
Upregulation of Vitamin C Transporter Functional Expression in 5xFAD Mouse Intestine




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Animals
2.3. AA Determination in Mice Serum
2.4. 14C-AA Uptake
2.5. RT-qPCR Analysis
2.6. Heterogeneous Nuclear RNA (hnRNA) Analysis
2.7. Western Blot Analysis
2.8. Immunohistochemistry
2.9. Statistical Analysis
3. Results
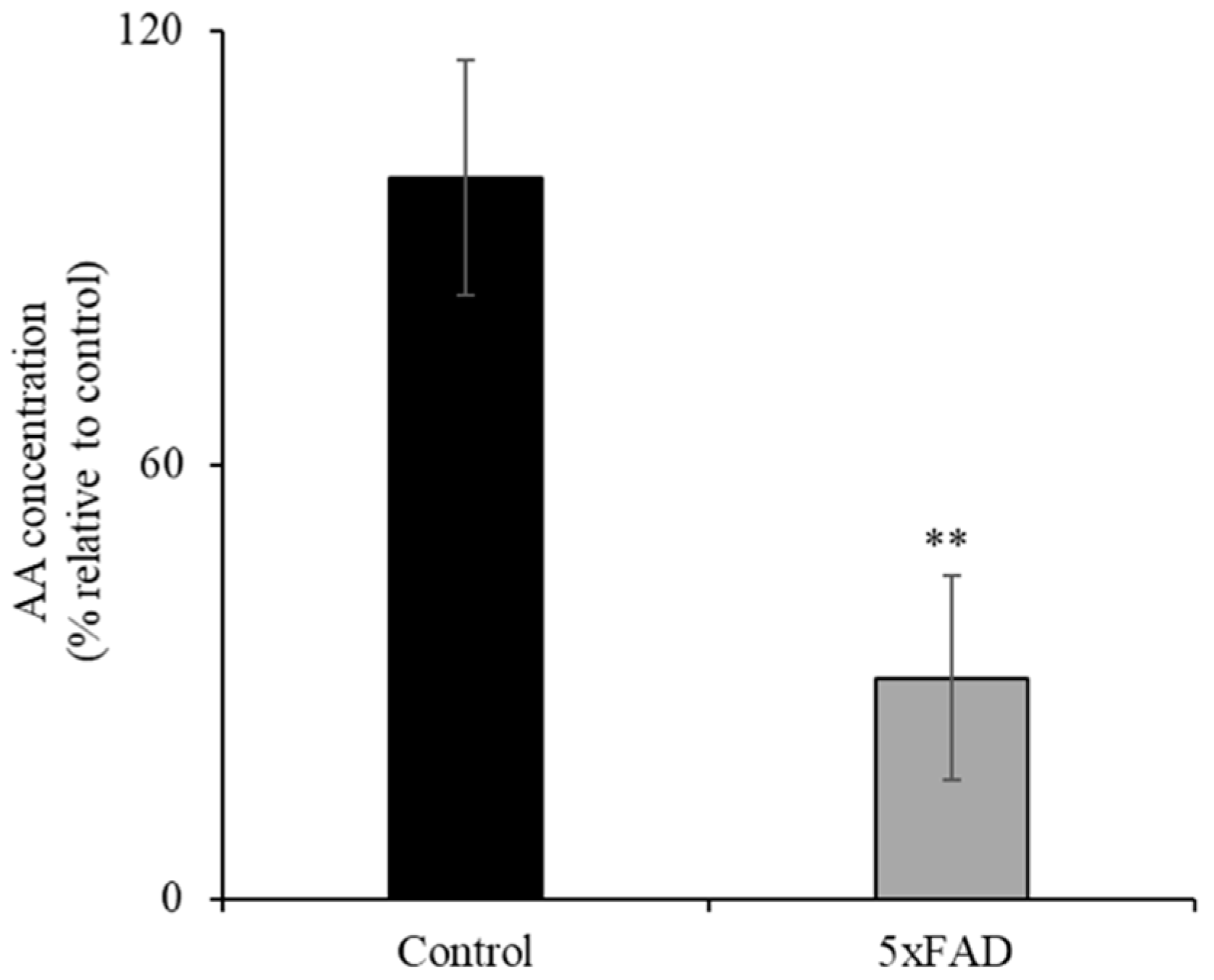
3.1. Level of AA in 5xFAD Mice Serum Samples
3.2. Levels of Oxidative Stress Response Genes Expression in the Intestine of the 5xFAD Mice
3.3. 14C-AA Uptake Upregulated in the Jejunum of 5xFAD Mice
3.4. SVCT1 and SVCT2 Protein and mRNA Expression Levels in the Jejunum of 5xFAD Mice
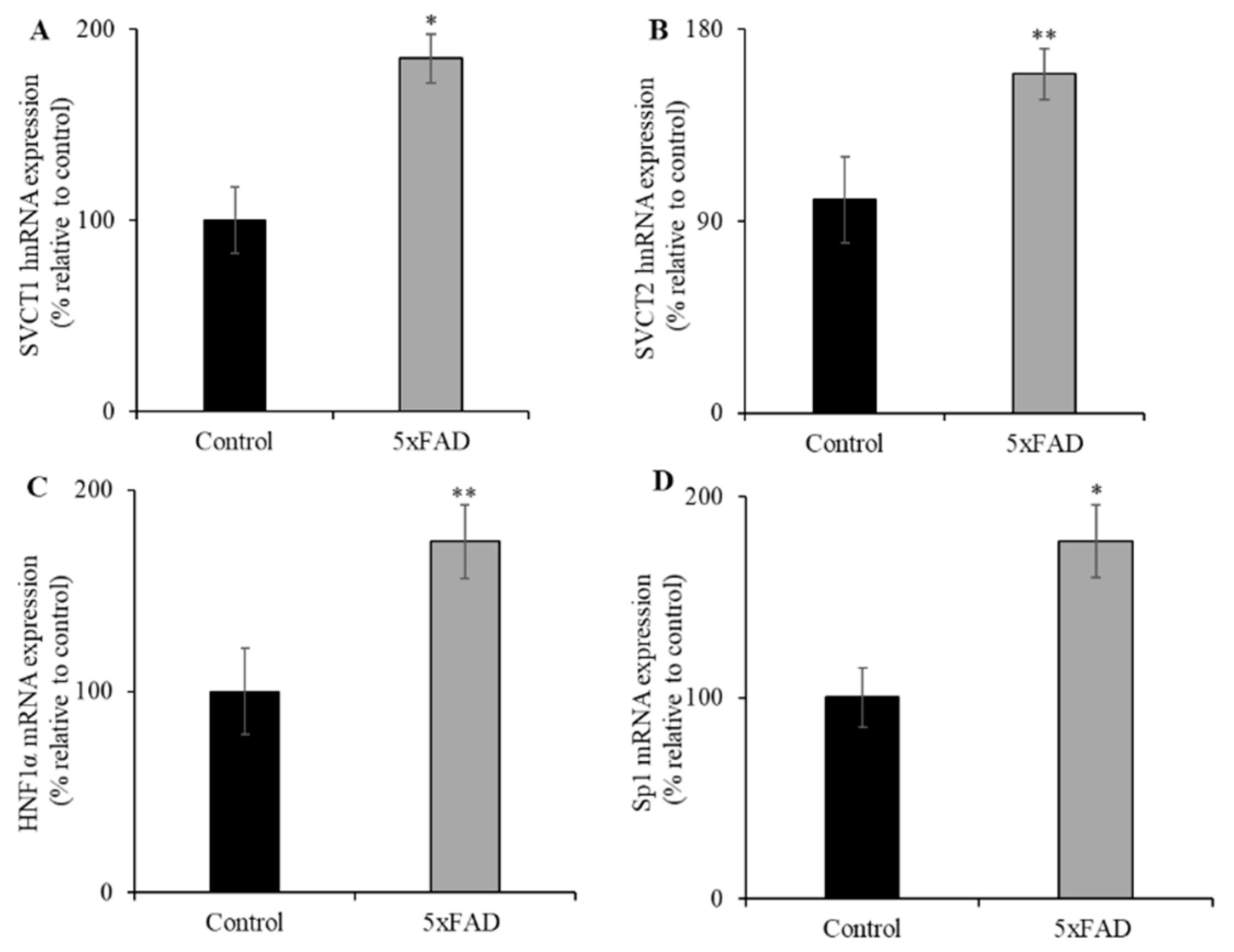
3.5. Transcriptional Upregulation of SVCT1 and SVCT2 in the Jejunum of 5xFAD Mice
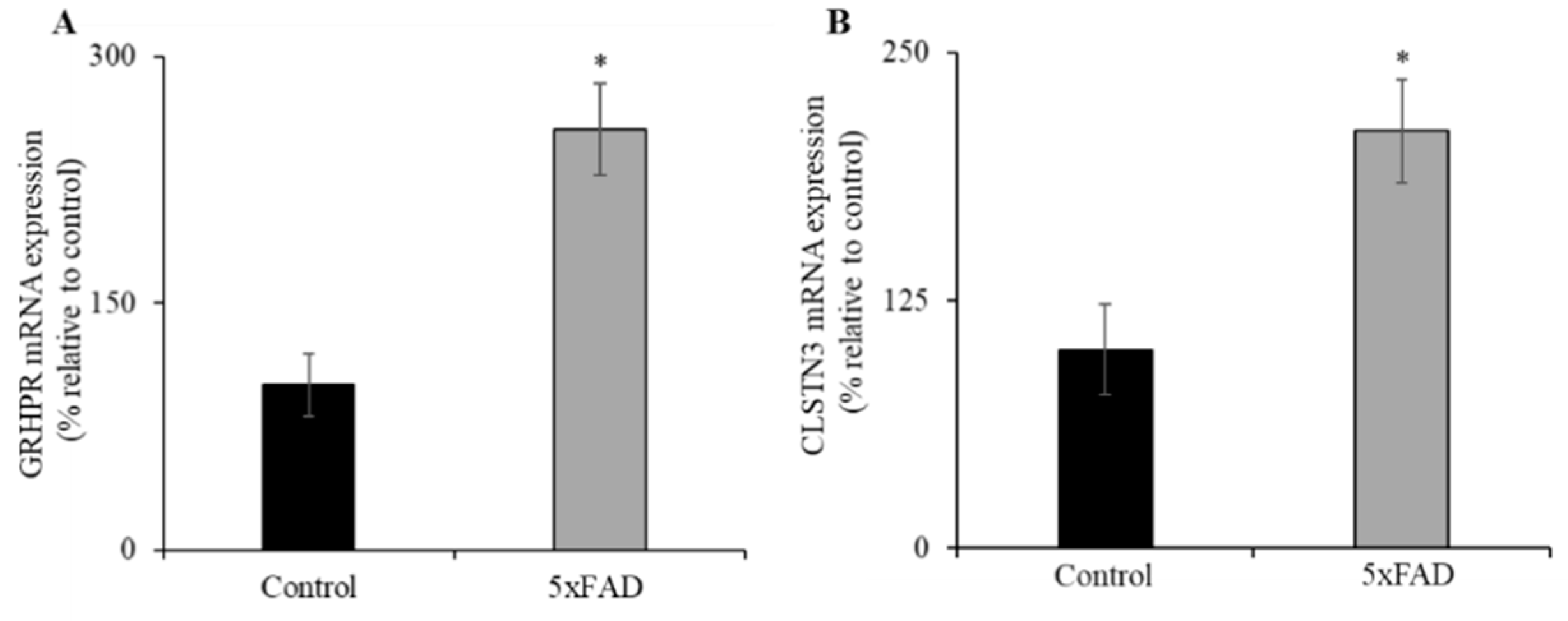
3.6. Expression of GRHPR and CLSTN3 mRNA in Jejunum of 5xFAD Mice
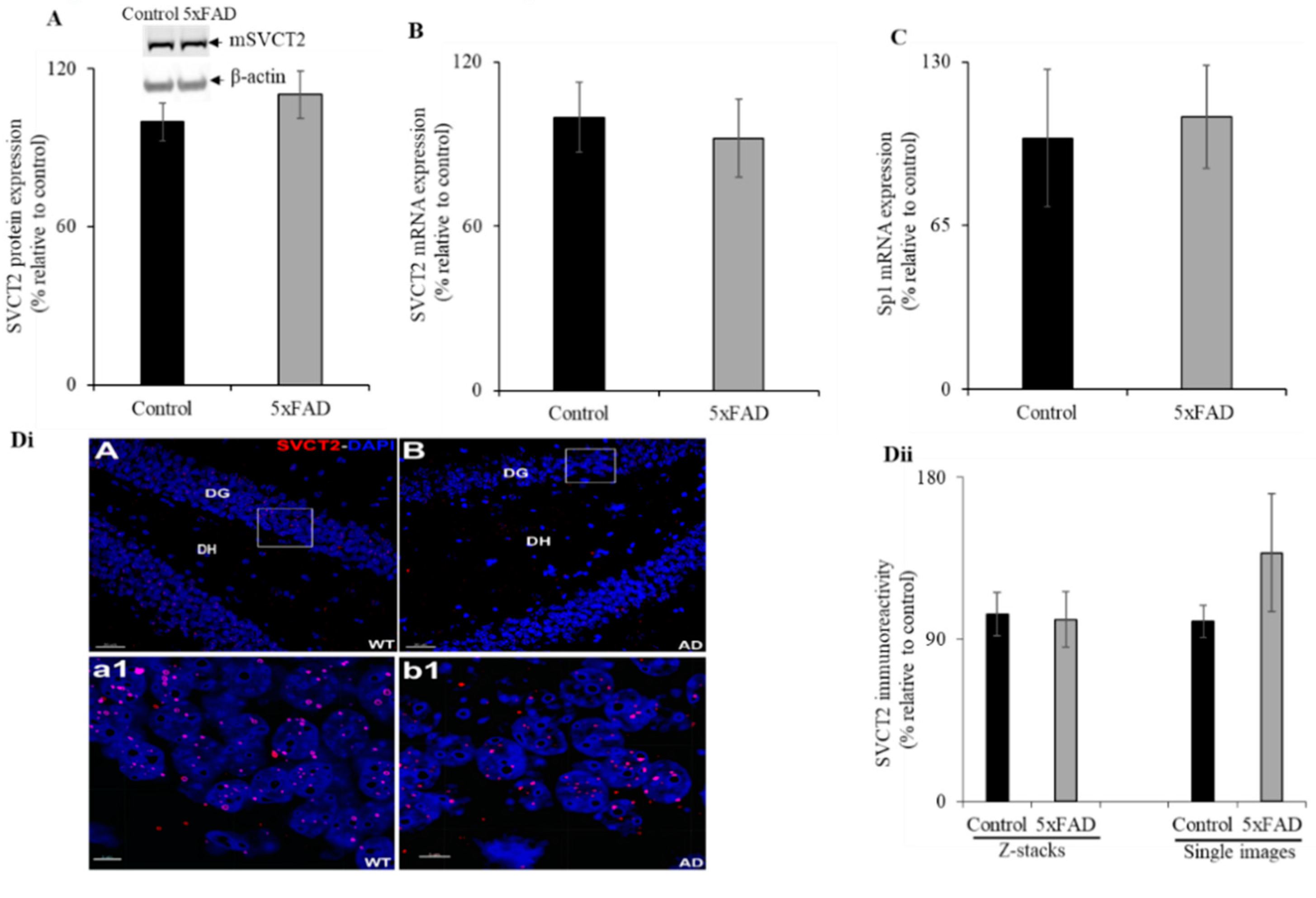
3.7. Hippocampal Expression of SVCT2 in the 5xFAD Mice
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carr, A.C.; Frei, B. Toward a new recommended dietary allowance for vitamin C based on antioxidant and health effects in humans. Am. J. Clin. Nutr. 1999, 69, 1086–1107. [Google Scholar] [CrossRef] [PubMed]
- Packer, L.; Fuchs, J. Vitamin C in Health and Disease; Marcel Dekker Inc: New York, NY, USA, 1997. [Google Scholar]
- Harrison, F.E. A Critical Review of Vitamin C for the Prevention of Age-Related Cognitive Decline and Alzheimer’s Disease. J. Alzheimer’s Dis. 2012, 29, 711–726. [Google Scholar] [CrossRef]
- Dixit, S.; Bernardo, A.; Walker, J.M.; Kennard, J.A.; Kim, G.Y.; Kessler, E.S.; Harrison, F.E. Vitamin C Deficiency in the Brain Impairs Cognition, Increases Amyloid Accumulation and Deposition, and Oxidative Stress in APP/PSEN1 and Normally Aging Mice. ACS Chem. Neurosci. 2015, 6, 570–581. [Google Scholar] [CrossRef] [PubMed]
- Mefford, I.N.; Oke, A.F.; Adams, R.N. Regional distribution of ascorbate in human brain. Brain Res. 1981, 212, 223–226. [Google Scholar] [CrossRef]
- Monacelli, F.; Acquarone, F.M.E.; Giannotti, C.; Borghi, R.; Nencioni, A. Vitamin C, Aging and Alzheimer’s Disease. Nutr. 2017, 9, 670. [Google Scholar] [CrossRef]
- Murakami, K.; Murata, N.; Ozawa, Y.; Kinoshita, N.; Irie, K.; Shirasawa, T.; Shimizu, T. Vitamin C Restores Behavioral Deficits and Amyloid-β Oligomerization without Affecting Plaque Formation in a Mouse Model of Alzheimer’s Disease. J. Alzheimer’s Dis. 2011, 26, 7–18. [Google Scholar] [CrossRef] [PubMed]
- E Charlton, K.; Rabinowitz, T.L.; Geffen, L.N.; A Dhansay, M. Lowered plasma vitamin C, but not vitamin E, concentrations in dementia patients. J. Nutr. Health Aging 2004, 8, 99–107. [Google Scholar]
- May, J.M. Vitamin C Transport and Its Role in the Central Nervous System. Cholest. Bind. Cholest. Transp. Proteins 2011, 56, 85–103. [Google Scholar] [CrossRef]
- Rose, R.C.; Nahrwold, D.L. Intestinal ascorbic acid transport following diets of high or low ascorbic acid content. Int. J. Vitam. Nutr. Res. 1978, 48, 382–386. [Google Scholar] [PubMed]
- Bürzle, M.; Suzuki, Y.; Ackermann, D.; Miyazaki, H.; Maeda, N.; Clémençon, B.; Burrier, R.; Hediger, M.A. The sodium-dependent ascorbic acid transporter family SLC23. Mol. Asp. Med. 2013, 34, 436–454. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, V.S.; Srinivasan, P.; Wildman, A.J.; Marchant, J.S.; Said, H.M. Molecular mechanism(s) involved in differential expression of vitamin C transporters along the intestinal tract. Am. J. Physiol. Liver Physiol. 2017, 312, G340–G347. [Google Scholar] [CrossRef]
- Corpe, C.P.; Tu, H.; Eck, P.; Wang, J.; Faulhaber-Walter, R.; Schnermann, J.; Margolis, S.; Padayatty, S.; Sun, H.; Wang, Y.; et al. Vitamin C transporter Slc23a1 links renal reabsorption, vitamin C tissue accumulation, and perinatal survival in mice. J. Clin. Investig. 2010, 120, 1069–1083. [Google Scholar] [CrossRef]
- Sotiriou, S.; Gispert, S.; Cheng, J.; Wang, Y.; Chen, A.; Hoogstraten-Miller, S.; Miller, G.F.; Kwon, O.; Levine, M.; Guttentag, S.H.; et al. Ascorbic-acid transporter Slc23a1 is essential for vitamin C transport into the brain and for perinatal survival. Nat. Med. 2002, 8, 514–517. [Google Scholar] [CrossRef] [PubMed]
- Rice, M.E. Ascorbate regulation and its neuroprotective role in the brain. Trends Neurosci. 2000, 23, 209–216. [Google Scholar] [CrossRef]
- Oakley, H.; Cole, S.L.; Logan, S.; Maus, E.; Shao, P.; Craft, J.; Guillozet-Bongaarts, A.; Ohno, M.; Disterhoft, J.; Van Eldik, L.; et al. Intraneuronal beta-Amyloid Aggregates, Neurodegeneration, and Neuron Loss in Transgenic Mice with Five Familial Alzheimer’s Disease Mutations: Potential Factors in Amyloid Plaque Formation. J. Neurosci. 2006, 26, 10129–10140. [Google Scholar] [CrossRef] [PubMed]
- Baulch, J.E.; Acharya, M.M.; Agrawal, S.; Apodaca, L.A.; Monteiro, C.; Agrawal, A. Immune and Inflammatory Determinants Underlying Alzheimer’s Disease Pathology. J. Neuroimmune Pharmacol. 2020, 15, 852–862. [Google Scholar] [CrossRef]
- Subramanian, V.S.; Sabui, S.; Moradi, H.; Marchant, J.S.; Said, H.M. Inhibition of intestinal ascorbic acid uptake by lipopolysaccharide is mediated via transcriptional mechanisms. Biochim. et Biophys. Acta Biomembr. 2018, 1860, 556–565. [Google Scholar] [CrossRef] [PubMed]
- Heskett, C.W.; Teafatiller, T.; Hennessey, C.; Gareau, M.G.; Marchant, J.S.; Said, H.M.; Subramanian, V.S. Enteropathogenic Escherichia coli Infection Inhibits Intestinal Ascorbic Acid Uptake via Dysregulation of Its Transporter Expression. Dig. Dis. Sci. 2020, 1–11. [Google Scholar] [CrossRef]
- Acharya, M.M.; Baulch, J.E.; Lusardi, T.A.; Allen, B.D.; Chmielewski, N.N.; Baddour, A.A.D.; Limoli, C.L.; Boison, D. Adenosine Kinase Inhibition Protects against Cranial Radiation-Induced Cognitive Dysfunction. Front. Mol. Neurosci. 2016, 9, 42. [Google Scholar] [CrossRef]
- Besler, H.T.; Çomoğlu, S.; Okçu, Z. Serum Levels of Antioxidant Vitamins and Lipid Peroxidation in Multiple Sclerosis. Nutr. Neurosci. 2002, 5, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Bae, S.; Yu, Y.; Kim, Y.; Kim, H.-R.; Hwang, Y.-I.; Kang, J.S.; Lee, W.J. The Analysis of Vitamin C Concentration in Organs ofGulo-/-Mice Upon Vitamin C Withdrawal. Immune Netw. 2012, 12, 18–26. [Google Scholar] [CrossRef]
- Harrison, F.E.; Meredith, M.E.; Dawes, S.M.; Saskowski, J.L.; May, J.M. Low ascorbic acid and increased oxidative stress in gulo(−/−) mice during development. Brain Res. 2010, 1349, 143–152. [Google Scholar] [CrossRef]
- Lykkesfeldt, J.; Trueba, G.P.; Poulsen, H.E.; Christen, S. Vitamin C deficiency in weanling guinea pigs: Differential expression of oxidative stress and DNA repair in liver and brain. Br. J. Nutr. 2007, 98, 1116–1119. [Google Scholar] [CrossRef] [PubMed]
- Paidi, M.D.; Schjoldager, J.G.; Lykkesfeldt, J.; Tveden-Nyborg, P. Prenatal vitamin C deficiency results in differential levels of oxidative stress during late gestation in foetal guinea pig brains. Redox Biol. 2014, 2, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Xiao, B.; Cui, S.; Song, H.; Qian, Y.; Dong, L.; An, H.; Cui, Y.; Zhang, W.; He, Y.; et al. Triptolide treatment reduces Alzheimer’s disease (AD)-like pathology through inhibition of BACE1 in a transgenic mouse model of AD. Dis. Model. Mech. 2014, 7, 1385–1395. [Google Scholar] [CrossRef]
- Kook, S.-Y.; Lee, K.-M.; Kim, Y.; Cha, M.-Y.; Kang, S.; Baik, S.H.; Lee, H.; Park, R.; Mook-Jung, I. High-dose of vitamin C supplementation reduces amyloid plaque burden and ameliorates pathological changes in the brain of 5XFAD mice. Cell Death Dis. 2014, 5, e1083. [Google Scholar] [CrossRef] [PubMed]
- Ferraris, R.P.; Diamond, J.M. Specific Regulation of Intestinal Nutrient Transporters by their Dietary Substrates. Annu. Rev. Physiol. 1989, 51, 125–141. [Google Scholar] [CrossRef]
- Karasov, W.H.; Diamond, J.M. Adaptive regulation of sugar and amino acid transport by vertebrate intestine. Am. J. Physiol. Liver Physiol. 1983, 245, G443–G462. [Google Scholar] [CrossRef] [PubMed]
- Reidling, J.C.; Nabokina, S.M.; Said, H.M. Molecular mechanisms involved in the adaptive regulation of human intestinal biotin uptake: A study of the hSMVT system. Am. J. Physiol. Liver Physiol. 2007, 292, G275–G281. [Google Scholar] [CrossRef]
- Reidling, J.C.; Said, H.M. Adaptive regulation of intestinal thiamin uptake: Molecular mechanism using wild-type and transgenic mice carrying hTHTR-1 and -2 promoters. Am. J. Physiol. Liver Physiol. 2005, 288, G1127–G1134. [Google Scholar] [CrossRef]
- Subramanian, V.S.; Chatterjee, N.; Said, H.M. Folate uptake in the human intestine: Promoter activity and effect of folate deficiency. J. Cell. Physiol. 2003, 196, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, V.S.; Ghosal, A.; Kapadia, R.; Nabokina, S.M.; Said, H.M. Molecular Mechanisms Mediating the Adaptive Regulation of Intestinal Riboflavin Uptake Process. PLOS ONE 2015, 10, e0131698. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, V.S.; Marchant, J.S.; Boulware, M.J.; Ma, T.Y.; Said, H.M. Membrane targeting and intracellular trafficking of the human sodium-dependent multivitamin transporter in polarized epithelial cells. Am. J. Physiol. Physiol. 2009, 296, C663–C671. [Google Scholar] [CrossRef] [PubMed]
- Elferink, C.J.; Reiners, J.J. Quantitative RT-PCR on CYP1A1 Heterogeneous Nuclear RNA: A Surrogate for the In Vitro Transcription Run-On Assay. Biotech. 1996, 20, 470–477. [Google Scholar] [CrossRef]
- Reidling, J.C.; Subramanian, V.S.; Dahhan, T.; Sadat, M.; Said, H.M. Mechanisms and regulation of vitamin C uptake: Studies of the hSVCT systems in human liver epithelial cells. Am. J. Physiol. Liver Physiol. 2008, 295, G1217–G1227. [Google Scholar] [CrossRef]
- Reidling, J.C.; Rubin, S.A. Promoter analysis of the human ascorbic acid transporters SVCT1 and 2: Mechanisms of adaptive regulation in liver epithelial cells. J. Nutr. Biochem. 2011, 22, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Rubin, S.A.; Dey, S.; Reidling, J.C. Functional analysis of two regulatory regions of the human Na+-dependent vitamin C transporter 2, SLC23A2, in human vascular smooth muscle cells. Biochim. Biophys. Acta Gene Struct. Expr. 2005, 1732, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, V.S.; Nabokina, S.M.; Patton, J.R.; Marchant, J.S.; Moradi, H.; Said, H.M. Glyoxalate reductase/hydroxypyruvate reductase interacts with the sodium-dependent vitamin C transporter-1 to regulate cellular vitamin C homeostasis. Am. J. Physiol. Liver Physiol. 2013, 304, G1079–G1086. [Google Scholar] [CrossRef]
- Marcos, P.; González-Fuentes, J.; Castro-Vázquez, L.; Lozano, M.V.; Santander-Ortega, M.J.; Rodríguez-Robledo, V.; Villaseca-González, N.; Arroyo-Jiménez, M.M. Vitamin transporters in mice brain with aging. J. Anat. 2018, 232, 699–715. [Google Scholar] [CrossRef]
- Rinaldi, P.; Polidori, M.; Metastasio, A.; Mariani, E.; Mattioli, P.; Cherubini, A.; Catani, M.; Cecchetti, R.; Senin, U.; Mecocci, P. Plasma antioxidants are similarly depleted in mild cognitive impairment and in Alzheimer’s disease. Neurobiol. Aging 2003, 24, 915–919. [Google Scholar] [CrossRef]
- Rivière, S.; Birlouez-Aragon, I.; Nourhashémi, F.; Vellas, B. Low plasma vitamin C in Alzheimer patients despite an adequate diet. Int. J. Geriatr. Psychiatry 1998, 13, 749–754. [Google Scholar] [CrossRef]
- Traebert, M.; Hattenhauer, O.; Murer, H.; Kaissling, B.; Biber, J. Expression of type II Na-Picotransporter in alveolar type II cells. Am. J. Physiol. Cell. Mol. Physiol. 1999, 277, L868–L873. [Google Scholar] [CrossRef]
- Harrison, F.E.; Hosseini, A.H.; McDonald, M.P.; May, J.M. Vitamin C reduces spatial learning deficits in middle-aged and very old APP/PSEN1 transgenic and wild-type mice. Pharmacol. Biochem. Behav. 2009, 93, 443–450. [Google Scholar] [CrossRef]
- Heo, J.-H.; Lee, H.-; Lee, K.-M. The Possible Role of Antioxidant Vitamin C in Alzheimer’s Disease Treatment and Prevention. Am. J. Alzheimer’s Dis. Other Dementiasr 2013, 28, 120–125. [Google Scholar] [CrossRef]
- Schippling, S.; Kontush, A.; Arlt, S.; Buhmann, C.; Stürenburg, H.-J.; Mann, U.; Müller-Thomsen, T.; Beisiegel, U. Increased lipoprotein oxidation in alzheimer’s disease. Free. Radic. Biol. Med. 2000, 28, 351–360. [Google Scholar] [CrossRef]
- Tveden-Nyborg, P.; Vogt, L.; Schjoldager, J.G.; Jeannet, N.; Hasselholt, S.; Paidi, M.D.; Christen, S.; Lykkesfeldt, J. Maternal Vitamin C Deficiency during Pregnancy Persistently Impairs Hippocampal Neurogenesis in Offspring of Guinea Pigs. PLOS ONE 2012, 7, e48488. [Google Scholar] [CrossRef]
- Harrison, F.E.; Green, R.J.; Dawes, S.M.; May, J.M. Vitamin C distribution and retention in the mouse brain. Brain Res. 2010, 1348, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Milby, K.; Oke, A.; Adams, R. Detailed mapping of ascorbate distribution in rat brain. Neurosci. Lett. 1982, 28, 169–174. [Google Scholar] [CrossRef]
- Tsukaguchi, H.; Tokui, T.; MacKenzie, B.; Berger, U.V.; Chen, X.-Z.; Wang, Y.; Brubaker, R.F.; Hediger, M.A. A family of mammalian Na+-dependent L-ascorbic acid transporters. Nat. Cell Biol. 1999, 399, 70–75. [Google Scholar] [CrossRef]
- Mun, G.H.; Kim, M.J.; Lee, J.H.; Kim, H.J.; Chung, Y.H.; Chung, Y.B.; Kang, J.S.; Hwang, Y.I.; Oh, S.H.; Kim, J.-G.; et al. Immunohistochemical study of the distribution of sodium-dependent vitamin C transporters in adult rat brain. J. Neurosci. Res. 2006, 83, 919–928. [Google Scholar] [CrossRef] [PubMed]
- Qiu, S.; Li, L.; Weeber, E.J.; May, J.M. Ascorbate transport by primary cultured neurons and its role in neuronal function and protection against excitotoxicity. J. Neurosci. Res. 2007, 85, 1046–1056. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teafatiller, T.; Heskett, C.W.; Agrawal, A.; Marchant, J.S.; Baulch, J.E.; Acharya, M.M.; Subramanian, V.S. Upregulation of Vitamin C Transporter Functional Expression in 5xFAD Mouse Intestine. Nutrients 2021, 13, 617. https://doi.org/10.3390/nu13020617
Teafatiller T, Heskett CW, Agrawal A, Marchant JS, Baulch JE, Acharya MM, Subramanian VS. Upregulation of Vitamin C Transporter Functional Expression in 5xFAD Mouse Intestine. Nutrients. 2021; 13(2):617. https://doi.org/10.3390/nu13020617
Chicago/Turabian StyleTeafatiller, Trevor, Christopher W. Heskett, Anshu Agrawal, Jonathan S. Marchant, Janet E. Baulch, Munjal M. Acharya, and Veedamali S. Subramanian. 2021. "Upregulation of Vitamin C Transporter Functional Expression in 5xFAD Mouse Intestine" Nutrients 13, no. 2: 617. https://doi.org/10.3390/nu13020617
APA StyleTeafatiller, T., Heskett, C. W., Agrawal, A., Marchant, J. S., Baulch, J. E., Acharya, M. M., & Subramanian, V. S. (2021). Upregulation of Vitamin C Transporter Functional Expression in 5xFAD Mouse Intestine. Nutrients, 13(2), 617. https://doi.org/10.3390/nu13020617