
Dietary Flavone Baicalein Combinate with Genipin Attenuates Inflammation Stimulated by Lipopolysaccharide in RAW264.7 Cells or Pseudomonas aeruginosa in Mice via Regulating the Expression and Phosphorylation of AKT

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
2.3. Target Fishing
2.4. Target Prediction
2.5. Microscale Thermophoresis (MST)
2.6. Fluorescence Quenching Assay (FQA)
2.7. Co-Immunoprecipitation
2.8. Co-Localization Test
2.9. Combination Index Analysis
2.10. Western Blotting
2.11. Acute Lung Injury Test
2.12. Statistical Analysis
3. Results
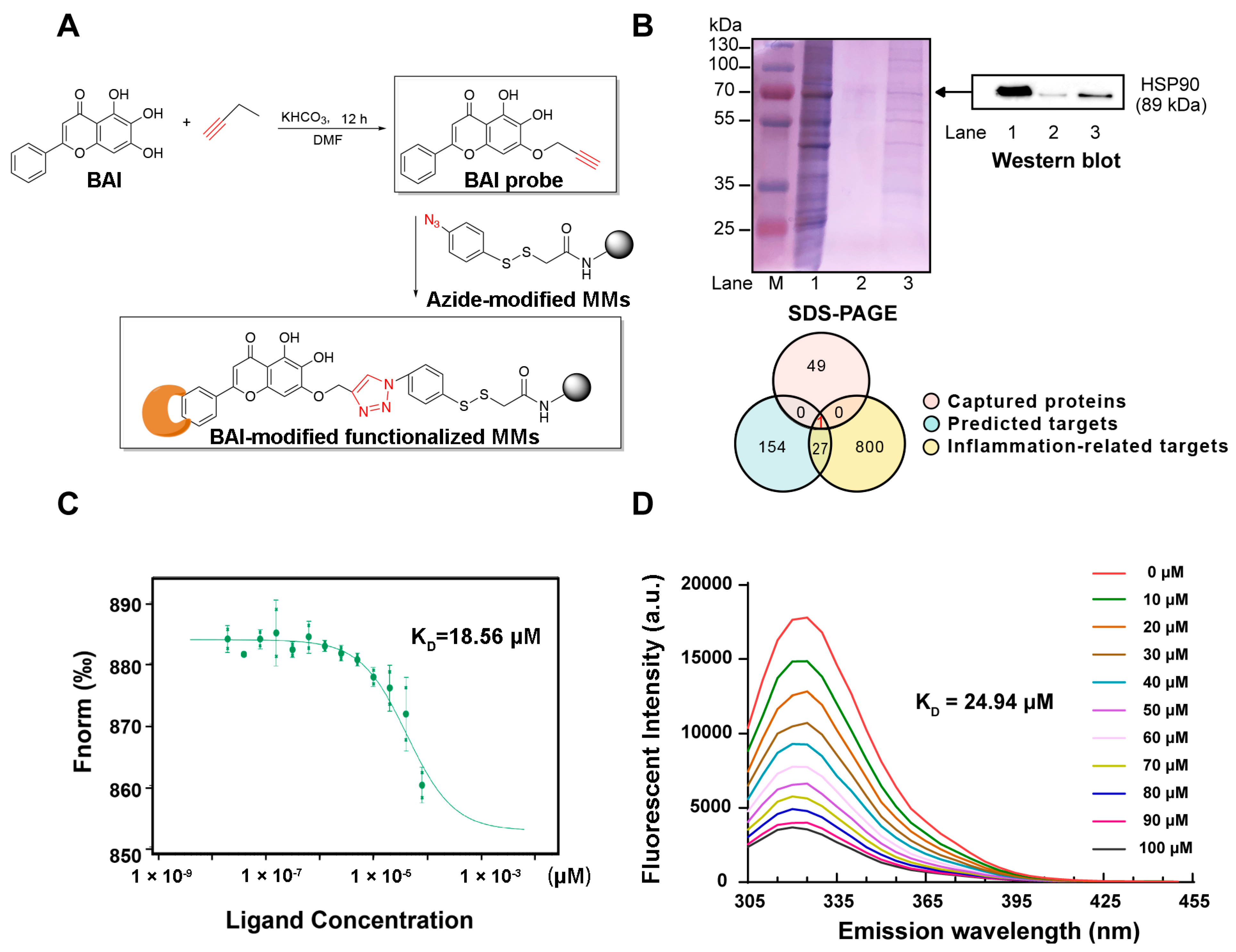
3.1. HSP90 as a Potential Target of BAI
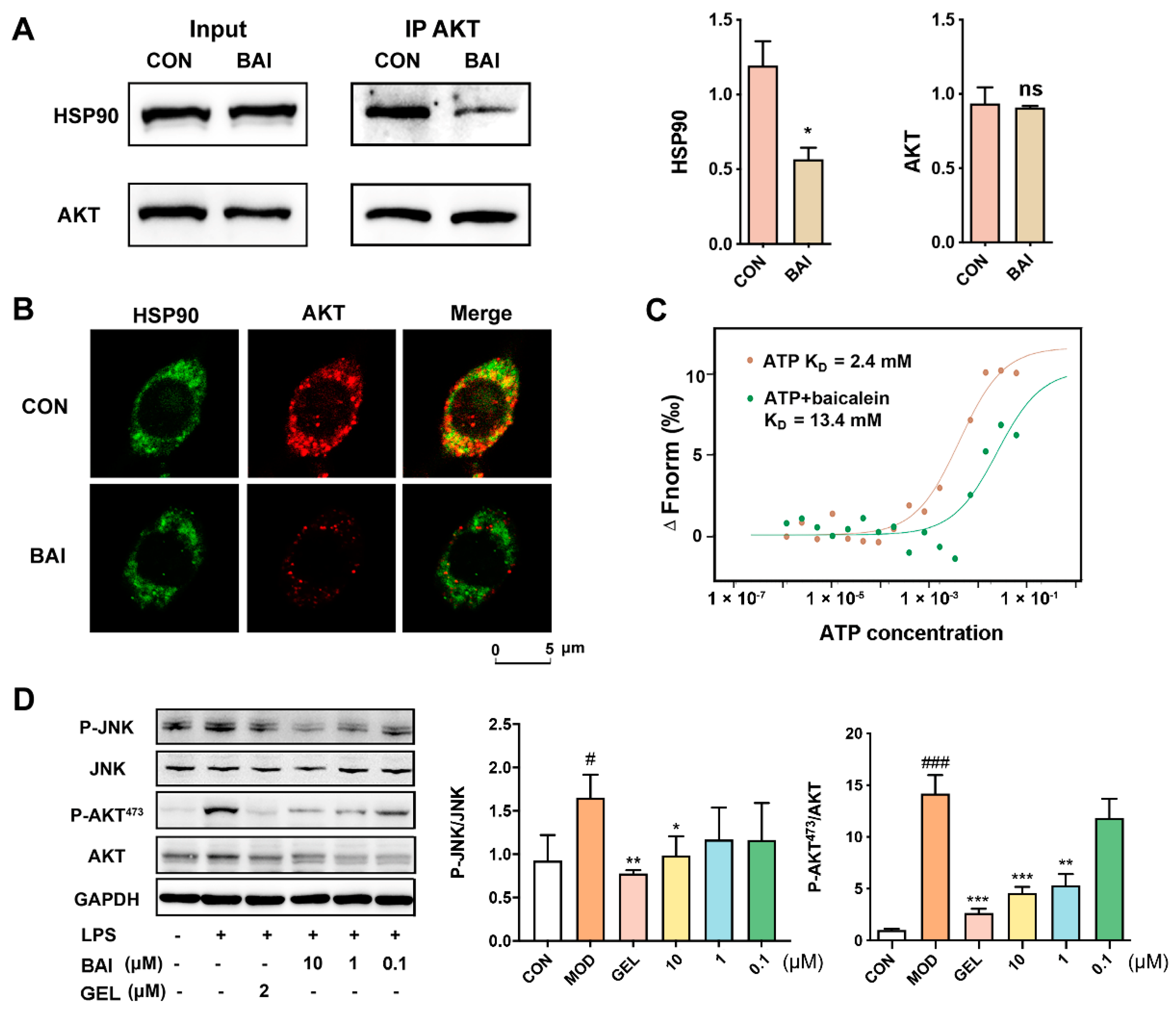
3.2. BAI Induced AKT Degradation and Decreased P-AKT
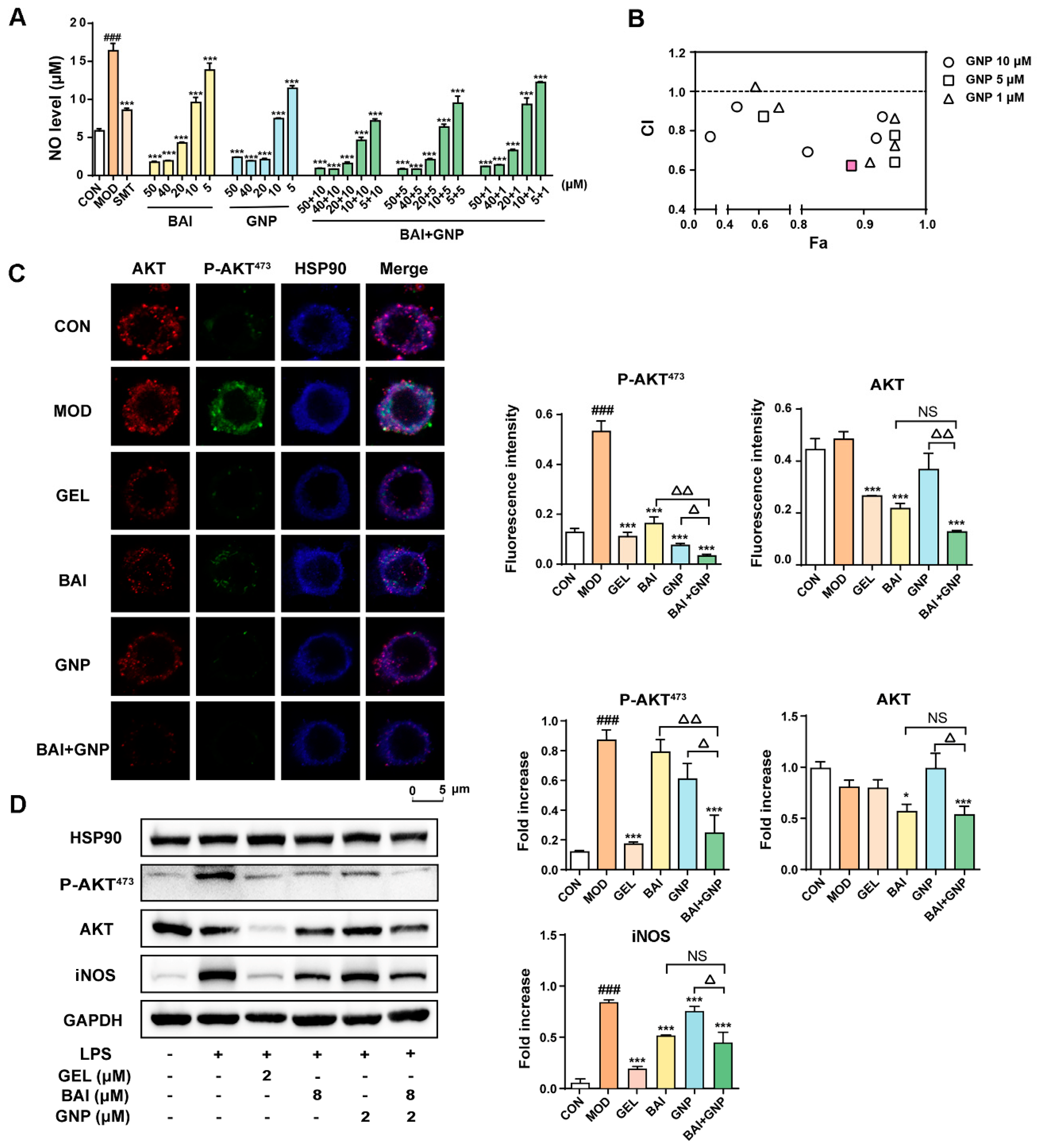
3.3. The CombInation of BAI and GNP Enhanced the Inhibition of AKT Activation
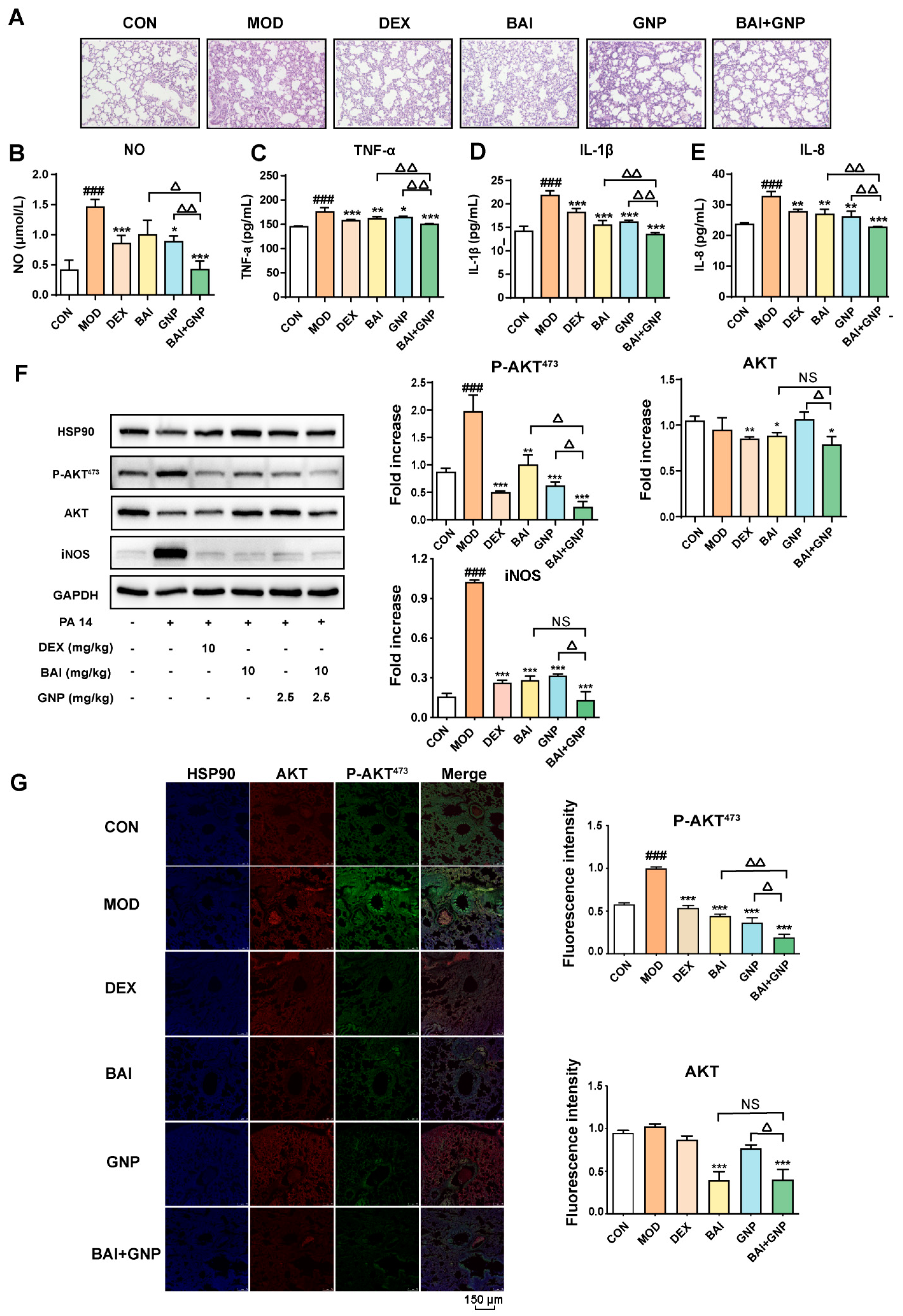
3.4. The Combination of BAI and GNP Alleviated Bacteria-Induced Acute Lung Injury in Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tang, Y.; Liu, J.; Zhang, D.; Xu, Z.; Ji, J.; Wen, C. Cytokine Storm in COVID-19: The Current Evidence and Treatment Strategies. Front. Immunol. 2020, 11, 1708. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, Y.; Qiao, W.; Zhang, J.; Qi, Z. Baricitinib, a drug with potential effect to prevent SARS-CoV-2 from entering target cells and control cytokine storm induced by COVID-19. Int. Immunopharmacol. 2020, 86, 106749. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.W.; Hu, X.; Jin, P.F. Cytokine Storm Induced by SARS-CoV-2 and the Drug Therapy. Chin. Pharmacol. J. 2020, 24, 333–336. [Google Scholar]
- Soy, M.; Keser, G.; Atagündüz, P.; Tabak, F.; Kayhan, S. Cytokine storm in COVID-19: Pathogenesis and overview of anti-inflammatory agents used in treatment. Clin. Rheumatol. 2020, 39, 2085–2094. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Hussain, T.; Murtaza, G.; Yang, H.; Kalhoro, M.; Kalhoro, D. Exploiting Anti-Inflammation Effects of Flavonoids in Chronic Inflammatory Diseases. Curr. Pharm. Design 2020, 26, 2610–2619. [Google Scholar] [CrossRef] [PubMed]
- Yeung, Y.T.; Aziz, F.; Guerrero-Castilla, A.; Arguelles, S. Signaling Pathways in Inflammation and Anti-Inflammatory Therapies. Curr Pharm Design. 2018, 24, 1449–1484. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Luo, L.; Namani, A.; Wang, X.J.; Tang, X. Nrf2 Signaling Pathway: Pivotal Roles in Inflammation. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef]
- Zusso, M.; Lunardi, V.; Franceschini, D.; Pagetta, A.; Lo, R.; Stifani, S.; Frigo, A.C.; Giusti, P.; Moro, S. Ciprofloxacin and levofloxacin attenuate microglia inflammatory response via TLR4/NF-kB pathway. J. Neuroinflammat. 2019, 16, 148. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.-D.; Zhang, J.-L.; Wang, X.-Y.; Hu, Z.-W.; Qian, Y.-B. The JAK2/STAT3 signaling pathway is required for inflammation and cell death induced by cerulein in AR42J cells. Eur. Rev. Med. Pharmacol Sci. 2019, 23, 1770–1777. [Google Scholar] [PubMed]
- Cianciulli, A.; Calvello, R.; Porro, C.; Trotta, T.; Salvatore, R.; Panaro, M.A. PI3k/Akt signalling pathway plays a crucial role in the anti-inflammatory effects of curcumin in LPS-activated microglia. Int. Immunopharmacol. 2016, 36, 282–290. [Google Scholar] [CrossRef]
- Cui, J.; Zhang, F.; Wang, Y.; Liu, J.; Ming, X.; Hou, J.; Lv, B.; Fang, S.; Yu, B. Macrophage migration inhibitory factor promotes cardiac stem cell proliferation and endothelial differentiation through the activation of the PI3K/Akt/mTOR and AMPK pathways. Int. J. Mol. Med. 2016, 37, 1229–1309. [Google Scholar] [CrossRef]
- He, S.Q.; Gao, M.; Fu, Y.F.; Zhang, Y.N. Glycyrrhizic acid inhibits leukemia cell growth and migration via blocking AKT/mTOR/STAT3 signaling. Int. J. Clin. Exp. Pathol. 2015, 8, 5175–5181. [Google Scholar] [PubMed]
- Duan, M.X.; Zhou, H.; Wu, Q.Q.; Liu, C.; Xiao, Y.; Deng, W.; Tang, Q.Z. Andrographolide Protects against HG-Induced Inflammation, Apoptosis, Migration, and Impairment of Angiogenesis via PI3K/AKT-eNOS Signalling in HUVECs. Mediat. Inflamm. 2019, 6168340. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Phong, B.; Wilson, D.C.; Hirsch, R.; Kane, L.P. Akt fine-tunes NF-κB-dependent gene expression during T cell activation. J. Biol. Chem. 2011, 286, 36076–36085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, T.; Ju, J.; Mo, L.; Ma, L.A.; Hu, W.; You, R.; Chen, X.; Chen, Y.; Liu, Z.; Qiu, S. Anti-inflammation action of xanthones from Swertia chirayita by regulating COX-2/NF-κB/MAPKs/Akt signaling pathways in RAW 264.7 macrophage cells. Phytomedicine 2019, 55, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Hassouni, B.E.; Mantini, G.; Petri, G.L.; Capula, M.; Peters, G.J. To Combine or Not Combine: Drug Interactions and Tools for Their Analysis. Reflections from the EORTC-PAMM Course on Preclinical and Early-phase Clinical Pharmacology. Anticancer Res. 2019, 39, 3303–3309. [Google Scholar] [CrossRef]
- Bush, K.; Bradford, P.A. β-Lactams and β-Lactamase Inhibitors: An Overview. Cold Spring Harb Perspect Med. 2016, 6, a025247. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Fu, J.N.; Chou, T.C. Synergistic combination of microtubule targeting anticancer fludelone with cytoprotective panaxytriol derived from panax ginseng against MX-1 cells in vitro: Experimental design and data analysis using the combination index method. Am. J. Cancer Res. 2016, 6, 97–104. [Google Scholar]
- Cao, B.; Wei, X.C.; Xu, X.R.; Zhang, H.Z.; Zhang, D.K. Seeing the Unseen of the Combination of Two Natural Resins, Frankincense and Myrrh: Changes in Chemical Constituents and Pharmacological Activities. Molecules 2019, 24, 3076. [Google Scholar] [CrossRef] [Green Version]
- Kandil, E.; Tsumagari, K.; Ma, J.; Abd Elmageed, Z.Y.; Li, X.; Slakey, D.; Mondal, D.; Abdel-Mageed, A.B. Synergistic inhibition of thyroid cancer by suppressing MAPK/PI3K/AKT pathways. J. Surg. Res. 2013, 184, 898–906. [Google Scholar] [CrossRef]
- Chang, C.H.; Chen, Y.S.; Chiou, M.T.; Su, C.H.; Hsu, Y.M. Application of Scutellariae radix, Gardeniae fructus, and Probiotics to Prevent Salmonella enterica Serovar Choleraesuis Infection in Swine. Evid Based Complement. Altern. Med. 2013, 2013, 568528. [Google Scholar] [CrossRef]
- Yu, X.; Liu, Y.; Wang, Y.; Mao, X.; Zhang, Y.; Xia, J. Baicalein induces cervical cancer apoptosis through the NF-κB signaling pathway. Mol. Med. Rep. 2018, 17, 5088–5094. [Google Scholar] [CrossRef]
- Dinda, B.; Dinda, S.; Dassharma, S.; Banik, R.; Chakraborty, A.; Dinda, M. Therapeutic potentials of baicalin and its aglycone, baicalein against inflammatory disorders. Eur. J. Med. Chem. 2017, 131, 68–80. [Google Scholar] [CrossRef]
- Li, X.; Hu, Y.F.; Wu, J.S.; Wang, L.; Huang, W.G.; Xu, C.S.; Meng, X.L.; Wang, P. Chen-Si Semi-Mechanism-Based Pharmacodynamic Model for the Anti-Inflammatory Effect of Baicalein in LPS-Stimulated RAW264.7 Macrophages. Front. Pharmacol. 2018, 9, 793. [Google Scholar]
- Yan, C.; Liu, T.; Wang, K.; Hou, C.; Cai, S.; Huang, Y.; Du, Z.; Huang, H.; Kong, J.; Chen, Y. Baicalein Inhibits Staphylococcus aureus Biofilm Formation and the Quorum Sensing System In Vitro. PLoS ONE 2016, 11, e0153468. [Google Scholar]
- Yan, W.J.; Ma, X.C.; Zhao, X.Y.; Zhang, S.Q. Baicalein induces apoptosis and autophagy of breast cancer cells via inhibiting PI3K/AKT pathway in vivo and vitro. Drug Des. Devel Ther. 2018, 12, 3961–3972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.S.; Lee, C.J.; Jin, Y.M. Enhancement of active compound, genipin, from Gardeniae Fructus using immobilized glycosyl hydrolase family 3 β-glucosidase from Lactobacillus antri. AMB Express 2017, 7, 64. [Google Scholar] [CrossRef] [Green Version]
- Koo, H.J.; Yun, S.S.; Kim, H.J.; Lee, Y.H.; Hong, S.M.; Kim, S.J.; Kim, B.C.; Jin, C.; Lim, C.J.; Park, E.H. Antiinflammatory effects of genipin, an active principle of gardenia. Eur. J. Pharmacol. 2004, 495, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, L.; Liang, Z.; Li, Y.; Yuan, F.; Liu, J.; Tian, Y.; Hao, Z.; Zhou, F.; Liu, X. Genipin Inhibits LPS-Induced Inflammatory Response in BV2 Microglial Cells. Neurochem. Res. 2017, 42, 2769–2776. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.C.; Tsai, S.Y.; Lai, P.Y.; Chen, Y.S.; Chao, P. Metabolism and pharmacokinetics of genipin and geniposide in rats. Food Chem. Toxicol. 2008, 46, 2764–2769. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Wang, Y.; Yao, W.; Xue, Y.; Liu, Z. Geniposide attenuates neonatal mouse brain injury after hypoxic-ischemia involving the activation of PI3K/Akt signaling pathway. J. Chem. Neuroanat. 2019, 102, 101687. [Google Scholar] [CrossRef]
- Lin, W.H.; Kuo, H.H.; Ho, L.H.; Tseng, M.L.; Siao, A.C.; Hung, C.T.; Jeng, K.C.; Hou, C.W. Gardenia jasminoides extracts and gallic acid inhibit lipopolysaccharide-induced inflammation by suppression of JNK2/1 signaling pathways in BV-2 cells. Iran. J. Basic Med. Sci. 2015, 18, 555–562. [Google Scholar]
- Zhang, Z.J.; Peng, L.; Wang, Z.; Li, P.T.; Zhang, W.S.; Sun, Z.H.; Zhang, X.J.; Wang, Y.Y. A comparative study on the individual and combined effects of baicalin and jasminoidin on focal cerebral ischemia-reperfusion injury. Brain Res. 2006, 1123, 188–195. [Google Scholar] [CrossRef]
- Cui, Q.; Hou, Y.; Hou, J.; Pan, P.; Li, L.Y.; Bai, G.; Luo, G. Preparation of functionalized alkynyl magnetic microspheres for the selective enrichment of cell glycoproteins based on click chemistry. Biomacromolecules 2013, 14, 124–131. [Google Scholar] [CrossRef]
- Cheng, C.J.; Liu, K.X.; Zhang, M.; Shen, F.K.; Bai, G. Okicamelliaside targets the N-terminal chaperone pocket of HSP90 disrupts the chaperone protein interaction of HSP90-CDC37 and exerts antitumor activity. Acta Pharmacol. Sin. 2021. online ahead of print. [Google Scholar] [CrossRef]
- Zhang, M.; Ma, X.; Xu, H.; Wu, W.; He, X.; Wang, X.; Jiang, M.; Hou, Y.; Bai, G. A natural AKT inhibitor swertiamarin targets AKT-PH domain, inhibits downstream signaling, and alleviates inflammation. FEBS J. 2020, 287, 1816–1829. [Google Scholar] [CrossRef] [PubMed]
- Dickey, C.A.; Koren, J.; Zhang, Y.J.; Xu, Y.F.; Jinwal, U.K.; Birnbaum, M.J.; Monks, B.; Mei, S.; Cheng, J.Q.; Patterson, C. Akt and CHIP coregulate tau degradation through coordinated interactions. Proc. Natl. Acad. Sci. USA 2008, 105, 3622–3627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stebbins, C.E.; Russo, A.A.; Schneider, C.; Rosen, N.; Rosen, F.U.; Pavletich, N.P. Crystal Structure of an Hsp90–Geldanamycin Complex: Targeting of a Protein Chaperone by an Antitumor Agent. Cell 1997, 89, 239–250. [Google Scholar] [CrossRef] [Green Version]
- Jiao, Y.; Shen, F.; Wang, Z.; Ye, L.; Bai, G. Genipin, a natural AKT inhibitor, targets the PH domain to affect downstream signaling and alleviates inflammation. Biochem. Pharmacol. 2019, 170, 113660. [Google Scholar] [CrossRef]
- Triantafilou, K.; Triantafilou, M.; Dedrick, R.L. A CD14-independent LPS receptor cluster. Nat. Immunol. 2001, 2, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Tarassishin, L.; Suh, H.S.; Lee, S.C. LPS and IL-1 differentially activate mouse and human astrocytes: Role of CD14. Glia 2014, 62, 999–1013. [Google Scholar] [CrossRef] [Green Version]
- Lester, S.N.; Li, K. Toll-Like Receptors in Antiviral Innate Immunity. J. Molecular Biol. 2013, 426, 1246–1264. [Google Scholar] [CrossRef] [PubMed]
- Manning, B.D.; Cantley, L.C. AKT/PKB signaling: Navigating downstream. Cell 2007, 129, 1261–1274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, K.Y.; Lee, H.J.; Woo, H.; Kang, R.J.; Hoe, H.S. Dasatinib regulates LPS-induced microglial and astrocytic neuroinflammatory responses by inhibiting AKT/STAT3 signaling. J. Neuroinflammat. 2019, 16, 190. [Google Scholar] [CrossRef]
- Riggs, D.L.; Cox, M.B.; Cheung-Flynn, J.; Prapapanich, V.; Carrigan, P.E.; Smith, D.F. Functional Specificity of Co-Chaperone Interactions with Hsp90 Client Proteins. Crit. Rev. Biochem. Mol. Biol. 2004, 39, 279–295. [Google Scholar] [CrossRef]
- Luke, W.; Susan, L. Lindquist HSP90 and the chaperoning of cancer. Nat. Rev. Cancer 2005, 5, 761–772. [Google Scholar]
- Prodromou Mechanisms of Hsp90 regulation. Biochem. J. 2016, 473, 2439–2452. [CrossRef] [Green Version]
- Pearl, L.; Prodromou, C.; Workman, P. The Hsp90 molecular chaperone: An open and shut case for treatment. Biochem. J. 2008, 410, 439–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartl, F.U.; Hayer-Hart, M. Molecular Chaperones in the Cytosol: From Nascent Chain to Folded Protein. Science 2002, 295, 1852–1860. [Google Scholar] [CrossRef] [Green Version]
- Rowlands, M.; Mcandrew, C.; Prodromou, C.; Pearl, L.; Kalusa, A.; Jones, K.; Workman, P.; Aherne, W. Detection of the ATPase Activity of the Molecular Chaperones Hsp90 and Hsp72 Using the Transcreener ADP Assay Kit. J. Biomol. Screen. 2010, 15, 279–286. [Google Scholar] [CrossRef] [Green Version]
- Sato, S.; Fujita, N.; Tsuruo, T. Modulation of Akt kinase activity by binding to Hsp90. Proc. Natl. Acad. Sci. USA 2000, 97, 10832–10837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Facchinetti, V.; Ouyang, W.M.; Wei, H.; Soto, N.; Lazorchak, A.; Gould, C.; Lowry, C.; Newton, A.C.; Mao, Y.; Miao, R.Q.; et al. The mammalian target of rapamycin complex 2 controls folding and stability of Akt and protein kinase C. EMBO J. 2008, 27, 1932–1943. [Google Scholar] [CrossRef] [PubMed]
- Uko, N.E.; Güner, O.; Matesic, D.F.; Bowen, J.P. Akt Pathway Inhibitors. Curr. Top. Med. Chem. 2020, 20, 883–900. [Google Scholar] [CrossRef]
- Liu, W.; Chu, G.; Chang, N.; Ma, X.; Jiang, M.; Bai, G. Phillygenin attenuates inflammatory responses and influences glucose metabolic parameters by inhibiting Akt activity. RSC Adv. 2017, 7, 40418–40426. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Sampath, D.; Nannini, M.A.; Lee, B.B.; Degtyarev, M.; Oeh, J.; Savage, H.; Guan, Z.; Hong, R.; Kassees, R. Targeting activated Akt with GDC-0068, a novel selective Akt inhibitor that is efficacious in multiple tumor models. Clin. Cancer Res. 2013, 19, 1760–1772. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.; Sanderson, P.E.; Zheng, W. Drug combination therapy increases successful drug repositioning. Drug Discov. Today 2016, 21, 1189–1195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dey, P. Targeting Gut Barrier Dysfunction with Phytotherapies: Effective Strategy Against Chronic Diseases. Pharmacol. Res. 2020, 161, 105135. [Google Scholar] [CrossRef] [PubMed]
- Dey, P. Gut Microbiota in Phytopharmacology: A Comprehensive Overview of Concepts, Reciprocal Interactions, Biotransformations And Mode of Actions. Pharmacol. Res. 2019, 147, 104367. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Ye, L.; Cheng, C.; Shen, F.; Niu, L.; Hou, Y.; Bai, G. Dietary Flavone Baicalein Combinate with Genipin Attenuates Inflammation Stimulated by Lipopolysaccharide in RAW264.7 Cells or Pseudomonas aeruginosa in Mice via Regulating the Expression and Phosphorylation of AKT. Nutrients 2021, 13, 4462. https://doi.org/10.3390/nu13124462
Zhang M, Ye L, Cheng C, Shen F, Niu L, Hou Y, Bai G. Dietary Flavone Baicalein Combinate with Genipin Attenuates Inflammation Stimulated by Lipopolysaccharide in RAW264.7 Cells or Pseudomonas aeruginosa in Mice via Regulating the Expression and Phosphorylation of AKT. Nutrients. 2021; 13(12):4462. https://doi.org/10.3390/nu13124462
Chicago/Turabian StyleZhang, Man, Lili Ye, Chuanjing Cheng, Fukui Shen, Lin Niu, Yuanyuan Hou, and Gang Bai. 2021. "Dietary Flavone Baicalein Combinate with Genipin Attenuates Inflammation Stimulated by Lipopolysaccharide in RAW264.7 Cells or Pseudomonas aeruginosa in Mice via Regulating the Expression and Phosphorylation of AKT" Nutrients 13, no. 12: 4462. https://doi.org/10.3390/nu13124462
APA StyleZhang, M., Ye, L., Cheng, C., Shen, F., Niu, L., Hou, Y., & Bai, G. (2021). Dietary Flavone Baicalein Combinate with Genipin Attenuates Inflammation Stimulated by Lipopolysaccharide in RAW264.7 Cells or Pseudomonas aeruginosa in Mice via Regulating the Expression and Phosphorylation of AKT. Nutrients, 13(12), 4462. https://doi.org/10.3390/nu13124462