Thyroid-Gut-Axis: How Does the Microbiota Influence Thyroid Function?



{kind=link}
Abstract
1. Introduction
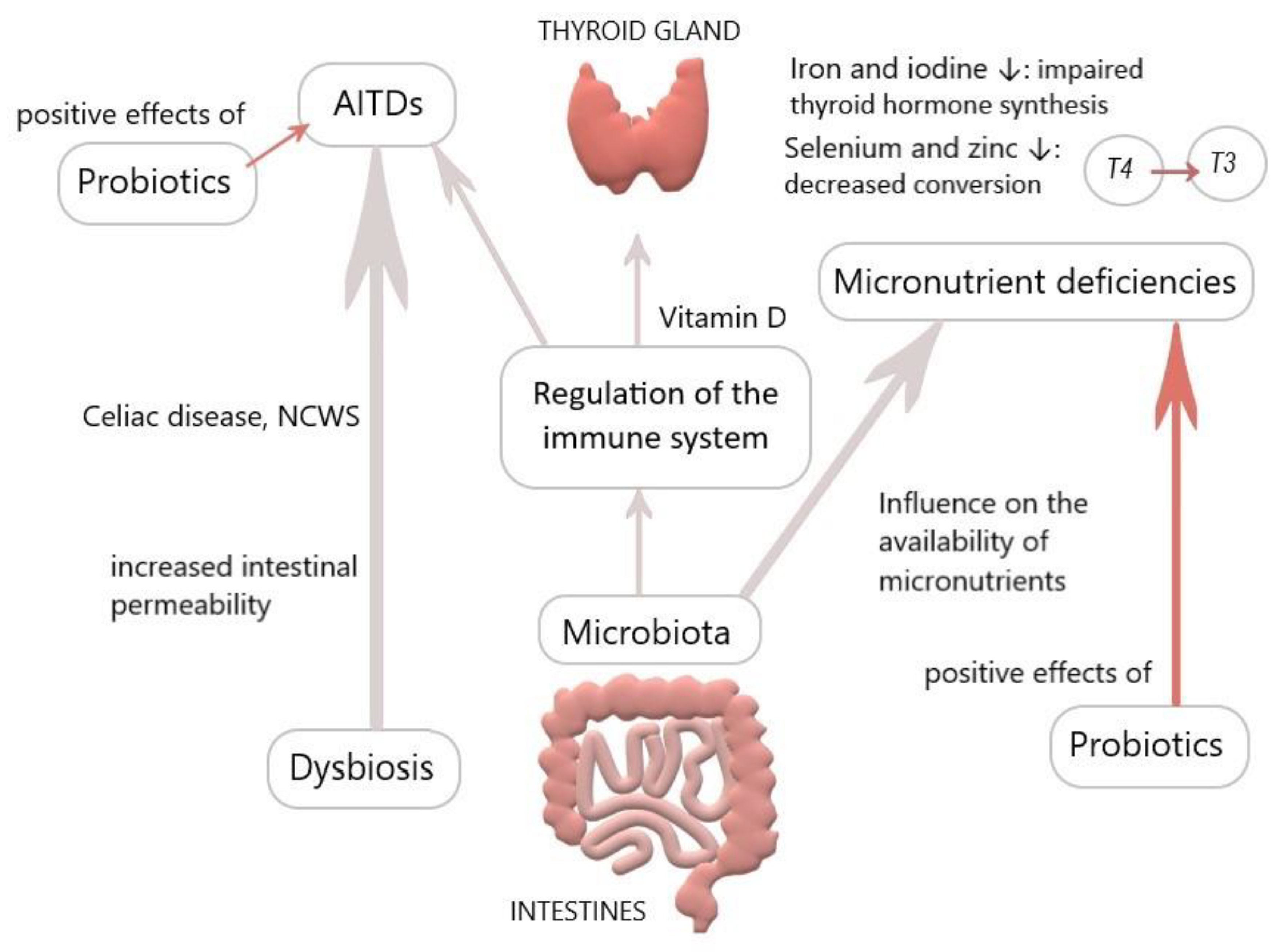
2. Intestinal Influences on the Thyroid
2.1. Celiac Disease
2.2. Non-Celiac Wheat Sensitivity (NCWS)
2.3. Micronutrients
2.3.1. Iodine
2.3.2. Iron
2.3.3. Zinc
2.3.4. Selenium
2.3.5. Vitamin D
2.4. Probiotics
2.5. Bariatric Surgery
2.6. Thyroid Cancer
3. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Virili, C.; Fallahi, P.; Antonelli, A.; Benvenga, S.; Centanni, M. Gut microbiota and Hashimoto’s thyroiditis. Rev. Endocr. Metab. Disord. 2018, 19, 293–300. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, G.L.V.; Leite, A.Z.; Higuchi, B.S.; Gonzaga, M.I.; Mariano, V.S. Intestinal dysbiosis and probiotic applications in autoimmune diseases. Immunology 2017, 152, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Rautava, S.; Luoto, R.; Salminen, S.; Isolauri, E. Microbial contact during pregnancy, intestinal colonization and human disease. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 565–576. [Google Scholar] [CrossRef]
- Fröhlich, E.; Wahl, R. Microbiota and thyroid interaction in health and disease. Trends Endocrinol. Metab. 2019, 30, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Maslowski, K.M.; MacKay, C.R. Diet, gut microbiota and immune responses. Nat. Immunol. 2011, 12, 5–9. [Google Scholar] [CrossRef]
- Fang, H.; Kang, J.; Zhang, D. Microbial production of vitamin B(12): A review and future perspectives. Microb. Cell Factories 2017, 16, 15. [Google Scholar] [CrossRef]
- Gu, Q. Biosynthesis of Vitamins by Probiotic Bacteria. Available online: https://doi.org/10.5772/63117 (accessed on 10 June 2020).
- Bastiaanssen, T.F.S.; Cowan, C.S.M.; Claesson, M.J.; Dinan, T.G.; Cryan, J.F. Making sense of … the microbiome in psychiatry. Int. J. Neuropsychopharmacol. 2019, 22, 37–52. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef]
- Köhling, H.L.; Plummer, S.F.; Marchesi, J.R.; Davidge, K.S.; Ludgate, M. The microbiota and autoimmunity: Their role in thyroid autoimmune diseases. Clin. Immunol. 2017, 183, 63–74. [Google Scholar] [CrossRef]
- Mori, K.; Nakagawa, Y.; Ozaki, H. Does the gut microbiota trigger hashimoto’s thyroiditis? Discov. Med. 2012, 14, 321–326. [Google Scholar]
- Caturegli, P.; De Remigis, A.; Rose, N.R. Hashimoto thyroiditis: Clinical and diagnostic criteria. Autoimmun. Rev. 2014, 13, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Hiromatsu, Y.; Satoh, H.; Amino, N. Hashimoto’s thyroiditis: History and future outlook. Hormones 2013, 12, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Morshed, S.A.; Latif, R.; Davies, T.F. Delineating the autoimmune mechanisms in Graves’ disease. Immunol. Res. 2012, 54, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Subekti, I.; Pramono, L.A. Current diagnosis and management of Graves’ disease. Acta Med. Indones. 2018, 50, 177–182. [Google Scholar]
- Covelli, D.; Ludgate, M. The thyroid, the eyes and the gut: A possible connection. J. Endocrinol. Invest. 2017, 40, 567–576. [Google Scholar] [CrossRef]
- Asarat, M.; Apostolopoulos, V.; Vasiljevic, T.; Donkor, O. Short-chain fatty acids regulate cytokines and Th17/Treg cells in human peripheral blood mononuclear cells in vitro short-chain fatty acids regulate cytokines and Th17/Treg. J. Mol. Cell. Immunol. 2016, 45, 205–222. [Google Scholar]
- Virili, C.; Centanni, M. Does microbiota composition affect thyroid homeostasis? Endocrine 2015, 49, 583–587. [Google Scholar] [CrossRef]
- Lerner, A.; Jeremias, P.; Matthias, T. Gut-thyroid axis and celiac disease. Endocr. Connect. 2017, 6, R52–R58. [Google Scholar] [CrossRef]
- Qin, J.; Zhou, J.; Fan, C.; Zhao, N.; Liu, Y.; Wang, S.; Cui, X.; Huang, M.; Guan, H.; Li, Y.; et al. Increased circulating Th17 but decreased CD4+Foxp3+ Treg and CD19+CD1dhiCD5+ Breg subsets in new-onset Graves’ disease. Biomed Res. Int. 2017, 2017, 1–8. [Google Scholar] [CrossRef]
- Caio, G.; Riegler, G.; Patt Urelli, M.; Facchiano, A.; De Magistris, L.; Sapone, A. Pathophysiology of non-celiac gluten sensitivity: Where are we now? Minerva Gastroenterol. E Dietol. 2017, 63, 16–21. [Google Scholar]
- Zhao, F.; Feng, J.; Li, J.; Zhao, L.; Liu, Y.; Chen, H.; Jin, Y.; Zhu, B.; Wei, Y. Alterations of the gut microbiota in hashimoto’s thyroiditis patients. Thyroid 2018, 28, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Ishaq, H.M.; Mohammad, I.S.; Guo, H.; Shahzad, M.; Hou, Y.J.; Ma, C.; Naseem, Z.; Wu, X.; Shi, P.; Xu, J. Molecular estimation of alteration in intestinal microbial composition in Hashimoto’s thyroiditis patients. Biomed. Pharmacother. 2017, 95, 865–874. [Google Scholar] [CrossRef] [PubMed]
- Lauritano, E.C.; Bilotta, A.L.; Gabrielli, M.; Scarpellini, E.; Lupascu, A.; Laginestra, A.; Novi, M.; Sottili, S.; Serricchio, M.; Cammarota, G.; et al. Association between hypothyroidism and small intestinal bacterial overgrowth. J. Clin. Endocrinol. Metab. 2007, 92, 4180–4184. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.; Distefano, J.J.; Huang, L.M.; Yamada, H.; Cahnmann, H.J. 5’- and 5-deiodinase activities in adult rat cecum and large bowel contents inhibited by intestinal microflora. Am. J. Physiol. Metab. 1993, 265, E521–E524. [Google Scholar] [CrossRef] [PubMed]
- Bernard, S.; Wostmann, D.S. Germfree and Gnotobiotic Animal Models, 1st ed.; Marsha Baker, A.D., Ed.; CRC Press: Notre Dame, IN, USA, 1996; ISBN 0-8493-400-X. [Google Scholar]
- Biesiekierski, J. What is gluten? J. Gastroenterol. Hepatol. 2017, 32, 78–81. [Google Scholar] [CrossRef]
- Valenti, S.; Corica, D.; Ricciardi, L.; Romano, C. Gluten-related disorders: Certainties, questions and doubts. Ann. Med. 2017, 49, 569–581. [Google Scholar] [CrossRef]
- Rotondi Aufiero, V.; Fasano, A.; Mazzarella, G. Non-celiac gluten sensitivity: How its gut immune activation and potential dietary management differ from celiac disease. Mol. Nutr. Food Res. 2018, 62, 1–7. [Google Scholar] [CrossRef]
- Carroccio, A.; D’Alcamo, A.; Cavataio, F.; Soresi, M.; Seidita, A.; Sciumè, C.; Geraci, G.; Iacono, G.; Mansueto, P. High proportions of people with Nonceliac wheat sensitivity have autoimmune disease or antinuclear antibodies. Gastroenterology 2015, 149, 596–603. [Google Scholar] [CrossRef]
- Junker, Y.; Zeissig, S.; Kim, S.J.; Barisani, D.; Wieser, H.; Leffler, D.A.; Zevallos, V.; Libermann, T.A.; Dillon, S.; Freitag, T.L.; et al. Wheat amylase trypsin inhibitors drive intestinal inflammation via activation of toll-like receptor 4. J. Exp. Med. 2012, 209, 2395–2408. [Google Scholar] [CrossRef]
- Losurdo, G.; Principi, M.; Iannone, A.; Amoruso, A.; Ierardi, E.; DiLeo, A.; Barone, M. Extra-intestinal manifestations of non-celiac gluten sensitivity: An expanding paradigm. World J. Gastroenterol. 2018, 24, 1521–1530. [Google Scholar] [CrossRef]
- Kahaly, G.J.; Frommer, L.; Schuppan, D. Celiac disease and glandular autoimmunity. Nutrients 2018, 10, 814. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Laszkowska, M.; Sundström, J.; Lebwohl, B.; Green, P.H.R.; Kämpe, O.; Ludvigsson, J.F. Prevalence of celiac disease in patients with autoimmune thyroid disease: A meta-analysis. Thyroid 2016, 26, 880–890. [Google Scholar] [CrossRef] [PubMed]
- Losurdo, G.; Piscitelli, D.; Giangaspero, A.; Principi, M.; Buffelli, F.; Giorgio, F.; Montenegro, L.; Sorrentino, C.; Amoruso, A.; Ierardi, E.; et al. Evolution of nonspecific duodenal lymphocytosis over 2 years of follow-up. World J. Gastroenterol. 2015, 21, 7545–7552. [Google Scholar] [CrossRef] [PubMed]
- Volta, U.; Bardella, M.T.; Calabrò, A.; Troncone, R.; Corazza, G.R.; Bagnato, C.; Belcari, C.; Bellantoni, A.; Caio, G.; Calella, F.; et al. An italian prospective multicenter survey on patients suspected of having non-celiac gluten sensitivity. BMC Med. 2014, 12, 85. [Google Scholar] [CrossRef]
- Mormile, R. Celiac disease and Hashimoto’s thyroiditis: A shared plot? Int. J. Color. Dis. 2016, 31, 947. [Google Scholar] [CrossRef]
- Pyzik, A.; Grywalska, E.; Matyjaszek-Matuszek, B.; Roliński, J. Immune disorders in Hashimoto’s thyroiditis: What do we know so far? J. Immunol. Res. 2015, 2015, 1–8. [Google Scholar] [CrossRef]
- Gianoukakis, A.G.; Khadavi, N.; Smith, T.J. Cytokines, graves’ disease, and thyroid-associated ophthalmopathy. Thyroid 2008, 18, 953–958. [Google Scholar] [CrossRef]
- Toscano, V.; Conti, F.G.; Anastasi, E.; Mariani, P.; Tiberti, C.; Poggi, M.; Montuori, M.; Monti, S.; Laureti, S.; Cipolletta, E.; et al. Importance of gluten in the induction of endocrine autoantibodies and organ dysfunction in adolescent celiac patients. Am. J. Gastroenterol. 2000, 95, 1742–1748. [Google Scholar] [CrossRef]
- Liontiris Michael, E.M. A concise review of Hashimoto thyroiditis (HT) and the importance of iodine, selenium, vitamin D and gluten on the autoimmunity and dietary management of HT patients.Points that need more investigation. Hell. J. Nucl. Med. 2017, 20, 51–56. [Google Scholar]
- Stazi, A.V.; Trinti, B. Selenium status and over-expression of interleukin-15 in celiac disease and autoimmune thyroid diseases. Ann. Ist. Super. Sanita. 2010, 46, 389–399. [Google Scholar]
- Freeman, H.J. Iron deficiency anemia in celiac disease. World J. Gastroenterol. 2015, 21, 9233–9238. [Google Scholar] [CrossRef] [PubMed]
- Zingone, F.; Ciacci, C. The value and significance of 25(OH) and 1,25(OH) vitamin D serum levels in adult coeliac patients: A review of the literature. Dig. Liver Dis. 2018, 50, 757–760. [Google Scholar] [CrossRef] [PubMed]
- Fasano, A.; Shea-Donohue, T. Mechanisms of disease: The role of intestinal barrier function in the pathogenesis of gastrointestinal autoimmune diseases. Nat. Clin. Pract. Gastroenterol. Hepatol. 2005, 2, 416–422. [Google Scholar] [CrossRef]
- Cereijido, M.; Contreras, R.G.; Flores-Benítez, D.; Flores-Maldonado, C.; Larre, I.; Ruiz, A.; Shoshani, L. New diseases derived or associated with the tight junction. Arch. Med. Res. 2007, 38, 465–478. [Google Scholar] [CrossRef]
- Fasano, A. Zonulin, regulation of tight junctions, and autoimmune diseases. Ann. N. Y. Acad Sci. 2012, 1258, 25–33. [Google Scholar] [CrossRef]
- Paterson, B.M.; Lammers, K.M.; Arrieta, M.C.; Fasano, A.; Meddings, J.B. The safety, tolerance, pharmacokinetic and pharmacodynamic effects of single doses of AT-1001 in coeliac disease subjects: A proof of concept study. Aliment. Pharmacol. Ther. 2007, 26, 757–766. [Google Scholar] [CrossRef] [PubMed]
- König, J.; Wells, J.; Cani, P.D.; García-Ródenas, C.L.; MacDonald, T.; Mercenier, A.; Whyte, J.; Troost, F.; Brummer, R.J. Human intestinal barrier function in health and disease. Clin. Transl. Gastroenterol. 2016, 7, e196. [Google Scholar] [CrossRef]
- Mu, Q.; Kirby, J.; Reilly, C.M.; Luo, X.M. Leaky gut as a danger signal for autoimmune diseases. Front. Immunol. 2017, 8, 1–10. [Google Scholar] [CrossRef]
- Achamrah, N.; Déchelotte, P.; Coëffier, M. Glutamine and the regulation of intestinal permeability: From bench to bedside. Curr. Opin. Clin. Nutr. Metab. Care 2017, 20, 86–91. [Google Scholar] [CrossRef]
- Yang, G.; Bibi, S.; Du, M.; Suzuki, T.; Zhu, M.-J. Regulation of the intestinal tight junction by natural polyphenols: A mechanistic perspective. Crit. Rev. Food Sci. Nutr. 2017, 57, 3830–3839. [Google Scholar] [CrossRef]
- Matthias, T.; Lerner, A. GUT-the trojan horse in remote organs’ autoimmunity. J. Clin. Cell. Immunol. 2016, 7, 1–10. [Google Scholar] [CrossRef]
- Kong, J.; Zhang, Z.; Musch, M.W.; Ning, G.; Sun, J.; Hart, J.; Bissonnette, M.; Yan, C.L. Novel role of the vitamin D receptor in maintaining the integrity of the intestinal mucosal barrier. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 294, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Bruno, R.; Giannasio, P.; Ronga, G.; Baudin, E.; Travagli, J.P.; Russo, D.; Filetti, S.; Schlumberger, M. Sodium iodide symporter expression and radioiodine distribution in extrathyroidal tissues. J. Endocrinol. Investig. 2004, 27, 1010–1014. [Google Scholar] [CrossRef] [PubMed]
- De La Vieja, A.; Santisteban, P. Role of iodide metabolism in physiology and cancer. Endocr. Relat. Cancer 2018, 25, R225–R245. [Google Scholar] [CrossRef] [PubMed]
- Frank, D.N.; Amand, A.L.S.; Feldman, R.A.; Boedeker, E.C.; Harpaz, N.; Pace, N.R. Molecular-phylogenetic characterization of microbial community imbalances in human inflammatory bowel diseases. Proc. Natl. Acad. Sci. USA 2007, 104, 13780–13785. [Google Scholar] [CrossRef]
- Coultate, T. Food: The Chemistry of Its Components: Edition 6, 6th ed.; The Royal Society of Chemistry: London, UK, 2016. [Google Scholar]
- Pearce, E.N. Iodine deficiency disorders and their elimination. Indian J. Med. Res. 2017, 148, 120. [Google Scholar]
- Zicker, S.; Schoenherr, B. The role of iodine in nutrition and metabolism. Compend. Contin. Educ. Pract. Vet. 2012, 34, 1–4. [Google Scholar]
- Manousou, S.; Carlsson, L.M.S.; Eggertsen, R.; Hulthén, L.; Jacobson, P.; Landin-Wilhelmsen, K.; Trimpou, P.; Svensson, P.A.; Nyström, H.F. Iodine status after bariatric surgery—A prospective 10-year report from the Swedish Obese Subjects (SOS) study. Obes. Surg. 2018, 28, 349–357. [Google Scholar] [CrossRef]
- Meletis, C.D. Iodine: Health implications of deficiency. J. Evid. Based Complement. Altern. Med. 2011, 16, 190–194. [Google Scholar] [CrossRef]
- Zimmermann, M.B.; Galetti, V. Iodine intake as a risk factor for thyroid cancer: A comprehensive review of animal and human studies. Thyroid Res. 2015, 8, 8. [Google Scholar] [CrossRef]
- Luo, J.; Hendryx, M.; Dinh, P.; He, K. Association of iodine and iron with thyroid function. Biol. Trace Element Res. 2017, 179, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Wolff, J.B.; Ciiaikoff, I.L. Plasma inorganic iodide as a homeostatic regulator of thyroid function. J. Biol. Chem. 1948, 174, 555–564. [Google Scholar] [PubMed]
- Leung, A.M.; Braverman, L.E. Consequences of excess iodine. Nat. Rev. Endocrinol. 2014, 10, 136–142. [Google Scholar] [CrossRef]
- Vela, D. Balance of cardiac and systemic hepcidin and its role in heart physiology and pathology. Lab. Investig. 2018, 98, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Chieppa, M.; Giannelli, G. Immune cells and microbiota response to iron starvation. Front. Med. 2018, 5, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Kortman, G.A.M.; Raffatellu, M.; Swinkels, D.W.; Tjalsma, H. Nutritional iron turned inside out: Intestinal stress from a gut microbial perspective. FEMS Microbiol. Rev. 2014, 38, 1202–1234. [Google Scholar] [CrossRef]
- Bullen, J.; Griffiths, E.; Rogers, H.; Ward, G. Sepsis: The critical role of iron. Microbes Infect. 2000, 2, 409–415. [Google Scholar] [CrossRef]
- Weinberg, E.D. The lactobacillus anomaly: Total iron abstinence. Perspect. Biol. Med. 1997, 40, 578–583. [Google Scholar] [CrossRef]
- Constante, M.; Fragoso, G.; Lupien-meilleur, J.; Calvé, A.; Santos, M.M. Iron supplements modulate colon microbiota composition and potentiate the protective effects of probiotics in dextran sodium sulfate-induced colitis. Inflamm. Bowel Dis. 2017, 23, 753–766. [Google Scholar] [CrossRef]
- Wu, W.; Song, Y.; He, C.; Liu, C.; Wu, R.; Fang, L.; Cong, Y.; Miao, Y.; Liu, Z. Divalent metal-ion transporter 1 is decreased in intestinal epithelial cells and contributes to the anemia in inflammatory bowel disease. Sci. Rep. 2015, 5, 1–12. [Google Scholar] [CrossRef]
- Paganini, D.; Zimmermann, M.B. The effects of iron fortification and supplementation on the gut microbiome and diarrhea in infants and children: A review. Am. J. Clin. Nutr. 2017, 106, 1688S–1693S. [Google Scholar] [CrossRef] [PubMed]
- Kawicka, A.; Regulska-Ilow, B. Metabolic disorders and nutritional status in autoimmune thyroid diseases. Postepy Hig. Med. Dosw. 2015, 69, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Rayman, M.P. Multiple nutritional factors and the risk of hashimoto’s thyroiditis. Thyroid 2017, 27, 597–610. [Google Scholar] [CrossRef] [PubMed]
- Velluzzi, F.; Caradonna, A.; Boy, M.F.; Pinna, M.A.; Cabula, R.; Lai, M.A.; Piras, E.; Corda, G.; Mossa, P.; Atzeni, F.; et al. Thyroid and celiac disease: Clinical, serological, and echographic study. Am. J. Gastroenterol. 1998, 93, 976–979. [Google Scholar] [CrossRef]
- Zimmermann, M.B. The influence of iron status on iodine utilization and thyroid function. Annu. Rev. Nutr. 2006, 26, 367–389. [Google Scholar] [CrossRef]
- Beard, J.L.; Brigham, D.E.; Kelley, S.K.; Green, M.H. Plasma thyroid hormone kinetics are altered in iron-deficient rats. J. Nutr. 1998, 128, 1401–1408. [Google Scholar] [CrossRef]
- Zhang, H.; Teng, X.; Shan, Z.; Wang, Z.; Li, C.; Yu, X.; Mao, J.; Wang, W.; Xie, X.; Teng, W. Association between iron deficiency and prevalence of thyroid autoimmunity in pregnant and non-pregnant women of childbearing age: A cross-sectional study. Chin. Med. J. 2019, 132, 2143–2149. [Google Scholar] [CrossRef]
- Yu, X.; Shan, Z.; Li, C.; Mao, J.; Wang, W.; Xie, X.; Liu, A.; Teng, X.; Zhou, W.; Li, C.; et al. Iron deficiency, an independent risk factor for isolated hypothyroxinemia in pregnant and nonpregnant women of childbearing age in China. J. Clin. Endocrinol. Metab. 2015, 100, 1594–1601. [Google Scholar] [CrossRef]
- Khatiwada, S.; Gelal, B.; Baral, N.; Lamsal, M. Association between iron status and thyroid function in Nepalese children. Thyroid Res. 2016, 9, 1–7. [Google Scholar] [CrossRef]
- Eftekhari, M.H.; Simondon, K.B.; Jalali, M.; Keshavarz, S.A.; Elguero, E.; Eshraghian, M.R.; Saadat, N. Effects of administration of iron, iodine and simultaneous iron-plus-iodine on the thyroid hormone profile in iron-deficient adolescent Iranian girls. Eur. J. Clin. Nutr. 2006, 60, 545–552. [Google Scholar] [CrossRef]
- Hess, S.Y.; Zimmermann, M.B.; Adou, P.; Torresani, T.; Hurrell, R.F. Treatment of iron deficiency in goitrous children improves the efficacy of iodized salt in Côte d’Ivoire. Am. J. Clin. Nutr. 2002, 75, 743–748. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zimmermann, M.B.; Wegmueller, R.; Zeder, C.; Chaouki, N.; Rohner, F.; Saïssi, M.; Torresani, T.; Hurrell, R.F. Dual fortification of salt with iodine and micronized ferric pyrophosphate: A randomized, double-blind, controlled trial. Am. J. Clin. Nutr. 2004, 80, 952–959. [Google Scholar] [CrossRef] [PubMed]
- Arora, M. Study of trace elements in patients of hypothyroidism with special reference to zinc and copper. Biomed. J. Sci. Tech. Res. 2018, 6, 11–16. [Google Scholar] [CrossRef]
- Aziz, M.A.; Diab, K.S. Effectiveness of zinc supplementation in regulating serum hormonal and inflammatory status in hypothyroidism patients. Med. J. Babylon 2016, 13, 347–353. [Google Scholar]
- Civitareale, D.; Saiardi, A.; Falasca, P. Purification and characterization of thyroid transcription factor 2. Biochem. J. 1994, 304, 981–985. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.; Zhao, Z.; Wang, Y. Preparation of selenium/zinc-enriched probiotics and their effect on blood selenium and zinc concentrations, antioxidant capacities, and intestinal microflora in canine. Biol. Trace Element Res. 2011, 141, 170–183. [Google Scholar] [CrossRef] [PubMed]
- Morley, E. Zinc deficiency, chronic hypothalamic-pituitary-thyroid and. Am. J. Clin. Nutr. 1980, 33, 1767–1770. [Google Scholar] [CrossRef]
- Pekary, A.E.; Bhasin, S.; Smith, V.; Sugawara, M.; Swerdloff, R.S.; Hershman, J.M. Thyroid hormone modulation of thyrotrophin-releasing hormone (TRH) and TRH-Gly levels in the male rat reproductive system. J. Endocrinol. 1987, 114, 271–277. [Google Scholar] [CrossRef]
- Anselmet, A.; Bismuth, J.; Torresani, J. Triiodothyronine nuclear receptor: Role of histones and dna in hormone binding. Biochim. Biophys. Acta 1983, 739, 291–300. [Google Scholar] [CrossRef]
- Baltaci, A.K.; Mogulkoc, R.; Belviranli, M. Serum levels of calcium, selenium, magnesium, phosphorus, chromium, copper and iron–their relation to zinc in rats with induced. Acta Clin. Croat. 2013, 52, 151–156. [Google Scholar]
- Yoshida, K.; Kiso, Y.; Watanabe, T.; Kaise, K.; Kaise, N.; Itagaki, Y.; Yamamoto, M.; Sakurada, T.; Yoshinaga, K. Erythrocyte zinc in hyperthyroidism: Reflection of integrated levels over the previous few months. Metabolism 1990, 39, 182–186. [Google Scholar] [CrossRef]
- Kralik, A.; Eder, K.; Kirchgessner, M. Influence of zinc and selenium deficiency on parameters relating to thyroid hormone metabolism. Horm. Metab. Res. 1996, 28, 223–226. [Google Scholar] [CrossRef] [PubMed]
- Regina, T. Effect of nutrients and dietary substances on thyroid function and hypothyroidism. Demetra 2016, 11, 427–444. [Google Scholar]
- Drutel, A.; Caron, P. Selenium and the thyroid gland: More good news for clinicians. Clin. Endocrinol. 2013, 78, 155–164. [Google Scholar] [CrossRef]
- Combs, G.F. Selenium in global food systems. Br. J. Nutr. 2001, 85, 517–547. [Google Scholar] [CrossRef]
- Burk, R.F.; Hill, K.E. Regulation of selenium metabolism and transport. Annu. Rev. Nutr. 2015, 35, 109–134. [Google Scholar] [CrossRef]
- Kasaikina, M.V.; Kravtsova, M.A.; Lee, B.C.; Seravalli, J.; Peterson, D.A.; Walter, J.; Legge, R.; Benson, A.K.; Hatfield, D.L.; Gladyshev, V.N. Dietary selenium affects host selenoproteome expression by influencing the gut microbiota. FASEB J. 2011, 25, 2492–2499. [Google Scholar] [CrossRef]
- Pizzulli, A.; Ranjbar, A. Selenium deficiency and hypothyroidism a new etiology in the differential diagnosis of hypothyroidism in children. Biol. Trace Element Res. 2000, 77, 199–208. [Google Scholar] [CrossRef]
- Hewison, M.; Burke, F.; Evans, K.N.; Lammas, D.A.; Sansom, D.M.; Liu, P.; Modlin, R.L.; Adams, J.S. Extra-renal 25-hydroxyvitamin D3-1a-hydroxylase in human health and disease. J. Steroid Biochem. Mol. Biol. 2007, 103, 316–321. [Google Scholar] [CrossRef]
- Hossein-Nezhad, A.; Holick, M.F. Vitamin D for health: A global perspective. Mayo Clin. Proc. 2013, 88, 720–755. [Google Scholar] [CrossRef]
- Barragan, M.; Good, M.; Kolls, J.K. Regulation of dendritic cell function by vitamin D. Nutrients 2015, 7, 8127–8151. [Google Scholar] [CrossRef]
- Mackawy, A.M.; Al-Ayed, B.M.; Al-Rashidi, B.M. Vitamin D deficiency and its association with thyroid disease. Int. J. Health Sci. 2013, 7, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Unal, A.D.; Tarcin, O.; Parildar, H.; Cigerli, O.; Eroglu, H.; Demirag, N.G. Vitamin D deficiency is related to thyroid antibodies in autoimmune thyroiditis. Central Eur. J. Immunol. 2014, 39, 493–497. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.Y.; Kim, K.J.; Kim, D.; Hwang, S.; Lee, E.J. Low serum vitamin D is associated with anti-thyroid peroxidase antibody in autoimmune thyroiditis. Yonsei Med. J. 2014, 55, 476–481. [Google Scholar] [CrossRef]
- Aurizio, F.D.; Villalta, D.; Metus, P.; Doretto, P.; Tozzoli, R. Vitamin D is a player or not in the pathophysiology of autoimmune thyroid diseases? Autoimmun. Rev. 2014, 25, 1–7. [Google Scholar]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document: The international scientific association for probiotics and prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Sohail, M.U.; Ijaz, A.; Yousaf, M.S.; Ashraf, K.; Zaneb, H.; Aleem, M.; Rehman, H. Alleviation of cyclic heat stress in broilers by dietary supplementation of mannan-oligosaccharide and lactobacillus-based probiotic: Dynamics of cortisol, thyroid hormones, cholesterol, C-reactive protein, and humoral immunity. Poult. Sci. 2010, 89, 1934–1938. [Google Scholar] [CrossRef]
- Chotinsky, D.; Mihaylov, R. Effect of probiotics and avotan on the level of thyroid hormones in the blood plasma of broiler chickens. Bulg. J. Agric. Sci. 2013, 19, 817–821. [Google Scholar]
- Talebi, S.; Karimifar, M.; Heidari, Z.; Mohammadi, H.; Askari, G. The effects of synbiotic supplementation on thyroid function and inflammation in hypothyroid patients: A randomized, double-blind, placebo-controlled trial. Complement. Ther. Med. 2020, 48, 102234. [Google Scholar] [CrossRef]
- Zhou, J.S.; Gill, H.S. Immunostimulatory probiotic Lactobacillus rhamnosus HN001 and Bifidobacterium lactis HN019 do not induce pathological inflammation in mouse model of experimental autoimmune thyroiditis. Int. J. Food Microbiol. 2005, 103, 97–104. [Google Scholar] [CrossRef]
- Spaggiari, G.; Brigante, G.; de Vincentis, S.; Cattini, U.; Roli, L.; de Santis, M.C.; Baraldi, E.; Tagliavini, S.; Varani, M.; Trenti, T.; et al. Probiotics ingestion does not directly affect thyroid hormonal parameters in hypothyroid patients on levothyroxine treatment. Front. Endocinol. 2017, 8, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Salvatore, G. Es hormones thyroidlennes et son mkcanisme ’. Neral Comp. Endocrinol. 1963, 3, 15–25. [Google Scholar] [CrossRef]
- Laurberg, P.; Knudsen, N.; Andersen, S.; Carlé, A.; Pedersen, I.B.; Karmisholt, J. Thyroid function and obesity. Eur. Thyroid J. 2012, 1, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Rotondi, M.; Leporati, P.; Rizza, M.I.; Clerici, A.; Groppelli, G.; Pallavicini, C.; La Manna, A.; Fonte, R.; Magri, F.; Biondi, B.; et al. Raised serum TSH in morbid-obese and non-obese patients: Effect on the circulating lipid profile. Endocrine 2014, 45, 92–97. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kok, P.; Roelfsema, F.; Langendonk, J.G.; Frölich, M.; Burggraaf, J.; Meinders, A.E.; Pijl, H. High circulating thyrotropin levels in obese women are reduced after body weight loss induced by caloric restriction. J. Clin. Endocrinol. Metab. 2005, 90, 4659–4663. [Google Scholar] [CrossRef]
- Guan, B.; Chen, Y.; Yang, J.; Yang, W.; Wang, C. Effect of bariatric surgery on thyroid function in obese patients: A systematic review and meta-analysis. Obes. Surg. 2017, 27, 3292–3305. [Google Scholar] [CrossRef]
- Duran, İ.D.; Gülçelik, N.E.; Bulut, B.; Balcı, Z.; Berker, D.; Güler, S. Differences in calcium metabolism and thyroid physiology after sleeve gastrectomy and Roux-En-Y gastric bypass. Obes. Surg. 2019, 29, 705–712. [Google Scholar] [CrossRef]
- Fazylov, R.; Soto, E.; Cohen, S.; Merola, S. Laparoscopic Roux-en-Y gastric bypass surgery on morbidly obese patients with hypothyroidism. Obes. Surg. 2008, 18, 644–647. [Google Scholar] [CrossRef]
- Janssen, I.M.C.; Homan, J.; Schijns, W.; Betzel, B.; Aarts, E.O.; Berends, F.J.; De Boer, H. Subclinical hypothyroidism and its relation to obesity in patients before and after Roux-en-Y gastric bypass. Surg. Obes. Relat. Dis. 2015, 11, 1257–1263. [Google Scholar] [CrossRef]
- Yu, H.; Li, Q.; Zhang, M.; Liu, F.; Pan, J.; Tu, Y.; Lu, J.; Zhang, P.; Han, J.; Jia, W.; et al. Decreased leptin is associated with alterations in thyroid-stimulating hormone levels after roux-en-y gastric bypass surgery in obese euthyroid patients with type 2 diabetes. Obes. Facts 2019, 12, 272–280. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, F.; Zhao, C.; Xu, Q.; Liang, C.; Yang, Y.; Wang, H.; Shang, Y.; Wang, Y.; Mu, X.; et al. Dysbiosis of the gut microbiome is associated with thyroid cancer and thyroid nodules and correlated with clinical index of thyroid function. Endocrine 2019, 64, 564–574. [Google Scholar] [CrossRef] [PubMed]
- Zitvogel, L.; Ayyoub, M.; Routy, B.; Kroemer, G. Microbiome and anticancer immunosurveillance. Cell 2016, 165, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Rajagopala, S.V.; Vashee, S.; Oldfield, L.M.; Suzuki, Y.; Venter, J.C.; Telenti, A.; Nelson, K.E. The human microbiome and cancer. Cancer Prev. Res. 2017, 10, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Meng, S.; Chen, B.; Yang, J.; Wang, J.; Zhu, D.; Meng, Q.; Zhang, L. Study of microbiomes in aseptically collected samples of human breast tissue using needle biopsy and the potential role of in situ tissue microbiomes for promoting malignancy. Front. Oncol. 2018, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Trapani, K.M.; Boghossian, L.J.; Caskey, E. Clostridium subterminale Septicemia in a Patient with Metastatic Gastrointestinal Adenocarcinoma. Case Rep. Infect. Dis. 2018, 2018, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Dahmus, J.D.; Kotler, D.L.; Kastenberg, D.M.; Kistler, C.A. The gut microbiome and colorectal cancer: A review of bacterial pathogenesis. J. Gastrointest. Oncol. 2018, 9, 769–777. [Google Scholar] [CrossRef]
- Benitez, A.J.; Hoffmann, C.; Muir, A.B.; Dods, K.K.; Spergel, J.M.; Bushman, F.D.; Wang, M.-L. Inflammation-associated microbiota in pediatric eosinophilic esophagitis. Microbiome 2015, 3, 23. [Google Scholar] [CrossRef]
- Memba, R.; Duggan, S.N.; Ni Chonchubhair, H.M.; Griffin, O.M.; Bashir, Y.; O’Connor, D.B.; Murphy, A.; McMahon, J.; Volcov, Y.; Ryan, B.M.; et al. The potential role of gut microbiota in pancreatic disease: A systematic review. Pancreatology 2017, 17, 867–874. [Google Scholar] [CrossRef]
- Soriano, O.; Delgado, G.; Anguiano, B.; Petrosyan, P.; Molina-Servín, E.D.; Gonsebatt, M.E.; Aceves, C. Antineoplastic effect of iodine and iodide in dimethylbenz[a]anthracene- induced mammary tumors: Association between lactoperoxidase and estrogen-adduct production. Endocr. Relat. Cancer 2011, 18, 529–539. [Google Scholar] [CrossRef]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Knezevic, J.; Starchl, C.; Tmava Berisha, A.; Amrein, K. Thyroid-Gut-Axis: How Does the Microbiota Influence Thyroid Function? Nutrients 2020, 12, 1769. https://doi.org/10.3390/nu12061769
Knezevic J, Starchl C, Tmava Berisha A, Amrein K. Thyroid-Gut-Axis: How Does the Microbiota Influence Thyroid Function? Nutrients. 2020; 12(6):1769. https://doi.org/10.3390/nu12061769
Chicago/Turabian StyleKnezevic, Jovana, Christina Starchl, Adelina Tmava Berisha, and Karin Amrein. 2020. "Thyroid-Gut-Axis: How Does the Microbiota Influence Thyroid Function?" Nutrients 12, no. 6: 1769. https://doi.org/10.3390/nu12061769
APA StyleKnezevic, J., Starchl, C., Tmava Berisha, A., & Amrein, K. (2020). Thyroid-Gut-Axis: How Does the Microbiota Influence Thyroid Function? Nutrients, 12(6), 1769. https://doi.org/10.3390/nu12061769