1998, 21, 2191–2192. [Google Scholar] [CrossRef]
- Christensen, K.L.; Hedemann, M.S.; Jorgensen, H.; Stagsted, J.; Knudsen, K.E. Liquid chromatography-mass spectrometry based metabolomics study of cloned versus normal pigs fed either restricted or ad libitum high-energy diets. J Proteome Res 2012, 11, 3573–3580. [Google Scholar] [CrossRef] [PubMed]
- Fisher, K.D.; Scheffler, T.L.; Kasten, S.C.; Reinholt, B.M.; van Eyk, G.R.; Escobar, J.; Scheffler, J.M.; Gerrard, D.E. Energy Dense, Protein Restricted Diet Increases Adiposity and Perturbs Metabolism in Young, Genetically Lean Pigs. PLoS ONE 2013, 8, e72320. [Google Scholar] [CrossRef] [PubMed]
- Ingerslev, A.K.; Mutt, S.J.; Lærke, H.N.; Hedemann, M.S.; Theil, P.K.; Nielsen, K.L.; Jørgensen, H.; Herzig, K.-H.; Bach Knudsen, K.E. Postprandial PYY increase by resistant starch supplementation is independent of net portal appearance of short-chain fatty acids in pigs. PLoS ONE 2017, 12, e0185927. [Google Scholar] [CrossRef] [PubMed]
- Karhunen, L.J.; Juvonen, K.R.; Huotari, A.; Purhonen, A.K.; Herzig, K.H. Effect of protein, fat, carbohydrate and fibre on gastrointestinal peptide release in humans. Regul. Pept. 2008, 149, 70–78. [Google Scholar] [CrossRef]
- de Leeuw, J.A.; Jongbloed, A.W.; Spoolder, H.A.M.; Verstegen, M.W.A. Effects of hindgut fermentation of non-starch polysaccharides on the stability of blood glucose and insulin levels and physical activity in empty sows. Livest. Prod. Sci. 2005, 96, 165–174. [Google Scholar] [CrossRef]
- Johnston, K.L.; Thomas, E.L.; Bell, J.D.; Frost, G.S.; Robertson, M.D. Resistant starch improves insulin sensitivity in metabolic syndrome. Diabet. Med. J. Br. Diabet. Assoc. 2010, 27, 391–397. [Google Scholar] [CrossRef]
- Maki, K.C.; Pelkman, C.L.; Finocchiaro, E.T.; Kelley, K.M.; Lawless, A.L.; Schild, A.L.; Rains, T.M. Resistant starch from high-amylose maize increases insulin sensitivity in overweight and obese men. J. Nutr. 2012, 142, 717–723. [Google Scholar] [CrossRef]
- Bollen, P.J.; Madsen, L.W.; Meyer, O.; Ritskes-Hoitinga, J. Growth differences of male and female Gottingen minipigs during ad libitum feeding: A pilot study. Lab. Anim. 2005, 39, 80–93. [Google Scholar] [CrossRef]
- Curtasu, M.V.; Knudsen, K.E.B.; Callesen, H.; Purup, S.; Stagsted, J.; Hedemann, M.S. Obesity Development in a Miniature Yucatan Pig Model: A Multi-compartmental Metabolomics Study on Cloned and Normal Pigs Fed Restricted or Ad Libitum High-Energy Diets. J. Proteome Res. 2019, 18, 30–47. [Google Scholar] [CrossRef]
- Stinson, E.J.; Piaggi, P.; Ibrahim, M.; Venti, C.; Krakoff, J.; Votruba, S.B. High Fat and Sugar Consumption During Ad Libitum Intake Predicts Weight Gain. Obesity 2018, 26, 689–695. [Google Scholar] [CrossRef]
- Perry, R.J.; Peng, L.; Barry, N.A.; Cline, G.W.; Zhang, D.; Cardone, R.L.; Petersen, K.F.; Kibbey, R.G.; Goodman, A.L.; Shulman, G.I. Acetate mediates a microbiome–brain–β-cell axis to promote metabolic syndrome. Nature 2016, 534, 213. [Google Scholar] [CrossRef]
- Souza da Silva, C.; Haenen, D.; Koopmans, S.J.; Hooiveld, G.J.; Bosch, G.; Bolhuis, J.E.; Kemp, B.; Muller, M.; Gerrits, W.J. Effects of resistant starch on behaviour, satiety-related hormones and metabolites in growing pigs. Animal 2014, 8, 1402–1411. [Google Scholar] [CrossRef]
- van Erp, R.J.J.; de Vries, S.; van Kempen, T.A.T.G.; Den Hartog, L.A.; Gerrits, W.J.J. Feed intake patterns nor growth rates of pigs are affected by dietary resistant starch, despite marked differences in digestion. Animal 2020, 14, 1402–1412. [Google Scholar] [CrossRef]
- Elliott, S.S.; Keim, N.L.; Stern, J.S.; Teff, K.; Havel, P.J. Fructose, weight gain, and the insulin resistance syndrome. Am. J. Clin. Nutr. 2002, 76, 911–922. [Google Scholar] [CrossRef]
- Jang, C.; Hui, S.; Lu, W.; Cowan, A.J.; Morscher, R.J.; Lee, G.; Liu, W.; Tesz, G.J.; Birnbaum, M.J.; Rabinowitz, J.D. The Small Intestine Converts Dietary Fructose into Glucose and Organic Acids. Cell Metab. 2018, 27, 351–361.e353. [Google Scholar] [CrossRef] [PubMed]
- Douard, V.; Ferraris, R.P. Regulation of the fructose transporter GLUT5 in health and disease. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E227–E237. [Google Scholar] [CrossRef]
- Janevski, M.; Ratnayake, S.; Siljanovski, S.; McGlynn, M.A.; Cameron-Smith, D.; Lewandowski, P. Fructose containing sugars modulate mRNA of lipogenic genes ACC and FAS and protein levels of transcription factors ChREBP and SREBP1c with no effect on body weight or liver fat. Food Funct. 2012, 3, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Alwahsh, S.M.; Dwyer, B.J.; Forbes, S.; Thiel, D.H.; Lewis, P.J.S.; Ramadori, G. Insulin Production and Resistance in Different Models of Diet-Induced Obesity and Metabolic Syndrome. Int. J. Mol. Sci. 2017, 18, 285. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, H.D.; Galsgaard, E.D.; Christoffersen, B.Ø.; Cirera, S.; Holst, D.; Fredholm, M.; Latta, M. NASH-inducing Diets in Göttingen Minipigs. J. Clin. Exp. Hepatol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Abel, E.D.; Peroni, O.; Kim, J.K.; Kim, Y.-B.; Boss, O.; Hadro, E.; Minnemann, T.; Shulman, G.I.; Kahn, B.B. Adipose-selective targeting of the GLUT4 gene impairs insulin action in muscle and liver. Nature 2001, 409, 729. [Google Scholar] [CrossRef]
- Carvalho, E.; Kotani, K.; Peroni, O.D.; Kahn, B.B. Adipose-specific overexpression of GLUT4 reverses insulin resistance and diabetes in mice lacking GLUT4 selectively in muscle. Am. J. Physiol. Endocrinol. Metab. 2005, 289, E551–E561. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zeng, X.; Ren, M.; Mao, X.; Qiao, S. Novel metabolic and physiological functions of branched chain amino acids: A review. J. Anim. Sci. Biotechnol. 2017, 8, 10. [Google Scholar] [CrossRef] [PubMed]
- Polakof, S.; Remond, D.; Bernalier-Donadille, A.; Rambeau, M.; Pujos-Guillot, E.; Comte, B.; Dardevet, D.; Savary-Auzeloux, I. Metabolic adaptations to HFHS overfeeding: How whole body and tissues postprandial metabolic flexibility adapt in Yucatan mini-pigs. Eur. J. Nutr. 2018, 57, 119–135. [Google Scholar] [CrossRef] [PubMed]
- Neeb, Z.P.; Edwards, J.M.; Alloosh, M.; Long, X.; Mokelke, E.A.; Sturek, M. Metabolic syndrome and coronary artery disease in Ossabaw compared with Yucatan swine. Comp. Med. 2010, 60, 300–315. [Google Scholar]
- Mentzel, C.M.J.; Cardoso, T.F.; Lex, A.M.J.; Sorensen, D.B.; Fredholm, M.; Cirera, S. Fat and carbohydrate content in the diet induces drastic changes in gene expression in young Gottingen minipigs. Mamm. Genome 2017, 28, 166–175. [Google Scholar] [CrossRef]
- Farnworth, E.R.; Kramer, J.K.G. Fat Metabolism in Growing Swine: A Review. Can. J. Anim. Sci. 1987, 67, 301–318. [Google Scholar] [CrossRef]
- Spalding, K.L.; Arner, E.; Westermark, P.O.; Bernard, S.; Buchholz, B.A.; Bergmann, O.; Blomqvist, L.; Hoffstedt, J.; Naslund, E.; Britton, T.; et al. Dynamics of fat cell turnover in humans. Nature 2008, 453, 783–787. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Contreras, C.; Vazquez-Gomez, M.; Torres-Rovira, L.; Gonzalez, J.; Porrini, E.; Gonzalez-Colaço, M.; Isabel, B.; Astiz, S.; Gonzalez-Bulnes, A. Characterization of Ageing- and Diet-Related Swine Models of Sarcopenia and Sarcopenic Obesity. Int. J. Mol. Sci. 2018, 19, 823. [Google Scholar] [CrossRef] [PubMed]
- Al-Mashhadi, A.L.; Poulsen, C.B.; von Wachenfeldt, K.; Robertson, A.K.; Bentzon, J.F.; Nielsen, L.B.; Thygesen, J.; Tolbod, L.P.; Larsen, J.R.; Moestrup, S.K.; et al. Diet-Induced Abdominal Obesity, Metabolic Changes, and Atherosclerosis in Hypercholesterolemic Minipigs. J. Diabetes Res. 2018, 2018, 6823193. [Google Scholar] [CrossRef]
- Maeda, N.; Takahashi, M.; Funahashi, T.; Kihara, S.; Nishizawa, H.; Kishida, K.; Nagaretani, H.; Matsuda, M.; Komuro, R.; Ouchi, N.; et al. PPAR Ligands Increase Expression and Plasma Concentrations of Adiponectin, an Adipose-Derived Protein. Diabetes 2001, 50, 2094–2099. [Google Scholar] [CrossRef]
- Kahn, S.E.; Hull, R.L.; Utzschneider, K.M. Mechanisms linking obesity to insulin resistance and type 2 diabetes. Nature 2006, 444, 840–846. [Google Scholar] [CrossRef] [PubMed]
- Despres, J.P.; Lemieux, I. Abdominal obesity and metabolic syndrome. Nature 2006, 444, 881–887. [Google Scholar] [CrossRef]
- Vieira-Potter, V.J.; Lee, S.; Bayless, D.S.; Scroggins, R.J.; Welly, R.J.; Fleming, N.J.; Smith, T.N.; Meers, G.M.; Hill, M.A.; Rector, R.S.; et al. Disconnect between adipose tissue inflammation and cardiometabolic dysfunction in Ossabaw pigs. Obesity 2015, 23, 2421–2429. [Google Scholar] [CrossRef] [PubMed]
- Kirovski, G.; Gäbele, E.; Dorn, C.; Moleda, L.; Niessen, C.; Weiss, T.S.; Wobser, H.; Schacherer, D.; Buechler, C.; Wasmuth, H.E.; et al. Hepatic steatosis causes induction of the chemokine RANTES in the absence of significant hepatic inflammation. Int. J. Clin. Exp. Pathol. 2010, 3, 675–680. [Google Scholar] [PubMed]
- Christoffersen, B.O.; Jensen, S.J.; Ludvigsen, T.P.; Nilsson, S.K.; Grossi, A.B.; Heegaard, P.M. Age- and Sex-Associated Effects on Acute-Phase Proteins in Gottingen Minipigs. Comp. Med. 2015, 65, 333–341. [Google Scholar] [PubMed]
- Schumacher-Petersen, C.; Christoffersen, B.Ø.; Kirk, R.K.; Ludvigsen, T.P.; Zois, N.E.; Pedersen, H.D.; Vyberg, M.; Olsen, L.H. Experimental non-alcoholic steatohepatitis in Göttingen Minipigs: Consequences of high fat-fructose-cholesterol diet and diabetes. J. Transl. Med. 2019, 17, 110. [Google Scholar] [CrossRef] [PubMed]
- Hsu, M.-C.; Wang, M.-E.; Jiang, Y.-F.; Liu, H.-C.; Chen, Y.-C.; Chiu, C.-H. Long-term feeding of high-fat plus high-fructose diet induces isolated impaired glucose tolerance and skeletal muscle insulin resistance in miniature pigs. Diabetol. Metab. Syndr. 2017, 9, 81. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.; Alloosh, M.; Saxena, R.; Van Alstine, W.; Watkins, B.A.; Klaunig, J.E.; Sturek, M.; Chalasani, N. Nutritional model of steatohepatitis and metabolic syndrome in the Ossabaw miniature swine. Hepatology 2009, 50, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Cirera, S.; Tasoz, E.; Juul Jacobsen, M.; Schumacher-Petersen, C.; Ostergaard Christoffersen, B.; Kaae Kirk, R.; Pagh Ludvigsen, T.; Hvid, H.; Duelund Pedersen, H.; Hoier Olsen, L.; et al. The expression signatures in liver and adipose tissue from obese Gottingen Minipigs reveal a predisposition for healthy fat accumulation. Nutr. Diabetes 2020, 10, 9. [Google Scholar] [CrossRef]
- Pullen, D.L.; Liesman, J.S.; Emery, R.S. A species comparison of liver slice synthesis and secretion of triacylglycerol from nonesterified fatty acids in media2. J. Anim. Sci. 1990, 68, 1395–1399. [Google Scholar] [CrossRef] [PubMed]
- Bergen, W.G.; Mersmann, H.J. Comparative Aspects of Lipid Metabolism: Impact on Contemporary Research and Use of Animal Models. J. Nutr. 2005, 135, 2499–2502. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LR | HR | |
|---|---|---|
| Chemical composition (g/kg DM 1) | ||
| DM (g/kg, as-fed basis) | 917 | 913 |
| Ash | 63 | 62 |
| Protein (N × 6.25) | 119 | 113 |
| Fat | 177 | 174 |
| Available carbohydrates | 424 | 555 |
| Sugars | 9 | 233 |
| Fructose | 0.6 | 225 |
| Glucose | 1.2 | 0.8 |
| Sucrose | 7 | 7 |
| Starch | 415 | 322 |
| Total dietary fiber 2 | 188 | 100 |
| Total NSP (soluble NSP) 3 | 73 (15) | 69 (8) |
| RS 4 | 89 | 2 |
| AXOS 5 | 3 | 5 |
| Fructans | 5 | 6 |
| Klason lignin | 18 | 18 |
| Gross energy (MJ/kg DM) | 20.3 | 20.7 |
| LR 1 | HR 1 | SEM | p-Value | |
|---|---|---|---|---|
| Nutrient intake (g/day) 2 | ||||
| Dry matter | 694 | 548 | 78 | 0.023 |
| Available carbohydrates | 295 | 304 | 39 | 0.775 |
| Protein | 83 | 62 | 9 | 0.006 |
| Fat | 123 | 95 | 14 | 0.015 |
| Total dietary fiber 3 | 130 | 55 | 13 | <0.0001 |
| Relative energy contribution (%) 4 | ||||
| Carbohydrates | 41.8 | 50.8 | ||
| Fat | 37.9 | 34.6 | ||
| Protein | 11.7 | 10.3 | ||
| Total dietary fiber | 8.7 | 4.3 | ||
| p-Value 2 | ||||||
|---|---|---|---|---|---|---|
| Week | LR 1 | HR 1 | Diet | Time | Diet × Time | |
| Creatinine (µmol/L) | 4 | 4.7 (3.6, 6.1) | 10 (7.4, 13.6) | 0.0002 | NS | 0.02 |
| 12 | 6.8 (5.3, 8.9) | 9.4 (7, 12.6) | ||||
| 20 | 5.9 (3.9, 6.8) | 12 (9.4, 15.3) | ||||
| Glucose (mmol/L) | 4 | 0.4 (0.1, 1.3) | 1.8 (0.4, 7.2) | 0.004 | 0.03 | NS |
| 12 | 0.4 (0.1, 1.4) | 4.7 (1.2, 17.8) | ||||
| 20 | 3.8 (1.2, 12.1) | 3.6 (1.3, 10.4) | ||||
| Total Protein (mg/L) | 4 | 74.9 (46, 123) | 136.1 (76, 243) | NS | NS | 0.03 |
| 12 | 169.9 (104, 278) | 75.6 (43, 134) | ||||
| 20 | 150.5 (92, 246) | 150 (97, 233) | ||||
| Glucose: Creatinine Ratio 3 | 4 | 0.1 (0.03, 0.3) | 0.2 (0.05, 0.7) | NS | NS | NS |
| 12 | 0.1 (0.02, 0.2) | 0.2 (0.06, 0.7) | ||||
| 20 | 0.4 (0.11, 1.1) | 0.3 (0.11, 0.8) | ||||
| Protein Creatinine Ratio | 4 | 17.1 (11.3, 22.9) | 17.2 (10.8, 23.6) | 0.004 | NS | 0.05 |
| 12 | 27 (18.2, 35.9) | 4.5 (−5.7, 14.8) | ||||
| 20 | 27.7 (18.6, 36.8) | 13.2 (4.6, 21.9) | ||||
| p-Value 4 | |||||||
|---|---|---|---|---|---|---|---|
| Week | LR 1 | HR 1 | SEM | Diet | Time | Diet × Time | |
| C-Peptide 2 | 4 | 28 | 27.2 | ||||
| 12 | 34.1 | 27.5 | 3.6 | NS | NS | NS | |
| 20 | 30 | 27.9 | |||||
| Ghrelin 2 | 4 | 11.3 | 15 | ||||
| 12 | 13.6 | 10.6 | 3.2 | NS | NS | NS | |
| 20 | 11.1 | 16.5 | |||||
| GIP 2 | 4 | 92.3 | 94.2 | ||||
| 12 | 70.8 | 55.7 | 8.4 | NS | <0.0001 | NS | |
| 20 | 43.8 | 50.3 | |||||
| GLP1T 3 | 4 | 207.6 | 234.6 | ||||
| 12 | 258.8 | 255.2 | 18.8 | NS | 0.04 | NS | |
| 20 | 236 | 236.1 | |||||
| Glucagon 2 | 4 | 163.1 | 202.5 | ||||
| 12 | 228.7 | 302.1 | 25.5 | 0.01 | <0.0001 | NS | |
| 20 | 213.8 | 295.5 | |||||
| PYY 2 | 4 | 281.8 | 234 | ||||
| 12 | 368.7 | 227.3 | 35.7 | 0.0005 | NS | NS | |
| 20 | 299.3 | 170.1 | |||||
| Insulin (pmol/L) | 4 | 28.0 | 31.2 | ||||
| 12 | 33.9 | 36.4 | 4.6 | NS | 0.03 | NS | |
| 20 | 38.9 | 43.5 | |||||
| HOMA-IR | 4 | 0.52 | 0.59 | ||||
| 12 | 0.66 | 0.71 | 0.09 | NS | 0.02 | NS | |
| 20 | 0.76 | 0.84 | |||||
| HOMA-B | 4 | 63.6 | 60.4 | ||||
| 12 | 52.3 | 60.3 | 8.7 | NS | NS | NS | |
| 20 | 56.2 | 67.8 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Curtasu, M.V.; Skou Hedemann, M.; Nygaard Lærke, H.; Bach Knudsen, K.E. Obesity Development and Signs of Metabolic Abnormalities in Young Göttingen Minipigs Consuming Energy Dense Diets Varying in Carbohydrate Quality. Nutrients 2021, 13, 1560. https://doi.org/10.3390/nu13051560
Curtasu MV, Skou Hedemann M, Nygaard Lærke H, Bach Knudsen KE. Obesity Development and Signs of Metabolic Abnormalities in Young Göttingen Minipigs Consuming Energy Dense Diets Varying in Carbohydrate Quality. Nutrients. 2021; 13(5):1560. https://doi.org/10.3390/nu13051560
Chicago/Turabian StyleCurtasu, Mihai Victor, Mette Skou Hedemann, Helle Nygaard Lærke, and Knud Erik Bach Knudsen. 2021. "Obesity Development and Signs of Metabolic Abnormalities in Young Göttingen Minipigs Consuming Energy Dense Diets Varying in Carbohydrate Quality" Nutrients 13, no. 5: 1560. https://doi.org/10.3390/nu13051560
APA StyleCurtasu, M. V., Skou Hedemann, M., Nygaard Lærke, H., & Bach Knudsen, K. E. (2021). Obesity Development and Signs of Metabolic Abnormalities in Young Göttingen Minipigs Consuming Energy Dense Diets Varying in Carbohydrate Quality. Nutrients, 13(5), 1560. https://doi.org/10.3390/nu13051560