The Seed of Industrial Hemp (Cannabis sativa L.): Nutritional Quality and Potential Functionality for Human Health and Nutrition





Abstract
1. Introduction
2. Biochemistry, Genetic, Taxonomy and Legal Status of C. sativa L.
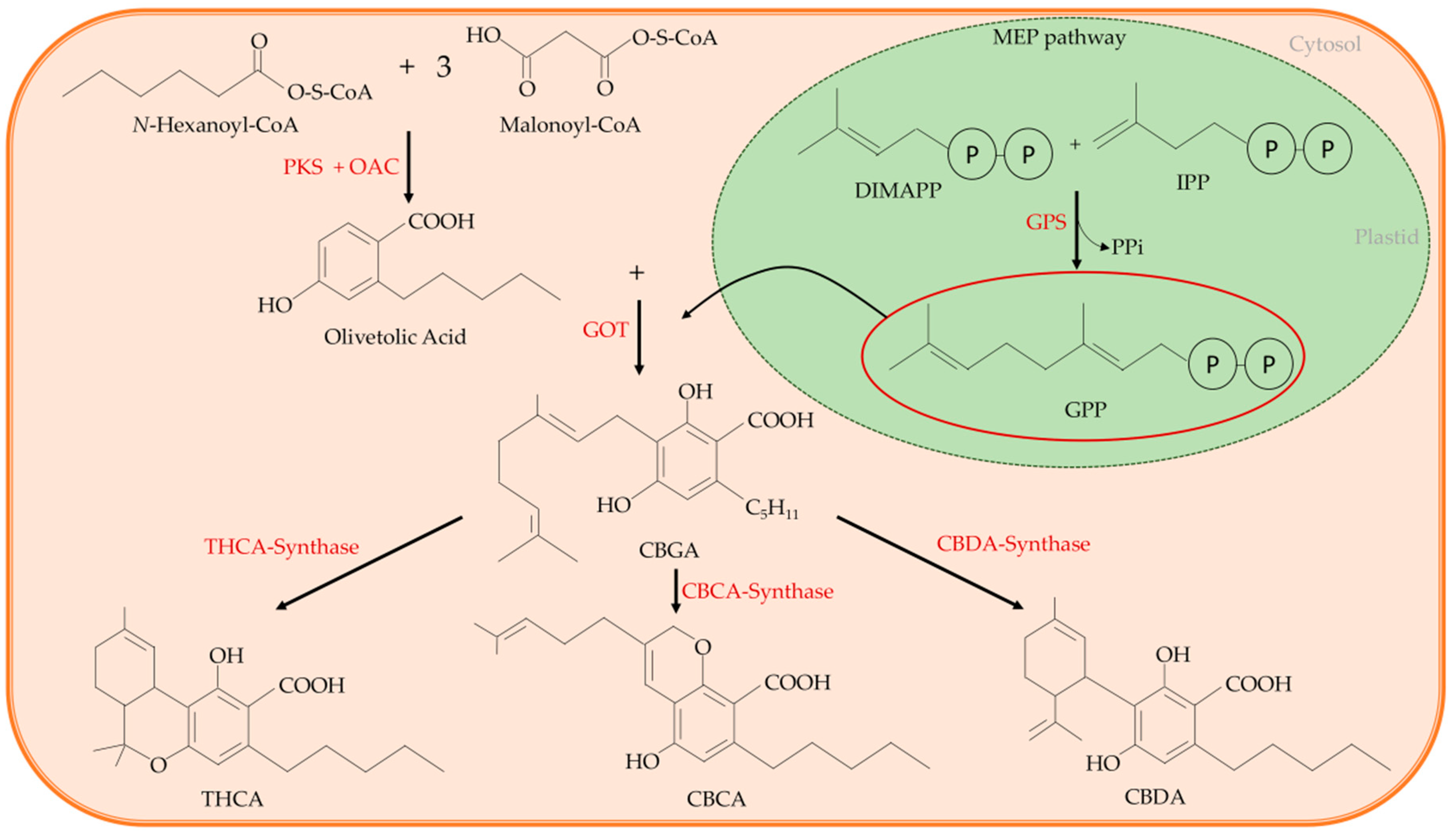
2.1. The Cannabinoids Synthesis in C. sativa L.
2.2. Chemical Phenotype and Taxonomy of C. sativa L.
- Chemotype I is characterized by a low CBD/THC ratio (0.00–0.05), due to high THC content (>0.3% of dry weight of the reproductive part of the female plant at flowering). This chemical phenotype is also known as “drug type”, “THC-predominant”, or C. sativa L. subsp. Indica and the varieties belonging to this chemotype are those commonly grown for narcotic/recreational purposes.
- Chemotype II has both the two main cannabinoids, CBD and THC, in a content ratio (CBD/THC) close to the unity (0.5–3.0), usually with a slight prevalence of CBD. This chemical phenotype is also named “intermediate type” or “THC-intermediate”, and the varieties belonging to this chemotype are mainly grown for medicinal use.
- Chemotype III is characterized by high CBD/THC ratio (15–25) due to high CBD amount and low THC content, not over than 0.3% of dry weight of the reproductive part of the female plant at flowering. This chemical phenotype is also known as “non-drug type”, “fibre-type”, “THC-predominant”, or C. sativa L. subsp. Sativa, and the varieties belonging to this chemotype are cultivated for industrial purposes, namely, for fibre, seeds, and their derivatives.
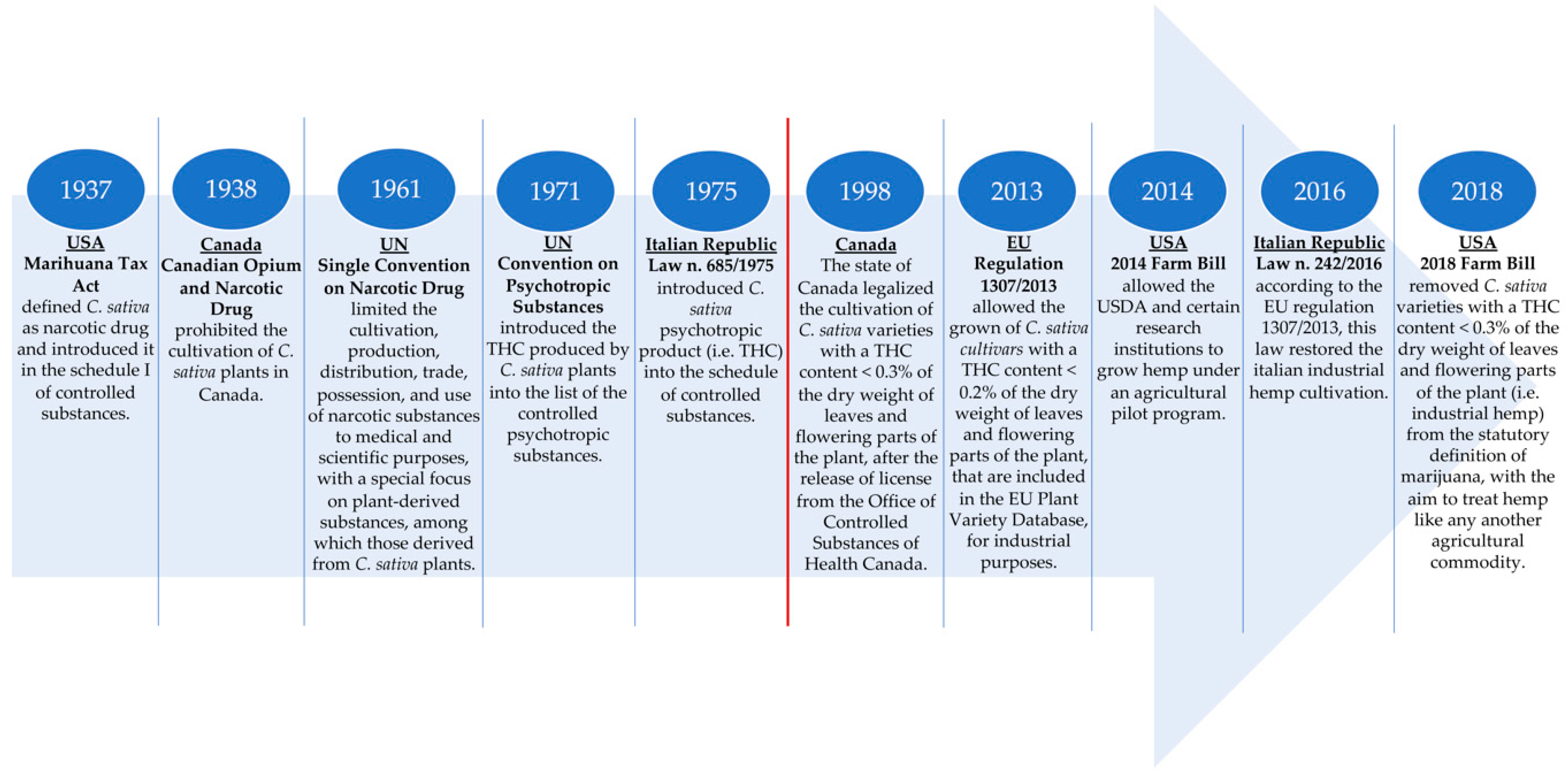
2.3. Legislation of C. sativa L.
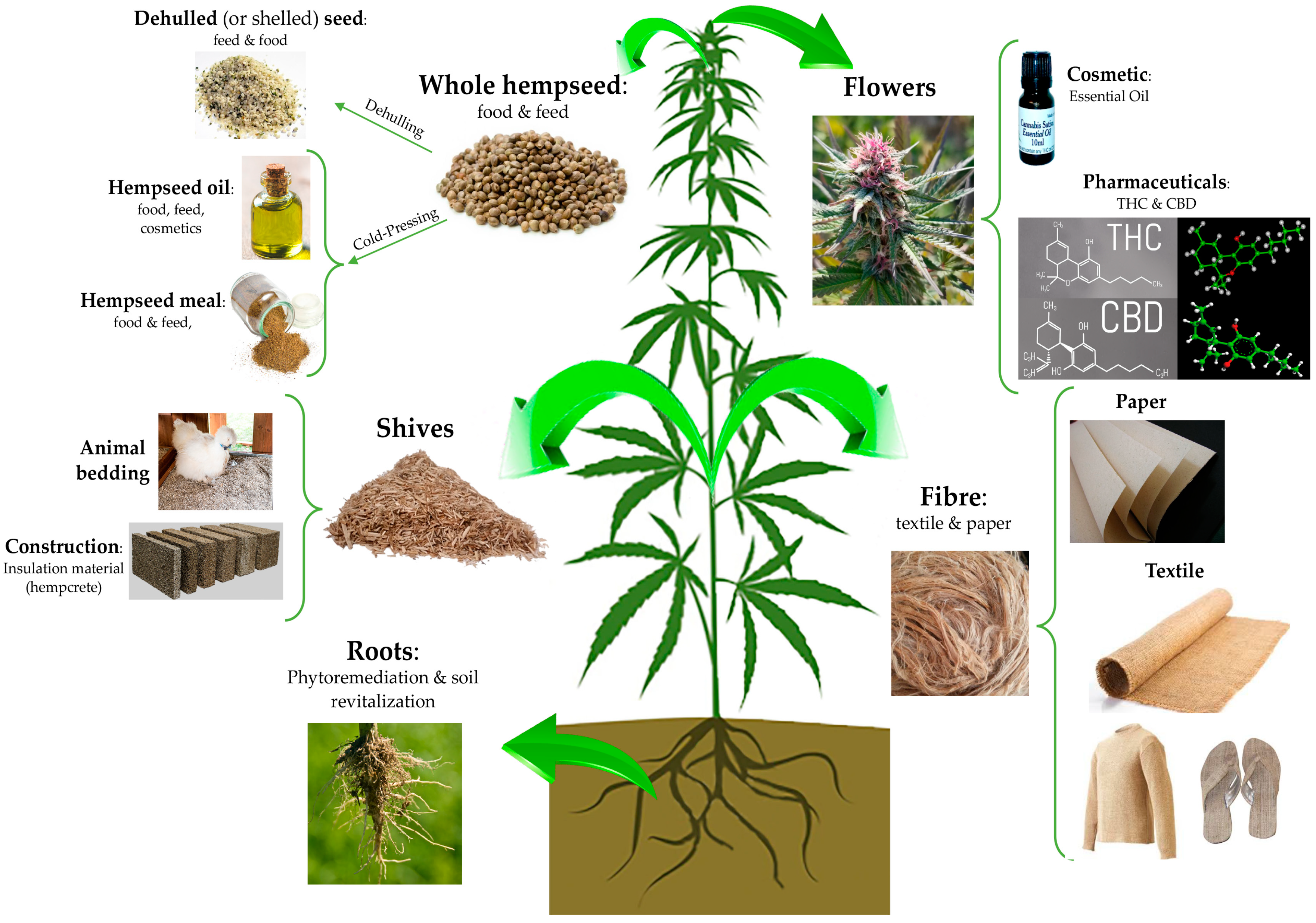
3. C. sativa L. Seed
3.1. Hempseed Nutritional Features
3.1.1. Hempseed Fat
Fatty Acid Composition
The Unsaponifiable Matter
3.1.2. Hempseed Proteins
3.1.3. Hempseed Carbohydrate and Dietary Fibre
3.1.4. Hempseed Minerals
3.1.5. Hempseed Antinutritional Compounds
- the highest content group including different soy (Glycine max) cvs with a TIs content ranging from 43 to 84 U/mg of the sample;
- the medium content group corresponding to common bean (Phaseolus vulgaris) varieties containing from 17 to 51 U/mg of the sample; and
- the lowest content group including lentil (Lens culinaris) and fava bean (Phaseolus vulgaris) cvs containing from 3 to 10 U/mg of the sample.
3.2. Hempseed Functional Features
3.2.1. Hempseed Phenolic Compounds
Composition Studies
Biological Effects
3.2.2. Hempseed Bioactive Peptides
4. Hempseed Dietary Supplementation
4.1. Animal Studies
4.2. Human Studies
5. Hempseed Foods
5.1. Hempseed Products as Livestock Supplement
5.2. Hempseed Products as Ingredient for Enriched/Fortified Foods
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mukherjee, A.; Roy, S.C.; De Bera, S.; Jiang, H.-E.; Li, X.; Li, C.-S.; Bera, S. Results of molecular analysis of an archaeological hemp (Cannabis sativa L.) DNA sample from North West China. Genet. Resour. Crop. Evol. 2008, 55, 481–485. [Google Scholar] [CrossRef]
- McPartland, J.M.; Guy, G.W.; Hegman, W. Cannabis is indigenous to Europe and cultivation began during the Copper or Bronze age: A probabilistic synthesis of fossil pollen studies. Veg. Hist. Archaeobotany 2018, 27, 635–648. [Google Scholar] [CrossRef]
- Irakli, M.; Tsaliki, E.; Kalivas, A.; Kleisiaris, F.; Sarrou, E.; Cook, C.M. Effect οf Genotype and Growing Year on the Nutritional, Phytochemical, and Antioxidant Properties of Industrial Hemp (Cannabis sativa L.) Seeds. Antioxidants 2019, 8, 491. [Google Scholar] [CrossRef] [PubMed]
- European Monitoring Centre for Drugs and Drug Addiction (EMCDDA). Cannabis Legislation in Europe: An Overview; Publications Office of the European Union: Luxembourg, 2018. [Google Scholar] [CrossRef]
- Marks, M.D.; Tian, L.; Wenger, J.P.; Omburo, S.N.; Soto-Fuentes, W.; He, J.; Gang, D.R.; Weiblen, G.D.; Dixon, R.A. Identification of candidate genes affecting Δ9-tetrahydrocannabinol biosynthesis in Cannabis sativa. J. Exp. Bot. 2009, 60, 3715–3726. [Google Scholar] [CrossRef]
- Sirikantaramas, S.; Taura, F.; Tanaka, Y.; Ishikawa, Y.; Morimoto, S.; Shoyama, Y. Tetrahydrocannabinolic Acid Synthase, the Enzyme Controlling Marijuana Psychoactivity, is Secreted into the Storage Cavity of the Glandular Trichomes. Plant Cell Physiol. 2005, 46, 1578–1582. [Google Scholar] [CrossRef]
- Onofri, C.; de Meijer, E.P.M.; Mandolino, G. Sequence heterogeneity of cannabidiolic- and tetrahydrocannabinolic acid-synthase in Cannabis sativa L. and its relationship with chemical phenotype. Phytochemistry 2015, 116, 57–68. [Google Scholar] [CrossRef]
- Staginnus, C.; Zörntlein, S.; de Meijer, E. A PCR marker Linked to a THCA synthase Polymorphism is a Reliable Tool to Discriminate Potentially THC-Rich Plants of Cannabis sativa L. J. Forensic Sci. 2014, 59, 919–926. [Google Scholar] [CrossRef]
- Citti, C.; Pacchetti, B.; Vandelli, M.A.; Forni, F.; Cannazza, G. Analysis of cannabinoids in commercial hemp seed oil and decarboxylation kinetics studies of cannabidiolic acid (CBDA). J. Pharm. Biomed. Anal. 2018, 149, 532–540. [Google Scholar] [CrossRef]
- Escrivá, Ú.; Andrés-Costa, M.J.; Andreu, V.; Picó, Y. Analysis of cannabinoids by liquid chromatography–mass spectrometry in milk, liver and hemp seed to ensure food safety. Food Chem. 2017, 228, 177–185. [Google Scholar] [CrossRef]
- Meng, Q.; Buchanan, B.; Zuccolo, J.; Poulin, M.-M.; Gabriele, J.; Baranowski, D.C. A reliable and validated LC-MS/MS method for the simultaneous quantification of 4 cannabinoids in 40 consumer products. PLoS ONE 2018, 13, e0196396. [Google Scholar] [CrossRef]
- Citti, C.; Linciano, P.; Panseri, S.; Vezzalini, F.; Forni, F.; Vandelli, M.A.; Cannazza, G. Cannabinoid Profiling of Hemp Seed Oil by Liquid Chromatography Coupled to High-Resolution Mass Spectrometry. Front. Plant Sci. 2019, 10, 120. [Google Scholar] [CrossRef] [PubMed]
- Christinat, N.; Savoy, M.-C.; Mottier, P. Development, validation and application of a LC-MS/MS method for quantification of 15 cannabinoids in food. Food Chem. 2020, 318, 126469. [Google Scholar] [CrossRef] [PubMed]
- Fellermeier, M.; Zenk, M.H. Prenylation of olivetolate by a hemp transferase yields cannabigerolic acid, the precursor of tetrahydrocannabinol. FEBS Lett. 1998, 427, 283–285. [Google Scholar] [CrossRef]
- Fellermeier, M.; Eisenreich, W.; Bacher, A.; Zenk, M.H. Biosynthesis of cannabinoids: Incorporation experiments with 13 C-labeled glucoses. Eur. J. Biochem. 2001, 268, 1596–1604. [Google Scholar] [CrossRef]
- Giroud, C. Analysis of Cannabinoids in Hemp Plants. CHIMIA Int. J. Chem. 2002, 56, 80–83. [Google Scholar] [CrossRef]
- Taura, F.; Tanaka, S.; Taguchi, C.; Fukamizu, T.; Tanaka, H.; Shoyama, Y.; Morimoto, S. Characterization of olivetol synthase, a polyketide synthase putatively involved in cannabinoid biosynthetic pathway. FEBS Lett. 2009, 583, 2061–2066. [Google Scholar] [CrossRef] [PubMed]
- Gagne, S.J.; Stout, J.M.; Liu, E.; Boubakir, Z.; Clark, S.M.; Page, J.E. Identification of olivetolic acid cyclase from Cannabis sativa reveals a unique catalytic route to plant polyketides. Proc. Natl. Acad. Sci. USA 2012, 109, 12811–12816. [Google Scholar] [CrossRef]
- Booth, J.K.; Page, J.E.; Bohlmann, J. Terpene synthases from Cannabis sativa. PLoS ONE 2017, 12, e0173911. [Google Scholar] [CrossRef]
- Pacifico, D.; Miselli, F.; Micheler, M.; Carboni, A.; Ranalli, P.; Mandolino, G. Genetics and Marker-assisted Selection of the Chemotype in Cannabis sativa L. Mol. Breed. 2006, 17, 257–268. [Google Scholar] [CrossRef]
- Rotherham, D.; Harbison, S.A. Differentiation of drug and non-drug Cannabis using a single nucleotide polymorphism (SNP) assay. Forensic Sci. Int. 2011, 207, 193–197. [Google Scholar] [CrossRef]
- Broséus, J.; Anglada, F.; Esseiva, P. The differentiation of fibre- and drug type Cannabis seedlings by gas chromatography/mass spectrometry and chemometric tools. Forensic Sci. Int. 2010, 200, 87–92. [Google Scholar] [CrossRef] [PubMed]
- de Meijer, E.P.M.; Bagatta, M.; Carboni, A.; Crucitti, P.; Moliterni, V.M.C.; Ranalli, P.; Mandolino, G. The inheritance of chemical phenotype in Cannabis sativa L. Genetics 2003, 163, 335–346. [Google Scholar] [PubMed]
- Cascini, F.; Farcomeni, A.; Migliorini, D.; Baldassarri, L.; Boschi, I.; Martello, S.; Amaducci, S.; Lucini, L.; Bernardi, J. Highly Predictive Genetic Markers Distinguish Drug-Type from Fiber-Type Cannabis sativa L. Plants 2019, 8, 496. [Google Scholar] [CrossRef] [PubMed]
- Kojoma, M.; Seki, H.; Yoshida, S.; Muranaka, T. DNA polymorphisms in the tetrahydrocannabinolic acid (THCA) synthase gene in “drug-type” and “fiber-type” Cannabis sativa L. Forensic Sci. Int. 2006, 159, 132–140. [Google Scholar] [CrossRef] [PubMed]
- De Meijer, E. The Chemical Phenotypes (Chemotypes) of Cannabis, 1st ed.; Handbooks in Psychopharmacology; Oxford University Press: Oxford, UK; New York, NY, USA, 2014; ISBN 978-0-19-966268-5. [Google Scholar]
- Hillig, K.W. Genetic evidence for speciation in Cannabis (Cannabaceae). Genet. Resour. Crop Evol. 2005, 52, 161–180. [Google Scholar] [CrossRef]
- Naraine, S.G.U.; Small, E.; Laursen, A.E.; Campbell, L.G. A multivariate analysis of morphological divergence of “seeds” (achenes) among ruderal, fibre, oilseed, dioecious/monoecious and marijuana variants of Cannabis sativa L. Genet. Resour. Crop Evol. 2020, 67, 703–714. [Google Scholar] [CrossRef]
- McPartland, J.M. Cannabis Systematics at the Levels of Family, Genus, and Species. Cannabis Cannabinoid Res. 2018, 3, 203–212. [Google Scholar] [CrossRef]
- Small, E.; Cronquist, A. A practical and natural taxonomy for Cannabis. Taxon 1976, 25, 405–435. [Google Scholar] [CrossRef]
- Johnson, R. Hemp as an Agricultural Commodity; CRS Report RL32725; Congressional Research Service: Washington, DC, USA, 2018; Available online: https://fas.org/sgp/crs/misc/RL32725.pdf (accessed on 12 March 2020).
- Porto, C.D.; Decorti, D.; Natolino, A. Potential Oil Yield, Fatty Acid Composition, and Oxidation Stability of the Hempseed Oil from Four Cannabis sativa L. Cultivars. J. Diet. Suppl. 2015, 12, 1–10. [Google Scholar] [CrossRef]
- Vera, C.L.; Hanks, A. Hemp Production in Western Canada. J. Ind. Hemp 2004, 9, 79–86. [Google Scholar] [CrossRef]
- Vonapartis, E.; Aubin, M.-P.; Seguin, P.; Mustafa, A.F.; Charron, J.-B. Seed composition of ten industrial hemp cultivars approved for production in Canada. J. Food Compos. Anal. 2015, 39, 8–12. [Google Scholar] [CrossRef]
- Sabet, K.A. Single Convention on Narcotic Drugs, 1961. In Encyclopedia of Drug Policy; SAGE Publications, Inc.: Thousand Oaks, CA, USA, 2011; ISBN 978-1-4129-7695-4. [Google Scholar] [CrossRef]
- United Nation Convention on Psychotropic Substances. Available online: https://www.unodc.org/unodc/en/treaties/psychotropics.html (accessed on 12 March 2020).
- European Commission EU Plant Variety Database (v.3.2). Available online: https://ec.europa.eu/food/plant/plant_propagation_material/plant_variety_catalogues_databases/search/public/index.cfm?event=SearchVariety&ctl_type=A&species_id=240&variety_name=&listed_in=0&show_current=on&show_deleted= (accessed on 15 March 2020).
- Pavlovic, R.; Panseri, S.; Giupponi, L.; Leoni, V.; Citti, C.; Cattaneo, C.; Cavaletto, M.; Giorgi, A. Phytochemical and Ecological Analysis of Two Varieties of Hemp (Cannabis sativa L.) Grown in a Mountain Environment of Italian Alps. Front. Plant Sci. 2019, 10, 1265. [Google Scholar] [CrossRef] [PubMed]
- Galasso, I.; Russo, R.; Mapelli, S.; Ponzoni, E.; Brambilla, I.M.; Battelli, G.; Reggiani, R. Variability in Seed Traits in a Collection of Cannabis sativa L. Genotypes. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Senate of Canada—The Cannabis Act in the Senate. Available online: https://www.sencanada.ca/en/sencaplus/news/cannabis-act/ (accessed on 15 January 2020).
- Kriese, U.; Schumann, E.; Weber, W.E.; Beyer, M.; Brühl, L.; Matthäus, B. Oil content, tocopherol composition and fatty acid patterns of the seeds of 51 Cannabis sativa L. genotypes. Euphytica 2004, 137, 339–351. [Google Scholar] [CrossRef]
- Karus, M.; Vogt, D. European hemp industry: Cultivation, processing and product lines. Euphytica 2004, 140, 7–12. [Google Scholar] [CrossRef]
- Mihoc, M.; Pop, G.; Alexa, E.; Radulov, I. Nutritive quality of romanian hemp varieties (Cannabis sativa L.) with special focus on oil and metal contents of seeds. Chem. Cent. J. 2012, 6, 122. [Google Scholar] [CrossRef]
- Lan, Y.; Zha, F.; Peckrul, A.; Hanson, B.; Johnson, B.; Rao, J.; Chen, B. Genotype x Environmental Effects on Yielding Ability and Seed Chemical Composition of Industrial Hemp (Cannabis sativa L.) Varieties Grown in North Dakota, USA. J. Am. Oil Chem. Soc. 2019, 96, 1417–1425. [Google Scholar] [CrossRef]
- Mattila, P.; Mäkinen, S.; Eurola, M.; Jalava, T.; Pihlava, J.-M.; Hellström, J.; Pihlanto, A. Nutritional Value of Commercial Protein-Rich Plant Products. Plant Foods Hum. Nutr. 2018, 73, 108–115. [Google Scholar] [CrossRef]
- House, J.D.; Neufeld, J.; Leson, G. Evaluating the Quality of Protein from Hemp Seed (Cannabis sativa L.) Products Through the use of the Protein Digestibility-Corrected Amino Acid Score Method. J. Agric. Food Chem. 2010, 58, 11801–11807. [Google Scholar] [CrossRef]
- Callaway, J.C. Hempseed as a nutritional resource: An overview. Euphytica 2004, 140, 65–72. [Google Scholar] [CrossRef]
- Oseyko, M.; Sova, N.; Lutsenko, M.; Kalyna, V. Chemical aspects of the composition of industrial hemp seed products. Ukr. Food J. 2019, 8, 544–559. [Google Scholar] [CrossRef]
- Siano, F.; Moccia, S.; Picariello, G.; Russo, G.; Sorrentino, G.; Di Stasio, M.; La Cara, F.; Volpe, M. Comparative Study of Chemical, Biochemical Characteristic and ATR-FTIR Analysis of Seeds, Oil and Flour of the Edible Fedora Cultivar Hemp (Cannabis sativa L.). Molecules 2018, 24, 83. [Google Scholar] [CrossRef] [PubMed]
- Vecka, M.; Staňková, B.; Kutová, S.; Tomášová, P.; Tvrzická, E.; Žák, A. Comprehensive sterol and fatty acid analysis in nineteen nuts, seeds, and kernel. SN Appl. Sci. 2019, 1, 1531. [Google Scholar] [CrossRef]
- da Silva Marineli, R.; Moraes, É.A.; Lenquiste, S.A.; Godoy, A.T.; Eberlin, M.N.; Maróstica, M.R., Jr. Chemical characterization and antioxidant potential of Chilean chia seeds and oil (Salvia hispanica L.). LWT Food Sci. Technol. 2014, 59, 1304–1310. [Google Scholar] [CrossRef]
- Teh, S.-S.; Birch, J. Physicochemical and quality characteristics of cold-pressed hemp, flax and canola seed oils. J. Food Compos. Anal. 2013, 30, 26–31. [Google Scholar] [CrossRef]
- Simopoulos, A.P. The Importance of the Omega-6/Omega-3 Fatty Acid Ratio in Cardiovascular Disease and Other Chronic Diseases. Exp. Biol. Med. (Maywood) 2008, 233, 674–688. [Google Scholar] [CrossRef]
- EFSA NDA Panel (EFSA Panel on Dietetic Products, Nutrition and Allergies). Scientific Opinion of the Panel on Dietetic products, Nutrition and Allergies on a request from European Commission related to labelling reference intake values for n-3 and n-6 polyunsaturated fatty acids. EFSA J. 2009, 8, 1461. [Google Scholar]
- Sergeant, S.; Rahbar, E.; Chilton, F.H. Gamma-linolenic acid, Dihommo-gamma linolenic, Eicosanoids and Inflammatory Processes. Eur. J. Pharmacol. 2016, 785, 77–86. [Google Scholar] [CrossRef]
- Guil-Guerrero, J.L.; Gómez-Mercado, F.; Ramos-Bueno, R.P.; González-Fernández, M.J.; Urrestarazu, M.; Jiménez-Becker, S.; de Bélair, G. Fatty acid profiles and sn -2 fatty acid distribution of γ-linolenic acid-rich Borago species. J. Food Compos. Anal. 2018, 66, 74–80. [Google Scholar] [CrossRef]
- Montserrat-de la Paz, S.; Marín-Aguilar, F.; García-Giménez, M.D.; Fernández-Arche, M.A. Hemp (Cannabis sativa L.) Seed Oil: Analytical and Phytochemical Characterization of the Unsaponifiable Fraction. J. Agric. Food Chem. 2014, 62, 1105–1110. [Google Scholar] [CrossRef]
- Uluata, S.; Özdemir, N. Antioxidant Activities and Oxidative Stabilities of Some Unconventional Oilseeds. J. Am. Oil Chem. Soc. 2012, 89, 551–559. [Google Scholar] [CrossRef]
- Oomah, B.D.; Busson, M.; Godfrey, D.V.; Drover, J.C.G. Characteristics of hemp (Cannabis sativa L.) seed oil. Food Chem. 2002, 76, 33–43. [Google Scholar] [CrossRef]
- Rezvankhah, A.; Emam-Djomeh, Z.; Safari, M.; Askari, G.; Salami, M. Investigation on the extraction yield, quality, and thermal properties of hempseed oil during ultrasound-assisted extraction: A comparative study. J. Food Process. Preserv. 2018, 42, e13766. [Google Scholar] [CrossRef]
- Rezvankhah, A.; Emam-Djomeh, Z.; Safari, M.; Askari, G.; Salami, M. Microwave-assisted extraction of hempseed oil: Studying and comparing of fatty acid composition, antioxidant activity, physiochemical and thermal properties with Soxhlet extraction. J. Food Sci. Technol. 2019, 56, 4198–4210. [Google Scholar] [CrossRef]
- EFSA NDA Panel (EFSA Panel on Dietetic Products, Nutrition and Allergies). Scientific Opinion on Dietary Reference Values for vitamin E as α-tocopherol. EFSA J. 2015, 13, 4149. [Google Scholar]
- Devi, V.; Khanam, S. Comparative study of different extraction processes for hemp (Cannabis sativa) seed oil considering physical, chemical and industrial-scale economic aspects. J. Clean. Prod. 2019, 207, 645–657. [Google Scholar] [CrossRef]
- Aladić, K. Cold Pressing and Supercritical CO2 Extraction of Hemp (Cannabis sativa) Seed Oil. Chem. Biochem. Eng. Q. 2015, 28, 481–490. [Google Scholar] [CrossRef]
- Latif, S.; Anwar, F. Physicochemical studies of hemp (Cannabis sativa) seed oil using enzyme-assisted cold-pressing. Eur. J. Lipid Sci. Technol. 2009, 111, 1042–1048. [Google Scholar] [CrossRef]
- Aladić, K.; Jarni, K.; Barbir, T.; Vidović, S.; Vladić, J.; Bilić, M.; Jokić, S. Supercritical CO2 extraction of hemp (Cannabis sativa L.) seed oil. Ind. Crop. Prod. 2015, 76, 472–478. [Google Scholar] [CrossRef]
- Tang, C.-H.; Ten, Z.; Wang, X.-S.; Yang, X.-Q. Physicochemical and Functional Properties of Hemp (Cannabis sativa L.) Protein Isolate. J. Agric. Food Chem. 2006, 54, 8945–8950. [Google Scholar] [CrossRef]
- Wang, X.-S.; Tang, C.-H.; Yang, X.-Q.; Gao, W.-R. Characterization, amino acid composition and in vitro digestibility of hemp (Cannabis sativa L.) proteins. Food Chem. 2008, 107, 11–18. [Google Scholar] [CrossRef]
- Ponzoni, E.; Brambilla, I.M.; Galasso, I. Genome-wide identification and organization of seed storage protein genes of Cannabis sativa. Biol. Plant 2018, 62, 693–702. [Google Scholar] [CrossRef]
- Mamone, G.; Picariello, G.; Ramondo, A.; Nicolai, M.A.; Ferranti, P. Production, digestibility and allergenicity of hemp (Cannabis sativa L.) protein isolates. Food Res. Int. 2019, 115, 562–571. [Google Scholar] [CrossRef] [PubMed]
- Raikos, V.; Duthie, G.; Ranawana, V. Denaturation and Oxidative Stability of Hemp Seed (Cannabis sativa L.) Protein Isolate as Affected by Heat Treatment. Plant Foods Hum. Nutr. 2015, 70, 304–309. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Pangloli, P.; Meng, X.; Dia, V.P. Effect of heating on the digestibility of isolated hempseed (Cannabis sativa L.) protein and bioactivity of its pepsin-pancreatin digests. Food Chem. 2020, 314, 126198. [Google Scholar] [CrossRef] [PubMed]
- Russo, R.; Reggiani, R. Evaluation of Protein Concentration, Amino Acid Profile and Antinutritional Compounds in Hempseed Meal from Dioecious and Monoecious Varieties. AJPS 2015, 06, 14–22. [Google Scholar] [CrossRef]
- Mattila, P.H.; Pihlava, J.-M.; Hellström, J.; Nurmi, M.; Eurola, M.; Mäkinen, S.; Jalava, T.; Pihlanto, A. Contents of phytochemicals and antinutritional factors in commercial protein-rich plant products. Food Qual. Saf. 2018, 2, 213–219. [Google Scholar] [CrossRef]
- Russo, R. Variability in Antinutritional Compounds in Hempseed Meal of Italian and French Varieties. Plant 2013, 1, 25. [Google Scholar] [CrossRef]
- Da Silva, B.P.; Anunciação, P.C.; da Silva Matyelka, J.C.; Della Lucia, C.M.; Martino, H.S.D.; Pinheiro-Sant’Ana, H.M. Chemical composition of Brazilian chia seeds grown in different places. Food Chem. 2017, 221, 1709–1716. [Google Scholar] [CrossRef]
- Multari, S.; Neacsu, M.; Scobbie, L.; Cantlay, L.; Duncan, G.; Vaughan, N.; Stewart, D.; Russell, W.R. Nutritional and Phytochemical Content of High-Protein Crops. J. Agric. Food Chem. 2016, 64, 7800–7811. [Google Scholar] [CrossRef]
- Lattimer, J.M.; Haub, M.D. Effects of Dietary Fiber and Its Components on Metabolic Health. Nutrients 2010, 2, 1266–1289. [Google Scholar] [CrossRef] [PubMed]
- Mueller, K.; Eisner, P.; Yoshie-Stark, Y.; Nakada, R.; Kirchhoff, E. Functional properties and chemical composition of fractionated brown and yellow linseed meal (Linum usitatissimum L.). J. Food Eng. 2010, 98, 453–460. [Google Scholar] [CrossRef]
- EFSA NDA Panel (EFSA Panel on Dietetic Products, Nutrition and Allergies). Overview on Dietary Reference Values for the EU population as derived by the EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Summary of Dietary Reference Values—Version 4 (September 2017). European Food Safety Authority, 2017. Available online: https://www.efsa.europa.eu/sites/default/files/assets/DRV_Summary_tables_jan_17.pdf (accessed on 16 June 2020).
- Deme, T.; Haki, G.; Retta, N.; Woldegiorgis, A.; Geleta, M. Mineral and Anti-Nutritional Contents of Niger Seed (Guizotia abyssinica (L.f.) Cass., Linseed (Linumusitatissimum L.) and Sesame (Sesamumindicum L.) Varieties Grown in Ethiopia. Foods 2017, 6, 27. [Google Scholar] [CrossRef] [PubMed]
- Köksal, A.I.; Artik, N.; Şimşek, A.; Güneş, N. Nutrient composition of hazelnut (Corylus avellana L.) varieties cultivated in Turkey. Food Chem. 2006, 99, 509–515. [Google Scholar] [CrossRef]
- Tapia, M.I.; Sánchez-Morgado, J.R.; García-Parra, J.; Ramírez, R.; Hernández, T.; González-Gómez, D. Comparative study of the nutritional and bioactive compounds content of four walnut (Juglans regia L.) cultivars. J. Food Compos. Anal. 2013, 31, 232–237. [Google Scholar] [CrossRef]
- Korkmaz, K.; Metin Kara, S.; Ozkutlu, F.; Gul, V. Monitoring of heavy metals and selected micronutrients in hempseeds from north-western Turkey. Afr. J. Agric. Res. 2010. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products, Nutrition and Allergies). Scientific opinion of the Panel on Contaminants in the Food Chain on a request from the European Commission on cadmium in food. EFSA J. 2009, 980, 1–139. [Google Scholar]
- Gupta, R.K.; Gangoliya, S.S.; Singh, N.K. Reduction of phytic acid and enhancement of bioavailable micronutrients in food grains. J. Food Sci. Technol. 2015, 52, 676–684. [Google Scholar] [CrossRef]
- Bohn, L.; Meyer, A.S.; Rasmussen, S.K. Phytate: Impact on environment and human nutrition. A challenge for molecular breeding. J. Zhejiang Univ. Sci. B 2008, 9, 165–191. [Google Scholar] [CrossRef]
- Weder, J.K.P.; Kahleyss, R. Reaction of Lentil Trypsin−Chymotrypsin Inhibitors with Human and Bovine Proteinases. J. Agric. Food Chem. 2003, 51, 8045–8050. [Google Scholar] [CrossRef]
- Guillamón, E.; Pedrosa, M.M.; Burbano, C.; Cuadrado, C.; de Sánchez, M.C.; Muzquiz, M. The trypsin inhibitors present in seed of different grain legume species and cultivar. Food Chem. 2008, 107, 68–74. [Google Scholar] [CrossRef]
- Bueno-Borges, L.B.; Sartim, M.A.; Gil, C.C.; Sampaio, S.V.; Rodrigues, P.H.V.; Regitano-d’Arce, M.A.B. Sacha inchi seeds from sub-tropical cultivation: Effects of roasting on antinutrients, antioxidant capacity and oxidative stability. J. Food Sci. Technol. 2018, 55, 4159–4166. [Google Scholar] [CrossRef]
- Russo, R.; Reggiani, R. Variability of antinutritive compounds in flaxseed flours. Int. J. Plant Biol. 2013, 4, 3. [Google Scholar] [CrossRef]
- Pojić, M.; Mišan, A.; Sakač, M.; Dapčević Hadnađev, T.; Šarić, B.; Milovanović, I.; Hadnađev, M. Characterization of Byproducts Originating from Hemp Oil Processing. J. Agric. Food Chem. 2014, 62, 12436–12442. [Google Scholar] [CrossRef] [PubMed]
- Francisco, I.A.; Pinotti, M.H.P. Cyanogenic glycosides in plants. Braz. Arch. Biol. Technol. 2000, 43, 487–492. [Google Scholar] [CrossRef]
- Joint FAO/WHO Food Standards Programme Proposed Draft Standard for Quinoa. 2018. Available online: http://www.fao.org/fao-who-codexalimentarius/sh-proxy/en/?lnk=1&url=https%253A%252F%252Fworkspace.fao.org%252Fsites%252Fcodex%252FCircular%252520Letters%252FCL%2525202018-25%252Fcl18_25e.pdf (accessed on 30 March 2020).
- Smeriglio, A.; Galati, E.M.; Monforte, M.T.; Lanuzza, F.; D’Angelo, V.; Circosta, C. Polyphenolic Compounds and Antioxidant Activity of Cold-Pressed Seed Oil from Finola Cultivar of Cannabis sativa L.: Polyphenolic Compounds and Antioxidant Activity of FHSO. Phytother. Res. 2016, 30, 1298–1307. [Google Scholar] [CrossRef]
- Yu, L.L.; Zhou, K.K.; Parry, J. Antioxidant properties of cold-pressed black caraway, carrot, cranberry, and hemp seed oils. Food Chem. 2005, 91, 723–729. [Google Scholar] [CrossRef]
- Faugno, S.; Piccolella, S.; Sannino, M.; Principio, L.; Crescente, G.; Baldi, G.M.; Fiorentino, N.; Pacifico, S. Can agronomic practices and cold-pressing extraction parameters affect phenols and polyphenols content in hempseed oils? Ind. Crop. Prod. 2019, 130, 511–519. [Google Scholar] [CrossRef]
- Moccia, S.; Siano, F.; Russo, G.L.; Volpe, M.G.; La Cara, F.; Pacifico, S.; Piccolella, S.; Picariello, G. Antiproliferative and antioxidant effect of polar hemp extracts (Cannabis sativa L., Fedora cv.) in human colorectal cell lines. Int. J. Food Sci. Nutr. 2019, 71, 410–423. [Google Scholar] [CrossRef]
- Frassinetti, S.; Moccia, E.; Caltavuturo, L.; Gabriele, M.; Longo, V.; Bellani, L.; Giorgi, G.; Giorgetti, L. Nutraceutical potential of hemp (Cannabis sativa L.) seeds and sprouts. Food Chem. 2018, 262, 56–66. [Google Scholar] [CrossRef]
- Chen, T.; He, J.; Zhang, J.; Li, X.; Zhang, H.; Hao, J.; Li, L. The isolation and identification of two compounds with predominant radical scavenging activity in hempseed (seed of Cannabis sativa L.). Food Chem. 2012, 134, 1030–1037. [Google Scholar] [CrossRef] [PubMed]
- Russo, R.; Reggiani, R. Phenolics and antioxidant activity in flax varieties with different productive attitude. Int. Food Res. J. 2015, 22, 1736–1739. [Google Scholar]
- Werz, O.; Seegers, J.; Schaible, A.M.; Weinigel, C.; Barz, D.; Koeberle, A.; Allegrone, G.; Pollastro, F.; Zampieri, L.; Grassi, G.; et al. Cannflavins from hemp sprouts, a novel cannabinoid-free hemp food product, target microsomal prostaglandin E2 synthase-1 and 5-lipoxygenase. PharmaNutrition 2014, 2, 53–60. [Google Scholar] [CrossRef]
- Yan, X.; Tang, J.; dos Santos Passos, C.; Nurisso, A.; Simões-Pires, C.A.; Ji, M.; Lou, H.; Fan, P. Characterization of Lignanamides from Hemp (Cannabis sativa L.) Seed and Their Antioxidant and Acetylcholinesterase Inhibitory Activities. J. Agric. Food Chem. 2015, 63, 10611–10619. [Google Scholar] [CrossRef] [PubMed]
- Bourjot, M.; Zedet, A.; Demange, B.; Pudlo, M.; Girard-Thernier, C. In Vitro Mammalian Arginase Inhibitory and Antioxidant Effects of Amide Derivatives Isolated from the Hempseed Cakes (Cannabis sativa). Planta Med. Int. Open 2017, 3, e64–e67. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, S.; Lou, H.; Fan, P. Chemical constituents of hemp (Cannabis sativa L.) seed with potential anti-neuroinflammatory activity. Phytochem. Lett. 2018, 23, 57–61. [Google Scholar] [CrossRef]
- Zhu, G.-Y.; Yang, J.; Yao, X.-J.; Yang, X.; Fu, J.; Liu, X.; Bai, L.-P.; Liu, L.; Jiang, Z.-H. (±)-Sativamides A and B, Two Pairs of Racemic Nor-Lignanamide Enantiomers from the Fruits of Cannabis sativa. J. Org. Chem. 2018, 83, 2376–2381. [Google Scholar] [CrossRef]
- Maiolo, S.A.; Fan, P.; Bobrovskaya, L. Bioactive constituents from cinnamon, hemp seed and polygonum cuspidatum protect against H2O2 but not rotenone toxicity in a cellular model of Parkinson’s disease. J. Tradit. Complementary Med. 2018, 8, 420–427. [Google Scholar] [CrossRef]
- Wang, S.; Luo, Q.; Zhou, Y.; Fan, P. CLG from Hemp Seed Inhibits LPS-Stimulated Neuroinflammation in BV2 Microglia by Regulating NF-κB and Nrf-2 Pathways. ACS Omega 2019, 4, 16517–16523. [Google Scholar] [CrossRef]
- Luo, Q.; Yan, X.; Bobrovskaya, L.; Ji, M.; Yuan, H.; Lou, H.; Fan, P. Anti-neuroinflammatory effects of grossamide from hemp seed via suppression of TLR-4-mediated NF-κB signaling pathways in lipopolysaccharide-stimulated BV2 microglia cells. Mol. Cell. Biochem. 2017, 428, 129–137. [Google Scholar] [CrossRef]
- Wang, S.; Luo, Q.; Fan, P. Cannabisin F from Hemp (Cannabis sativa) Seed Suppresses Lipopolysaccharide-Induced Inflammatory Responses in BV2 Microglia as SIRT1 Modulator. IJMS 2019, 20, 507. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wang, S.; Ji, J.; Lou, H.; Fan, P. Hemp (Cannabis sativa L.) Seed Phenylpropionamides Composition and Effects on Memory Dysfunction and Biomarkers of Neuroinflammation Induced by Lipopolysaccharide in Mice. ACS Omega 2018, 3, 15988–15995. [Google Scholar] [CrossRef]
- Logarušić, M.; Slivac, I.; Radošević, K.; Bagović, M.; Redovniković, I.R.; Srček, V.G. Hempseed protein hydrolysates’ effects on the proliferation and induced oxidative stress in normal and cancer cell lines. Mol. Biol. Rep. 2019, 46, 6079–6085. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Martin, N.M.; Toscano, R.; Villanueva, A.; Pedroche, J.; Millan, F.; Montserrat-de la Paz, S.; Millan-Linares, M.C. Neuroprotective protein hydrolysates from hemp (Cannabis sativa L.) seeds. Food Funct. 2019, 10, 6732–6739. [Google Scholar] [CrossRef] [PubMed]
- Teh, S.-S.; Bekhit, A.E.-D.A.; Carne, A.; Birch, J. Antioxidant and ACE-inhibitory activities of hemp (Cannabis sativa L.) protein hydrolysates produced by the proteases AFP, HT, Pro-G, actinidin and zingibain. Food Chem. 2016, 203, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.-H.; Wang, X.-S.; Yang, X.-Q. Enzymatic hydrolysis of hemp (Cannabis sativa L.) protein isolate by various proteases and antioxidant properties of the resulting hydrolysates. Food Chem. 2009, 114, 1484–1490. [Google Scholar] [CrossRef]
- Wang, X.-S.; Tang, C.-H.; Chen, L.; Yang, X.-Q. Characterization and Antioxidant Properties of Hemp Protein Hydrolysates Obtained with Neutrase®. Food Technol. Biotechnol. 2009, 47, 428–434. [Google Scholar]
- Girgih, A.; Alashi, A.; He, R.; Malomo, S.; Raj, P.; Netticadan, T.; Aluko, R. A Novel Hemp Seed Meal Protein Hydrolysate Reduces Oxidative Stress Factors in Spontaneously Hypertensive Rats. Nutrients 2014, 6, 5652–5666. [Google Scholar] [CrossRef]
- Girgih, A.T.; Udenigwe, C.C.; Aluko, R.E. In Vitro Antioxidant Properties of Hemp Seed (Cannabis sativa L.) Protein Hydrolysate Fractions. J. Am. Oil Chem. Soc. 2011, 88, 381–389. [Google Scholar] [CrossRef]
- Malomo, S.; Onuh, J.; Girgih, A.; Aluko, R. Structural and Antihypertensive Properties of Enzymatic Hemp Seed Protein Hydrolysates. Nutrients 2015, 7, 7616–7632. [Google Scholar] [CrossRef]
- Zanoni, C.; Aiello, G.; Arnoldi, A.; Lammi, C. Hempseed Peptides Exert Hypocholesterolemic Effects with a Statin-Like Mechanism. J. Agric. Food Chem. 2017, 65, 8829–8838. [Google Scholar] [CrossRef] [PubMed]
- Aiello, G.; Lammi, C.; Boschin, G.; Zanoni, C.; Arnoldi, A. Exploration of Potentially Bioactive Peptides Generated from the Enzymatic Hydrolysis of Hempseed Proteins. J. Agric. Food Chem. 2017, 65, 10174–10184. [Google Scholar] [CrossRef] [PubMed]
- Malomo, S.A.; Aluko, R.E. In Vitro Acetylcholinesterase-Inhibitory Properties of Enzymatic Hemp Seed Protein Hydrolysates. J. Am. Oil Chem. Soc. 2016, 93, 411–420. [Google Scholar] [CrossRef]
- Girgih, A.T.; He, R.; Malomo, S.; Offengenden, M.; Wu, J.; Aluko, R.E. Structural and functional characterization of hemp seed (Cannabis sativa L.) protein-derived antioxidant and antihypertensive peptides. J. Funct. Foods 2014, 6, 384–394. [Google Scholar] [CrossRef]
- Orio, L.P.; Boschin, G.; Recca, T.; Morelli, C.F.; Ragona, L.; Francescato, P.; Arnoldi, A.; Speranza, G. New ACE-Inhibitory Peptides from Hemp Seed (Cannabis sativa L.) Proteins. J. Agric. Food Chem. 2017, 65, 10482–10488. [Google Scholar] [CrossRef]
- Girgih, A.T.; He, R.; Aluko, R.E. Kinetics and Molecular Docking Studies of the Inhibitions of Angiotensin Converting Enzyme and Renin Activities by Hemp Seed (Cannabis sativa L.) Peptides. J. Agric. Food Chem. 2014, 62, 4135–4144. [Google Scholar] [CrossRef]
- Lu, R.-R.; Qian, P.; Sun, Z.; Zhou, X.-H.; Chen, T.-P.; He, J.-F.; Zhang, H.; Wu, J. Hempseed protein derived antioxidative peptides: Purification, identification and protection from hydrogen peroxide-induced apoptosis in PC12 cells. Food Chem. 2010, 123, 1210–1218. [Google Scholar] [CrossRef]
- Girgih, A.T.; Udenigwe, C.C.; Aluko, R.E. Reverse-phase HPLC Separation of Hemp Seed (Cannabis sativa L.) Protein Hydrolysate Produced Peptide Fractions with Enhanced Antioxidant Capacity. Plant Foods Hum. Nutr. 2013, 68, 39–46. [Google Scholar] [CrossRef]
- Ren, Y.; Liang, K.; Jin, Y.; Zhang, M.; Chen, Y.; Wu, H.; Lai, F. Identification and characterization of two novel α-glucosidase inhibitory oligopeptides from hemp (Cannabis sativa L.) seed protein. J. Funct. Foods 2016, 26, 439–450. [Google Scholar] [CrossRef]
- World Health Organization Cardiovascular Diseases (CVDs). Available online: https://www.who.int/news-room/fact-sheets/detail/cardiovascular-diseases-(cvds) (accessed on 15 April 2020).
- Mendis, S.; Puska, P.; Norrving, B.; World Health Organization; World Heart Federation; World Stroke Organization. Global Atlas on Cardiovascular Disease Prevention and Control; World Health Organization in collaboration with the World Heart Federation and the World Stroke Organization: Geneva, Switzerland, 2011; ISBN 978-92-4-156437-3. [Google Scholar]
- Kaushal, N.; Dhadwal, S.; Kaur, P. Ameliorative effects of hempseed (Cannabis sativa) against hypercholesterolemia associated cardiovascular changes. Nutr. Metab. Cardiovasc. Dis. 2020, 30, 330–338. [Google Scholar] [CrossRef]
- Karimi, I.; Hayatghaibi, H. Effect of Cannabis sativa L. Seed (Hempseed) on Serum Lipid and Protein Profiles of Rat. Pak. J. Nutr. 2006, 5, 585–588. [Google Scholar] [CrossRef]
- Seo, J.-H.; Jeong, E.-S.; Lee, K.-S.; Heo, S.-H.; Jeong, D.-G.; Lee, S.-J.; Kim, E.S.; Choi, Y.-K. Hempseed water extract ameliorates atherosclerosis in apolipoprotein E knockout mice. Food Sci. Biotechnol. 2012, 21, 927–932. [Google Scholar] [CrossRef]
- Lee, M.J.; Park, S.H.; Han, J.H.; Hong, Y.K.; Hwang, S.; Lee, S.; Kim, D.; Han, S.Y.; Kim, E.S.; Cho, K.S. The effects of hempseed meal intake and linoleic acid on Drosophila models of neurodegenerative diseases and hypercholesterolemia. Mol. Cells 2011, 31, 337–342. [Google Scholar] [CrossRef]
- Prociuk, M.A.; Edel, A.L.; Richard, M.N.; Gavel, N.T.; Ander, B.P.; Dupasquier, C.M.C.; Pierce, G.N. Cholesterol-induced stimulation of platelet aggregation is prevented by a hempseed-enriched dietThis article is one of a selection of papers published in the special issue Bridging the Gap: Where Progress in Cardiovascular and Neurophysiologic Research Meet. Can. J. Physiol. Pharmacol. 2008, 86, 153–159. [Google Scholar] [CrossRef]
- Gavel, N.; Edel, A.; Bassett, C.; Weber, A.; Merchant, M.; Rodriguez-Leyva, D.; Pierce, G. The effect of dietary hempseed on atherogenesis and contractile function in aortae from hypercholesterolemic rabbits. Acta Physiol. Hung. 2011, 98, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Saberivand, A.; Karimi, I.; Becker, L.A.; Moghaddam, A.; Azizi-Mahmoodjigh, S.; Yousefi, M.; Zavareh, S. The effects of Cannabis sativa L. seed (hempseed) in the ovariectomized rat model of menopause. Methods Find. Exp. Clin. Pharmacol. 2010, 32, 467. [Google Scholar] [CrossRef]
- Girgih, A.T.; Alashi, A.; He, R.; Malomo, S.; Aluko, R.E. Preventive and treatment effects of a hemp seed (Cannabis sativa L.) meal protein hydrolysate against high blood pressure in spontaneously hypertensive rats. Eur. J. Nutr. 2014, 53, 1237–1246. [Google Scholar] [CrossRef]
- Chen, N.-Y.; Liu, C.-W.; Lin, W.; Ding, Y.; Bian, Z.; Huang, L.; Huang, H.; Yu, K.-H.; Chen, S.-B.; Sun, Y.; et al. Extract of Fructus Cannabis Ameliorates Learning and Memory Impairment Induced by D-Galactose in an Aging Rats Model. Evid. Based Complementary Altern. Med. 2017, 2017, 1–13. [Google Scholar] [CrossRef]
- Li, X.-Y.; Liu, Y.-H.; Wang, B.; Chen, C.-Y.; Zhang, H.-M.; Kang, J.X. Identification of a sustainable two-plant diet that effectively prevents age-related metabolic syndrome and extends lifespan in aged mice. J. Nutr. Biochem. 2018, 51, 16–26. [Google Scholar] [CrossRef]
- Rezapour-Firouzi, S.; Shahabi, S.; Mohammadzadeh, A.; Tehrani, A.; Kheradmand, F.; Mazloomi, E. The potential effects of hemp seed/evening primrose oils on the mammalian target of rapamycin complex 1 and interferon-gamma genes expression in experimental autoimmune encephalomyelitis. Res. Pharm. Sci. 2018, 13, 523. [Google Scholar] [CrossRef]
- Rezapour-Firouzi, S.; Kheradmand, F.; Shahabi, S.; Tehrani, A.; Mazloomi, E.; Mohammadzadeh, A. Regulatory effects of hemp seed/evening primrose oil supplement in comparison with rapamycin on the expression of the mammalian target of rapamycin-complex 2 and interleukin-10 genes in experimental autoimmune encephalomyelitis. Res. Pharm. Sci. 2019, 14, 36. [Google Scholar] [CrossRef] [PubMed]
- Callaway, J.; Schwab, U.; Harvima, I.; Halonen, P.; Mykkänen, O.; Hyvönen, P.; Järvinen, T. Efficacy of dietary hempseed oil in patients with atopic dermatitis. J. Dermatol. Treat. 2005, 16, 87–94. [Google Scholar] [CrossRef]
- Schwab, U.S.; Callaway, J.C.; Erkkilä, A.T.; Gynther, J.; Uusitupa, M.I.J.; Järvinen, T. Effects of hempseed and flaxseed oils on the profile of serum lipids, serum total and lipoprotein lipid concentrations and haemostatic factors. Eur. J. Nutr. 2006, 45, 470–477. [Google Scholar] [CrossRef]
- Kaul, N.; Kreml, R.; Austria, J.A.; Richard, M.N.; Edel, A.L.; Dibrov, E.; Hirono, S.; Zettler, M.E.; Pierce, G.N. A Comparison of Fish Oil, Flaxseed Oil and Hempseed Oil Supplementation on Selected Parameters of Cardiovascular Health in Healthy Volunteers. J. Am. Coll. Nutr. 2008, 27, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Del Bo’, C.; Deon, V.; Abello, F.; Massini, G.; Porrini, M.; Riso, P.; Guardamagna, O. Eight-week hempseed oil intervention improves the fatty acid composition of erythrocyte phospholipids and the omega-3 index, but does not affect the lipid profile in children and adolescents with primary hyperlipidemia. Food Res. Int. 2019, 119, 469–476. [Google Scholar] [CrossRef]
- Rezapour-Firouzi, S.; Arefhosseini, S.R.; Mehdi, F.; Mehrangiz, E.-M.; Baradaran, B.; Sadeghihokmabad, E.; Mostafaei, S.; Fazljou, S.M.B.; Torbati, M.; Sanaie, S.; et al. Immunomodulatory and therapeutic effects of Hot-nature diet and co-supplemented hemp seed, evening primrose oils intervention in multiple sclerosis patients. Complementary Ther. Med. 2013, 21, 473–480. [Google Scholar] [CrossRef]
- Rezapour-Firouzi, S.; Arefhosseini, S.R.; Farhoudi, M.; Ebrahimi-Mamaghani, M.; Rashidi, M.-R.; Torbati, M.-A.; Baradaran, B. Association of Expanded Disability Status Scale and Cytokines after Intervention with Co-supplemented Hemp Seed, Evening Primrose Oils and Hot-natured Diet in Multiple Sclerosis Patients.pdf. BioImpacts 2013, 3, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Rezapour-Firouzi, S.; Arefhosseini, S.R.; Ebrahimi-Mamaghani, M.; Baradaran, B.; Sadeghihokmabad, E.; Torbati, M.; Mostafaei, S.; Chehreh, M.; Zamani, F. Activity of liver enzymes in multiple sclerosis patients with Hot-nature diet and co-supplemented hemp seed, evening primrose oils intervention. Complementary Ther. Med. 2014, 22, 986–993. [Google Scholar] [CrossRef]
- Jing, M.; Zhao, S.; House, J.D. Performance and tissue fatty acid profile of broiler chickens and laying hens fed hemp oil and HempOmegaTM. Poult. Sci. 2017, 96, 1809–1819. [Google Scholar] [CrossRef]
- Neijat, M.; Gakhar, N.; Neufeld, J.; House, J.D. Performance, egg quality, and blood plasma chemistry of laying hens fed hempseed and hempseed oil. Poult. Sci. 2014, 93, 2827–2840. [Google Scholar] [CrossRef]
- Gakhar, N.; Goldberg, E.; Jing, M.; Gibson, R.; House, J.D. Effect of feeding hemp seed and hemp seed oil on laying hen performance and egg yolk fatty acid content: Evidence of their safety and efficacy for laying hen diets. Poult. Sci. 2012, 91, 701–711. [Google Scholar] [CrossRef]
- Mierliță, D. Fatty acids profile and oxidative stability of eggs from laying hens fed diets containing hemp seed or hempseed cake. S. Afr. J. Anim. Sci. 2019, 49, 310. [Google Scholar] [CrossRef]
- Bazdidi, H.; Afzali, N.; Hosseini-Vashan, S.; Ghiase, S.; Malehaneh, M. Evaluation of Dietary Hempseed and Hempseed Oil on Performance, Egg Quality and Some Blood Parameters in Laying Hens after Peak Period. Poult. Sci. J. 2016, 4. [Google Scholar] [CrossRef]
- Konca, Y.; Yuksel, T.; Yalcin, H.; Beyzi, S.B.; Kaliber, M. Effects of heat-treated hempseed supplementation on performance, egg quality, sensory evaluation and antioxidant activity of laying hens. Br. Poult. Sci. 2019, 60, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Neijat, M.; Suh, M.; Neufeld, J.; House, J.D. Increasing Levels of Dietary Hempseed Products Leads to Differential Responses in the Fatty Acid Profiles of Egg Yolk, Liver and Plasma of Laying Hens. Lipids 2016, 51, 615–633. [Google Scholar] [CrossRef] [PubMed]
- Neijat, M.; Suh, M.; Neufeld, J.; House, J.D. Hempseed Products Fed to Hens Effectively Increased n-3 Polyunsaturated Fatty Acids in Total Lipids, Triacylglycerol and Phospholipid of Egg Yolk. Lipids 2016, 51, 601–614. [Google Scholar] [CrossRef]
- Raza, T.; Chand, N.; Khan, R.U.; Shahid, M.S.; Abudabos, A.M. Improving the fatty acid profile in egg yolk through the use of hempseed (Cannabis sativa), ginger (Zingiber officinale), and turmeric (Curcuma longa) in the diet of Hy-Line White Leghorns Arch. Anim. Breed. 2016, 59, 183–190. [Google Scholar] [CrossRef]
- Goldberg, E.M.; Gakhar, N.; Ryland, D.; Aliani, M.; Gibson, R.A.; House, J.D. Fatty Acid Profile and Sensory Characteristics of Table Eggs from Laying Hens Fed Hempseed and Hempseed Oil. J. Food Sci. 2012, 77, S153–S160. [Google Scholar] [CrossRef]
- Shahid, S.; Chand, N.; Khan, R.U.; Suhail, S.M.; Khan, N.A. Alternations in Cholesterol and Fatty Acids Composition in Egg Yolk of Rhode Island Red x Fyoumi Hens Fed with Hemp Seeds (Cannabis sativa L.). J. Chem. 2015, 2015, 1–6. [Google Scholar] [CrossRef]
- Park, S.-O.; Hwangbo, J.; Yuh, I.-S.; Park, B.-S. Gamma-linolenic acid egg production enriched with hemp seed oil and evening primrose oil in diet of laying hens. J. Environ. Biol. 2014, 35, 635–640. [Google Scholar]
- Juodka, R.; Juska, R.; Juskiene, V.; Leikus, R.; Stankeviciene, D.; Nainiene, R. The effect of feeding with hemp and Camelina cakes on the fatty acid profile of duck muscles. Arch. Anim. Breed. 2018, 61, 293–303. [Google Scholar] [CrossRef]
- Yalcin, H.; Konca, Y.; Durmuscelebi, F. Effect of dietary supplementation of hemp seed (Cannabis sativa L.) on meat quality and egg fatty acid composition of Japanese quail (Coturnix coturnix japonica). J. Anim. Physiol. Anim. Nutr. 2018, 102, 131–141. [Google Scholar] [CrossRef]
- Mierliță, D. Fatty acid profile and health lipid indices in the raw milk of ewes grazing part-time and hemp seed supplementation of lactating ewes. S. Afr. J. Anim. Sci. 2016, 46, 237–246. [Google Scholar] [CrossRef]
- Iannaccone, M.; Ianni, A.; Contaldi, F.; Esposito, S.; Martino, C.; Bennato, F.; De Angelis, E.; Grotta, L.; Pomilio, F.; Giansante, D.; et al. Whole blood transcriptome analysis in ewes fed with hemp seed supplemented diet. Sci. Rep. 2019, 9, 16192. [Google Scholar] [CrossRef] [PubMed]
- Pojić, M.; Dapčević Hadnađev, T.; Hadnađev, M.; Rakita, S.; Brlek, T. Bread Supplementation with Hemp Seed Cake: A By-Product of Hemp Oil Processing: Hemp Seed Cake in Bread Making. J. Food Qual. 2015, 38, 431–440. [Google Scholar] [CrossRef]
- Mierliță, D. Effects of diets containing hemp seeds or hemp cake on fatty acid composition and oxidative stability of sheep milk. S. Afr. J. Anim. Sci. 2018, 48, 504. [Google Scholar] [CrossRef]
- Cozma, A.; Andrei, S.; Pintea, A.; Miere, D.; Filip, L.; Loghin, F.; Ferlay, A. Effect of hemp seed oil supplementation on plasma lipid profile, liver function, milk fatty acid, cholesterol, and vitamin A concentrations in Carpathian goats. Czech J. Anim. Sci. 2016, 60, 289–301. [Google Scholar] [CrossRef]
- Švec, I.; Hrušková, M. Properties and nutritional value of wheat bread enriched by hemp products. Potravin. Slovak J. Food Sci. 2015, 9, 304–308. [Google Scholar] [CrossRef]
- Švec, I.; Hrušková, M. The Mixolab parameters of composite wheat/hemp flour and their relation to quality features. LWT Food Sci. Technol. 2015, 60, 623–629. [Google Scholar] [CrossRef]
- Hrušková, M.; Švec, I.; Jurinová, I. Chemometrics of Wheat Composites with Hemp, Teff, and Chia Flour: Comparison of Rheological Features. Int. J. Food Sci. 2013, 2013, 1–6. [Google Scholar] [CrossRef]
- Mikulec, A.; Kowalski, S.; Sabat, R.; Skoczylas, Ł.; Tabaszewska, M.; Wywrocka-Gurgul, A. Hemp flour as a valuable component for enriching physicochemical and antioxidant properties of wheat bread. LWT 2019, 102, 164–172. [Google Scholar] [CrossRef]
- Wang, Y.-Y.; Norajit, K.; Ryu, G.-H. Influence of Extruded Hemp-Rice Flour Addition on the Physical Properties of Wheat Bread. J. Food Sci. Nutr. 2011, 16, 62–66. [Google Scholar] [CrossRef]
- Wang, Y.-Y.; Norajit, K.; Kim, M.-H.; Kim, Y.-H.; Ryu, G.-H. Influence of extrusion condition and hemp addition on wheat dough and bread properties. Food Sci. Biotechnol. 2013, 22, 89–97. [Google Scholar] [CrossRef]
- Korus, J.; Witczak, M.; Ziobro, R.; Juszczak, L. Hemp (Cannabis sativa subsp. sativa) flour and protein preparation as natural nutrients and structure forming agents in starch based gluten-free bread. LWT 2017, 84, 143–150. [Google Scholar] [CrossRef]
- Korus, A.; Gumul, D.; Krystyjan, M.; Juszczak, L.; Korus, J. Evaluation of the quality, nutritional value and antioxidant activity of gluten-free biscuits made from corn-acorn flour or corn-hemp flour composites. Eur. Food Res. Technol. 2017, 243, 1429–1438. [Google Scholar] [CrossRef]
- Radočaj, O.; Dimić, E.; Tsao, R. Effects of Hemp (Cannabis sativa L.) Seed Oil Press-Cake and Decaffeinated Green Tea Leaves (Camellia sinensis) on Functional Characteristics of Gluten-Free Crackers. J. Food Sci. 2014, 79, C318–C325. [Google Scholar] [CrossRef]
- Norajit, K.; Gu, B.-J.; Ryu, G.-H. Effects of the addition of hemp powder on the physicochemical properties and energy bar qualities of extruded rice. Food Chem. 2011, 129, 1919–1925. [Google Scholar] [CrossRef]
- Zając, M.; Świątek, R. The effect of hemp seed and linseed addition on the quality of liver pâtés. Acta Sci. Pol. Technol. Aliment. 2018, 17, 169–176. [Google Scholar] [CrossRef]
- Zając, M.; Guzik, P.; Kulawik, P.; Tkaczewska, J.; Florkiewicz, A.; Migdał, W. The quality of pork loaves with the addition of hemp seeds, de-hulled hemp seeds, hemp protein and hemp flour. LWT 2019, 105, 190–199. [Google Scholar] [CrossRef]
- Dabija, A.; Codină, G.G.; Gâtlan, A.-M.; Sănduleac, E.T.; Rusu, L. Effects of some vegetables proteins addition on yogurt quality. Sci. Study Res. Chem. Chem. Eng. Biotechnol. Food Ind. 2018, 13, 181–192. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| No | Registered Varieties | Origin | No | Registered Varieties | Origin |
|---|---|---|---|---|---|
| 1 | Adzelvieši | Latvia | 33 | Ivory | Nederland |
| 2 | Armanca | Romania | 34 | KC Bonusz | Hungary |
| 3 | Asso | Italy | 35 | KC Dora | Hungary |
| 4 | Austa SK | Latvia | 36 | KC Virtus | Hungary |
| 5 | Balaton | Hungary | 37 | KC Zuzana | Hungary |
| 6 | Beniko | Switzerland | 38 | KCA Borana | Hungary |
| Czech Republic | |||||
| Nederland | |||||
| Poland | |||||
| 7 | Bialobrzeskie | Czech Republic Polan | 39 | Kompoliti | Hungary |
| 8 | Cannakomp | Hungary | 40 | Kompoliti Hibrid TC | Hungary |
| 9 | Carma | Italy | 41 | Lipko | Hungary |
| 10 | Carmaleonte | Italy | 42 | Lovrin 110 | Romania |
| 11 | Chamaeleon | Nederland | 43 | Marcello | Nederland |
| 12 | Codimono | Italy | 44 | Markant | Nederland |
| 13 | Dacia Secuieni | Romania | 45 | Monoica | Switzerland |
| Hungary | |||||
| 14 | Delta-405 | Spanish | 46 | Orion 33 | France |
| 15 | Delta-Ilosa | Spanish | 47 | Rajan | Poland |
| 16 | Dioica 88 | France | 48 | Ratza | Romania |
| 17 | Earlina 8 FC | France | 49 | Santhica 23 | France |
| 18 | Eletta Campana | Italy | 50 | Santhica 27 | France |
| 19 | Epsilon 68 | France | 51 | Santhica 70 | France |
| 20 | Fedora 17 | Switzerland | 52 | Secuieni Jubileu | Romania |
| France | |||||
| 21 | Felina 32 | France | 53 | Silvana | Romania |
| 22 | Fibrante | Italy | 54 | Succesiv | Romania |
| 23 | Fibrol | Hungary | 55 | Teodora | Romania |
| 24 | Fibror 79 | France | 56 | Tiborszallasi | HungaryItaly |
| 25 | Finola | Finland | 57 | Tisza | Hungary |
| 26 | Futura 75 | France | 58 | Tygra | Poland |
| 27 | Futura 83 | France | 59 | Uniko B | Hungary |
| 28 | Fèrimon | Deutschland France | 60 | Uso-31 | Nederland |
| 29 | Glecia | Italy | 61 | Villanova | Italy |
| 30 | Gilana | Italy | 62 | Wielkopolskie | Poland |
| 31 | Glyana | Poland | 63 | Wojko | Poland |
| 32 | Henola | Poland | 64 | Zenit | Romania |
| Moisture | Fat | Proteins | CHO | Total DF | Insoluble DF | Soluble DF | Ash | Ref |
|---|---|---|---|---|---|---|---|---|
| 1.1–7.2 | 26.9–30.6 | 23.8–28.0 | n.a. | n.a. | n.a. | n.a. | 5.1–5.8 | [34] |
| 4.1–4.3 | 32.8–35.9 | 24.3–28.1 | 32.5–37.5 | n.a. | n.a. | n.a. | 4.9–6.1 | [44] |
| 6.7 ± 0.5 | 34.6 ± 1.2 | 25.6 ± 0.6 | 34.4 ± 1.5 | 33.8 ± 1.9 | 30.9 ± 1.5 | 2.9 ± 0.4 | 5.4 ± 0.3 | [45] |
| 4.0–9.2 | 25.4–33.0 | 21.3–27.5 | n.a. | n.a. | n.a. | n.a. | 3.7–5.9 | [46] |
| 6.5 | 35.5 | 24.8 | 27.6 | 27.6 | 22.2 | 5.4 | 5.6 | [47] |
| 8.4 ± 0.02 | 33.3 ± 0.1 | 22.5 ± 0.2 | n.a. | n.a. | n.a. | n.a. | 5.9 ± 0.03 | [48] |
| 7.3 ± 0.1 | 24.5 ± 2.0 | 24.8 ± 1.1 | 38.1 ± 2.5 | n.a. | n.a. | n.a. | 5.3 ± 0.6 | [49] |
| PA | SA | OA | LA | GLA | ALA | SDA | ∑ SFA | ∑ MUFA | ∑ PUFA | n-6/n-3 | Ref. |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 6.7–7.0 | 2.1–2.8 | 9.4–13.0 | 55.6–56.6 | 2.6–4.5 | 14.7–17.3 | n.a. | 9.6–10.3 | n.a. | n.a. | n.a. | [34] |
| 6.0–7.3 | 2.3–3.5 | 9.2–15.7 | 55.0–58.2 | 0.6–4.5 | 12.6–19.6 | 0.2–1.5 | 9.4–11.7 | 9.7–16.1 | 72.2–80.7 | 2.8–4.5 | [39] |
| 7.1–9.1 | 2.1–2.8 | 10.3–17.9 | 51.6–54.2 | 1.9–5.0 | 10.5–15.3 | n.a. | n.a. | n.a. | n.a. | 3.9–5.5 | [3] |
| 6.1–7.8 | 2.3–4.0 | 12.2–18.8 | 53.9–59.0 | 3.5–6.2 | 12.3–18.9 | n.a. | n.a. | n.a. | 72.0–78.6 | 3.2–5.0 | [44] |
| 5.0 | 2.0 | 9.0 | 56.0 | 4.0 | 22.0 | 2 | n.a. | n.a. | 84.0 | 2.5 | [47] |
| 7.0 ± 0.3 | 2.8 ± 0.4 | 12.7 ± 1.3 | 56.2 ± 3.5 | 2.9 ± 0.4 | 15.0 ± 1.1 | n.a. | 10.9 ± 0.5 | 13.1 ± 1.3 | 75.1 ± 3.7 | 4.0 | [49] |
| 5.6 ± 0.5 | 3.9 ± 0.4 | 16.2 ± 6.2 | 54.7 ± 4.1 | n.a. | 16.2 ± 4.0 | 0.5 ± 0.01 | 10.9 ± 0.7 | 17.5 ± 6.3 | 72 ± 4.3 | 3.2 | [50] |
| Type of Analyzed Sample | Total Tocopherols | α-Tocopherol | β-Tocopherol | γ-Tocopherol | δ-Tocopherol | Ref. |
|---|---|---|---|---|---|---|
| mg/100 g seed | ||||||
| Whole seed | n.a. | n.a. | n.a. | 21.21–294.9 | 62.0–115.7 | [34] |
| Oil | 14.33–34.04 | 0.70–3.05 | 0.06–0.30 | 15.42–29.40 | 0.51–2.49 | [41] |
| Whole seed | n.a. | n.a. | n.a. | 4.6–11.3 | 0.6–1.3 | [3] |
| Whole seed | 90 | 5 | n.a. | 85 | n.a. | [47] |
| Whole seed | n.a. | 0.35 ± 0.09 | n.a. | 0.55 ± 0.13 | n.a. | [49] |
| mg/100 g oil | ||||||
| Whole seed | 61–135 | n.a. | n.a. | n.a. | n.a. | [39] |
| Oil | 56.28–58.22 | 23.4–24.62 | 31.6–32.20 1 | 1.28–1.40 | [48] | |
| Oil | n.a. | 2.78 ± 0.01 | n.a. | 56.41 ± 0.02 | n.a. | [52] |
| Oil | 80.28 ± 4.50 | 3.22 ± 0.65 | 0.81 ± 0.16 | 73.38 ± 2.86 | 2.87 ± 0.83 | [57] |
| Oil | n.a. | 2.56 ± 0.06 | 0.60 ± 0.005 | 59.79 ± 1.21 | 3.97 ± 0.15 | [58] |
| Oil | 79.7 | 3.4 | 0.6 | 73.3 | 2.5 | [59] |
| Oil | 97.13 | 4.32 ± 0.65 | n.a. | 89.26 ± 1.92 | 3.55 ± 0.25 | [60] |
| Oil | 92.97 | 4.71 ± 0.54 | n.a. | 84.11 ± 1.74 | 4.15 ± 0.44 | [61] |
| Type of Analysed Sample | Total Phytosterols | β-Sitosterol | Stigmasterol | Campesterol | Ref. |
|---|---|---|---|---|---|
| mg/100 g seed | |||||
| Whole seed | n.a. | 53.61 ± 3.15 | 2.47 ± 0.25 | 11.54 ± 1.03 | [49] |
| Whole seed | 124 ± 12 | 79.7 ± 0.1 | 3.4 ± 0.9 | 7.3 ± 0.6 | [50] |
| mg/100 g oil | |||||
| Oil | 279.37 ± 12.43 | 190.5 ± 5.93 | 10.02 ± 0.75 | 50.57 ± 3.20 | [57] |
| Ala | Arg | Asp | Cys | Glu | Gly | His * | Ile * | Leu * | Lys * | Met * | Phe * | Pro | Ser | Thr * | Trp * | Tyr | Val * | Ref. |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1.28 | 3.10 | 2.78 | 0.41 | 4.57 | 1.14 | 0.71 | 0.98 | 1.72 | 1.03 | 0.58 | 1.17 | 1.15 | 1.27 | 0.88 | 0.20 | 0.86 | 1.28 | [47] |
| 1.23 | 2.76 | 2.13 | 0.31 | 4.58 | 1.24 | 0.64 | 0.62 | 1.53 | 1.28 | 0.50 | 1.02 | 1.14 | 1.27 | 0.85 | n.a. | 0.73 | 0.68 | [48] |
| 0.94 | 2.69 | 2.33 | 0.36 | 3.83 | 1.02 | 0.58 | 0.86 | 1.47 | 0.84 | 0.56 | 1.01 | 0.89 | 1.12 | 0.79 | n.a. | 0.78 | 1.13 | [45] |
| 0.96 | 2.28 | 2.39 | 0.41 | 3.74 | 1.06 | 0.55 | 0.80 | 1.49 | 0.86 | 0.56 | 1.03 | 0.90 | 1.19 | 1.01 | 0.23 | 0.68 | 1.14 | [46] |
| P 1 | K 1 | Mg 1 | Ca 1 | Na 1 | Fe 2 | Mn 2 | Zn 2 | Cu 2 | Cd 2 | Ref. |
|---|---|---|---|---|---|---|---|---|---|---|
| 1160 | 859 | 483 | 145 | 12 | 14 | 7 | 7 | 2 | n.a. | [47] |
| 890 | n.a. | 240 | 90 | n.a. | 7 | 6 | 6 | n.a. | n.a. | [48] |
| n.a. | 252 | 268 | 94 | 6.8 | 10 | 4 | 5 | 0.5 | n.a. | [49] |
| n.a. | 463–2821 | 237–694 | 144–955 | n.a. | 113–240 | 6–11 | 4–9 | n.a. | 0.1–0.4 | [43] |
| 1014–910 | 727–866 | 430–482 | 94–121 | 22–27 | 11–13 | 12–15 | 10–11 | 0.8–0.9 | n.a. | [44] |
| 1170 | 921 | 496 | 127 | n.a. | 4 | 11 | 7 | 1.9 | 0.0015 | [45] |
| Phytic Acid 1 | Condensed Tannins 2 | Trypsin Inhibitors 3 | Cyanogenic Glycosides 4 | Sapoins 2 | Ref. |
|---|---|---|---|---|---|
| 4.5–7.6 * | n.a. | 10.8–27.8 * | n.a. | n.a. | [39] |
| 6.2–7.7 * | 214.0–456.0 * | 10.8–27.7 * | 50.0–170.0 * | 47.0–70.0 * | [73] |
| 5.3 ± 0.4 * | 105.0 ± 4.0 ** | n.a. | n.a. | n.a. | [74] |
| 6.1–7.4 * | 135.0–214.0 * | 18.2–28.4 * | 60.0–240.0 * | 5.9–8.1 * | [75] |
| Type of Sample | TPC | Ref. |
|---|---|---|
| Whole hempseed | 13.68–51.60 | [34] |
| Whole hempseed | 3.82–7.80 | [3] |
| Whole hempseed | 0.77 ± 0.04 | [49] |
| Cold-pressed oil | 1.88 ± 0.003 | [52] |
| Whole hempseed | 0.96 ± 0.35 | [74] |
| Cold-pressed oil | 2.68 ± 0.09 | [95] |
| Cold-pressed oil | 0.44 ± 0.001 | [96] |
| Whole hempseed | 0.92 ± 0.04 | [98] |
| Whole hempseed | 2.33 ± 0.07 | [99] |
| Compound | Biological Effect | Ref. |
|---|---|---|
| N-trans-caffeolyltyramine | Strong in vitro scavenging DPPH radical and ORAC antioxidant activity; LDL protection against in vitro oxidation; In vitro AChE and arginase inhibitory activity; In vivo anti-neuroinflammatory action; In vitro prevention of H2O2-induced cell death. | [100] [103] [104] [107] [111] |
| N-trans-feryoryltyramine | In vitro scavenging DPPH radical activity; In vivo anti-neuroinflammatory action. | [103] [111] |
| N-trans-caffeolyoctopamine | In vivo anti-neuroinflammatory action. | [111] |
| N-trans-cumaroyltyramine | In vivo anti-neuroinflammatory action. | [111] |
| Cannabisin A | In vitro scavenging DPPH radical activity; In vivo anti-neuroinflammatory action. | [103] [111] |
| Cannabisin B | Strong in vitro scavenging DPPH radical and ORAC antioxidant activity; LDL protection against in vitro oxidation; In vitro arginase inhibitory activity. | [100] [104] [111] |
| Cannabisin C | In vitro scavenging DPPH radical activity; In vivo anti-neuroinflammatory action. | [103] [111] |
| Cannabisin D | In vitro scavenging DPPH radical activity; In vivo anti-neuroinflammatory action. | [103] [111] |
| Cannabisin E | In vivo anti-neuroinflammatory action | [111] |
| Cannabisin F | In vitro scavenging DPPH radical and ORAC antioxidant activity; In vivo anti-neuroinflammatory action; In vitro inhibition of the NFκB inflammatory pathway, activation of Nfr-2 antioxidant pathway and upregulation of SIRT1. | [104] [110] [111] |
| Cannabisin M | In vitro scavenging DPPH radical activity; In vivo anti-neuroinflammatory action. | [103] [111] |
| Isocannabisin N | In vivo anti-neuroinflammatory action | [111] |
| Grossamide | In vitro inhibition of the NFκB inflammatory pathway. In vivo anti-neuroinflammatory action; | [109] [111] |
| 3,3′-demethyl-grossamide | In vitro scavenging DPPH radical activity; In vitro AChE inhibitory activity; In vivo anti-neuroinflammatory action. | [103] [111] |
| 3,3′-demethylheliotropamide | In vitro AChE and arginase inhibitory activity. | [103] |
| Sativamides A | In vitro reduction of ER stress-induced cell death. | [106] |
| Sativamides B | In vitro reduction of ER stress-induced cell death. | [106] |
| 4-((E)-p-coumaroylamino)butan-1-ol | In vitro inhibition of TNF-α release. | [105] |
| Cumaroylaminobutanol glucopyranoside | In vitro inhibition of TNF-α release; In vitro inhibition of the NFκB inflammatory pathway and activation of Nfr-2 antioxidant pathway. | [105] [108] |
| Supplement Type | Purpose | Experimental Design | Results | Ref. |
|---|---|---|---|---|
| Hempseed meal hydrolysate (HMH) | Evaluation of the antioxidant ability of an HMH-containing diet to attenuate the plasma levels of some oxidative stress markers in growing and adult SHRs rats. | 8-week old SHRs (n = 8) fed control or 1% HMH-containing diets ad libitum for 8 weeks (for analysing the preventive effect). 20-week old SHRs (n = 4) fed control or HPH-containing diets ad libitum for 4 weeks (for analysing the treatment effect). | Dietary intervention with a specific HMH on young and adults SHRs exerted an antioxidant action by: • The increase of the SOD and CAT activities in both growing and adult SHRs; • The significant decrease of the TPx plasma level in both growing and adult SHRs and NTRs. | [117] |
| Hempseed | Investigation of the aspects of dietary hempseed against hyperlipidemia-associated CV risks. | 1 month of 10% hempseed supplemented high fat-diet on Wistar rats. | The 10% hempseed dietary treatment had an anti-hyperlipidaemic effect by inducing: • The serum level decreases of the cholesterol, LDL and triglycerides; • The increase of the HDL serum level; • A significant reduction in the aorta thickness; • Less intima damage foam cells formation and smooth muscle cell proliferation; • The remarkable decrease in the lipid deposition, plaque size, and lesion surface area distribution; • The decrease of the pro-inflammatory PGEs expression; • The increase of the anti-inflammatory PGDs expression. | [131] |
| Hempseed | Evaluation of the effect of hempseed feeding on lipid and protein profiles. | 20-days of hempseed supplementation (free access) on Wistar rats. | Short-term hempseed dietary intake can have hepatoprotective properties and beneficial effect on the prevention of CVD by favouring: • The decrease of the mean fasting serum LDL level; • The increase of the mean fasting serum HDL and total protein levels. | [132] |
| Hempseed water extract (HWE) | Evaluation of the anti-atherosclerotic activity of hempseed water extract in ApoE KO mice. | 14-days of intragastric inoculation of 300 µL of hempseed water extract or distilled water (control) to six-week-old, male ApoE KO (n = 7). | Dietary HWE ingestion exerted an anti-atherosclerotic effect through: • The reduction of the atherosclerotic plaque formation in the aortic sinuses; • The reduction of the serum GOT concentration, demonstrating less liver damage after the treatment; • The decrease of total serum cholesterol and LDL, and the increase of serum HDL; • The significant decrease of TNF-α serum level. | [133] |
| Hempseed Meal (HSM) | Investigation of the protective effects of the dietary intake of HSM and its LA and ALA components on AD and CVD Drosophila melanogaster models. | GMR-Aβ42 Drosophila melanogaster line (AD model) reared in cornmeal-soybean standard media with the LA, ALA, and GLA amounts present in the HSM (for the effect on AD phenotype). Wild-type Drosophila melanogaster larvae reared in HSM media with 35.1 μg/mL of cholesterol (for the CV effect). | Thanks to their LA and ALA contents, HSM dietary uptake may exert beneficial effects on AD and CVD through: • The suppression of suppressed Aβ42 cytotoxicity in the GMR-Aβ42 flies’ line reared on HSM mean (beneficial effect on AD); • The reduction of dietary cholesterol uptake observed in the wild-type flies reared on the cholesterol media (beneficial effect on CVD). | [134] |
| Hempseed | Investigation of the ability of dietary hempseed to reduce platelet aggregation under hypercholesterolemia. | Male New Zealand rabbits fed 125 g/day of control diet supplemented with 10% of hempseed and 0.5% of cholesterol for 8 weeks. | Hempseed supplementation to a hypercholesterolemic diet is able to normalize the platelet aggregation by increasing the GLA plasma level. | [135] |
| Hempseed | Investigation of the dietary hempseed ability to inhibit atherosclerosis and the associated vascular contractile dysfunction in a cholesterol-induced rabbit model. | Male albino New Zealand rabbits fed 125 g/day of control diet supplemented with 10% of hempseed and 0.5% of cholesterol (to induce atherosclerotic plaque formation) for 8 weeks. | Dietary hempseed supplementation did not generate protection against atherosclerotic plaque formation in hypercholesterolemic diet, but it provided mildly beneficial effects against contractile dysfunction associated with atherosclerotic vessels. | [136] |
| Hempseed | Investigation of the ability of hempseed dietary intake on the prevention of ovarian hormone deficiency-induced hypercholesterolemia in ovariectomized rats and on the complications of estrogen deficiency such as depression, anxiety, and bone loss problems. | Bilateral ovariectomized six-month-old female Wistar rats (n = 5) fed 1%, 2%, or 10% hempseed supplemented diets or a control diet (standard commercial diet) for 3 weeks. | Until 10% of hempseed dietary supplementation improved the post-ovariectomized complications in rats since it: • Prevented the accrual of blood calcium, preventing the bone loss; • Prevented the body weight gain induced by ovariectomy; • Conserved the TGs and TC plasma levels as those of the non-ovariectomized rats, preventing dyslipidaemia; • Resulted in an antidepressant activity. | [137] |
| Hempseed meal hydrolysate (HMH) | Evaluation of the effect of HMH-containing diet on the prevention and treatment of hypertension in growing SHRs and in SHRs with established hypertension. | 8-week old SHRs (n = 8) fed control or 1% HMH-containing diets ad libitum for 8 weeks (for the preventive effect). 20-week old SHRs (n = 4) fed control (without HPH) or HPH-containing diets, ad libitum for 4 weeks (for analysing the treatment effect). | Dietary intervention with a specific HMH on young and adult SHRs exerted a preventive and therapeutic effects since it led to: • The attenuation of the normal increases of SBP in young growing SHRs; • The significant reduction of SBP in adult SHRs; • The significant reduction of the plasma ACE and renin activities in both young and adult SHRs. | [138] |
| Hempseed and bitter vegetable (HB) | Evaluation of the beneficial effect of the HB diet on chronic diseases and on the slowing of the aging process. | 15-month-old C57BL/6 female mice (n = 10) fed low CHO, low SFA, and high PUFA content-HB diets containing hempseed and bitter vegetable (Sonchus oleraceus) (2:1) or control (western diet), ad libitum until only 20% of the mice in one group survived. | The HB diet is capable of promoting health and longevity since it: • Significantly increased the mice lifespan; • Improved the spatial learning and memory, and the locomotory activity; • Exerted beneficial effects on hepatic steatosis, hepatotoxicity, and plasma lipid profiles including the normalization of the size, weight, and colour of the liver and the reduction of hepatic lipid accumulation and of the ALT, AST, and TG levels. • Provided protective effects against the aging-related abnormalities in spleen morphology and inflammation; • Increased the antioxidant defences and decreased the oxidative stress; • Decreased the systemic inflammation by lowering the plasma level of the inflammatory cytokines (i.e., TNF-α, IL-1β, MCP-1 and IL-6), and by increasing the plasma level of anti-inflammatory IL-10; • Improved the gut microbiota profile leading to a significant decrease in the gut levels of E. coli and a significant increase in the gut levels of Bifidobacterium and Lactobacillum species after 11 weeks of dietary intervention; • Increased the sensitivity to insulin; • Increased the hepatic expression of genes associated with longevity (i.e., AMPK, Sirt1, Nrf-1, and FOXO3). | [140] |
| Hempseed oil and Evening primrose oil (HSO/EPO) | Evaluation of the MS’ therapeutic effect of co-supplementation with HSO/EPO. | 14 days of HSO/EPO oral administration on adult female C57BL/6 (n = 6) EAE mice (MS model) | The HSO/EPO dietary treatment can exert a beneficial effect on MS treatment since it is able to: • Act as an immunomodulator, down-regulating the mRNA expression of mTORC1 (which promotes Th1 and Th2 cells differentiation) and of the pro-inflammatory cytokines IFN- in lymph node; • Exert an anti-inflammatory action through increasing the mRNA expression of mTORC2 (which promotes Th2 cells differentiation) and of the anti-inflammatory cytokines IL-10 in lymph nodes; • Reduce the infiltration of the inflammatory cells in brain; • Promote the re-myelination of neurons’ myelin sheath. | [140,141] |
| Supplement Type | Purpose | Experimental Design | Results | Ref. |
|---|---|---|---|---|
| Cold-pressed hempseed oil | Evaluation of the effect of dietary hempseed oil and olive oil on plasma lipid profiles, TEWL, skin quality, and dermal medication usage in patients with atopic dermatitis. | Controlled, randomized single-blind crossover study, based on two intervention periods of 8-weeks last, separated by a 4-weeks washout period. Patients (n = 20) with a diagnosis of atopic dermatitis were randomly divided into two groups. Every patient of each group orally consumed 30 mL/day of olive or hempseed oils. • Inclusion criteria: BMI < 30 Kg/m2; 25–60 years old. • Exclusion criteria: assumption of lipid-lowering, anti-hypertensive or asthma medications, nutrient supplements, steroid-containing skin creams, oral cyclosporine, and use of solarium during the study and in the previous month. | A relatively short period of hempseed oil dietary consumption significantly improved skin quality and the atopic symptomology of the patients without any negative side effects or adverse reactions. The dietary treatment significantly increased the plasma level of LA, ALA, and GLA; significantly decreased the TEWL values and patients’ skin dryness, itchiness, and dermal medication usage. | [143] |
| Cold-pressed under nitrogen atmosphere hempseed oil | Evaluation and comparison of the effects of dietary hempseed oil and flaxseed oil on the profile of serum lipids, fasting serum total and lipoprotein lipid concentrations, plasma glucose and insulin concentrations, and hemostatic factors in healthy volunteers. | Controlled, randomized single-blind crossover study, based on two intervention periods of 4-weeks last, separated by a 4-weeks washout period. Male (n = 8) and female (n = 6) healthy volunteers were randomly divided into two groups. Every volunteer of each group orally consumed 30 mL/day of flax or hempseed oils. • Inclusion criteria: BMI < 30 Kg/m2; 25–60 years old; fasting serum concentration of TG < 3.5 mmol/L; fasting serum concentration of TC 5.0–7.5 mmol/L; fasting plasma glucose < 6.0 mmol/L. • Exclusion criteria: assumption of lipid-lowering, anti-hypertensive medications and nutrient supplements during the study and in the previous month. | In healthy subjects, 4-weeks daily of orally ingestion of hempseed oil: • Reduced the total plasma TGs concentration; • Significantly decreased the TC/HDL-C ratio, lowering the risk of coronary heart diseases; • Significantly changed the FA composition of serum TG and CE, increasing the LA, ALA, and GLA amounts and decreasing the OA content, whereas the AA and DHA amount did not differ; • Did not modify the plasma glucose, insulin, and hemostatic factor like fibrinogen, FVIIa, and PAI-1. | [144] |
| Hempseed oil capsule | Evaluation and comparison of the effects of the hempseed, flaxseed, and fish dietary oils on cardiovascular parameters (lipid profile, LDL oxidation, inflammatory markers and platelet aggregation) in healthy volunteers. | Double-blinded, placebo controlled clinical trial. Male (n = 34) and female (n = 54) healthy volunteers were randomly divided into 4 groups (placebo, fish oil, flaxseed oil, and hempseed oil). The volunteers of each group were orally daily supplemented with 2 capsules each containing 1 g of the relative treatment for 12 weeks. • Inclusion criteria: fasting TC levels <5.2 mM; non-smokers; consumption of fish not more than once a week and not more than 29.6 mL/day of alcohol consumption. • Exclusion criteria: usage of aspirin; ibuprofen; and/or other non-steroidal, anti-inflammatory medications as well as thyroid, anticoagulant, or lipid-lowering medication and the presence of menopause or chronic illness. | 2 × 1g hempseed oil capsule daily supplementation, administrated for 12 weeks did not significantly affect the ALA and LA plasma levels; TC, TG, LDL, and HDL amounts, or inflammatory (CRP, and TNF-α) and platelet aggregation analyzed markers (collagen and thrombin-induced platelet aggregation). | [145] |
| Hempseed oil soft capsule | Evaluation of the role of hempseed oil in the modulation of hyperlipidaemia and CVD risk in children and adolescents. | Randomized, controlled, two-arm parallel-group study. Children and adolescents (n = 36) with diagnosis of primary hyperlipidemia were randomly divided into two groups. Each subject of the test group was dietary supplemented with one hempseed oil soft capsule containing 3 g of oil for 8 weeks. The other group was used as control (without control). • Inclusion criteria: assessment of primary hyperlipidaemia; 6–16 years old. • Exclusion criteria: diagnosis of secondary hyperlipidaemia, and/or renal, endocrine, lipid, neurologic, onco-hematologic disorders requiring drug treatment; BMI > 85th percentile; smokers. | The 8-weeks daily intake of 3 g of hempseed oil: • Did not affect the levels of TC, LDL-C, HDL-C and TGs; • Did not change the values of body weight, BMI, and blood pressure; • Significantly modified the RBC FAs composition increasing the total n-3 PUFAs, n-3 LCPUFAs, and n-6 LCPUFAs, and decreasing the n-6/n-3 PUFAs ratio as well as the SFAs and MUFAs. | [146] |
| Hempseed oil and evening primrose oil (HSO/EPO, 9:1) co-supplementation | Evaluation of the therapeutic and protective effect of the Hot-natured diet co-supplemented with HSO/EPO or of the HSO/EPO dietary co-supplementation on MS patients. | Double blind, randomized trial. Female (n = 42) and male (n = 23) patients with diagnosis of RMSS were randomly divided into three groups. Each group (A, B, and C) was subjected to a Hot-natured diet co-supplemented with 18–21 g of HSO/EPO (9:1) (group A) or to 18–21 g olive oil supplementation (group B), or to 18–21 g of HSE/EPO (9:1) co-supplementation (group C) for 6 months. • Inclusion criteria: diagnosis of RRMS (EDSS < 6); 14–55 years old. • Exclusion criteria: diagnosis of secondary or primary progressive MS; pregnancy; corticosteroid treatment; and diagnosis of other chronic neurological and inflammatory diseases such as cancer, rheumatic diseases, and heart diseases. | 6 months of Hot-natured diet co-supplemented with HSO/EPO or of HSO/EPO co-supplementation significantly improved the MS clinical condition and inflammatory status of the patients, without any adverse effects. • Both treatments significantly improved the degree of Th2/Th1 ratio and the value of EDSS and of relapse rate; • Both treatments significantly increased the level of anti-inflammatory IL-4 cytokine; • Only the Hot-natured diet co-supplemented with HSO/EPO significantly decreased the level of pro-inflammatory IFN-γ and IL-17 cytokines; • The co-supplementation of HSO/EPO led to a decreasing (but not statistically significant) trend in the level of IFN-γ and IL-17 cytokines. | [147] |
| Hempseed oil and evening primrose oil (HSO/EPO, 9:1) co-supplementation | Evaluation of the Hot-natured diet co-supplemented with HSO/EPO in patients with RRMS on the type and level of produced cytokines. | Females (n = 16) and males (n = 7) patients with diagnosis of RRMS were subjected to a Hot-natured dietary regimen co-supplemented with 18–21 g of HSE/EPO (9:1), for 6 months. • Inclusion criteria: diagnosis of RRMS (EDSS < 6); 14–55 years old. • Exclusion criteria: diagnosis of secondary or primary progressive MS; pregnancy; corticosteroid treatment; and diagnosis of other chronic neurological and inflammatory diseases such as cancer, rheumatic diseases, and heart diseases. | 6 months of Hot-natured diet co-supplemented with HSO/EPO had a beneficial effect in RRMS patients by decreasing the Th1 cells and by promoting the Th2 cell responses. This dietary treatment led to: • A significant improvement of EDSS; • A significant decrease of the production of the pro-inflammatory cytokines (IFN-γ and IL-17); •A significant increase of the production of anti-inflammatory IL-4. | [148] |
| Hempseed oil and evening primrose oil (HSO/EPO, 9:1) co-supplementation | Investigation of the effect of the Hot-natured diet co-supplemented with HSO/EPO or of the solely HSO/EPO dietary co-supplementation on MS patient’s liver dysfunction. | Double blind, randomized trial. Female (n = 42) and male (n = 23) patients with diagnosis of RMSS were randomly divided into three groups. Each group (A, B, and C) was subjected to a Hot-natured diet co-supplemented with 18–21 g of HSO/EPO (9:1) (group A) or to 18–21 g olive oil supplementation (group B), or to 18-21 g of HSE/EPO (9:1) co-supplementation (group C) for 6 months. • Inclusion criteria: diagnosis of RRMS (EDSS < 6); 14–55 years old. • Exclusion criteria: diagnosis of secondary or primary progressive MS; pregnancy; corticosteroid treatment; and diagnosis of other chronic neurological and inflammatory diseases such as cancer, rheumatic diseases, and heart diseases. | 6 months of Hot-natured diet co-supplemented with HSO/EPO or of HSO/EPO co-supplementation significantly improved the MS patients’ liver dysfunction, without any adverse effects. • Both treatments significantly improved the EDSS value; • Both treatments significantly decrease the AST enzyme serum level; • Only the Hot-natured diet co-supplemented with HSO/EPO significantly decreased the ALT and GGT enzyme serum levels. | [149] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farinon, B.; Molinari, R.; Costantini, L.; Merendino, N. The Seed of Industrial Hemp (Cannabis sativa L.): Nutritional Quality and Potential Functionality for Human Health and Nutrition. Nutrients 2020, 12, 1935. https://doi.org/10.3390/nu12071935
Farinon B, Molinari R, Costantini L, Merendino N. The Seed of Industrial Hemp (Cannabis sativa L.): Nutritional Quality and Potential Functionality for Human Health and Nutrition. Nutrients. 2020; 12(7):1935. https://doi.org/10.3390/nu12071935
Chicago/Turabian StyleFarinon, Barbara, Romina Molinari, Lara Costantini, and Nicolò Merendino. 2020. "The Seed of Industrial Hemp (Cannabis sativa L.): Nutritional Quality and Potential Functionality for Human Health and Nutrition" Nutrients 12, no. 7: 1935. https://doi.org/10.3390/nu12071935
APA StyleFarinon, B., Molinari, R., Costantini, L., & Merendino, N. (2020). The Seed of Industrial Hemp (Cannabis sativa L.): Nutritional Quality and Potential Functionality for Human Health and Nutrition. Nutrients, 12(7), 1935. https://doi.org/10.3390/nu12071935