
Behavioral and Metabolic Effects of a Calorie-Restricted Cafeteria Diet and Oleuropein Supplementation in Obese Male Rats

 ,
,  , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
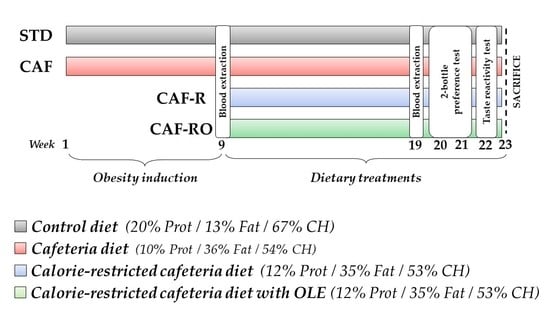
2.1. Animals and General Procedures
2.2. Diets
2.3. Oleuropein Supplementation
2.4. Body Mass Index
2.5. Serum Analyses
2.6. Behavioural Procedures
2.6.1. Two-Bottle Preference Test
2.6.2. Taste Reactivity Test
2.7. Statistical Analyses
3. Results
3.1. Effects of Diets on the Biometric, Food Intake, and Serum Parameters
3.2. Two-Bottle Preference Test
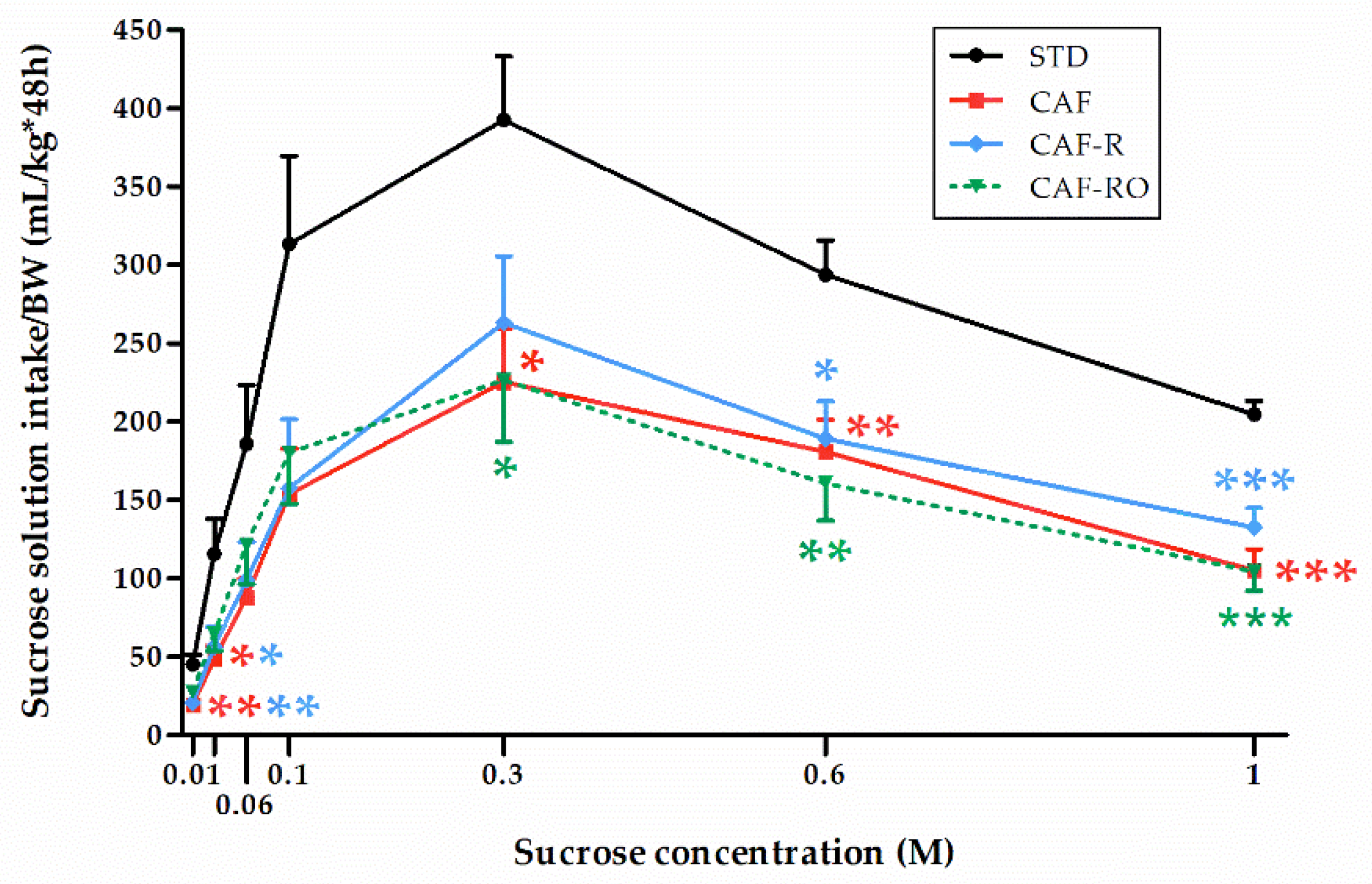
3.2.1. Sucrose Solution Intake and Preference
3.2.2. Total Fluid and Total Sucrose Intakes
3.3. Taste Reactivity Test
3.4. Correlations between Serum Leptin Levels and Sucrose Preference and Total Sucrose Intake
4. Discussion
5. Conclusions and Final Remarks
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Leitner, D.R.; Frühbeck, G.; Yumuk, V.; Schindler, K.; Micic, D.; Woodward, E.; Toplak, H. Obesity and type 2 diabetes: Two diseases with a need for combined treatment strategies—EASO can lead the way. Obes. Facts 2017, 10, 483–492. [Google Scholar] [CrossRef]
- World Health Organization. Obesity and Overweight. Key Facts. Available online: www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 25 October 2021).
- Bartoshuk, L.M.; Duffy, V.B.; Hayes, J.E.; Moskowitz, H.R.; Snyder, D.J. Psychophysics of sweet and fat perception in obesity: Problems, solutions and new perspectives. Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 1137–1148. [Google Scholar] [CrossRef]
- Melis, M.; Pintus, S.; Mastinu, M.; Fantola, G.; Moroni, R.; Pepino, M.Y.; Tomassini Barbarossa, I. Changes of Taste, Smell and Eating Behavior in Patients Undergoing Bariatric Surgery: Associations with PROP Phenotypes and Polymorphisms in the Odorant-Binding Protein OBPIIa and CD36 Receptor Genes. Nutrients 2021, 13, 250. [Google Scholar] [CrossRef]
- Trius-Soler, M.; Santillán-Alarcón, D.A.; Martínez-Huélamo, M.; Lamuela-Raventós, R.M.; Moreno, J.J. Effect of physiological factors, pathologies, and acquired habits on the sweet taste threshold: A systematic review and meta-analysis. Compr. Rev. Food Sci. Food Saf. 2020, 19, 3755–3773. [Google Scholar] [CrossRef]
- Gutierrez, R.; Fonseca, E.; Simon, S.A. The neuroscience of sugars in taste, gut-reward, feeding circuits, and obesity. Cell. Mol. Life Sci. 2020, 77, 3469–3502. [Google Scholar] [CrossRef]
- May, C.E.; Dus, M. Confection Confusion: Interplay Between Diet, Taste, and Nutrition. Trends Endocrinol. Metab. 2021, 32, 95–105. [Google Scholar] [CrossRef]
- Horio, N.; Jyotaki, M.; Yoshida, R.; Sanematsu, K.; Shigemura, N.; Ninomiya, Y. New Frontiers in Gut Nutrient Sensor Research: Nutrient Sensors in the Gastrointestinal Tract: Modulation of Sweet Taste Sensitivity by Leptin. J. Pharmacol. Sci. 2010, 112, 8–12. [Google Scholar] [CrossRef]
- Yoshida, R.; Niki, M.; Jyotaki, M.; Sanematsu, K.; Shigemura, N.; Ninomiya, Y. Modulation of sweet responses of taste receptor cells. Semin. Cell Dev. Biol. 2013, 24, 226–231. [Google Scholar] [CrossRef]
- Belza, A.; Frandsen, E.; Kondrup, J. Body fat loss achieved by stimulation of thermogenesis by a combination of bioactive food ingredients: A placebo-controlled, double-blind 8-week intervention in obese subjects. Int. J. Obes. 2007, 31, 121–130. [Google Scholar] [CrossRef]
- Llaha, F.; Zamora-Ros, R. The Effects of Polyphenol Supplementation in Addition to Calorie Restricted Diets and/or Physical Activity on Body Composition Parameters: A Systematic Review of Randomized Trials. Front. Nutr. 2020, 7, 84. [Google Scholar] [CrossRef] [PubMed]
- Konstantinidi, M.; Koutelidakis, A.E. Functional Foods and Bioactive Compounds: A Review of Its Possible Role on Weight Management and Obesity’s Metabolic Consequences. Medicines 2019, 6, 94. [Google Scholar] [CrossRef]
- Ahamad, J.; Toufeeq, I.; Khan, M.A.; Ameen, M.S.M.; Anwer, E.T.; Uthirapathy, S.; Mir, S.R.; Ahmad, J. Oleuropein: A natural antioxidant molecule in the treatment of metabolic syndrome. Phyther. Res. 2019, 33, 3112–3128. [Google Scholar] [CrossRef]
- Castejón, M.L.; Montoya, T.; Alarcón-de-la-Lastra, C.; Sánchez-Hidalgo, M. Potential Protective Role Exerted by Secoiridoids from Olea europaea L. in Cancer, Cardiovascular, Neurodegenerative, Aging-Related, and Immunoinflammatory Diseases. Antioxidants 2020, 9, 149. [Google Scholar] [CrossRef]
- Kleinert, M.; Clemmensen, C.; Hofmann, S.M.; Moore, M.C.; Renner, S.; Woods, S.C.; Huypens, P.; Beckers, J.; de Angelis, M.H.; Schürmann, A.; et al. Animal models of obesity and diabetes mellitus. Nat. Rev. Endocrinol. 2018, 14, 140–162. [Google Scholar] [CrossRef]
- Lalanza, J.F.; Caimari, A.; del Bas, J.M.; Torregrosa, D.; Cigarroa, I.; Pallàs, M.; Capdevila, L.; Arola, L.; Escorihuela, R.M. Effects of a post-weaning cafeteria diet in young rats: Metabolic syndrome, reduced activity and low anxiety-like behaviour. PLoS ONE 2014, 9, e85049. [Google Scholar] [CrossRef]
- Lalanza, J.F.; Snoeren, E.M.S. The cafeteria diet: A standardized protocol and its effects on behavior. Neurosci. Biobehav. Rev. 2021, 122, 92–119. [Google Scholar] [CrossRef]
- Leigh, S.-J.; Kendig, M.D.; Morris, M.J. Palatable Western-style Cafeteria Diet as a Reliable Method for Modeling Diet-induced Obesity in Rodents. J. Vis. Exp. 2019, 153, e60262. [Google Scholar] [CrossRef]
- Sampey, B.P.; Vanhoose, A.M.; Winfield, H.M.; Freemerman, A.J.; Muehlbauer, M.J.; Fueger, P.T.; Newgard, C.B.; Makowski, L. Cafeteria Diet Is a Robust Model of Human Metabolic Syndrome with Liver and Adipose Inflammation: Comparison to High-Fat Diet. Obesity 2011, 19, 1109–1117. [Google Scholar] [CrossRef] [PubMed]
- Caimari, A.; del Bas, J.M.; Boqué, N.; Crescenti, A.; Puiggròs, F.; Chenoll, E.; Martorell, P.; Ramón, D.; Genovés, S.; Arola, L. Heat-killed Bifidobacterium animalis subsp. Lactis CECT 8145 increases lean mass and ameliorates metabolic syndrome in cafeteria-fed obese rats. J. Funct. Foods 2017, 38, 251–263. [Google Scholar] [CrossRef]
- Mayneris-Perxachs, J.; Alcaide-Hidalgo, J.M.; de la Hera, E.; del Bas, J.M.; Arola, L.; Caimari, A. Supplementation with biscuits enriched with hesperidin and naringenin is associated with an improvement of the Metabolic Syndrome induced by a cafeteria diet in rats. J. Funct. Foods 2019, 61, 103504. [Google Scholar] [CrossRef]
- Lepore, S.M.; Morittu, V.M.; Celano, M.; Trimboli, F.; Oliverio, M.; Procopio, A.; Di Loreto, C.; Damante, G.; Britti, D.; Bulotta, S.; et al. Oral Administration of Oleuropein and Its Semisynthetic Peracetylated Derivative Prevents Hepatic Steatosis, Hyperinsulinemia, and Weight Gain in Mice Fed with High Fat Cafeteria Diet. Int. J. Endocrinol. 2015, 2015, 431453. [Google Scholar] [CrossRef]
- Kuem, N.; Song, S.J.; Yu, R.; Yun, J.W.; Park, T. Oleuropein attenuates visceral adiposity in high-fat diet-induced obese mice through the modulation of WNT10b- and galanin-mediated signalings. Mol. Nutr. Food Res. 2014, 58, 2166–2176. [Google Scholar] [CrossRef]
- Poudyal, H.; Campbell, F.; Brown, L. Olive leaf extract attenuates cardiac, hepatic, and metabolic changes in high carbohydrate-, high fat-fed rats. J. Nutr. 2010, 140, 946–953. [Google Scholar] [CrossRef]
- Ebaid, G.M.X.; Seiva, F.R.F.; Rocha, K.K.H.R.; Souza, G.A.; Novelli, E.L.B. Effects of olive oil and its minor phenolic constituents on obesity-induced cardiac metabolic changes. Nutr. J. 2010, 9, 46. [Google Scholar] [CrossRef]
- Araki, R.; Fujie, K.; Yuine, N.; Watabe, Y.; Nakata, Y.; Suzuki, H.; Isoda, H.; Hashimoto, K. Olive leaf tea is beneficial for lipid metabolism in adults with prediabetes: An exploratory randomized controlled trial. Nutr. Res. 2019, 67, 60–66. [Google Scholar] [CrossRef]
- Subias-Gusils, A.; Boqué, N.; Caimari, A.; Del Bas, J.M.; Mariné-Casadó, R.; Solanas, M.; Escorihuela, R.M. A restricted cafeteria diet ameliorates biometric and metabolic profile in a rat diet-induced obesity model. Int. J. Food Sci. Nutr. 2021, 71, 767–780. [Google Scholar] [CrossRef]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and β-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef]
- Loney, G.C.; Torregrossa, A.M.; Smith, J.C.; Sclafani, A.; Eckel, L.A. Rats display a robust bimodal preference profile for sucralose. Chem. Senses 2011, 36, 733–745. [Google Scholar] [CrossRef]
- Berridge, K.C. Measuring hedonic impact in animals and infants: Microstructure of affective taste reactivity patterns. Neurosci. Biobehav. Rev. 2000, 24, 173–198. [Google Scholar] [CrossRef]
- Shin, A.C.; Townsend, R.L.; Patterson, L.M.; Berthoud, H.-R. “Liking” and “wanting” of sweet and oily food stimuli as affected by high-fat diet-induced obesity, weight loss, leptin, and genetic predisposition. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, R1267–R1280. [Google Scholar] [CrossRef]
- Vezza, T.; Rodríguez-Nogales, A.; Algieri, F.; Garrido-Mesa, J.; Romero, M.; Sánchez, M.; Toral, M.; Martín-García, B.; Gómez-Caravaca, A.M.; Arráez-Román, D.; et al. The metabolic and vascular protective effects of olive (Olea europaea L.) leaf extract in diet-induced obesity in mice are related to the amelioration of gut microbiota dysbiosis and to its immunomodulatory properties. Pharmacol. Res. 2019, 150, 104487. [Google Scholar] [CrossRef] [PubMed]
- van der Stelt, I.; Hoek-van den Hil, E.F.; Swarts, H.J.M.; Vervoort, J.J.M.; Hoving, L.; Skaltsounis, L.; Lemonakis, N.; Andreadou, I.; van Schothorst, E.M.; Keijer, J. Nutraceutical oleuropein supplementation prevents high fat diet-induced adiposity in mice. J. Funct. Foods 2015, 14, 702–715. [Google Scholar] [CrossRef]
- Duca, F.A.; Swartz, T.D.; Covasa, M. Effect of Diet on Preference and Intake of Sucrose in Obese Prone and Resistant Rats. PLoS ONE 2014, 9, e111232. [Google Scholar] [CrossRef]
- Gac, L.; Kanaly, V.; Ramirez, V.; Teske, J.A.; Pinto, M.P.; Perez-Leighton, C.E. Behavioral characterization of a model of differential susceptibility to obesity induced by standard and personalized cafeteria diet feeding. Physiol. Behav. 2015, 152, 315–322. [Google Scholar] [CrossRef]
- Bernard, A.; Ancel, D.; Neyrinck, A.M.; Dastugue, A.; Bindels, L.B.; Delzenne, N.M.; Besnard, P. A preventive prebiotic supplementation improves the sweet taste perception in diet-induced obese mice. Nutrients 2019, 11, 549. [Google Scholar] [CrossRef]
- Maliphol, A.B.; Garth, D.J.; Medler, K.F. Diet-induced obesity reduces the responsiveness of the peripheral taste receptor cells. PLoS ONE 2013, 8, e79403. [Google Scholar] [CrossRef]
- Chen, K.; Yan, J.; Suo, Y.; Li, J.; Wang, Q.; Lv, B. Nutritional status alters saccharin intake and sweet receptor mRNA expression in rat taste buds. Brain Res. 2010, 1325, 53–62. [Google Scholar] [CrossRef]
- Casper, R.C.; Sullivan, E.L.; Tecott, L. Relevance of animal models to human eating disorders and obesity. Psychopharmacology 2008, 199, 313–329. [Google Scholar] [CrossRef] [PubMed]
- Goldschmidt, A.B.; Jones, M.; Manwaring, J.L.; Luce, K.H.; Osborne, M.I.; Cunning, D.; Taylor, K.L.; Doyle, A.C.; Wilfley, D.E.; Taylor, C.B. The clinical significance of loss of control over eating in overweight adolescents. Int. J. Eat. Disord. 2008, 41, 153–158. [Google Scholar] [CrossRef]
- Stafford, L.D.; Whittle, A. Obese individuals have higher preference and sensitivity to odor of chocolate. Chem. Senses 2015, 40, 279–284. [Google Scholar] [CrossRef]
- Connolly, L.; Coveleskie, K.; Kilpatrick, L.A.; Labus, J.S.; Ebrat, B.; Stains, J.; Jiang, Z.; Tillisch, K.; Raybould, H.E.; Mayer, E.A. Differences in brain responses between lean and obese women to a sweetened drink. Neurogastroenterol. Motil. 2013, 25, 579-e460. [Google Scholar] [CrossRef]
- Hajnal, A.; Norgren, R. Accumbens dopamine mechanisms in sucrose intake. Brain Res. 2001, 904, 76–84. [Google Scholar] [CrossRef]
- Hakim, J.D.; Keay, K.A. Prolonged ad libitum access to low-concentration sucrose changes the neurochemistry of the nucleus accumbens in male Sprague-Dawley rats. Physiol. Behav. 2019, 201, 95–103. [Google Scholar] [CrossRef]
- Muscat, R.; Kyprianou, T.; Osman, M.; Phillips, G.; Willner, P. Sweetness-dependent facilitation of sucrose drinking by raclopride is unrelated to calorie content. Pharmacol. Biochem. Behav. 1991, 40, 209–213. [Google Scholar] [CrossRef]
- de Araujo, I.E.; Oliveira-Maia, A.J.; Sotnikova, T.D.; Gainetdinov, R.R.; Caron, M.G.; Nicolelis, M.A.L.; Simon, S.A. Food Reward in the Absence of Taste Receptor Signaling. Neuron 2008, 57, 930–941. [Google Scholar] [CrossRef]
- Cabanac, M. Physiological Role of Pleasure. Science 1971, 173, 1103–1107. [Google Scholar] [CrossRef]
- Cabanac, M. Sensory Pleasure. Q. Rev. Biol. 1979, 54, 1–29. [Google Scholar] [CrossRef]
- Laeng, B.; Berridge, K.C.; Butter, C.M. Pleasantness of a Sweet Taste during Hunger and Satiety: Effects of Gender and “Sweet Tooth”. Appetite 1993, 21, 247–254. [Google Scholar] [CrossRef]
- Berridge, K.C. Modulation of taste affect by hunger, caloric satiety, and sensory-specific satiety in the rat. Appetite 1991, 16, 103–120. [Google Scholar] [CrossRef]
- Grill, H.J.; Roitman, M.F.; Kaplan, J.M. A new taste reactivity analysis of the integration of taste and physiological state information. Am. J. Physiol. Integr. Comp. Physiol. 1996, 271, R677–R687. [Google Scholar] [CrossRef]
- Johnson, P.M.; Kenny, P.J. Dopamine D2 receptors in addiction-like reward dysfunction and compulsive eating in obese rats. Nat. Neurosci. 2010, 13, 635–641. [Google Scholar] [CrossRef]
- Vucetic, Z.; Kimmel, J.; Reyes, T.M. Chronic high-fat diet drives postnatal epigenetic regulation of-opioid receptor in the brain. Neuropsychopharmacology 2011, 36, 1199–1206. [Google Scholar] [CrossRef]
- Volkow, N.D.; Wang, G.-J.; Tomasi, D.; Baler, R.D. Obesity and addiction: Neurobiological overlaps. Obes. Rev. 2013, 14, 2–18. [Google Scholar] [CrossRef]
- Arcego, D.M.; Krolow, R.; Lampert, C.; Toniazzo, A.P.; dos Santos Garcia, E.; Lazzaretti, C.; Costa, G.; Scorza, C.; Dalmaz, C. Chronic high-fat diet affects food-motivated behavior and hedonic systems in the nucleus accumbens of male rats. Appetite 2020, 153, 104739. [Google Scholar] [CrossRef]
- Dutheil, S.; Ota, K.T.; Wohleb, E.S.; Rasmussen, K.; Duman, R.S. High-Fat Diet Induced Anxiety and Anhedonia: Impact on Brain Homeostasis and Inflammation. Neuropsychopharmacology 2016, 41, 1874–1887. [Google Scholar] [CrossRef]
- Blum, K.; Thanos, P.K.; Gold, M.S. Dopamine and glucose, obesity, and reward deficiency syndrome. Front. Psychol. 2014, 5, 919. [Google Scholar] [CrossRef]
- Achat, S.; Tomao, V.; Madani, K.; Chibane, M.; Elmaataoui, M.; Dangles, O.; Chemat, F. Direct enrichment of olive oil in oleuropein by ultrasound-assisted maceration at laboratory and pilot plant scale. Ultrason. Sonochem. 2012, 19, 777–786. [Google Scholar] [CrossRef]
- Hirai, M.; Kobori, R.; Doge, R.; Tsuji, I.; Saito, A. Efficient Concentration of Functional Polyphenols Using Their Interaction with Gelatin. Foods 2021, 10, 698. [Google Scholar] [CrossRef]
- Bonacci, S.; Paonessa, R.; Costanzo, P.; Salerno, R.; Maiuolo, J.; Nardi, M.; Procopio, A.; Manuela, O. Peracetylation as a strategy to improve oleuropein stability and its affinity to fatty foods. Food Funct. 2018, 9, 5759–5767. [Google Scholar] [CrossRef]
- Alirezaei, M.; Rezaei, M.; Hajighahramani, S.; Sookhtehzari, A.; Kiani, K. Oleuropein attenuates cognitive dysfunction and oxidative stress induced by some anesthetic drugs in the hippocampal area of rats. J. Physiol. Sci. 2017, 67, 131–139. [Google Scholar] [CrossRef]
- Sueishi, Y.; Nii, R. A comparative study of the antioxidant profiles of olive fruit and leaf extracts against five reactive oxygen species as measured with a multiple free-radical scavenging method. J. Food Sci. 2020, 85, 2737–2744. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, R.; Margolskee, R.F.; Ninomiya, Y. Phosphatidylinositol-3 kinase mediates the sweet suppressive effect of leptin in mouse taste cells. J. Neurochem. 2021, 158, 233–245. [Google Scholar] [CrossRef]
- Kawai, K.; Sugimoto, K.; Nakashima, K.; Miura, H.; Ninomiya, Y. Leptin as a modulator of sweet taste sensitivities in mice. Proc. Natl. Acad. Sci. USA 2000, 97, 11044–11049. [Google Scholar] [CrossRef] [PubMed]
- Shigemura, N.; Ohta, R.; Kusakabe, Y.; Miura, H.; Hino, A.; Koyano, K.; Nakashima, K.; Ninomiya, Y. Leptin Modulates Behavioral Responses to Sweet Substances by Influencing Peripheral Taste Structures. Endocrinology 2004, 145, 839–847. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CAF Diet-Induced Obesity Period | Dietary Treatments Period | |||||
|---|---|---|---|---|---|---|
| STD | CAF | STD | CAF | CAF-R | CAF-RO | |
| Biometric parameters | Weeks 1–8 | Weeks 9–19 | ||||
| Body weight gain (g) | 255 ± 6 | 313 ± 5 +++ | 121 ± 5 | 158 ± 9 ** | 124 ± 5 $$ | 132 ± 6 $$ |
| BMI (g/cm2) 1 | 0.64 ± 0.01 | 0.70 ± 0.01 ++ | 0.74 ± 0.01 | 0.83 ± 0.01 *** | 0.78 ± 0.01 $ | 0.78 ± 0.02 $ |
| Daily food intake parameters | ||||||
| Food intake (g/kg) | 101 ± 1 | 309 ± 2 +++ | 46.3 ± 0.6 | 140 ± 4 *** | 77.2 ± 2.1 *** $$$ | 75.9 ± 1.3 *** $$$ |
| Energy intake (kcal/kg) | 292 ± 3 | 667 ± 5 +++ | 134 ± 2 | 289 ± 6 *** | 183 ± 4 *** $$$ | 177 ± 4 *** $$$ |
| Chow intake (kcal/kg) | 292 ± 3 | 123 ± 3 +++ | 134 ± 2 | 44.1 ± 2.9 *** | 58.8 ± 4.2 *** $ | 53.8 ± 4.5 *** |
| Simple sugars (kcal/kg) | 196 ± 2 | 93.7 ± 3.6 | 36.8 ± 1.3 $$$ | 36.7 ± 1.1 $$$ | ||
| Serum parameters | Week 9 | Week 19 | ||||
| Triacylglycerides (mmol/L) | 0.93 ± 0.06 | 1.31 ± 0.09 + | 1.07 ± 0.08 | 1.79 ± 0.22 ** | 1.58 ± 0.14 * | 1.54 ± 0.13 |
| Insulin (ng/mL) | 0.51 ± 0.07 | 0.70 ± 0.05 + | 0.61 ± 0.08 | 1.03 ± 0.17 * | 0.82 ± 0.06 | 0.83 ± 0.10 |
| HOMA-IR | 3.20 ± 0.54 | 4.73 ± 0.37 + | 3.68 ± 0.57 | 6.92 ± 1.31 * | 5.65 ± 0.58 * | 5.36 ± 0.76 |
| Leptin (ng/mL) | 4.23 ± 0.40 | 12.5 ± 1.0 +++ | 7.05 ± 0.78 | 23.7 ± 4.3 *** | 19.1 ± 1.9 *** | 17.0 ± 2.0 *** |
| 0.01 M | 0.03 M | 0.06 M | 0.1 M | 0.3 M | 0.6 M | 1 M | |
|---|---|---|---|---|---|---|---|
| rs value | −0.12 | −0.38 * | −0.26 | −0.13 | −0.13 | −0.44 ** | −0.45 ** |
| p value | 0.45 | 0.02 | 0.10 | 0.44 | 0.43 | 0.01 | 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Subias-Gusils, A.; Álvarez-Monell, A.; Boqué, N.; Caimari, A.; Del Bas, J.M.; Mariné-Casadó, R.; Solanas, M.; Escorihuela, R.M. Behavioral and Metabolic Effects of a Calorie-Restricted Cafeteria Diet and Oleuropein Supplementation in Obese Male Rats. Nutrients 2021, 13, 4474. https://doi.org/10.3390/nu13124474
Subias-Gusils A, Álvarez-Monell A, Boqué N, Caimari A, Del Bas JM, Mariné-Casadó R, Solanas M, Escorihuela RM. Behavioral and Metabolic Effects of a Calorie-Restricted Cafeteria Diet and Oleuropein Supplementation in Obese Male Rats. Nutrients. 2021; 13(12):4474. https://doi.org/10.3390/nu13124474
Chicago/Turabian StyleSubias-Gusils, Alex, Adam Álvarez-Monell, Noemí Boqué, Antoni Caimari, Josep M. Del Bas, Roger Mariné-Casadó, Montserrat Solanas, and Rosa M. Escorihuela. 2021. "Behavioral and Metabolic Effects of a Calorie-Restricted Cafeteria Diet and Oleuropein Supplementation in Obese Male Rats" Nutrients 13, no. 12: 4474. https://doi.org/10.3390/nu13124474
APA StyleSubias-Gusils, A., Álvarez-Monell, A., Boqué, N., Caimari, A., Del Bas, J. M., Mariné-Casadó, R., Solanas, M., & Escorihuela, R. M. (2021). Behavioral and Metabolic Effects of a Calorie-Restricted Cafeteria Diet and Oleuropein Supplementation in Obese Male Rats. Nutrients, 13(12), 4474. https://doi.org/10.3390/nu13124474