Onco-Preventive and Chemo-Protective Effects of Apple Bioactive Compounds

 , , ,
, , ,  and
and
Abstract
1. Introduction
2. Cancer
3. Importance of Phytochemicals from Diet in Management of Cancer
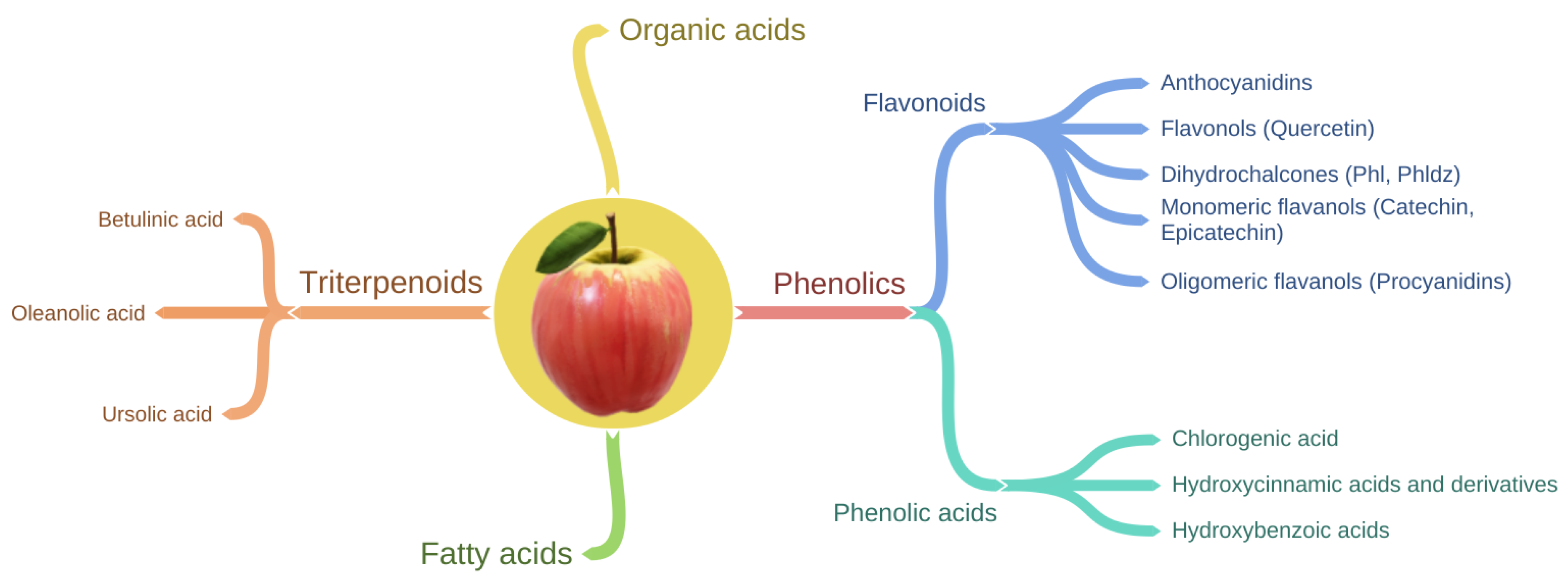
4. Apple Phytochemical Profile and Bioavailability
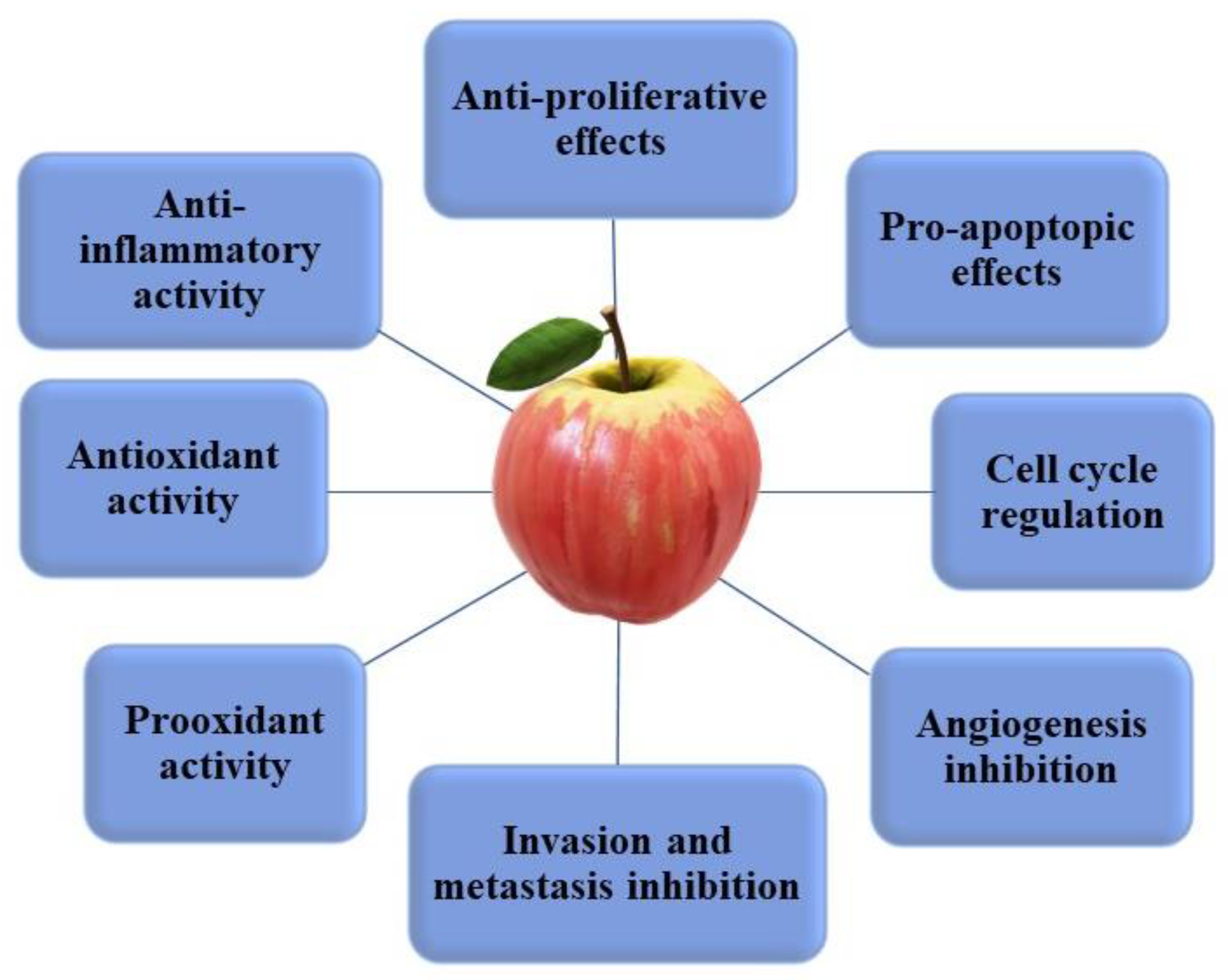
5. Health Benefits of Apple Phytochemicals: Cancer
5.1. Epidemiological Evidence of Apple Consumption and Cancer Incidence
5.2. In Vitro and In Vivo Evidence of the Anticancer Properties of Apple Phytochemicals
5.1.1. Quercetin Anticancer Properties
5.1.2. Phloretin and Phloridzin Anticancer Properties
5.1.3. Chlorogenic Acid Anticancer Properties
5.1.4. Catechins and Epicatechins Anticancer Properties
5.1.5. Procyanidins Anticancer Properties
5.1.6. Triterpenoids Anticancer Properties

{kind=link}
{kind=link}
{kind=link}
| In Vitro | Effect | Expression Markers Affected | Effective Concentration | Cell Line | Ref. |
|---|---|---|---|---|---|
| QUERCETIN | Lung cancer | ||||
| Anti-proliferative | ↓ PDK3 | 55.90 ± 2.25 µM | A549 | [180] | |
| Anti-proliferative, pro-apoptotic, autophagy inhibition | ↑LC3-II, SIRT 1, AMPK, beclin 1 ↓ p62 | 100 µM | A549, H1299 | [181] | |
| Breast cancer | |||||
| Anti-proliferative, cell cycle arrest | ↑cyclin B1 and CDK-1 ↓p21 | 10 µM | SK-BR3, MDA-MB-453 | [175] | |
| Anti-proliferative, pro-apoptotic | ↑ miR-146a, bax, caspase-3 ↓ EGFR | 80 µM/mL, 50 µM/mL (respectively) | MCF-7, MDA-MB-231 | [177] | |
| Anti-proliferative, pro-apoptotic | ↓ survivin | 40 mg/mL | MCF-7 | [178] | |
| Pro-apoptotic, cell cycle arrest | ↓ Foxo3a, p53, GADD45 | 20 µM | MDA-MB-231 | [182] | |
| Metastasis and invasion inhibition | ↓ MMP-2,9, VEGF, PKM2, GLUT1, LDHA, Akt, mTOR | 30 µM | MCF-7, MDA-MB-231 | [186] | |
| Angiogenesis inhibition | ↓ VEGF, Pin1 | 30 µM | MCF-7 | [188] | |
| Colorectal cancer | |||||
| Metastasis and invasion inhibition, anti-inflammatory | ↑ E-cadherin ↓ MMP-2,9, p65, TLR4, TNF-α, COX-2, IL-6 | 5,10, 20 μM | Caco-2 | [187] | |
| Liver cancer | |||||
| Anti-proliferative | ↓ PDK3 | 49.10 ± 1.45 µM | HepG2 | [180] | |
| Ovarian cancer | |||||
| Pro-apoptotic | ↑ phospho-eIF2α, p53 ↓ Rad51 | 25, 50, 75, 100 µM | OV2008, A2780, GM9607 | [183] | |
| Anti-proliferative, pro-apoptotic, cell cycle arrest | ↓ survivin | 30 mg/ml | SKOV-3 | [179] | |
| Prostate cancer | |||||
| Anti-proliferative, angiogenesis inhibition | ↓ Akt, mTOR, VEGFR2, S6 kinase | 10-40 mmol/L | HUVECs | [189] | |
| Retinoblastoma | |||||
| Angiogenesis inhibition | ↓ VEGFR | 25, 50, 200 µM | Y79 | [190] | |
| PHLORETIN AND PHLORIDZIN | Lung cancer | ||||
| Anti-proliferative, pro-apoptotic, invasion and migration inhibition | ↑ caspase-3,-9 ↓ Bcl-2, MMP-2,-9 | 25, 50, 75 µg/mL | A549, H838, H520, Calu-1 | [205] | |
| Pro-apoptotic, cell cycle arrest | ↑ Bax, caspase-3, -9 ↓ Bcl-2 | 50, 100, 200 µM | A549 | [199] | |
| Breast cancer | |||||
| Anti-proliferative, cell cycle arrest | ↓ GLUT-2 | 25, 50, 100, 150 µM | MDA-MB-231 | [195] | |
| Anti-proliferative, autophagy inhibition | ↓ mTOR, ULK1, LC3B-II | 100, 200 µM | MDA-MB-231, MCF7, ERα+ | [204] | |
| Colorectal cancer | |||||
| Anti-proliferative, cell cycle arrest | ↑ E-cadherin, p53 ↓ GLUT-2 | 100, 200 µM | Colo 205, HT-29 | [196] | |
| Pro-apoptotic | ↑ caspase-3,-7, -9, Bax, cytochrome C ↓ Bcl-2 | 100 μmol/L | HT-29 | [206] | |
| Anti-inflammatory | ↓ PGE2, IL-8, AGEs | 50 μM | CCD-18Co | [207] | |
| Liver cancer | |||||
| Pro-apoptotic, invasion and migration inhibition | ↓ GLUT-2, Bcl-2, Akt | 200 μM | HepG2 | [197] | |
| Pro-apoptotic | ↑ SHP-1 ↓ p-Akt, pERK, mTOR, VEGFR2, p-JNK | 25, 50, 100 μM | SK-Hep1, Hep3B2.1-7, Huh7, PLC5, HepG2 | [201] | |
| Prostate cancer | |||||
| Prooxidant, anti-proliferative, migration inhibition | ↑ ROS ↓ β-catenin, TCF4, FoxA2, c-Myc, CISD2 | 20, 50, 100 μM | PC3, DU145 | [200] | |
| Gastric cancer | |||||
| Anti-proliferative, pro-apoptotic, cell cycle arrest | ↓ p-JNK, p-38 | 4, 8, 16 μM | AGS | [198] | |
| Esophageal cancer | |||||
| Anti-proliferative, pro-apoptotic | ↑ BAX, p53 ↓ Bcl-2 | 60, 70, 80, 90, 100 μg/mL | EC-109 | [203] | |
| Brain cancer | |||||
| Anti-proliferative, pro-apoptotic, cell cycle arrest | ↑ p27 ↓ CDK-2,-4,-6, cyclin-D,-E | 100, 200, 300 μM | U87, U251 | [202] | |
| CHLOROGENIC ACID | Lung cancer | ||||
| Anti-proliferative, cell cycle arrest | ↑ p21, p53, and KHSRP ↓ c-Myc, miR-17 family | 25, 50 μM | H446 | [214] | |
| Anti-proliferative, pro-apoptotic | ↓ cIAP1, cIAP2, binding of annexin A2 to p50 and actin => ↓ NF-κB | 25, 50, 100, 200, 400, 800 μM | A549 | [220] | |
| Anti-proliferative, pro-apoptotic | ↑ Bax, caspase-3, p38, JNK, annexin V ↓ Bcl-2, SOX2 | 30, 50 μM | A549 | [215] | |
| Breast cancer | |||||
| Anti-proliferative, pro-apoptotic, migration and invasion inhibition | ↓ annexin, NF-κB, p65 | 10,20 μM | MDA-MB-231, MDA-MB-453 | [221] | |
| Colorectal cancer | |||||
| Anti-proliferative, cell cycle arrest, prooxidant | ↑ ROS, p53 ↓ ERK | 125, 250, 500, 1000 μmol/L | HCT116, HT29 | [217] | |
| Cell cycle arrest, pro-apoptotic | ↑ caspase-3 | 250, 500, 1000 μM | Caco-2 | [219] | |
| Liver cancer | |||||
| Anti-proliferative, cell cycle arrest, invasion, and metastasis inhibition | ↓ MMP-2,-9, ERK1/2 | 250, 500, 1000 μM | HepG2 | [216] | |
| Anti-proliferative, cell cycle arrest | ↑ p21, p53, and KHSRP ↓ c-Myc, miR-17 family | 25, 50 μM | Huh7 | [214] | |
| Kidney cancer | |||||
| Anti-proliferative, pro-apoptotic | ↑ caspase, Bax ↓ Bcl-2, PI3K, Akt, mTOR | 40 μM | A498 | [26] | |
| Osteosarcoma | |||||
| Anti-proliferative, pro-apoptotic, cell cycle arrest | ↑ ERK1/2 | 200, 400 μM | U2OS, Saos-2 | [218] | |
| CATECHIN AND EPICATECHIN | Breast cancer | ||||
| Pro-apoptotic | ↑ ZIP9 ↓ cAMP agonists to membrane androgen receptors | 200 nM | MDA-MB-468 | [230] | |
| Pro-apoptotic | agonists to membrane androgen receptors | 21.4 nM | T47D | [231] | |
| Anti-proliferative, pro-apoptotic, antioxidant | ↑ IRK ↓ ROS | 40, 100 μg/mL | MCF-10A | [227] | |
| Pro-apoptotic, prooxidant | ↑ ROS, Bad, Bax | 150, 200, 250, 300, 350, 400, 450, 500 μM | MDA-MB-231 | [232] | |
| Colorectal cancer | |||||
| Pro-apoptotic, migration and invasion reduction | ↑ E-cadherin ↓ ERK1/2, c-Myc, β-catenin | 12.5, 20 μM | HT-29 | [233] | |
| Liver cancer | |||||
| Anti-proliferative, cell cycle arrest | ↑ p21, waf1/cip1 ↓ CDC25A | 50, 75, 100, 125, 150 μM | HepG2, Huh7 | [226] | |
| Biliary tract cancer | |||||
| Pro-apoptotic, cell cycle arrest | ↑ caspase, p21, gene dr5 | 20, 50 μM | CCSW-1, BDC, EGI-1, SkChA-1, TFK-1, MzChA-1, MzChA-2, GBC | [229] | |
| Prostate cancer | |||||
| Pro-apoptotic | ↑ ZIP9 ↓ cAMP agonists to membrane androgen receptors | 200 nM | PC-3 | [230] | |
| Pancreatic cancer | |||||
| Anti-proliferative, pro-apoptotic, cell cycle arrest | ↑ Bax ↓ Ras, NF-κB, p65, Bcl-2, Pi3K, Akt | 25, 50 μM | E6E7-Kras-st | [228] | |
| PROCYANIDINS | Breast cancer | ||||
| Pro-apoptotic | ↑ cytochrome-c, caspase-3,-9 | 25 μg/m | B16, BALB-MC.E12 | [239] | |
| Pro-apoptotic, migration and invasion reduction | ↑ maspin, E-cadherin, BRCA1 ↓ DNA methyltransferases | 50, 100, 150, 200, 250 μM | MDA-MB-231 | [243] | |
| Pro-apoptotic | n/a | 50 μM | MCF-7 | [244] | |
| Pro-apoptotic, cell cycle arrest | ↑ Bax, caspase-3,-9 ↓ Bcl-2 | 31.5, 36.6 mg/mL | MDA-MB-231, MCF-7 | [274] | |
| Colorectal cancer | |||||
| Anti-proliferative, pro-apoptotic, cell cycle arrest | ↑ caspase-3, ERK1/2, JNK ↓ PKC | 45 μg/mL | SW620 | [238] | |
| Anti-proliferative, pro-apoptotic, cell cycle arrest | ↑ MMP-2,-9, caspase-3,-9, ERK 1/2, MEK, Akt, PI3K ↓ EGFR | 10–60 μM | Caco-2, HT-29, HCT-15, HCT-116 | [240] | |
| Pro-apoptotic, prooxidant | ↑ caspase-3,-8,-9, Bax, ROS, cytochrome-c ↓Bcl-2 | 80 µg/mL | SW480 and SW620 | [246] | |
| Anti-proliferative, cell cycle arrest, pro-apoptotic | ↑ ERK1/2, MEK, PI3K, Akt ↓ EGFR | 10, 20, 30 μM | Caco-2 | [241] | |
| Pro-apoptotic | ↑ PKB, Akt, ERK1/2, p38 | 2.5–20 μM | Caco-2 | [275] | |
| Pro-apoptotic | ↑ caspase-3,-9, cytochrome-c ↓ PI3K, Akt, bad | 2.5–50 μM | Caco-2 | [276] | |
| Liver cancer | |||||
| Anti-proliferative, migration inhibition | ↓ Kv10.1 | 10, 100, 1000 μM | HepG2 | [242] | |
| Prostate cancer | |||||
| Pro-apoptotic, prooxidant | ↑ ROS, ERK1/2, AMPKα | 25, 50 μM (PCa LNCaP); 50, 100, 200 μM (22Rv1) | PCa LNCaP, 22Rv1 | [245] | |
| TRITERPENOIDS | Breast cancer | ||||
| Anti-proliferative | n/a | n/a | MCF-7 | [260] | |
| Anti-proliferative, pro-apoptotic | ↑ Bax, cytochrome-c, p53 ↓ Bcl-2 | 2.57, 5.45 μM (respectively) | MDA-MB-231, MCF-7 | [254] | |
| Anti-proliferative, migration and invasion inhibition | ↑ caspase-3 ↓ MMP-2,-9, TIMP-2 | 5, 10, 20 μM | MCF-7, 4T1, MDA-MB-231 | [256] | |
| Anti-proliferative, pro-apoptotic, migration and invasion inhibition | ↓ aerobic glycolysis, c-Myc, lactate dehydrogenase A (LDH-A), p-PDK1, Caveloin-1 | 48.55, 19.06 μM (respectively) | MDA-MB-231, MCF-7 | [255] | |
| Anti-proliferative, pro-apoptotic, migration and invasion inhibition | ↑ GRP78, PERK ↓ aerobic glycolysis, c-Myc, β-catenin | 5-50 μM | MDA-MB-231, BT-549, HBL-100 | [257] | |
| Anti-proliferative, pro-apoptotic, autophagy inhibition, anti-inflammatory | ↓ PI3K, Akt, NF-κB | 232, 221, 240 μg/mL (respectively) | T47D, MCF-7, MDA-MB-231 | [258] | |
| Cell cycle arrest, pro-apoptotic, autophagy | ↑ p53, p21, AMPK ↓ ERK1/2, glycolysis, PKM2, HK2 | 20 μM | MCF-7, MDA-MB-231, SK-BR-3 | [80] | |
| Lung cancer | |||||
| Pro-apoptotic, angiogenesis inhibition | ↑ Bax ↓ VEGF | 25, 50 μg/ml | A549, H460 | [270] | |
| Colorectal cancer | |||||
| Anti-proliferative | n/a | n/a | Caco-2 | [260] | |
| Invasion and metastasis inhibition | ↓ cadherins, integrins | 10, 20, 40, 80 μM | SW620 | [272] | |
| Liver cancer | |||||
| Anti-proliferative, prooxidant | ↑ ROS ↓ PI3K, Akt1, mTOR | 10, 30, 100 μM | [268] | ||
| Anti-proliferative | n/a | n/a | HepG2 | [260] | |
| Invasion and metastasis inhibition | ↓ cadherins, integrins | 10, 20, 40, 80 μM | HepG2 | [272] | |
| Anti-proliferative, pro-apoptotic | ↑ p53, caspase-3 ↓ Bcl-2, Mcl-a mRNA | 10, 20, 30 μM | HUH7, PLC/PRF/5, L02 | [265] | |
| Pancreatic cancer | |||||
| Cell cycle arrest, pro-apoptotic, autophagy induction | ↑ Bax, ATG5, LC3-II ↓ Bcl-2, RAGE | 25, 50, 75, 100 μM | MIA Paca-2 | [266] | |
| Prostate cancer | |||||
| Pro-apoptotic | ↑ cytochrome-c, PARP, p21, p53 ↓ NF-κB, Bcl-2, p65 | 10, 25 μM | LNCaP, DU145 | [263] | |
| Anti-proliferative, pro-apoptotic |
↑ survivin ↓ Bcl-2, Bcl-xl, survivin, PI3K, Akt, mTOR | LNCaP, PC-3 | [259] | ||
| Cervical cancer | |||||
| Pro-apoptotic, cell cycle arrest, prooxidant | ↑ ROS, p21, Bad, caspase-9 ↓ PI3K, Akt | 30 μmol/: | HeLa | [262] | |
| Ovarian cancer | |||||
| Anti-proliferative, pro-apoptotic | ↑ Bax, caspase-3,-8,-9 ↓ Bcl-2 | 44.47 μM | A2780 | [264] | |
| Gallbladder cancer | |||||
| Anti-proliferative, cell cycle arrest, pro-apoptotic | ↑ Bax, cytochrome-c, caspase-3,-9 ↓ Bcl-2 | 50 μmol/L | GBC-SD, NOZ | [267] | |
| Brain cancer | |||||
| Pro-apoptotic, migration and invasion reduction | ↑ JNK signaling pathway, caspases ↓ enzyme MGMT | 20 μM | U373MG | [273] | |
| Anti-proliferative, pro-apoptotic | ↓ enzyme MGMT, STAT3 | 20, 30, 40, 50 μM | LN229, LN18, T98G | [253] | |
| Osteosarcoma | |||||
| Anti-proliferative, pro-apoptotic, antioxidant | ↑ caspase-3 ↓ Notch signaling pathway, Bcl-2, ROS | 50, 80 μM (respectively) | Saos-2, MG63 | [269] | |
| Melanoma | |||||
| Invasion and metastasis inhibition | ↓ cadherins, integrins | 10, 20, 40, 80 μM | B16-F10 | [272] | |
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AGEs | Advanced glycation end products |
| Akt | Protein kinase B |
| AMPK | Activated protein kinase |
| ATG5 | Autophagy protein 5 |
| Bad | Bcl2 Associated Agonist Of Cell Death protein |
| Bax | Bcl2 Associated X protein |
| Bcl-2 | B-cell lymphoma 2 |
| BRCA1 | Breast cancer 1 gene |
| cAMP | Cyclic adenosine monophosphate |
| CDC25A | Cell division cycle 25 A gene |
| CDK-1,-2,-4,-6 | Cyclin dependent kinase |
| cIAP1,2 | Cellular Inhibitor of Apoptosis Protein 1 |
| cip1 | CDK- interacting protein |
| CISD2 | CISD2 gene |
| c-Myc | Cellular myelocytomatosis oncogene |
| COX-2 | Cyclooxygenase-2 |
| EGFR | Epidermal growth factor receptor |
| ERK, ERK1/2 | Extracellular regulated kinase |
| FoxA2 | Forkhead box protein A2 |
| FoxO3a | Forkhead box protein o3a |
| GADD45 | Growth arrest and DNA damage-inducible protein |
| Glut1 | Glucose transporter 1 |
| GRP78 (BiP) | Binding immunoglobulin protein |
| HK2 | Hexokinase-2 |
| IL-6,-8 | Interleukin-6,-8 |
| IRK | The arabidopsis receptor kinase |
| JNK | C-Jun N-terminal Kinase |
| KHSRP | Kh-type splicing regulatory protein |
| Kv10.1 | Kv10.1 potassium channel |
| LC3-II | Light chain membrane protein |
| LDHA | Lactate dehydrogenase A |
| LDH-A | Lactate dehydrogenase A |
| Mcl-1 | Induced myeloid leukemia cell differentiation protein |
| MEK | Mitogen-activated protein kinase kinase |
| MGMT | Methylated-DNA—protein-cysteine methyltransferase |
| miR-146a | Micro Ribonucleic Acid 146a |
| MMP-2,-9 | Matrix metalloproteinase-2,-9 |
| mTOR | Mammalian target of rapamycin protein |
| NF-κB | Nuclear Factor kappa B |
| p21, p65, p62, p53, p38, p27 | Protein 21,65,62,53,38,27 |
| PARP | Poly [ADP-ribose] polymerase 1 |
| PDK3,1 | Pyruvate dehydrogenase |
| pERK | R-like endoplasmic reticulum kinase |
| PGE2 | Prostaglandin E2 |
| Phosphor-eIF2α | Phosphorylation of eukaryotic initiation factor-2 alpha |
| PI3K | Phosphoinositide 3-kinase |
| Pin1 | Prolyl isomerase |
| p-JNK | Phosphorylated c-Jun N-terminal Kinase (JNK) |
| PKB | Protein kinase B |
| PKC | Protein kinase C |
| PKM2 | Pyruvate kinase m2 |
| rad51 | RAD51 Recombinase (gene) |
| RAGE | Advanced glycosylation end product-specific receptor |
| Ras | Ras protein |
| ROS | Reactive oxygen species |
| SHP-1 | Protein tyrosine phosphatase shp 1 |
| SIRT-1 | Sirtiuin 1 |
| SOX2 | Sry-box transcription factor 2 |
| STAT3 | Signal transducer and activator of transcription 3 |
| TCF4 | Transcription factor 4 (gene) |
| TIMP-2 | Tissue Inhibitor of Metalloproteinase 2 |
| TLR4 | Toll-like receptor 4 |
| TNF- α | Tumor necrosis factor alpha |
| ULK-1 | Unc-51 like autophagy activating kinase 1 |
| VEGF | Vascular endothelial growth factor |
| VEGFR | Vascular endothelial growth factor receptor |
| Waf1 | Wild type p53 activated protein-1 |
| ZIP9 | Zinc transporter 9 |
References
- Harris, R.E. Epidemiology of Chronic Disease: Global Perspectives; Jones & Bartlett Learning: Burlington, MA, USA, 2019. [Google Scholar]
- Wilkins, E.; Wilson, L.; Wickramasinghe, K.; Bhatnagar, P.; Leal, J.; Luengo-Fernandez, R.; Burns, R.; Rayner, M.; Townsend, N. European Cardiovascular Disease Statistics 2017; European Heart Network: Brussels, Belgium, 2017. [Google Scholar]
- Bowry, A.D.; Lewey, J.; Dugani, S.B.; Choudhry, N.K. The Burden of Cardiovascular Disease in Low- and Middle-Income Countries: Epidemiology and Management. Can. J. Cardiol. 2015, 31, 1151–1159. [Google Scholar] [CrossRef] [PubMed]
- Magliano, D.J.; Islam, R.M.; Barr, E.L.M.; Gregg, E.; Pavkov, M.E.; Harding, J.L.; Tabesh, M.; Koye, D.N.; Shaw, J.E. Trends in incidence of total or type 2 diabetes: Systematic review. BMJ 2019, 366, l5003. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, O.; Kayode, A.A.; Olusola, Y.O.; Joshua, A.I.; Chinedu, O.V. Lung Cancer: A Chronic Disease Epidemiology; Prevalence Study. Asian J. Adv. Res. Rep. 2019, 3, 1–7. [Google Scholar] [CrossRef]
- World Health Organization. Noncommunicable Diseases: Progress Monitor 2020; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Ferlay, J.; Ervik, M.; Lam, F.; Colombet, M.; Mery, L.; Piñeros, M.; Znaor, A.; Soerjomataram, I.; Bray, F. Global Cancer Observatory: Cancer Today; International Agency for Research on Cancer: Lyon, France, 2018; Available online: https://gco.iarc.fr/today (accessed on 23 July 2021).
- Reiss, R.; Johnston, J.; Tucker, K.; DeSesso, J.M.; Keen, C.L. Estimation of cancer risks and benefits associated with a potential increased consumption of fruits and vegetables. Food Chem. Toxicol. 2012, 50, 4421–4427. [Google Scholar] [CrossRef] [PubMed]
- Mathur, G.; Nain, S.; Sharma, P.K. Cancer: An Overview. Acad. J. Cancer Res. 2015, 8, 1. [Google Scholar]
- Sun, Y.S.; Zhao, Z.; Yang, Z.N.; Xu, F.; Lu, H.J.; Zhu, Z.Y.; Shi, W.; Jiang, J.; Yao, P.P.; Zhu, H.P. Risk factors and preventions of breast cancer. Int. J. Biol. Sci. 2017, 13, 1387. [Google Scholar] [CrossRef]
- Rawla, P.; Barsouk, A. Epidemiology of gastric cancer: Global trends, risk factors and prevention. Prz. Gastroenterol. 2019, 14, 26. [Google Scholar] [CrossRef]
- Botelho, M.C.; Teixeira, J.P.; Oliveira, P.A. Carcinogenesis. In Encyclopedia of Toxicology, 3rd ed.; Wexler, P., Ed.; Academic Press: Oxford, UK, 2014; pp. 713–729. [Google Scholar]
- Fan, A.M. Chapter 11—Cancer. In Information Resources in Toxicology, 4th ed.; Wexler, P., Gilbert, S.G., Hakkinen, P.J., Mohapatra, A., Eds.; Academic Press: San Diego, CA, USA, 2009; pp. 103–121. [Google Scholar]
- Basu, A.K. DNA damage, mutagenesis and cancer. Int. J. Mol. Sci. 2018, 19, 970. [Google Scholar] [CrossRef]
- Bajaj, J.; Diaz, E.; Reya, T. Stem cells in cancer initiation and progression. J. Cell Biol. 2019, 219, e201911053. [Google Scholar] [CrossRef]
- Mäki-Nevala, S.; Valo, S.; Ristimäki, A.; Sarhadi, V.; Knuutila, S.; Nyström, M.; Renkonen-Sinisalo, L.; Lepistö, A.; Mecklin, J.P.; Peltomäki, P. DNA methylation changes and somatic mutations as tumorigenic events in Lynch syndrome-associated adenomas retaining mismatch repair protein expression. EBioMedicine 2019, 39, 280–291. [Google Scholar] [CrossRef]
- Takeshima, H.; Ushijima, T. Accumulation of genetic and epigenetic alterations in normal cells and cancer risk. Npj Precis. Oncol. 2019, 3, 1–8. [Google Scholar] [CrossRef]
- Pogribny, I.P.; Rusyn, I. Environmental Toxicants, Epigenetics, and Cancer. In Epigenetic Alterations in Oncogenesis; Springer: New York, NY, USA, 2013; pp. 215–232. [Google Scholar]
- Botezatu, A.; Iancu, I.V.; Popa, O.; Plesa, A.; Manda, D.; Huica, I.; Vladoiu, S.; Anton, G.; Badiu, C. Mechanisms of Oncogene Activation. New Aspects in Molecular and Cellular Mechanisms of Human Carcinogenesis; Bulgin, D., Ed.; BoD—Books on Demand: Norderstedt, Germany, 2016; pp. 1–52. [Google Scholar]
- Kinsella, A.R. Multistage Carcinogenesis and the Biological Effects of Tumor Promoters. In Naturally Occurring Phorbol Esters; CRC Press: Boca Raton, FL, USA, 2018; pp. 33–61. [Google Scholar]
- Mulshine, J.L.; Treston, A.M.; Brown, P.H.; Birrer, M.J.; Shaw, G.L. Initiators and promoters of lung cancer. Chest 1993, 103, 4S–11S. [Google Scholar] [CrossRef] [PubMed]
- Weston, A.; Harris, C.C. Multistage Carcinogenesis. In Cancer Medicine; Kufe, D.W., Pollock, R.E., Weichselbaum, R.R., Bast, R.C., Jr., Gansler, T.S., Holland, J.F., Frei, E., III, Eds.; BC Decker: Hamilton, ON, Canada, 2003. [Google Scholar]
- Klein, C.A. Cancer progression and the invisible phase of metastatic colonization. Nat. Rev. Cancer 2020, 20, 681–694. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Divisi, D.; Di Tommaso, S.; Salvemini, S.; Garramone, M.; Crisci, R. Diet and cancer. Acta Bio-Med. Atenei Parm. 2006, 77, 118–123. [Google Scholar]
- Wallace, T.C.; Bailey, R.L.; Blumberg, J.B.; Burton-Freeman, B.; Chen, C.-Y.O.; Crowe-White, K.M.; Drewnowski, A.; Hooshmand, S.; Johnson, E.; Lewis, R.; et al. Fruits, vegetables, and health: A comprehensive narrative, umbrella review of the science and recommendations for enhanced public policy to improve intake. Crit. Rev. Food Sci. Nutr. 2019, 60, 2174–2211. [Google Scholar] [CrossRef] [PubMed]
- Schulze, M.B.; A Martínez-González, M.; Fung, T.T.; Lichtenstein, A.H.; Forouhi, N. Food based dietary patterns and chronic disease prevention. BMJ 2018, 361, k2396. [Google Scholar] [CrossRef] [PubMed]
- Afshin, A.; Sur, P.J.; Fay, K.A.; Cornaby, L.; Ferrara, G.; Salama, J.S.; Mullany, E.C.; Abate, K.H.; Abbafati, C.; Abebe, Z.; et al. Health effects of dietary risks in 195 countries, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2019, 393, 1958–1972. [Google Scholar] [CrossRef]
- Lippi, G.; Mattiuzzi, C.; Cervellin, G. Meat consumption and cancer risk: A critical review of published meta-analyses. Crit. Rev. Oncol. 2015, 97, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Johnson, I. The cancer risk related to meat and meat products. Br. Med Bull. 2016, 121, 73–81. [Google Scholar] [CrossRef]
- Yu, H.; Xu, Q.; Xiong, W.; Liu, Z.; Cai, L.; He, F. Association of pickled food, fired food and smoked food combined with smoking and alcohol drinking with lung cancer: A case-control study. Wei Sheng Yan Jiu J. Hyg. Res. 2019, 48, 925–931. [Google Scholar]
- WHO; FAO. Diet, Nutrition, and the Prevention of Chronic Diseases: Report of a Joint Who/Fao Expert Consultation; World Health Organization: Geneva, Switzerland, 2003. [Google Scholar]
- WHO. Healthy Diet—Fact Sheet No. 394 2018; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Hurtado-Barroso, S.; Trius-Soler, M.; Lamuela-Raventós, R.M.; Zamora-Ros, R. Vegetable and Fruit Consumption and Prognosis among Cancer Survivors: A Systematic Review and Meta-Analysis of Cohort Studies. Adv. Nutr. 2020, 11, 1569–1582. [Google Scholar] [CrossRef] [PubMed]
- Aune, D.; Giovannucci, E.; Boffetta, P.; Fadnes, L.T.; Keum, N.; Norat, T.; Greenwood, D.C.; Riboli, E.; Vatten, L.J.; Tonstad, S. Fruit and vegetable intake and the risk of cardiovascular disease, total cancer and all-cause mortality—A systematic review and dose-response meta-analysis of prospective studies. Int. J. Epidemiol. 2017, 46, 1029–1056. [Google Scholar] [CrossRef] [PubMed]
- Basli, A.; Belkacem, N.; Amrani, I. Health Benefits of Phenolic Compounds against Cancers. In Phenolic Compounds–Biological Activity; IntechOpen: London, UK, 2017; pp. 193–210. [Google Scholar]
- Davidson, K.T.; Zhu, Z.; Fang, Y. Phytochemicals in the Fight against Cancer. Pathol. Oncol. Res. 2016, 22, 655–660. [Google Scholar] [CrossRef] [PubMed]
- Johnson, I.T. Phytochemicals and cancer. Proc. Nutr. Soc. 2007, 66, 207–215. [Google Scholar] [CrossRef]
- Shree, T.J.; Poompavai, S.; Begum SM, F.M.; Gowrisree, V.; Hemalatha, S. Cancer-fighting phytochemicals: Another look. J. Nanomedine Biother. Discov. 2019, 8, 162. [Google Scholar]
- Scarpa, E.-S.; Ninfali, P. Phytochemicals as Innovative Therapeutic Tools against Cancer Stem Cells. Int. J. Mol. Sci. 2015, 16, 15727–15742. [Google Scholar] [CrossRef] [PubMed]
- Talalay, P.; Fahey, J.W. Phytochemicals from Cruciferous Plants Protect against Cancer by Modulating Carcinogen Metabolism. J. Nutr. 2001, 131, 3027S–3033S. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.-F.; Weng, C.-J.; Sethi, G.; Hu, D.-N. Natural Bioactives and Phytochemicals Serve in Cancer Treatment and Prevention. Evid.-Based Complement. Altern. Med. 2013, 2013, 698190. [Google Scholar] [CrossRef]
- Meybodi, N.M.; Mortazavian, A.M.; Monfared, A.B.; Sohrabvandi, S.; Meybodi, F.A. Phytochemicals in Cancer Prevention: A Review of the Evidence. Iran. J. Cancer Prev. 2017, 10, e7219. [Google Scholar] [CrossRef]
- Ranjan, A.; Ramachandran, S.; Gupta, N.; Kaushik, I.; Wright, S.; Srivastava, S.; Das, H.; Srivastava, S.; Prasad, S.; Srivastava, S.K. Role of phytochemicals in cancer prevention. Int. J. Mol. Sci. 2019, 20, 4981. [Google Scholar] [CrossRef] [PubMed]
- Zubair, H.; Azim, S.; Ahmad, A.; Khan, M.A.; Patel, G.K.; Singh, S.; Singh, A.P. Cancer Chemoprevention by Phytochemicals: Nature’s Healing Touch. Molecules 2017, 22, 395. [Google Scholar] [CrossRef] [PubMed]
- Boyer, J.; Liu, R.H. Apple phytochemicals and their health benefits. Nutr. J. 2004, 3, 5. [Google Scholar] [CrossRef]
- Gallus, S.; Talamini, R.; Giacosa, A.; Montella, M.; Ramazzotti, V.; Franceschi, S.; Negri, E.; La Vecchia, C. Does an apple a day keep the oncologist away? Ann. Oncol. 2005, 16, 1841–1844. [Google Scholar] [CrossRef] [PubMed]
- Rupasinghe, H.V.; Thilakarathna, S.; Nair, S. Polyphenols of Apples and Their Potential Health Benefits. In Polyphenols: Chemistry, Dietary Sources and Health Benefits; Nova Science Publishers: Hauppauge, NY, USA, 2013; pp. 333–368. [Google Scholar]
- Rana, S.; Bhushan, S. Apple phenolics as nutraceuticals: Assessment, analysis and application. J. Food Sci. Technol. 2016, 53, 1727–1738. [Google Scholar] [CrossRef] [PubMed]
- Konopacka, D.; Jesionkowska, K.; Kruczyńska, D.; Stehr, R.; Schoorl, F.; Buehler, A.; Egger, S.; Codarin, S.; Hilaire, C.; Höller, I.; et al. Apple and peach consumption habits across European countries. Appetite 2010, 55, 478–483. [Google Scholar] [CrossRef]
- Kidoń, M.; Grabowska, J. Bioactive compounds, antioxidant activity, and sensory qualities of red-fleshed apples dried by different methods. LWT 2020, 136, 110302. [Google Scholar] [CrossRef]
- Dashbaldan, S.; Pączkowski, C.; Szakiel, A. Variations in triterpenoid deposition in cuticular Waxes during development and maturation of selected fruits of Rosaceae family. Int. J. Mol. Sci. 2020, 21, 9762. [Google Scholar] [CrossRef] [PubMed]
- Sut, S.; Poloniato, G.; Malagoli, M.; Dall’Acqua, S. Fragmentation of the main triterpene acids of apple by LC-APCI-MSn. J. Mass Spectrom. 2018, 53, 882–892. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.A.; Bynum, J.P.; Sirovich, B.E. Association between apple consumption and physician visits: Appealing the conventional wisdom that an apple a day keeps the doctor away. JAMA Intern. Med. 2015, 175, 777–783. [Google Scholar] [CrossRef]
- Vinson, J.A.; Su, X.; Zubik, L.; Bose, P. Phenol Antioxidant Quantity and Quality in Foods: Fruits. J. Agric. Food Chem. 2001, 49, 5315–5321. [Google Scholar] [CrossRef]
- Scalbert, A.; Williamson, G. Dietary Intake and Bioavailability of Polyphenols. J. Nutr. 2000, 130, 2073S–2085S. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Chu, Y.-F.; Wu, X.; Liu, R.H. Antioxidant and Antiproliferative Activities of Common Fruits. J. Agric. Food Chem. 2002, 50, 7449–7454. [Google Scholar] [CrossRef]
- McCann, M.; Gill, C.; Brien, G.O.; Rao, J.; McRoberts, W.; Hughes, P.; McEntee, R.; Rowland, I. Anti-cancer properties of phenolics from apple waste on colon carcinogenesis in vitro. Food Chem. Toxicol. 2007, 45, 1224–1230. [Google Scholar] [CrossRef]
- Kalinowska, M.; Bielawska, A.; Lewandowska-Siwkiewicz, H.; Priebe, W.; Lewandowski, W. Apples: Content of phenolic compounds vs. variety, part of apple and cultivation model, extraction of phenolic compounds, biological properties. Plant Physiol. Biochem. 2014, 84, 169–188. [Google Scholar] [CrossRef] [PubMed]
- Imeh, U.; Khokhar, S. Distribution of conjugated and free phenols in fruits: Antioxidant activity and cultivar variations. J. Agric. Food Chem. 2002, 50, 6301–6306. [Google Scholar] [CrossRef] [PubMed]
- Almeida, D.P.; Gião, M.S.; Pintado, M.; Gomes, M.H. Bioactive phytochemicals in apple cultivars from the Portuguese protected geographical indication “Maçã de Alcobaça:” Basis for market segmentation. Int. J. Food Prop. 2017, 20, 2206–2214. [Google Scholar] [CrossRef]
- McGhie, T.K.; Hunt, M.; Barnett, L.E. Cultivar and growing region determine the antioxidant polyphenolic concentration and composition of apples grown in New Zealand. J. Agric. Food Chem. 2005, 53, 3065–3070. [Google Scholar] [CrossRef]
- Honda, C.; Moriya, S. Anthocyanin Biosynthesis in Apple Fruit. Hortic. J. 2018, 87, 305–314. [Google Scholar] [CrossRef]
- De Paepe, D.; Valkenborg, D.; Noten, B.; Servaes, K.; Diels, L.; De Loose, M.; Van Droogenbroeck, B.; Voorspoels, S. Variability of the phenolic profiles in the fruits from old, recent and new apple cultivars cultivated in Belgium. Metabolomics 2015, 11, 739–752. [Google Scholar] [CrossRef]
- Kruger, M.J.; Davies, N.; Myburgh, K.H.; Lecour, S. Proanthocyanidins, anthocyanins and cardiovascular diseases. Food Res. Int. 2014, 59, 41–52. [Google Scholar] [CrossRef]
- Wang, X.; Li, C.; Liang, D.; Zou, Y.; Li, P.; Ma, F. Phenolic compounds and antioxidant activity in red-fleshed apples. J. Funct. Foods 2015, 18, 1086–1094. [Google Scholar] [CrossRef]
- Rana, S.; Rana, A.; Gupta, S.; Bhushan, S. Varietal influence on phenolic constituents and nutritive characteristics of pomace obtained from apples grown in western Himalayas. J. Food Sci. Technol. 2020, 58, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Laaksonen, O.; Kuldjärv, R.; Paalme, T.; Virkki, M.; Yang, B. Impact of apple cultivar, ripening stage, fermentation type and yeast strain on phenolic composition of apple ciders. Food Chem. 2017, 233, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Alberti, A.; Zielinski, A.; Couto, M.; Judacewski, P.; Igarashi-Mafra, L.; Nogueira, A. Distribution of phenolic compounds and antioxidant capacity in apples tissues during ripening. J. Food Sci. Technol. 2017, 54, 1511–1518. [Google Scholar] [CrossRef]
- Van der Sluis, A.A.; Dekker, M.; de Jager, A.; Jongen, W.M. Activity and concentration of polyphenolic antioxidants in apple: Effect of cultivar, harvest year, and storage conditions. J. Agric. Food Chem. 2001, 49, 3606–3613. [Google Scholar] [CrossRef]
- Łata, B.; Trampczynska, A.; Paczesna, J. Cultivar variation in apple peel and whole fruit phenolic composition. Sci. Hortic. 2009, 121, 176–181. [Google Scholar] [CrossRef]
- Kalinowska, M.; Gryko, K.; Wróblewska, A.M.; Jabłońska-Trypuć, A.; Karpowicz, D. Phenolic content, chemical composition and anti-/pro-oxidant activity of Gold Milenium and Papierowka apple peel extracts. Sci. Rep. 2020, 10, 14951. [Google Scholar] [CrossRef]
- Kschonsek, J.; Wolfram, T.; Stöckl, A.; Böhm, V. Polyphenolic Compounds Analysis of Old and New Apple Cultivars and Contribution of Polyphenolic Profile to the In Vitro Antioxidant Capacity. Antioxidants 2018, 7, 20. [Google Scholar] [CrossRef] [PubMed]
- Shehzadi, K.; Rubab, Q.; Asad, L.; Ishfaq, M.; Shafique, B.; Modassar, M.; Ranjha, A.N.; Mahmood, S.; Mueen-Ud-Din, G.; Javaid, T. A critical review on presence of polyphenols in commercial varieties of apple peel, their extraction and Health benefits. Open Access J. Biog. Sci. Res. 2020, 6, 18. [Google Scholar]
- Veberic, R.; Trobec, M.; Herbinger, K.; Hofer, M.; Grill, D.; Stampar, F. Phenolic compounds in some apple (Malus domestica Borkh) cultivars of organic and integrated production. J. Sci. Food Agric. 2005, 85, 1687–1694. [Google Scholar] [CrossRef]
- Francini, A.; Sebastiani, L. Phenolic compounds in apple (Malus × domestica Borkh): Compounds characterization and stability during postharvest and after processing. Antioxidants 2013, 2, 181–193. [Google Scholar] [CrossRef]
- Raudone, L.; Raudonis, R.; Liaudanskas, M.; Janulis, V.; Viskelis, P. Phenolic antioxidant profiles in the whole fruit, flesh and peel of apple cultivars grown in Lithuania. Sci. Hortic. 2017, 216, 186–192. [Google Scholar] [CrossRef]
- Tsao, R.; Yang, R.; Young, A.J.C.; Zhu, H. Polyphenolic Profiles in Eight Apple Cultivars Using High-Performance Liquid Chromatography (HPLC). J. Agric. Food Chem. 2003, 51, 6347–6353. [Google Scholar] [CrossRef] [PubMed]
- Łata, B. Relationship between Apple Peel and the Whole Fruit Antioxidant Content: Year and Cultivar Variation. J. Agric. Food Chem. 2007, 55, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Lewinska, A.; Adamczyk-Grochala, J.; Kwasniewicz, E.; Deregowska, A.; Wnuk, M. Ursolic acid-mediated changes in glycolytic pathway promote cytotoxic autophagy and apoptosis in phenotypically different breast cancer cells. Apoptosis 2017, 22, 800–815. [Google Scholar] [CrossRef] [PubMed]
- Le Bourvellec, C.; Bouzerzour, K.; Ginies, C.; Regis, S.; Plé, Y.; Renard, C.M. Phenolic and polysaccharidic composition of applesauce is close to that of apple flesh. J. Food Compos. Anal. 2011, 24, 537–547. [Google Scholar] [CrossRef]
- Selby-Pham, S.N.; Miller, R.B.; Howell, K.; Dunshea, F.; Bennett, L.E. Physicochemical properties of dietary phytochemicals can predict their passive absorption in the human small intestine. Sci. Rep. 2017, 7, 1931. [Google Scholar] [CrossRef]
- Marín, L.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Bioavailability of dietary polyphenols and gut microbiota metabolism: Antimicrobial properties. BioMed Res. Int. 2015, 2015, 905215. [Google Scholar] [CrossRef]
- Bondonno, N.P.; Bondonno, C.P.; Ward, N.C.; Hodgson, J.M.; Croft, K.D. The cardiovascular health benefits of apples: Whole fruit vs. isolated compounds. Trends Food Sci. Technol. 2017, 69, 243–256. [Google Scholar] [CrossRef]
- Cosme, P.; Rodríguez, A.B.; Espino, J.; Garrido, M. Plant phenolics: Bioavailability as a key determinant of their potential health-promoting applications. Antioxidants 2020, 9, 1263. [Google Scholar] [CrossRef] [PubMed]
- Anantharaju, P.G.; Gowda, P.C.; Vimalambike, M.G.; Madhunapantula, S.V. An overview on the role of dietary phenolics for the treatment of cancers. Nutr. J. 2016, 15, 1–16. [Google Scholar] [CrossRef]
- Aprikian, O.; Duclos, V.; Guyot, S.; Besson, C.; Manach, C.; Bernalier, A.; Morand, C.; Rémésy, C.; Demigné, C. Apple pectin and a polyphenol-rich apple concentrate are more effective together than separately on cecal fermentations and plasma lipids in rats. J. Nutr. 2003, 133, 1860–1865. [Google Scholar] [CrossRef] [PubMed]
- Selma, M.V.; Espín, J.C.; Tomas-Barberan, F. Interaction between Phenolics and Gut Microbiota: Role in Human Health. J. Agric. Food Chem. 2009, 57, 6485–6501. [Google Scholar] [CrossRef] [PubMed]
- Stracke, B.A.; Rüfer, C.E.; Bub, A.; Seifert, S.; Weibel, F.P.; Kunz, C.; Watzl, B. No effect of the farming system (organic/conventional) on the bioavailability of apple (Malus domestica Bork., cultivar Golden Delicious) polyphenols in healthy men: A comparative study. Eur. J. Nutr. 2010, 49, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Wruss, J.; Lanzerstorfer, P.; Huemer, S.; Himmelsbach, M.; Mangge, H.; Höglinger, O.; Weghuber, D.; Weghuber, J. Differences in pharmacokinetics of apple polyphenols after standardized oral consumption of unprocessed apple juice. Nutr. J. 2015, 14, 1–11. [Google Scholar] [CrossRef] [PubMed]
- DuPont, M.S.; Bennett, R.N.; Mellon, F.A.; Williamson, G. Polyphenols from alcoholic apple cider are absorbed, metabolized and excreted by humans. J. Nutr. 2002, 132, 172–175. [Google Scholar] [CrossRef]
- Koutsos, A.; Lima, M.; Conterno, L.; Gasperotti, M.; Bianchi, M.; Fava, F.; Vrhovsek, U.; Lovegrove, J.A.; Tuohy, K.M. Effects of Commercial Apple Varieties on Human Gut Microbiota Composition and Metabolic Output Using an In Vitro Colonic Model. Nutrients 2017, 9, 533. [Google Scholar] [CrossRef]
- Shinohara, K.; Ohashi, Y.; Kawasumi, K.; Terada, A.; Fujisawa, T. Effect of apple intake on fecal microbiota and metabolites in humans. Anaerobe 2010, 16, 510–515. [Google Scholar] [CrossRef]
- Williams, B.A.; Grant, L.J.; Gidley, M.J.; Mikkelsen, D. Gut fermentation of dietary fibres: Physico-chemistry of plant cell walls and implications for health. Int. J. Mol. Sci. 2017, 18, 2203. [Google Scholar] [CrossRef]
- Efimtseva, E.A.; Chelpanova, T.I. Apples as a Source of Soluble and Insoluble Dietary Fibers: Effect of Dietary Fibers on Appetite. Hum. Physiol. 2020, 46, 224–234. [Google Scholar] [CrossRef]
- Andoh, A.; Tsujikawa, T.; Fujiyama, Y. Role of dietary fiber and short-chain fatty acids in the colon. Curr. Pharm. Des. 2003, 9, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Starowicz, M.; Achrem-Achremowicz, B.; Piskula, M.; Zielinski, H. Phenolic compounds from apples: Reviewing their occurrence, absorption, bioavailability, processing, and antioxidant activity—A review. Pol. J. Food Nutr. Sci. 2020, 70, 321–336. [Google Scholar] [CrossRef]
- Zou, T.; Wang, B.; Li, S.; Liu, Y.; You, J. Dietary apple polyphenols promote fat browning in high-fat diet-induced obese mice through activation of adenosine monophosphate-activated protein kinase α. J. Sci. Food Agric. 2020, 100, 2389–2398. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.-N.; Meng, X.; Li, Y.; Li, S.; Liu, Q.; Tang, G.-Y.; Li, H.-B. Fruits for Prevention and Treatment of Cardiovascular Diseases. Nutrients 2017, 9, 598. [Google Scholar] [CrossRef] [PubMed]
- Gayer, B.A.; Avendano, E.E.; Edelson, E.; Nirmala, N.; Johnson, E.J.; Raman, G. Effects of Intake of Apples, Pears, or Their Products on Cardiometabolic Risk Factors and Clinical Outcomes: A Systematic Review and Meta-Analysis. Curr. Dev. Nutr. 2019, 3, nzz109. [Google Scholar] [CrossRef]
- Hyun, T.K.; Jang, K.I. Apple as a source of dietary phytonutrients: An update on the potential health benefits of apple. EXCLI J. 2016, 15, 565. [Google Scholar]
- Mahmud, L.S.M.; Tisha, A.; Sagor, A.T. A comprehensive review on effective role of Apple polyphenols in the treatment of obesity, diabetes, and liver dysfunctions with some possible molecular mechanisms. Oxid. Antioxid. Med Sci. 2018, 7, 9–27. [Google Scholar]
- Guo, X.F.; Yang, B.; Tang, J.; Jiang, J.J.; Li, D. Apple and pear consumption and type 2 diabetes mellitus risk: A meta-analysis of prospective cohort studies. Food Funct. 2017, 8, 927–934. [Google Scholar] [CrossRef]
- Alam, M.N.; Almoyad, M.; Huq, F. Polyphenols in colorectal cancer: Current state of knowledge including clinical trials and molecular mechanism of action. BioMed Res. Int. 2018, 2018, 4154185. [Google Scholar] [CrossRef]
- Martínez-Rodríguez, O.P.; Thompson-Bonilla, M.D.R.; Jaramillo-Flores, M.E. Association between obesity and breast cancer: Molecular bases and the effect of flavonoids in signaling pathways. Crit. Rev. Food Sci. Nutr. 2020, 60, 3770–3792. [Google Scholar] [CrossRef]
- Linseisen, J.; Rohrmann, S.; Miller, A.B.; Bueno-De-Mesquita, H.B.; Büchner, F.; Vineis, P.; Agudo, A.; Gram, I.T.; Janson, L.; Krogh, V.; et al. Fruit and vegetable consumption and lung cancer risk: Updated information from the European Prospective Investigation into Cancer and Nutrition (EPIC). Int. J. Cancer 2007, 121, 1103–1114. [Google Scholar] [CrossRef]
- Büchner, F.L.; Bueno-de-Mesquita, H.B.; Ros, M.M.; Kampman, E.; Egevad, L.; Overvad, K.; Raaschou-Nielsen, O.; Tjønneland, A.; Roswall, N.; Clavel-Chapelon, F.; et al. Consumption of vegetables and fruit and the risk of bladder cancer in the European Prospective Investigation into Cancer and Nutrition. Int. J. Cancer 2009, 125, 2643–2651. [Google Scholar] [CrossRef] [PubMed]
- Feskanich, D.; Ziegler, R.G.; Michaud, D.S.; Giovannucci, E.L.; Speizer, F.E.; Willett, W.C.; Colditz, G. Prospective Study of Fruit and Vegetable Consumption and Risk of Lung Cancer Among Men and Women. J. Natl. Cancer Inst. 2000, 92, 1812–1823. [Google Scholar] [CrossRef] [PubMed]
- Knekt, P.; Järvinen, R.; Seppänen, R.; Heliövaara, M.; Teppo, L.; Pukkala, E.; Aromaa, A. Dietary flavonoids and the risk of lung cancer and other malignant neoplasms. Am. J. Epidemiol. 1997, 146, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Le Marchand, L.; Murphy, S.P.; Hankin, J.H.; Wilkens, L.R.; Kolonel, L.N. Intake of flavonoids and lung cancer. J. Natl. Cancer Inst. 2000, 92, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Arts, I.C.; Hollman, P.C.; Bueno de Mesquita, H.B.; Feskens, E.J.; Kromhout, D. Dietary catechins and epithelial cancer incidence: The Zutphen elderly study. Int. J. Cancer 2001, 92, 298–302. [Google Scholar] [CrossRef]
- Farvid, M.S.; Chen, W.Y.; Rosner, B.A.; Tamimi, R.M.; Willett, W.C.; Eliassen, A.H. Fruit and vegetable consumption and breast cancer incidence: Repeated measures over 30 years of follow-up. Int. J. Cancer 2018, 144, 1496–1510. [Google Scholar] [CrossRef]
- Rossi, M.; Lugo, A.; Lagiou, P.; Zucchetto, A.; Polesel, J.; Serraino, D.; Negri, E.; Trichopoulos, D.; La Vecchia, C. Proanthocyanidins and other flavonoids in relation to pancreatic cancer: A case–control study in Italy. Ann. Oncol. 2012, 23, 1488–1493. [Google Scholar] [CrossRef]
- Smith-Warner, S.A.; Spiegelman, D.; Yaun, S.S.; Adami, H.O.; Beeson, W.L.; Van Den Brandt, P.A.; Folsom, A.R.; Fraser, G.E.; Freudenheim, J.L.; Goldbohm, R.A.; et al. Intake of fruits and vegetables and risk of breast cancer: A pooled analysis of cohort studies. JAMA 2001, 285, 769–776. [Google Scholar] [CrossRef] [PubMed]
- Torres-Sánchez, L.; López-Carrillo, L.; Ló-Cervantes, M.; Rueda-Neria, C.; Wolff, M.S. Food Sources of Phytoestrogens and Breast Cancer Risk in Mexican Women. Nutr. Cancer 2000, 37, 134–139. [Google Scholar] [CrossRef]
- Jedrychowski, W.; Maugeri, U.; Popiela, T.; Kulig, J.; Sochacka-Tatara, E.; Pac, A.; Sowa, A.; Musial, A. Case–control study on beneficial effect of regular consumption of apples on colorectal cancer risk in a population with relatively low intake of fruits and vegetables. Eur. J. Cancer Prev. 2010, 19, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Kreimer, A.R.; Randi, G.; Herrero, R.; Castellsagué, X.; Vecchia, C.L.; Franceschi, S. Diet and body mass, and oral and oropharyngeal squamous cell carcinomas: Analysis from the IARC multinational case–control study. Int. J. Cancer 2006, 118, 2293–2297. [Google Scholar] [CrossRef] [PubMed]
- Rashidkhani, B.; Lindblad, P.; Wolk, A. Fruits, vegetables and risk of renal cell carcinoma: A prospective study of Swedish women. Int. J. Cancer 2004, 113, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Lindblad, P.; Wolk, A.; Bergström, R.; Adami, H.O. Diet and risk of renal cell cancer: A population-based case-control study. Cancer Epidemiol. Biomark. Prev. 1997, 6, 215–223. [Google Scholar]
- Askari, F.; Parizi, M.K.; Jessri, M.; Rashidkhani, B. Fruit and Vegetable Intake in Relation to Prostate Cancer in Iranian Men: A Case-Control Study. Asian Pac. J. Cancer Prev. 2014, 15, 5223–5227. [Google Scholar] [CrossRef] [PubMed]
- Malin, A.S.; Qi, D.; Shu, X.O.; Gao, Y.T.; Friedmann, J.M.; Jin, F.; Zheng, W. Intake of fruits, vegetables and selected micronutrients in relation to the risk of breast cancer. Int. J. Cancer 2003, 105, 413–418. [Google Scholar] [CrossRef]
- Fabiani, R.; Minelli, L.; Rosignoli, P. Apple intake and cancer risk: A systematic review and meta-analysis of observational studies. Public Health Nutr. 2016, 19, 2603–2617. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Farias, M.; Carrasco-Pozo, C. The Anti-Cancer Effect of Quercetin: Molecular Implications in Cancer Metabolism. Int. J. Mol. Sci. 2019, 20, 3177. [Google Scholar] [CrossRef]
- Li, C.X.; Zhao, X.H.; Zuo, W.F.; Zhang, T.L.; Zhang, Z.Y.; Chen, X.S. Phytochemical profiles, antioxidant, and antiproliferative activities of four red-fleshed apple varieties in China. J. Food Sci. 2020, 85, 718–726. [Google Scholar] [CrossRef]
- Sun, J.; Liu, R.H. Apple Phytochemical Extracts Inhibit Proliferation of Estrogen-Dependent and Estrogen-Independent Human Breast Cancer Cells through Cell Cycle Modulation. J. Agric. Food Chem. 2008, 56, 11661–11667. [Google Scholar] [CrossRef] [PubMed]
- Schiavano, G.F.; De Santi, M.; Brandi, G.; Fanelli, M.; Bucchini, A.; Giamperi, L.; Giomaro, G.M. Inhibition of Breast Cancer Cell Proliferation and In Vitro Tumorigenesis by a New Red Apple Cultivar. PLoS ONE 2015, 10, e0135840. [Google Scholar] [CrossRef]
- Reagan-Shaw, S.; Eggert, D.; Mukhtar, H.; Ahmad, N. Antiproliferative Effects of Apple Peel Extract Against Cancer Cells. Nutr. Cancer 2010, 62, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Quagliuolo, L.; D’Angelo, S. Annurca Apple Biophenols’ Effects in Combination with Cisplatin on A549 Cells. Curr. Nutr. Food Sci. 2020, 17, 111–120. [Google Scholar] [CrossRef]
- Rana, S.; Kumar, S.; Rana, A.; Padwad, Y.; Bhushan, S. Biological activity of phenolics enriched extracts from industrial apple pomace. Ind. Crop. Prod. 2020, 160, 113158. [Google Scholar] [CrossRef]
- Miura, D.; Miura, Y.; Yagasaki, K. Effect of apple polyphenol extract on hepatoma proliferation and invasion in culture and on tumor growth, metastasis, and abnormal lipoprotein profiles in hepatoma-bearing rats. Biosci. Biotechnol. Biochem. 2007, 71, 2743–2750. [Google Scholar] [CrossRef]
- Almatroodi, S.; Alsahli, M.; Almatroudi, A.; Verma, A.; Aloliqi, A.; Allemailem, K.; Khan, A.; Rahmani, A. Potential Therapeutic Targets of Quercetin, a Plant Flavonol, and Its Role in the Therapy of Various Types of Cancer through the Modulation of Various Cell Signaling Pathways. Molecules 2021, 26, 1315. [Google Scholar] [CrossRef] [PubMed]
- Tu, S.-H.; Chen, L.-C.; Ho, Y.-S. An apple a day to prevent cancer formation: Reducing cancer risk with flavonoids. J. Food Drug Anal. 2016, 25, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Koh, Y.-C.; Ho, C.-T.; Pan, M.-H. Recent advances in cancer chemoprevention with phytochemicals. J. Food Drug Anal. 2019, 28, 14–37. [Google Scholar] [CrossRef]
- Bars-Cortina, D.; Martínez-Bardají, A.; Macià, A.; Motilva, M.J.; Piñol-Felis, C. Consumption evaluation of one apple flesh a day in the initial phases prior to adenoma/adenocarcinoma in an azoxymethane rat colon carcinogenesis model. J. Nutr. Biochem. 2020, 83, 108418. [Google Scholar] [CrossRef]
- Liu, R.H.; Liu, J.; Chen, B. Apples prevent mammary tumors in rats. J. Agric. Food Chem. 2005, 53, 2341–2343. [Google Scholar] [CrossRef] [PubMed]
- Veeriah, S.; Kautenburger, T.; Habermann, N.; Sauer, J.; Dietrich, H.; Will, F.; Pool-Zobel, B.L. Apple flavonoids inhibit growth of HT29 human colon cancer cells and modulate expression of genes involved in the biotransformation of xenobiotics. Mol. Carcinog. 2006, 45, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Leng, E.; Xiao, Y.; Mo, Z.; Li, Y.; Zhang, Y.; Deng, X.; Li, W. Synergistic effect of phytochemicals on cholesterol metabolism and lipid accumulation in HepG2 cells. BMC Complementary Altern. Med. 2018, 18, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Virgous, C.; Si, H. Synergistic anti-inflammatory effects and mechanisms of combined phytochemicals. J. Nutr. Biochem. 2019, 69, 19–30. [Google Scholar] [CrossRef]
- Yang, J.; Liu, R.H. Synergistic effect of apple extracts and quercetin 3-β-d-glucoside combination on antiproliferative activity in MCF-7 human breast cancer cells in vitro. J. Agric. Food Chem. 2009, 57, 8581–8586. [Google Scholar] [CrossRef]
- Moskovitz, J.; Yim, M.B.; Chock, P.B. Free radicals and disease. Arch. Biochem. Biophys. 2002, 397, 354–359. [Google Scholar] [CrossRef]
- Adom, K.K.; Liu, R.H. Antioxidant activity of grains. J. Agric. Food Chem. 2002, 50, 6182–6187. [Google Scholar] [CrossRef]
- Lee, K.W.; Kim, Y.J.; Kim, D.O.; Lee, H.J.; Lee, C.Y. Major phenolics in apple and their contribution to the total antioxidant capacity. J. Agric. Food Chem. 2003, 51, 6516–6520. [Google Scholar] [CrossRef]
- Wandjou, J.G.N.; Lancioni, L.; Barbalace, M.C.; Hrelia, S.; Papa, F.; Sagratini, G.; Vittori, S.; Dall’Acqua, S.; Caprioli, G.; Beghelli, D.; et al. Comprehensive characterization of phytochemicals and biological activities of the Italian ancient apple ‘Mela Rosa dei Monti Sibillini’. Food Res. Int. 2020, 137, 109422. [Google Scholar] [CrossRef]
- Sies, H. Total Antioxidant Capacity: Appraisal of a Concept. J. Nutr. 2007, 137, 1493–1495. [Google Scholar] [CrossRef]
- Wolfe, K.; Wu, X.; Liu, R.H. Antioxidant Activity of Apple Peels. J. Agric. Food Chem. 2003, 51, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.H.; Sun, J. Antiproliferative Activity of Apples Is Not Due to Phenolic-Induced Hydrogen Peroxide Formation. J. Agric. Food Chem. 2003, 51, 1718–1723. [Google Scholar] [CrossRef] [PubMed]
- Eberhardt, M.V.; Lee, C.Y.; Liu, R.H. Antioxidant activity of fresh apples. Nat. Cell Biol. 2000, 405, 903–904. [Google Scholar] [CrossRef] [PubMed]
- Martino, E.; Vuoso, D.C.; D’Angelo, S.; Mele, L.; D’Onofrio, N.; Porcelli, M.; Cacciapuoti, G. Annurca apple polyphenol extract selectively kills MDA-MB-231 cells through ROS generation, sustained JNK activation and cell growth and survival inhibition. Scientific reports 2019, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- León-González, A.J.; Auger, C.; Schini-Kerth, V.B. Pro-oxidant activity of polyphenols and its implication on cancer chemoprevention and chemotherapy. Biochem. Pharmacol. 2015, 98, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Decker, E.A. Phenolics: Prooxidants or antioxidants? Nutr. Rev. 1997, 55, 396–398. [Google Scholar] [CrossRef]
- Rahal, A.; Kumar, A.; Singh, V.; Yadav, B.; Tiwari, R.; Chakraborty, S.; Dhama, K. Oxidative stress, prooxidants, and antioxidants: The interplay. BioMed Res. Int. 2014, 2014, 761264. [Google Scholar] [CrossRef]
- Sotler, R.; Poljšak, B.; Dahmane, R.; Jukić, T.; Jukić, D.P.; Rotim, C.; Trebše, P.; Starc, A. Prooxidant Activities of Antioxidants and Their Impact on Health. Acta Clin. Croat. 2019, 58, 726–736. [Google Scholar] [CrossRef]
- Azmi, A.S.; Sarkar, F.H.; Hadi, S.M. Pro-oxidant activity of dietary chemopreventive agents: An under-appreciated anti-cancer property. F1000Research 2013, 2, 135. [Google Scholar] [CrossRef][Green Version]
- Fernando, W.; Rupasinghe, H.V.; Hoskin, D.W. Dietary phytochemicals with anti-oxidant and pro-oxidant activities: A double-edged sword in relation to adjuvant chemotherapy and radiotherapy? Cancer Lett. 2019, 452, 168–177. [Google Scholar] [CrossRef]
- Eghbaliferiz, S.; Iranshahi, M. Prooxidant activity of polyphenols, flavonoids, anthocyanins and carotenoids: Updated review of mechanisms and catalyzing metals. Phytother. Res. 2016, 30, 1379–1391. [Google Scholar] [CrossRef] [PubMed]
- Moldovan, B.; Hosu, A.; David, L.; Cimpoiu, C. Total phenolics, anthocyanins, antioxidant and pro-oxidant activity of some red fruits teas. Acta Chim. Slov. 2016, 63, 213–219. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yordi, E.G.; Pérez, E.M.; Matos, M.J.; Villares, E.U. Antioxidant and pro-oxidant effects of polyphenolic compounds and structure-activity relationship evidence. Nutr. Well-Being Health 2012, 2, 23–48. [Google Scholar]
- Mendoza-Wilson, A.M.; Castro-Arredondo, S.I.; Espinosa-Plascencia, A.; del Refugio Robles-Burgueño, M.; Balandrán-Quintana, R.R.; del Carmen Bermúdez-Almada, M. Chemical composition and antioxidant-prooxidant potential of a polyphenolic extract and a proanthocyanidin-rich fraction of apple skin. Heliyon 2016, 2, e00073. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.-T.; Ha, J.; Park, I.-J.; Lee, S.-K.; Baik, H.W.; Kim, Y.M.; Park, O.J. Apoptotic effect of EGCG in HT-29 colon cancer cells via AMPK signal pathway. Cancer Lett. 2007, 247, 115–121. [Google Scholar] [CrossRef]
- Lambert, J.D.; Elias, R.J. The antioxidant and pro-oxidant activities of green tea polyphenols: A role in cancer prevention. Arch. Biochem. Biophys. 2010, 501, 65–72. [Google Scholar] [CrossRef]
- Kocyigit, A.; Guler, E.M.; Dikilitas, M. Role of Antioxidant Phytochemicals in Prevention, Formation and Treatment of Cancer. In Reactive Oxygen Species (ROS) in Living Cells; InterchOpen: London, UK, 2018; pp. 21–45. [Google Scholar]
- Almeida, A.F.; Borge, G.I.A.; Piskula, M.; Tudose, A.; Tudoreanu, L.; Valentová, K.; Williamson, G.; Santos, C.N. Bioavailability of Quercetin in Humans with a Focus on Interindividual Variation. Compr. Rev. Food Sci. Food Saf. 2018, 17, 714–731. [Google Scholar] [CrossRef]
- Karakaya, S. Bioavailability of phenolic compounds. Crit. Rev. Food Sci. Nutr. 2004, 44, 453–464. [Google Scholar] [CrossRef]
- Petersen, B.; Egert, S.; Bosy-Westphal, A.; Müller, M.J.; Wolffram, S.; Hubbermann, E.M.; Rimbach, G.; Schwarz, K. Bioavailability of quercetin in humans and the influence of food matrix comparing quercetin capsules and different apple sources. Food Res. Int. 2016, 88, 159–165. [Google Scholar] [CrossRef]
- Hollman, P.C.; Tijburg, L.B.; Yang, C.S. Bioavailability of flavonoids from tea. Crit. Rev. Food Sci. Nutr. 1997, 37, 719–738. [Google Scholar] [CrossRef]
- Hollman, P.C.; Van Trijp, J.; Buysman, M.N.; Gaag, M.S.V.; Mengelers, M.J.; De Vries, J.H.; Katan, M.B. Relative bioavailability of the antioxidant flavonoid quercetin from various foods in man. FEBS Lett. 1997, 418, 152–156. [Google Scholar] [CrossRef]
- Biedrzycka, E.; Amarowicz, R. Diet and Health: Apple Polyphenols as Antioxidants. Food Rev. Int. 2008, 24, 235–251. [Google Scholar] [CrossRef]
- Nishijima, T.; Iwai, K.; Saito, Y.; Takida, Y.; Matsue, H. Chronic Ingestion of Apple Pectin Can Enhance the Absorption of Quercetin. J. Agric. Food Chem. 2009, 57, 2583–2587. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, G.; Turco, I.; Bacchetti, T. Apple as a source of dietary phytonutrients: Bioavailability and evidence of protective effects against human cardiovascular disease. Food Nutr. Sci. 2014, 2014, 13. [Google Scholar] [CrossRef]
- Graefe, E.U.; Wittig, J.; Mueller, S.; Riethling, A.K.; Uehleke, B.; Drewelow, B.; Pforte, H.; Jacobasch, G.; Derendorf, H.; Veit, M. Pharmacokinetics and bioavailability of quercetin glycosides in humans. J. Clin. Pharmacol. 2001, 41, 492–499. [Google Scholar] [CrossRef] [PubMed]
- Erlund, I.; Freese, R.; Marniemi, J.; Hakala, P.; Alfthan, G. Bioavailability of Quercetin from Berries and the Diet. Nutr. Cancer 2006, 54, 13–17. [Google Scholar] [CrossRef]
- Manach, C.; Morand, C.; Texier, O.; Favier, M.L.; Agullo, G.; Demigné, C.; Régérat, F.; Rémésy, C. Quercetin metabolites in plasma of rats fed diets containing rutin or quercetin. J. Nutr. 1995, 125, 1911–1922. [Google Scholar] [CrossRef]
- Batiha, G.E.-S.; Beshbishy, A.M.; Ikram, M.; Mulla, Z.S.; El-Hack, M.E.A.; Taha, A.E.; Algammal, A.M.; Elewa, Y.H.A. The Pharmacological Activity, Biochemical Properties, and Pharmacokinetics of the Major Natural Polyphenolic Flavonoid: Quercetin. Foods 2020, 9, 374. [Google Scholar] [CrossRef]
- Rauf, A.; Imran, M.; Khan, I.A.; Ur-Rehman, M.; Gilani, S.A.; Mehmood, Z.; Mubarak, M.S. Anticancer potential of quercetin: A comprehensive review. Phytother. Res. 2018, 32, 2109–2130. [Google Scholar] [CrossRef]
- Jeong, J.-H.; An, J.Y.; Kwon, Y.T.; Rhee, J.G.; Lee, Y.J. Effects of low dose quercetin: Cancer cell-specific inhibition of cell cycle progression. J. Cell. Biochem. 2008, 106, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Ezzati, M.; Yousefi, B.; Velaei, K.; Safa, A. A review on anti-cancer properties of Quercetin in breast cancer. Life Sci. 2020, 248, 117463. [Google Scholar] [CrossRef]
- Tao, S.-F.; He, H.-F.; Chen, Q. Quercetin inhibits proliferation and invasion acts by up-regulating miR-146a in human breast cancer cells. Mol. Cell. Biochem. 2015, 402, 93–100. [Google Scholar] [CrossRef]
- Deng, X.-H.; Song, H.-Y.; Zhou, Y.-F.; Yuan, G.-Y.; Zheng, F.-J. Effects of quercetin on the proliferation of breast cancer cells and expression of survivin in vitro. Exp. Ther. Med. 2013, 6, 1155–1158. [Google Scholar] [CrossRef] [PubMed]
- Ren, M.X.; Deng, X.H.; Ai, F.; Yuan, G.Y.; Song, H.Y. Effect of quercetin on the proliferation of the human ovarian cancer cell line SKOV-3 in vitro. Exp. Ther. Med. 2015, 10, 579–583. [Google Scholar] [CrossRef] [PubMed]
- Dahiya, R.; Mohammad, T.; Roy, S.; Anwar, S.; Gupta, P.; Haque, A.; Khan, P.; Kazim, S.N.; Islam, A.; Ahmad, F.; et al. Investigation of inhibitory potential of quercetin to the pyruvate dehydrogenase kinase 3: Towards implications in anticancer therapy. Int. J. Biol. Macromol. 2019, 136, 1076–1085. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Ding, H.; Tang, X.; Liang, M.; Li, S.; Zhang, J.; Cao, J. Quercetin induces pro-apoptotic autophagy via SIRT1 / AMPK signaling pathway in human lung cancer cell lines A549 and H1299 in vitro. Thorac. Cancer 2021, 12, 1415–1422. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Lee, Y.H.; Sharma, A.R.; Park, J.B.; Jagga, S.; Sharma, G.; Lee, S.S.; Nam, J.S. Quercetin induces apoptosis and cell cycle arrest in triple-negative breast cancer cells through modulation of Foxo3a activity. Korean J. Physiol. Pharmacol. 2017, 21, 205–213. [Google Scholar] [CrossRef]
- Gong, C.; Yang, Z.; Zhang, L.; Wang, Y.; Gong, W.; Liu, Y. Quercetin suppresses DNA double-strand break repair and enhances the radiosensitivity of human ovarian cancer cells via p53-dependent endoplasmic reticulum stress pathway. OncoTargets Ther. 2018, 11, 17–27. [Google Scholar] [CrossRef]
- Kim, H.-J.; Kim, S.-K.; Kim, B.-S.; Lee, S.-H.; Park, Y.-S.; Park, B.-K.; Kim, S.-J.; Kim, J.; Choi, C.; Kim, J.-S.; et al. Apoptotic Effect of Quercetin on HT-29 Colon Cancer Cells via the AMPK Signaling Pathway. J. Agric. Food Chem. 2010, 58, 8643–8650. [Google Scholar] [CrossRef]
- Glick, D.; Barth, S.; MacLeod, K.F. Autophagy: Cellular and molecular mechanisms. J. Pathol. 2010, 221, 3–12. [Google Scholar] [CrossRef]
- Jia, L.; Huang, S.; Yin, X.; Zan, Y.; Guo, Y.; Han, L. Quercetin suppresses the mobility of breast cancer by suppressing glycolysis through Akt-mTOR pathway mediated autophagy induction. Life Sci. 2018, 208, 123–130. [Google Scholar] [CrossRef]
- Song, Y.; Han, M.; Zhang, X. Quercetin suppresses the migration and invasion in human colon cancer Caco-2 cells through regulating toll-like receptor 4/nuclear factor-kappa B pathway. Pharmacogn. Mag. 2016, 12, S237. [Google Scholar] [CrossRef]
- Oh, S.J.; Kim, O.; Lee, J.S.; Kim, J.-A.; Kim, M.R.; Choi, H.S.; Shim, J.-H.; Kang, K.W.; Kim, Y.C. Inhibition of angiogenesis by quercetin in tamoxifen-resistant breast cancer cells. Food Chem. Toxicol. 2010, 48, 3227–3234. [Google Scholar] [CrossRef] [PubMed]
- Pratheeshkumar, P.; Budhraja, A.; Son, Y.-O.; Wang, X.; Zhang, Z.; Ding, S.; Wang, L.; Hitron, A.; Lee, J.-C.; Xu, M.; et al. Quercetin Inhibits Angiogenesis Mediated Human Prostate Tumor Growth by Targeting VEGFR- 2 Regulated AKT/mTOR/P70S6K Signaling Pathways. PLoS ONE 2012, 7, e47516. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Zhao, X.; Xu, J.; Zhang, H. Quercetin inhibits angiogenesis-mediated human retinoblastoma growth by targeting vascular endothelial growth factor receptor. Oncol. Lett. 2017, 14, 3343–3348. [Google Scholar] [CrossRef]
- Monge, P.; Solheim, E.; Scheline, R.R. Dihydrochalcone metabolism in the rat: Phloretin. Xenobiotica 1984, 14, 917–924. [Google Scholar] [CrossRef] [PubMed]
- Crespy, V.; Aprikian, O.; Morand, C.; Besson, C.; Manach, C.; Demigné, C.; Rémésy, C. Bioavailability of Phloretin and Phloridzin in Rats. J. Nutr. 2001, 131, 3227–3230. [Google Scholar] [CrossRef]
- Marks, S.C.; Mullen, W.; Borges, G.; Crozier, A. Absorption, Metabolism, and Excretion of Cider Dihydrochalcones in Healthy Humans and Subjects with an Ileostomy. J. Agric. Food Chem. 2009, 57, 2009–2015. [Google Scholar] [CrossRef] [PubMed]
- Kahle, K.; Huemmer, W.; Kempf, M.; Scheppach, W.; Erk, T.; Richling, E. Polyphenols Are Intensively Metabolized in the Human Gastrointestinal Tract after Apple Juice Consumption. J. Agric. Food Chem. 2007, 55, 10605–10614. [Google Scholar] [CrossRef]
- Wu, K.-H.; Ho, C.-T.; Chen, Z.-F.; Chen, L.-C.; Whang-Peng, J.; Lin, T.-N.; Ho, Y.-S. The apple polyphenol phloretin inhibits breast cancer cell migration and proliferation via inhibition of signals by type 2 glucose transporter. J. Food Drug Anal. 2017, 26, 221–231. [Google Scholar] [CrossRef]
- Lin, S.-T.; Tu, S.-H.; Yang, P.-S.; Hsu, S.-P.; Lee, W.H.; Ho, C.-T.; Wu, C.-H.; Lai, Y.-H.; Chen, M.-Y.; Chen, L.-C. Apple Polyphenol Phloretin Inhibits Colorectal Cancer Cell Growth via Inhibition of the Type 2 Glucose Transporter and Activation of p53-Mediated Signaling. J. Agric. Food Chem. 2016, 64, 6826–6837. [Google Scholar] [CrossRef]
- Wu, C.H.; Ho, Y.S.; Tsai, C.Y.; Wang, Y.J.; Tseng, H.; Wei, P.L.; Lee, C.H.; Liu, R.S.; Lin, S.Y. In vitro and in vivo study of phloretin—Induced apoptosis in human liver cancer cells involving inhibition of type II glucose transporter. Int. J. Cancer 2009, 124, 2210–2219. [Google Scholar] [CrossRef]
- Xu, M.; Gu, W.; Shen, Z.; Wang, F. Anticancer activity of phloretin against human gastric cancer cell lines involves apoptosis, cell cycle arrest, and inhibition of cell invasion and JNK signalling pathway. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2018, 24, 6551. [Google Scholar] [CrossRef]
- Min, J.; Li, X.; Huang, K.; Tang, H.; Ding, X.; Qi, C.; Qin, X.; Xu, Z. Phloretin induces apoptosis of non-small cell lung carcinoma A549 cells via JNK1/2 and p38 MAPK pathways. Oncol. Rep. 2015, 34, 2871–2879. [Google Scholar] [CrossRef]
- Kim, U.; Kim, C.Y.; Lee, J.M.; Oh, H.; Ryu, B.; Kim, J.; Park, J.H. Phloretin inhibits the human prostate cancer cells through the generation of reactive oxygen species. Pathol. Oncol. Res. 2020, 26, 977–984. [Google Scholar] [CrossRef] [PubMed]
- Saraswati, S.; Alhaider, A.; Abdelgadir, A.M.; Tanwer, P.; Korashy, H.M. Phloretin attenuates STAT-3 activity and overcomes sorafenib resistance targeting SHP-1–mediated inhibition of STAT3 and Akt/VEGFR2 pathway in hepatocellular carcinoma. Cell Commun. Signal. 2019, 17, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Fan, C.; Pu, L.; Wei, C.; Jin, H.; Teng, Y.; Zhao, M.; Yu, A.C.H.; Jiang, F.; Shu, J.; et al. Phloretin induces cell cycle arrest and apoptosis of human glioblastoma cells through the generation of reactive oxygen species. J. Neuro-Oncol. 2016, 128, 217–223. [Google Scholar] [CrossRef]
- Duan, H.; Wang, R.; Yan, X.; Liu, H.; Zhang, Y.; Mu, D.; Han, J.; Li, X. Phloretin induces apoptosis of human esophageal cancer via a mitochondria-dependent pathway. Oncol. Lett. 2017, 14, 6763–6768. [Google Scholar] [CrossRef]
- Chen, M.; Gowd, V.; Wang, M.; Chen, F.; Cheng, K.-W. The apple dihydrochalcone phloretin suppresses growth and improves chemosensitivity of breast cancer cells via inhibition of cytoprotective autophagy. Food Funct. 2020, 12, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Wang, R.; Nan, Y.; Li, W.; Wang, Q.; Jin, F. Phloretin exhibits an anticancer effect and enhances the anticancer ability of cisplatin on non-small cell lung cancer cell lines by regulating expression of apoptotic pathways and matrix metalloproteinases. Int. J. Oncol. 2015, 48, 843–853. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Kim, E.J.; Shin, H.-K.; Kwon, D.Y.; Kim, M.S.; Surh, Y.-J.; Park, J.H.Y. Induction of Apoptosis in HT-29 Colon Cancer Cells by Phloretin. J. Med. Food 2007, 10, 581–586. [Google Scholar] [CrossRef]
- Zielinska, D.; Laparra-Llopis, J.M.; Zielinski, H.; Szawara-Nowak, D.; Giménez-Bastida, J.A. Role of apple phytochemicals, phloretin and phloridzin, in modulating processes related to intestinal inflammation. Nutrients 2019, 11, 1173. [Google Scholar] [CrossRef]
- Yang, K.-C.; Tsai, C.-Y.; Wang, Y.-J.; Wei, P.-L.; Lee, C.-H.; Chen, J.-H.; Wu, C.-H.; Ho, Y.-S. Apple polyphenol phloretin potentiates the anticancer actions of paclitaxel through induction of apoptosis in human hep G2 cells. Mol. Carcinog. 2008, 48, 420–431. [Google Scholar] [CrossRef]
- Azuma, K.; Ippoushi, K.; Nakayama, M.; Ito, H.; Higashio, H.; Terao, J. Absorption of Chlorogenic Acid and Caffeic Acid in Rats after Oral Administration. J. Agric. Food Chem. 2000, 48, 5496–5500. [Google Scholar] [CrossRef] [PubMed]
- Gonthier, M.-P.; Verny, M.-A.; Besson, C.; Rémésy, C.; Scalbert, A. Chlorogenic Acid Bioavailability Largely Depends on Its Metabolism by the Gut Microflora in Rats. J. Nutr. 2003, 133, 1853–1859. [Google Scholar] [CrossRef] [PubMed]
- Olthof, M.R.; Hollman, P.C.; Katan, M.B. Chlorogenic acid and caffeic acid are absorbed in humans. J. Nutr. 2001, 131, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Nardini, M.; Cirillo, E.; Natella, F.; Scaccini, C. Absorption of Phenolic Acids in Humans after Coffee Consumption. J. Agric. Food Chem. 2002, 50, 5735–5741. [Google Scholar] [CrossRef]
- Olthof, M.R.; Hollman, P.C.H.; Buijsman, M.N.; Van Amelsvoort, J.M.M.; Katan, M.B. Chlorogenic Acid, Quercetin-3-Rutinoside and Black Tea Phenols Are Extensively Metabolized in Humans. J. Nutr. 2003, 133, 1806–1814. [Google Scholar] [CrossRef]
- Huang, S.; Wang, L.-L.; Xue, N.-N.; Li, C.; Guo, H.-H.; Ren, T.-K.; Zhan, Y.; Li, W.-B.; Zhang, J.; Chen, X.-G.; et al. Chlorogenic acid effectively treats cancers through induction of cancer cell differentiation. Theranostics 2019, 9, 6745–6763. [Google Scholar] [CrossRef]
- Yamagata, K.; Izawa, Y.; Onodera, D.; Tagami, M. Chlorogenic acid regulates apoptosis and stem cell marker-related gene expression in A549 human lung cancer cells. Mol. Cell. Biochem. 2017, 441, 9–19. [Google Scholar] [CrossRef]
- Yan, Y.; Liu, N.; Hou, N.; Dong, L.; Li, J. Chlorogenic acid inhibits hepatocellular carcinoma in vitro and in vivo. J. Nutr. Biochem. 2017, 46, 68–73. [Google Scholar] [CrossRef]
- Hou, N.; Liu, N.; Han, J.; Yan, Y.; Li, J. Chlorogenic acid induces reactive oxygen species generation and inhibits the viability of human colon cancer cells. Anti-Cancer Drugs 2017, 28, 59–65. [Google Scholar] [CrossRef]
- Sapio, L.; Salzillo, A.; Illiano, M.; Ragone, A.; Spina, A.; Chiosi, E.; Pacifico, S.; Catauro, M.; Naviglio, S. Chlorogenic acid activates ERK1/2 and inhibits proliferation of osteosarcoma cells. J. Cell. Physiol. 2020, 235, 3741–3752. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi Ekbatan, S.; Li, X.Q.; Ghorbani, M.; Azadi, B.; Kubow, S. Chlorogenic acid and its microbial metabolites exert anti-proliferative effects, S-phase cell-cycle arrest and apoptosis in human colon cancer Caco-2 cells. Int. J. Mol. Sci. 2018, 19, 723. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Du, H.; Chen, P. Chlorogenic acid inhibits the proliferation of human lung cancer A549 cell lines by targeting annexin A2 in vitro and in vivo. Biomed. Pharmacother. 2020, 131, 110673. [Google Scholar] [CrossRef]
- Zeng, A.; Liang, X.; Zhu, S.; Liu, C.; Wang, S.; Zhang, Q.; Zhao, J.; Song, L. Chlorogenic acid induces apoptosis, inhibits metastasis and improves antitumor immunity in breast cancer via the NF-κB signaling pathway. Oncol. Rep. 2021, 45, 717–727. [Google Scholar] [CrossRef]
- Cai, Z.-Y.; Li, X.-M.; Liang, J.-P.; Xiang, L.-P.; Wang, K.-R.; Shi, Y.-L.; Yang, R.; Shi, M.; Ye, J.-H.; Lu, J.-L.; et al. Bioavailability of Tea Catechins and Its Improvement. Molecules 2018, 23, 2346. [Google Scholar] [CrossRef] [PubMed]
- Warden, B.A.; Smith, L.S.; Beecher, G.R.; Balentine, D.A.; Clevidence, B.A. Catechins Are Bioavailable in Men and Women Drinking Black Tea throughout the Day. J. Nutr. 2001, 131, 1731–1737. [Google Scholar] [CrossRef]
- Lin, L.-C.; Wang, M.-N.; Tseng, T.-Y.; Sung, A.J.-S.; Tsai, T.-H. Pharmacokinetics of (−)-Epigallocatechin-3-gallate in Conscious and Freely Moving Rats and Its Brain Regional Distribution. J. Agric. Food Chem. 2007, 55, 1517–1524. [Google Scholar] [CrossRef]
- Catterall, F.; King, L.J.; Clifford, M.N.; Ioannides, C. Bioavailability of dietary doses of3H-labelled tea antioxidants (+)-catechin and (−)-epicatechin in rat. Xenobiotica 2003, 33, 743–753. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Cao, J.; Cai, Z.; An, H.; Li, Y.; Peng, Y.; Chen, N.; Luo, A.; Tao, H.; Li, K. Epigallocatechin gallate induces chemopreventive effects on rats with diethylnitrosamine-induced liver cancer via inhibition of cell division cycle 25A. Mol. Med. Rep. 2020, 22, 3873–3885. [Google Scholar] [CrossRef]
- Rathore, K.; Choudhary, S.; Odoi, A.; Wang, H.-C.R. Green tea catechin intervention of reactive oxygen species-mediated ERK pathway activation and chronically induced breast cell carcinogenesis. Carcinogenesis 2011, 33, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Siddique, H.R.; Liao, D.J.; Mishra, S.K.; Schuster, T.; Wang, L.; Matter, B.; Campbell, P.M.; Villalta, P.; Nanda, S.; Deng, Y.; et al. Epicatechin-rich cocoa polyphenol inhibits Kras-activated pancreatic ductal carcinoma cell growth in vitro and in a mouse model. Int. J. Cancer 2011, 131, 1720–1731. [Google Scholar] [CrossRef] [PubMed]
- Mayr, C.; Wagner, A.; Neureiter, D.; Pichler, M.; Jakab, M.; Illig, R.; Berr, F.; Kiesslich, T. The green tea catechin epigallocatechin gallate induces cell cycle arrest and shows potential synergism with cisplatin in biliary tract cancer cells. BMC Complement. Altern. Med. 2015, 15, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.; Dong, J. (-)-Epicatechin acts as a potent agonist of the membrane androgen receptor, ZIP9 (SLC39A9), to promote apoptosis of breast and prostate cancer cells. J. Steroid Biochem. Mol. Biol. 2021, 211, 105906. [Google Scholar] [CrossRef]
- Nifli, A.P.; Bosson-Kouamé, A.; Papadopoulou, N.; Kogia, C.; Kampa, M.; Castagnino, C.; Stournaras, C.; Vercauteren, J.; Castanas, E. Monomeric and oligomeric flavanols are agonists of membrane androgen receptors. Exp. Cell Res. 2005, 309, 329–339. [Google Scholar] [CrossRef]
- Pereyra-Vergara, F.; Olivares-Corichi, I.M.; Perez-Ruiz, A.G.; Luna-Arias, J.P.; García-Sánchez, J.R. Apoptosis induced by (−)-epicatechin in human breast cancer cells is mediated by reactive oxygen species. Molecules 2020, 25, 1020. [Google Scholar] [CrossRef]
- Ju, J.; Hong, J.; Zhou, J.N.; Pan, Z.; Bose, M.; Liao, J.; Yang, G.Y.; Liu, Y.Y.; Hou, Z.; Lin, Y.; et al. Inhibition of intestinal tumorigenesis in Apcmin/+ mice by (−)-epigallocatechin-3-gallate, the major catechin in green tea. Cancer Res. 2005, 65, 10623–10631. [Google Scholar] [CrossRef]
- Holt, R.R.; Lazarus, S.A.; Sullards, M.C.; Zhu, Q.Y.; Schramm, D.D.; Hammerstone, J.F.; Fraga, C.G.; Schmitz, H.H.; Keen, C.L. Procyanidin dimer B2 [epicatechin-(4β-8)-epicatechin] in human plasma after the consumption of a flavanol-rich cocoa. Am. J. Clin. Nutr. 2002, 76, 798–804. [Google Scholar] [CrossRef] [PubMed]
- Sano, A.; Yamakoshi, J.; Tokutake, S.; Tobe, K.; Kubota, Y.; Kikuchi, M. Procyanidin B1 Is Detected in Human Serum after Intake of Proanthocyanidin-rich Grape Seed Extract. Biosci. Biotechnol. Biochem. 2003, 67, 1140–1143. [Google Scholar] [CrossRef] [PubMed]
- Donovan, J.L.; Lee, A.; Manach, C.; Rios, L.; Morand, C.; Scalbert, A.; Remesy, C. Procyanidins are not bioavailable in rats fed a single meal containing a grapeseed extract or the procyanidin dimer B3. Br. J. Nutr. 2002, 87, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Deprez, S.; Mila, I.; Huneau, J.-F.; Tome, D.; Scalbert, A. Transport of Proanthocyanidin Dimer, Trimer, and Polymer Across Monolayers of Human Intestinal Epithelial Caco-2 Cells. Antioxidants Redox Signal. 2001, 3, 957–967. [Google Scholar] [CrossRef] [PubMed]
- Gossé, F.; Guyot, S.; Roussi, S.; Lobstein, A.; Fischer, B.; Seiler, N.; Raul, F. Chemopreventive properties of apple procyanidins on human colon cancer-derived metastatic SW620 cells and in a rat model of colon carcinogenesis. Carcinogenesis 2005, 26, 1291–1295. [Google Scholar] [CrossRef]
- Miura, T.; Chiba, M.; Kasai, K.; Nozaka, H.; Nakamura, T.; Shoji, T.; Kanda, T.; Ohtake, Y.; Sato, T. Apple procyanidins induce tumor cell apoptosis through mitochondrial pathway activation of caspase-3. Carcinogenesis 2007, 29, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Li, M.C.; Wang, F.R.; Mackenzie, G.G.; Oteiza, P.I. The inhibitory effect of ECG and EGCG dimeric procyanidins on colorectal cancer cells growth is associated with their actions at lipid rafts and the inhibition of the epidermal growth factor receptor signaling. Biochem. Pharmacol. 2020, 175, 113923. [Google Scholar] [CrossRef]
- Daveri, E.; Adamo, A.M.; Alfine, E.; Zhu, W.; Oteiza, P.I. Hexameric procyanidins inhibit colorectal cancer cell growth through both redox and non-redox regulation of the epidermal growth factor signaling pathway. Redox Biol. 2020, 38, 101830. [Google Scholar] [CrossRef]
- Na, W.; Ma, B.; Shi, S.; Chen, Y.; Zhang, H.; Zhan, Y.; An, H. Procyanidin B1, a novel and specific inhibitor of Kv10. 1 channel, suppresses the evolution of hepatoma. Biochem. Pharmacol. 2020, 178, 114089. [Google Scholar] [CrossRef]
- Shilpi, A.; Parbin, S.; Sengupta, D.; Kar, S.; Deb, M.; Rath, S.K.; Pradhan, N.; Rakshit, M.; Patra, S.K. Mechanisms of DNA methyltransferase–inhibitor interactions: Procyanidin B2 shows new promise for therapeutic intervention of cancer. Chem.-Biol. Interact. 2015, 233, 122–138. [Google Scholar] [CrossRef]
- Gouvêa, C.C.P.; Avelar, M. Procyanidin B2 cytotoxicity to MCF-7 human breast adenocarcinoma cells. Indian J. Pharm. Sci. 2012, 74, 351–355. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Deep, G.; Wempe, M.F.; Surek, J.; Kumar, A.; Agarwal, R.; Agarwal, C. Procyanidin B2 3, 3 ″-di-O-gallate induces oxidative stress-mediated cell death in prostate cancer cells via inhibiting MAP kinase phosphatase activity and activating ERK1/2 and AMPK. Mol. Carcinog. 2018, 57, 57–69. [Google Scholar] [CrossRef]
- Maldonado-Celis, M.E.; Bousserouel, S.; Gossé, F.; Minker, C.; Lobstein, A.; Raul, F. Differential induction of apoptosis by apple procyanidins in TRAIL-sensitive human colon tumor cells and derived TRAIL-resistant metastatic cells. J. Cancer Mol. 2009, 5, 21–30. [Google Scholar]
- Moyle, C.W.A.; Cerezo, A.B.; Winterbone, M.S.; Hollands, W.J.; Alexeev, Y.; Needs, P.W.; Kroon, P.A. Potent inhibition of VEGFR-2 activation by tight binding of green tea epigallocatechin gallate and apple procyanidins to VEGF: Relevance to angiogenesis. Mol. Nutr. Food Res. 2014, 59, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Furtado, N.A.J.C.; Pirson, L.; Edelberg, H.; Miranda, L.M.; Loira-Pastoriza, C.; Preat, V.; Larondelle, Y.; André, C.M. Pentacyclic Triterpene Bioavailability: An Overview of In Vitro and In Vivo Studies. Molecules 2017, 22, 400. [Google Scholar] [CrossRef] [PubMed]
- Patlolla, J.M.R.; Rao, C.V. Triterpenoids for cancer prevention and treatment: Current status and future prospects. Curr. Pharm. Biotechnol. 2012, 13, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Wang, G.J.; Wang, S.J.; Li, X.T.; Xu, Y.P.; Wang, S.P.; De Xiang, J.; Pan, S.R.; Cao, G.X.; Ye, W.C. Quantitative analysis of 23-hydroxybetulinic acid in mouse plasma using electrospray liquid chromatography/mass spectrometry. Rapid Commun. Mass Spectrom. 2005, 19, 1619–1623. [Google Scholar] [CrossRef]
- Jeong, D.W.; Kim, Y.H.; Kim, H.H.; Ji, H.Y.; Yoo, S.D.; Choi, W.R.; Lee, S.M.; Han, C.K.; Lee, H.S. Dose-linear pharmacokinetics of oleanolic acid after intravenous and oral administration in rats. Biopharm. Drug Dispos. 2007, 28, 51–57. [Google Scholar] [CrossRef]
- Kanellos, P.T.; Kaliora, A.C.; Gioxari, A.; Christopoulou, G.O.; Kalogeropoulos, N.; Karathanos, V.T. Absorption and Bioavailability of Antioxidant Phytochemicals and Increase of Serum Oxidation Resistance in Healthy Subjects Following Supplementation with Raisins. Plant Foods Hum. Nutr. 2013, 68, 411–415. [Google Scholar] [CrossRef]
- Zhu, Z.; Du, S.; Ding, F.; Guo, S.; Ying, G.; Yan, Z. Ursolic acid attenuates temozolomide resistance in glioblastoma cells by downregulating O6-methylguanine-DNA methyltransferase (MGMT) expression. Am. J. Transl. Res. 2016, 8, 3299–3308. [Google Scholar]
- Qiao, A.; Wang, Y.; Xiang, L.; Wang, C.; He, X. A novel triterpenoid isolated from apple functions as an anti-mammary tumor agent via a mitochondrial and caspase-independent apoptosis pathway. J. Agric. Food Chem. 2015, 63, 185–191. [Google Scholar] [CrossRef]
- Jiao, L.; Wang, S.; Zheng, Y.; Wang, N.; Yang, B.; Wang, D.; Yang, D.; Mei, W.; Zhao, Z.; Wang, Z. Betulinic acid suppresses breast cancer aerobic glycolysis via caveolin-1/NF-κB/c-Myc pathway. Biochem. Pharmacol. 2019, 161, 149–162. [Google Scholar] [CrossRef]
- Zeng, A.-Q.; Yu, Y.; Yao, Y.-Q.; Yang, F.-F.; Liao, M.; Song, L.-J.; Li, Y.-L.; Li, Y.-J.; Deng, Y.-L.; Yang, S.-P.; et al. Betulinic acid impairs metastasis and reduces immunosuppressive cells in breast cancer models. Oncotarget 2017, 9, 3794–3804. [Google Scholar] [CrossRef]
- Zheng, Y.; Liu, P.; Wang, N.; Wang, S.; Yang, B.; Li, M.; Chen, J.; Situ, H.; Xie, M.; Lin, Y.; et al. Betulinic Acid Suppresses Breast Cancer Metastasis by Targeting GRP78-Mediated Glycolysis and ER Stress Apoptotic Pathway. Oxidative Med. Cell. Longev. 2019, 2019, 8781690. [Google Scholar] [CrossRef]
- Luo, J.; Hu, Y.L.; Wang, H. Ursolic acid inhibits breast cancer growth by inhibiting proliferation, inducing autophagy and apoptosis, and suppressing inflammatory responses via the PI3K/AKT and NF-κB signaling pathways in vitro. Exp. Ther. Med. 2017, 14, 3623–3631. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Lin, Z.-M.; Ge, N.; Zhang, D.-L.; Huang, J.; Kong, F. Ursolic Acid Induces Apoptosis of Prostate Cancer Cells via the PI3K/Akt/mTOR Pathway. Am. J. Chin. Med. 2015, 43, 1471–1486. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Liu, R.H. Triterpenoids isolated from apple peels have potent antiproliferative activity and may be partially responsible for apple’s anticancer activity. J. Agric. Food Chem. 2007, 55, 4366–4370. [Google Scholar] [CrossRef]
- Qiu, B.; Simon, M.C. Oncogenes strike a balance between cellular growth and homeostasis. Semin. Cell Dev. Biol. 2015, 43, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Pang, Q.; Wang, Y.; Yan, X. Betulinic acid induces apoptosis by regulating PI3K/Akt signaling and mitochondrial pathways in human cervical cancer cells. Int. J. Mol. Med. 2017, 40, 1669–1678. [Google Scholar] [CrossRef]
- Shankar, E.; Zhang, A.; Franco, D.; Gupta, S. Betulinic acid-mediated apoptosis in human prostate cancer cells involves p53 and nuclear factor-kappa B (NF-κB) pathways. Molecules 2017, 22, 264. [Google Scholar] [CrossRef]
- Lee, D.; Lee, S.R.; Kang, K.S.; Ko, Y.; Pang, C.; Yamabe, N.; Kim, K.H. Betulinic Acid Suppresses Ovarian Cancer Cell Proliferation through Induction of Apoptosis. Biomolecules 2019, 9, 257. [Google Scholar] [CrossRef]
- Xu, Y.; Li, J.; Li, Q.J.; Feng, Y.L.; Pan, F. Betulinic acid promotes TRAIL function on liver cancer progression inhibition through p53/Caspase-3 signaling activation. Biomed. Pharmacother. 2017, 88, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.H.; Chen, S.Y.; Lu, C.C.; Lin, J.A.; Yen, G.C. Ursolic acid promotes apoptosis, autophagy, and chemosensitivity in gemcitabine-resistant human pancreatic cancer cells. Phytother. Res. 2020, 34, 2053–2066. [Google Scholar] [CrossRef]
- Li, H.-F.; Wang, X.-A.; Xiang, S.-S.; Hu, Y.-P.; Jiang, L.; Shu, Y.-J.; Li, M.-L.; Wu, X.-S.; Zhang, F.; Ye, Y.-Y.; et al. Oleanolic acid induces mitochondrial-dependent apoptosis and G0/G1 phase arrest in gallbladder cancer cells. Drug Des. Dev. Ther. 2015, 9, 3017–3030. [Google Scholar] [CrossRef]
- Shi, Y.; Song, Q.; Hu, D.; Zhuang, X.; Yu, S.; Teng, D. Oleanolic acid induced autophagic cell death in hepatocellular carcinoma cells via PI3K/Akt/mTOR and ROS-dependent pathway. Korean J. Physiol. Pharmacol. 2016, 20, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Shu, B.; Tian, Y.; Wang, G.; Wang, Y.; Wang, J.; Dong, Y. Oleanolic acid induces osteosarcoma cell apoptosis by inhibition of Notch signaling. Mol. Carcinog. 2018, 57, 896–902. [Google Scholar] [CrossRef] [PubMed]
- Lúcio, K.A.; Rocha, G.D.G.; Monção-Ribeiro, L.C.; Fernandes, J.; Takiya, C.M.; Gattass, C.R. Oleanolic Acid Initiates Apoptosis in Non-Small Cell Lung Cancer Cell Lines and Reduces Metastasis of a B16F10 Melanoma Model In Vivo. PLoS ONE 2011, 6, e28596. [Google Scholar] [CrossRef]
- Liao, L.; Liu, C.; Xie, X.; Zhou, J. Betulinic acid induces apoptosis and impairs migration and invasion in a mouse model of ovarian cancer. J. Food Biochem. 2020, 44, e13278. [Google Scholar] [CrossRef]
- Xiang, L.; Chi, T.; Tang, Q.; Yang, X.; Ou, M.; Chen, X.; Yu, X.; Chen, J.; Ho, R.J.; Shao, J.; et al. A pentacyclic triterpene natural product, ursolic acid and its prodrug US597 inhibit targets within cell adhesion pathway and prevent cancer metastasis. Oncotarget 2015, 6, 9295–9312. [Google Scholar] [CrossRef] [PubMed]
- Conway, G.; Zizyte, D.; Mondala, J.; He, Z.; Lynam, L.; Lecourt, M.; Barcia, C.; Howe, O.; Curtin, J. Ursolic Acid Inhibits Collective Cell Migration and Promotes JNK-Dependent Lysosomal Associated Cell Death in Glioblastoma Multiforme Cells. Pharmaceuticals 2021, 14, 91. [Google Scholar] [CrossRef]
- Koteswari, L.L.; Kumari, S.; Kumar, A.B.; Malla, R.R. A comparative anticancer study on procyanidin C1 against receptor positive and receptor negative breast cancer. Nat. Prod. Res. 2019, 34, 3267–3274. [Google Scholar] [CrossRef]
- Da Silva, M.; Jaggers, G.K.; Verstraeten, S.V.; Erlejman, A.G.; Fraga, C.G.; Oteiza, P.I. Large procyanidins prevent bile-acid-induced oxidant production and membrane-initiated ERK1/2, p38, and Akt activation in Caco-2 cells. Free. Radic. Biol. 2012, 52, 151–159. [Google Scholar] [CrossRef]
- Choy, Y.Y.; Fraga, M.; Mackenzie, G.G.; Waterhouse, A.L.; Cremonini, E.; Oteiza, P.I. The PI3K/Akt pathway is involved in procyanidin-mediated suppression of human colorectal cancer cell growth. Mol. Carcinog. 2016, 55, 2196–2209. [Google Scholar] [CrossRef] [PubMed]



| Phenolics Group | Phenolic Subgroup | Phenolic Compounds |
|---|---|---|
| Flavonoids | Anthocyanidins | Cyanidin 3-O-arabinoside |
| Cyanidin 3-O-galactoside | ||
| Cyanidin 3-O-xyloside | ||
| Cyanidin 3-O-xylgalactoside | ||
| Flavonols | Quercetin | |
| Quercetin 3-arabinopyranoside | ||
| Quercetin-3-arabinofuranoside | ||
| Quercetin 3-galactoside | ||
| Quercetin 3-glucoside | ||
| Quercetin 3-rhamnoside | ||
| Quercetin 3-rutinoside | ||
| Quercetin 3-xyloside | ||
| Dihydrochalcones | Phloretin | |
| Phloretin-2′-O-xyloglucoside | ||
| Phloridzin | ||
| 3-hydroxyphloridzin | ||
| Flavan-3-ols | Monomeric | |
| (+)-Catechin | ||
| (−)-Epicatechin | ||
| Oligomeric (Procyanidins) | ||
| Procyanidin B1 | ||
| Procyanidin B2 | ||
| Procyanidin B5 | ||
| Procyanidin B7 | ||
| Procyanidin C1 | ||
| Phenolic acids | Chlorogenic acid Hydroxy benzoic acid Hydroxy cinnamic acid |
| Monomeric Flavanols | Procyanidins | Flavonols | Dihydrochalcones | Chlorogenic Acid | Anthocyanins | Total Phenolics | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cultivar | Skin (%) | Flesh (%) | Skin (%) | Flesh (%) | Skin (%) | Flesh (%) | Skin (%) | Flesh (%) | Skin (%) | Flesh (%) | Skin (%) | Flesh (%) | Skin (%) | Flesh (%) |
| Monty’ Surprise * | 33 | 67 | 29 | 71 | 94 | 6 | 42 | 58 | 10 | 90 | 100 | n.e. | 37 | 63 |
| Braeburn | 19 | 81 | 21 | 79 | 99 | 1 | 8 | 92 | 1 | 99 | 100 | n.e. | 31 | 69 |
| Red Delicious | 37 | 63 | 36 | 64 | 94 | 6 | 40 | 60 | 2 | 98 | 100 | n.e. | 46 | 54 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nezbedova, L.; McGhie, T.; Christensen, M.; Heyes, J.; Nasef, N.A.; Mehta, S. Onco-Preventive and Chemo-Protective Effects of Apple Bioactive Compounds. Nutrients 2021, 13, 4025. https://doi.org/10.3390/nu13114025
Nezbedova L, McGhie T, Christensen M, Heyes J, Nasef NA, Mehta S. Onco-Preventive and Chemo-Protective Effects of Apple Bioactive Compounds. Nutrients. 2021; 13(11):4025. https://doi.org/10.3390/nu13114025
Chicago/Turabian StyleNezbedova, Linda, Tony McGhie, Mark Christensen, Julian Heyes, Noha Ahmed Nasef, and Sunali Mehta. 2021. "Onco-Preventive and Chemo-Protective Effects of Apple Bioactive Compounds" Nutrients 13, no. 11: 4025. https://doi.org/10.3390/nu13114025
APA StyleNezbedova, L., McGhie, T., Christensen, M., Heyes, J., Nasef, N. A., & Mehta, S. (2021). Onco-Preventive and Chemo-Protective Effects of Apple Bioactive Compounds. Nutrients, 13(11), 4025. https://doi.org/10.3390/nu13114025