Anti-Pathogenic Functions of Non-Digestible Oligosaccharides In Vitro

 ,
,  ,
,
Abstract
1. Introduction
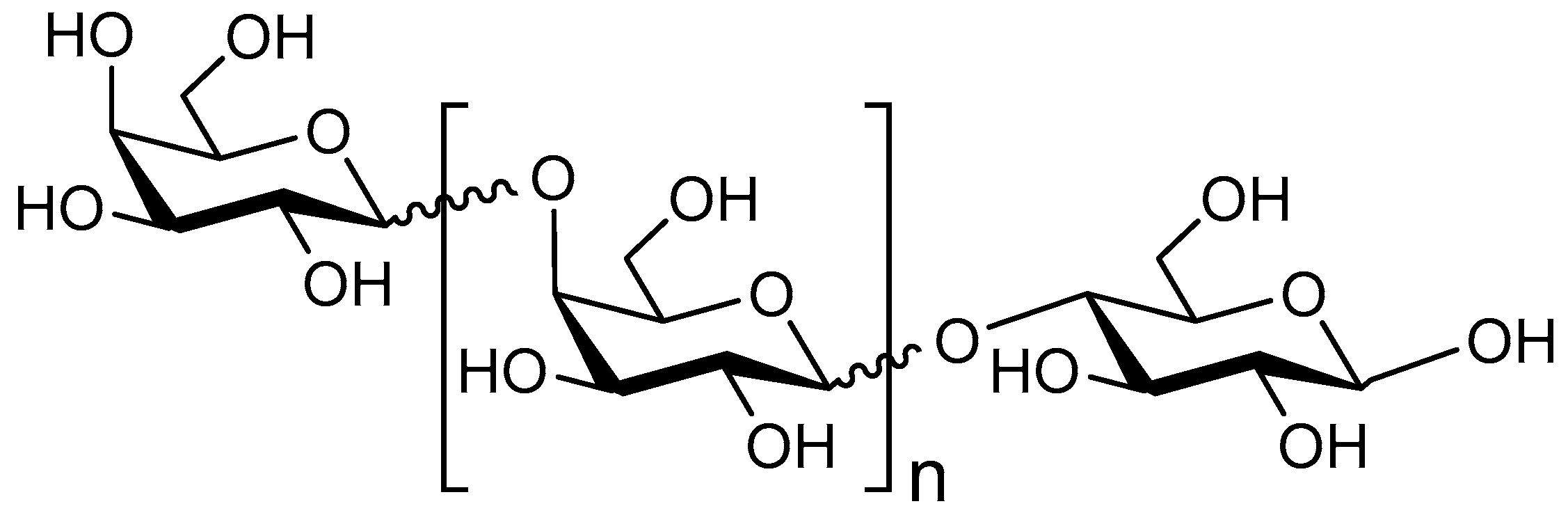
2. Human Milk Oligosaccharides
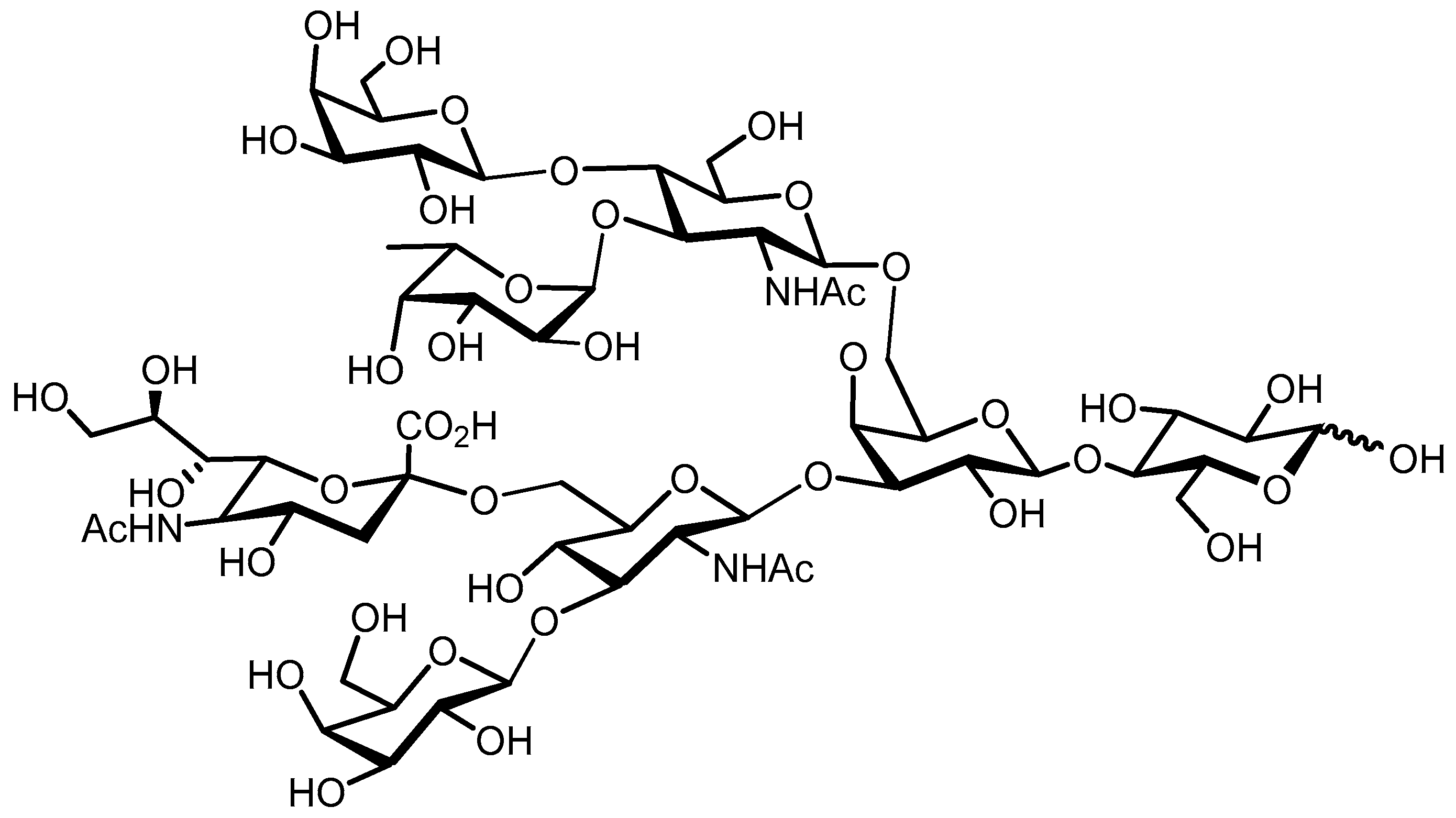
2.1. Structure
2.2. Anti-Pathogenic Functionalities HMO Mixtures
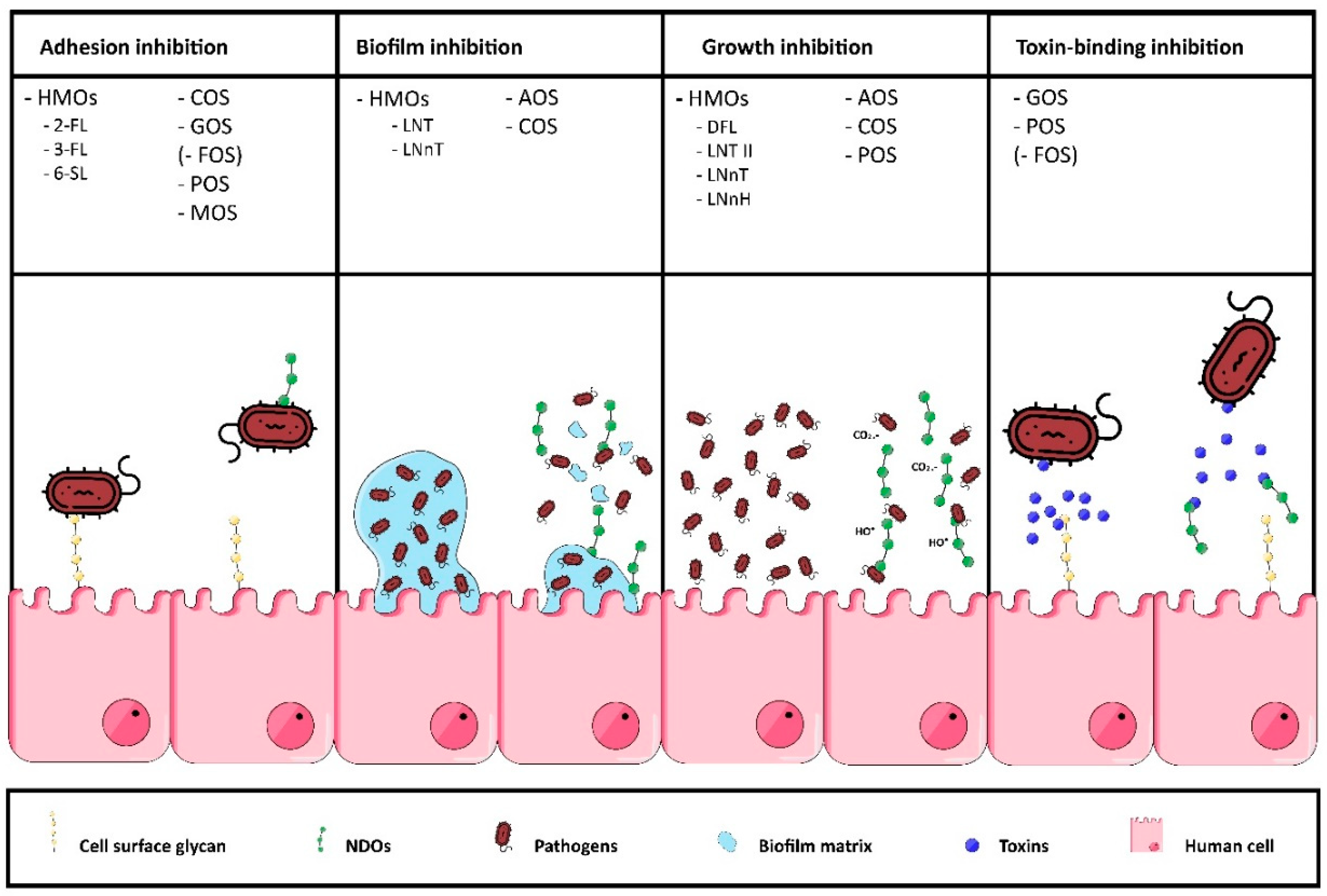
2.2.1. Anti-Adhesion HMO Mixtures
Neutral HMO Fraction—Fucosylated and Non-Fucosylated
Acidic HMO Fraction
2.2.2. Other Anti-Pathogenic Mechanisms of HMO Mixtures
2.3. Anti-Pathogenic Functionalities Isolated HMO Structures
2.3.1. Neutral Isolated HMO Structures
2.3.2. Acidic Isolated HMO Structures
2.3.3. Fucosylated Oligosaccharides (FO)
3. Alginate Oligosaccharides
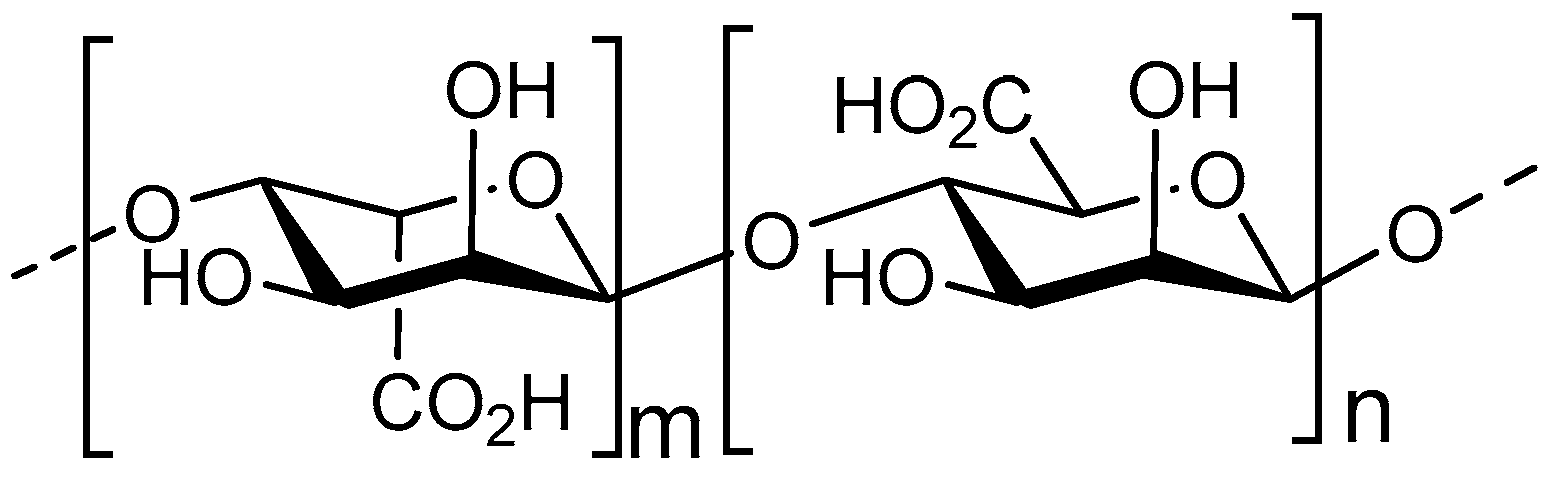
3.1. Structure
3.2. Anti-Pathogenic Functionalities
3.2.1. Biofilm Inhibition
3.2.2. Metal ion Scavenging
3.2.3. Quorum Sensing (QS) System Inhibition
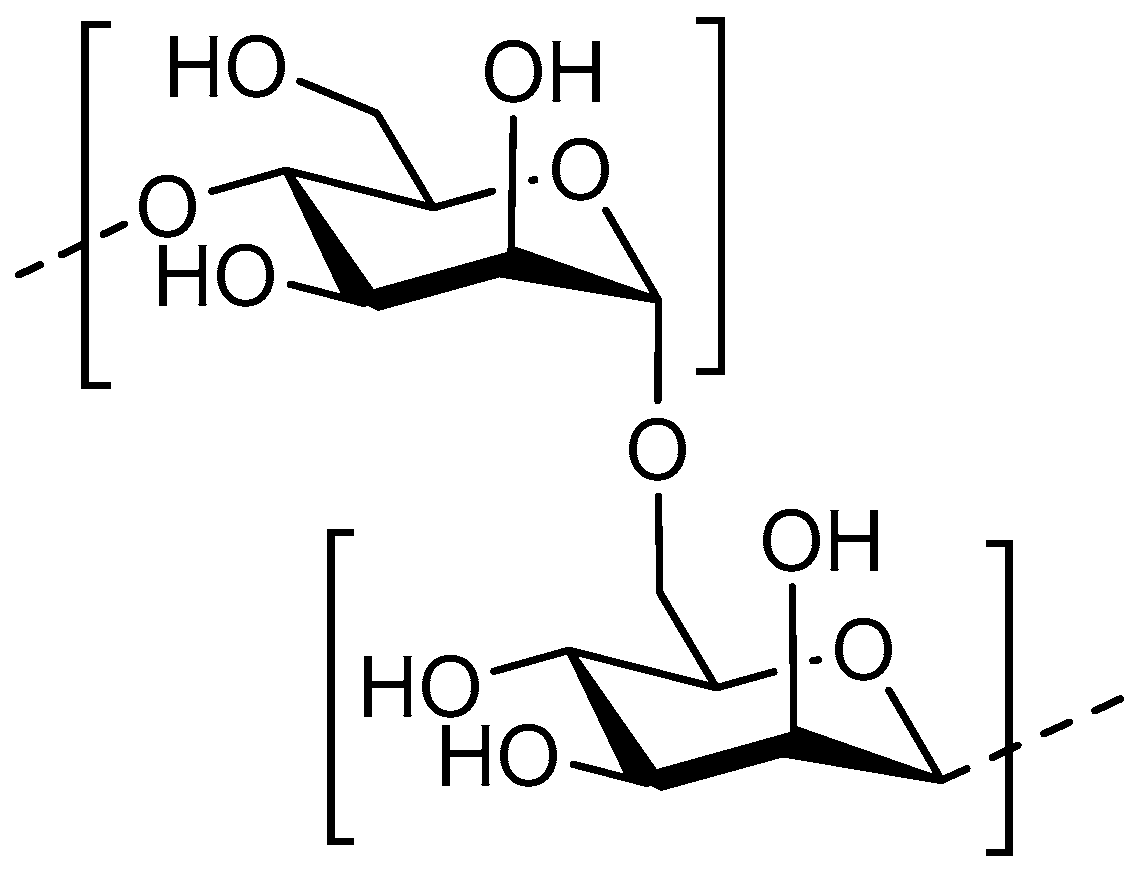
4. Chito-Oligosaccharides/Chitosan Oligosaccharides
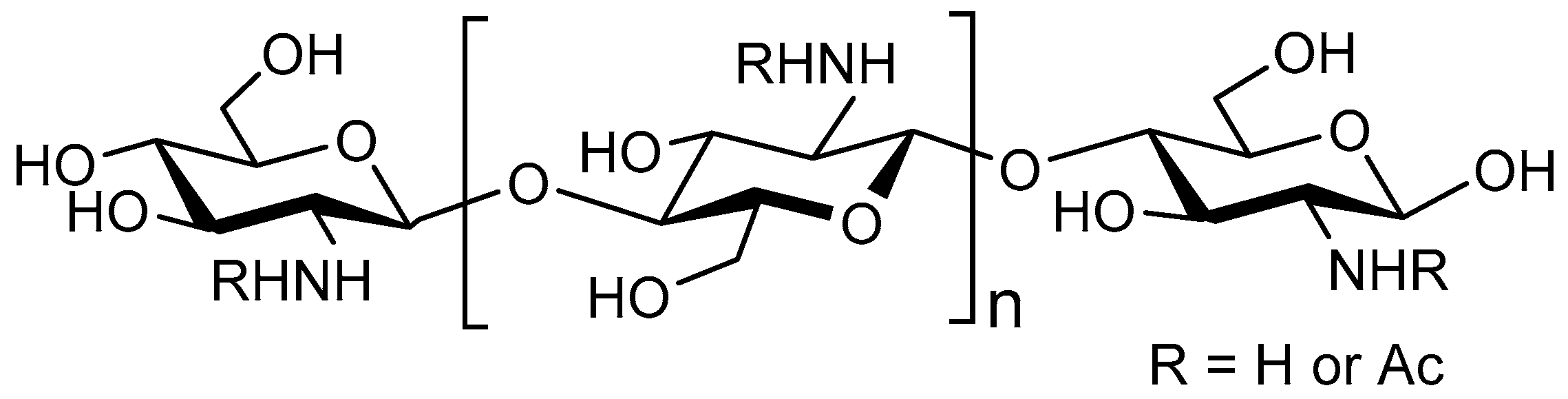
4.1. Structure
4.2. Anti-Pathogenic Functionalities
4.2.1. Cell Membrane Disruption
4.2.2. Adhesion Inhibition
4.2.3. Association with Bacterial DNA
4.2.4. Synergy with Antibiotic Treatment
5. Fructo-Oligosaccharides
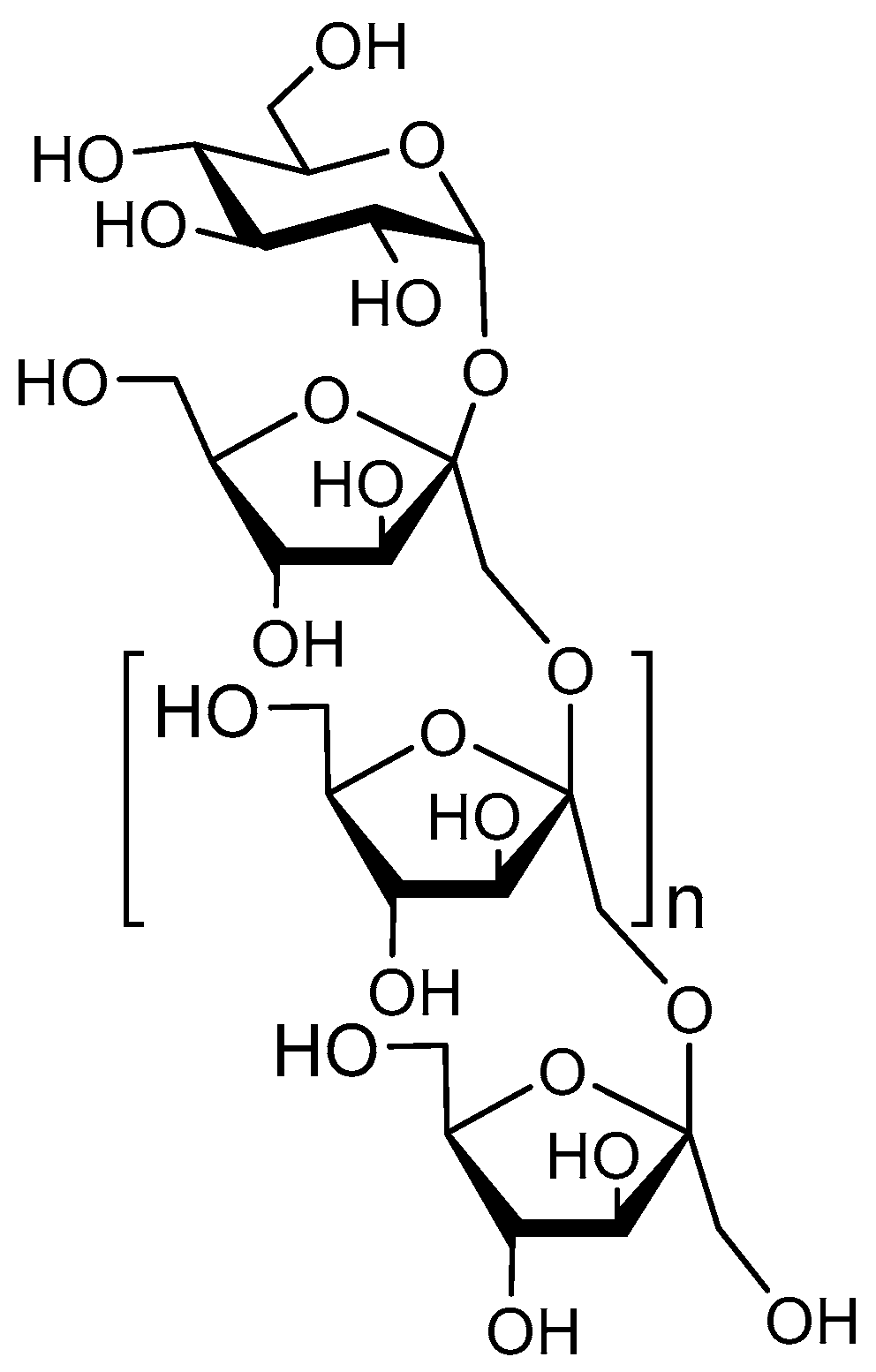
5.1. Structure
5.2. Anti-Pathogenic Functionalities
6. Galacto-Oligosaccharides
6.1. Structure
6.2. Anti-Pathogenic Functionalities
6.2.1. Adhesion Inhibition
6.2.2. Anti-Toxin Binding
7. Mannan-Oligosaccharides
7.1. Structure
7.2. Anti-Pathogenic Functionalities
Adhesion Inhibition
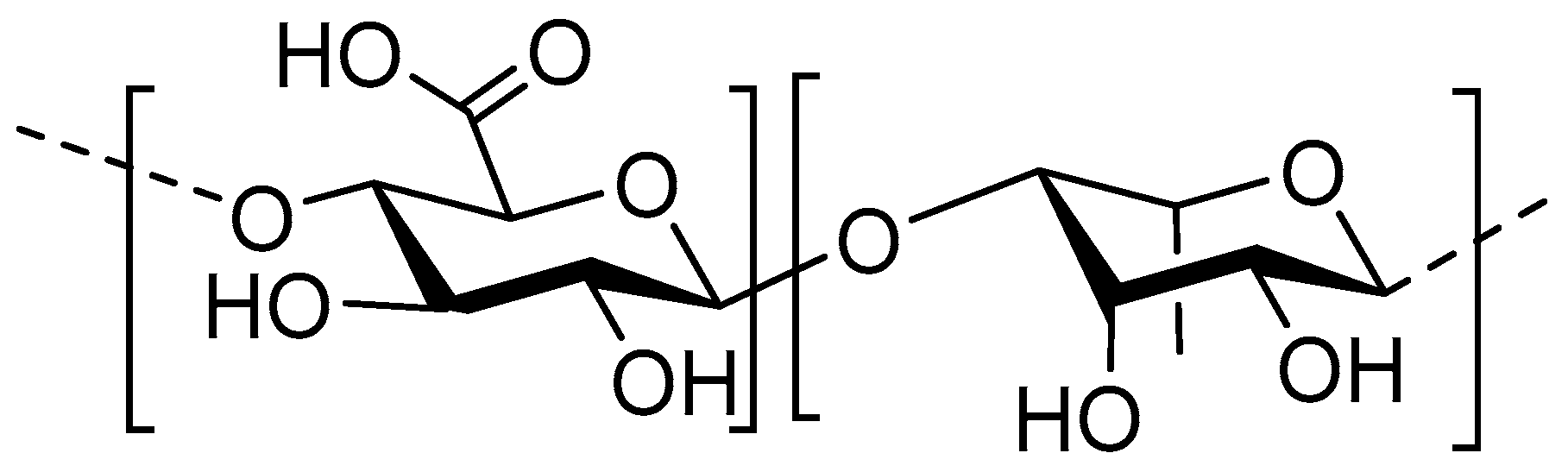
8. Pectic Oligosaccharides
8.1. Structure
8.2. Anti-Pathogenic Functionalities
8.2.1. Growth Inhibition
8.2.2. CO2 Radical Production
8.2.3. Adhesion Inhibition
8.2.4. Inhibition of Toxin-Binding
9. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 2-FL | 2-Fucosyllactose |
| 3-FL | 3-Fucosyllactose |
| 3-SL | 3-Sialyllactose |
| 6-SL | 6-Sialyllactose |
| α1,2-fucose | FUT2 |
| α1,4-fucose | FUT3 |
| AHL | Acyl homoserine lactones |
| AOS | Alginate oligosaccharide |
| AP | Acetylation pattern |
| COS | Chito-oligosaccharides/chitosan oligosaccharides |
| Ctx | Vibrio cholerae toxin |
| DA | Degree of acetylation |
| DD | Degree of deacetylation |
| DP | Degree of polymerization |
| EPEC | Enteropathogenic Escherichia coli |
| EPS | Extracellular polymeric substances |
| FO | Fucosylated oligosaccharides |
| FOS | Fructo-oligosaccharides |
| Gal | Galactose |
| GBS | Group B Streptococcus |
| Gb3 | Globotriaosylceramide |
| GC | Guluronic content |
| Glc | Glucose |
| GlcN | β-1,4-linked d-glucosamine |
| GlcNAc | N-acetylglucosamine |
| GOS | Galacto-oligosaccharides |
| HMOs | Human milk oligosaccharides |
| HMWF | High-molecular weight fraction |
| LMWF | Low-molecular weight fraction |
| LNFP I | Lacto-N-fucopentaose I |
| LNT | Lacto-N-tetraose |
| LSTa | LS-tetrasaccharide a |
| MOS | Mannan-oligosaccharides |
| NDOs | Non-digestible oligosaccharides |
| POS | Pectic oligosaccharides |
| QS | Quorum-Sensing |
| Stx | Shiga toxin |
| UPEC | UroPathogenic Escherichia coli |
| VTEC | Verocytotoxin-producing Escherichia coli |
References
- Mussatto, S.I.; Mancilha, I.M. Non-digestible oligosaccharides: A review. Carbohydr. Polym. 2007, 68, 587–597. [Google Scholar] [CrossRef]
- Akbari, P.; Fink-Gremmels, J.; Willems, R.H.A.M.; Difilippo, E.; Schols, H.A.; Schoterman, M.H.C.; Garssen, J.; Braber, S. Characterizing microbiota-independent effects of oligosaccharides on intestinal epithelial cells: Insight into the role of structure and size: Structure–activity relationships of non-digestible oligosaccharides. Eur. J. Nutr. 2017, 56, 1919–1930. [Google Scholar] [CrossRef] [PubMed]
- Schouten, B.; van Esch, B.C.A.M.; Kormelink, T.G.; Moro, G.E.; Arslanoglu, S.; Boehm, G.; Knippels, L.M.J.; Redegeld, F.A.; Willemsen, L.E.M.; Garssen, J. Non-digestible oligosaccharides reduce immunoglobulin free light-chain concentrations in infants at risk for allergy. Pediatr. Allergy Immunol. 2011, 22, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Algieri, F.; Rodríguez-Nogales, A.; Garrido-Mesa, N.; Vezza, T.; Garrido-Mesa, J.; Utrilla, M.P.; Montilla, A.; Cardelle-Cobas, A.; Olano, A.; Corzo, N.; et al. Intestinal anti-inflammatory effects of oligosaccharides derived from lactulose in the trinitrobenzenesulfonic acid model of rat colitis. J. Agric. Food Chem. 2014, 62, 4285–4297. [Google Scholar] [CrossRef] [PubMed]
- Okeke, I.N.; Aboderin, O.A.; Byarugaba, D.K.; Ojo, K.K.; Opintan, J.A. Growing problem of multidrug-resistant enteric pathogens in Africa. Emerg. Infect. Dis. 2007, 13, 1640–1646. [Google Scholar] [CrossRef]
- Humphries, R.M.; Schuetz, A.N. Antimicrobial susceptibility testing of bacteria that cause gastroenteritis. Clin. Lab. Med. 2015, 35, 313–331. [Google Scholar] [CrossRef]
- Willyard, C. The drug-resistant bacteria that pose the greatest health threats. Nature 2017, 543, 15. [Google Scholar] [CrossRef]
- Flemming, H.-C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Qiang, X.; YongLie, C.; QianBing, W. Health benefit application of functional oligosaccharides. Carbohydr. Polym. 2009, 77, 435–441. [Google Scholar] [CrossRef]
- Cai, Y.; Folkerts, G.; Maurer, M. Microbiota—Dependent and—Independent effects of dietary fibre on human health. Br. J. Pharmacol. 2020, 177, 1363–1381. [Google Scholar] [CrossRef]
- Haak, B.W.; Prescott, H.C.; Wiersinga, W.J. Therapeutic Potential of the Gut Microbiota in the Prevention and Treatment of Sepsis. Front. Imunnol. 2018, 9, 1–8. [Google Scholar] [CrossRef] [PubMed]
- 1Vitetta, L.; Vitetta, G.; Hall, S. Immunological Tolerance and Function: Associations Between Intestinal Bacteria. Probiotics 2018, 9, 1–15. [Google Scholar]
- Bernardi, A.; Jiménez-barbero, J.; Casnati, A.; Castro, C. De Multivalent glycoconjugates as anti-pathogenic agents. Chem. Soc. Rev. 2015, 42, 4709–4727. [Google Scholar] [CrossRef] [PubMed]
- Shoaf-Sweeney, K.D.; Hutkins, R.W. Chapter 2 Adherence, Anti-Adherence, and Oligosaccharides. Preventing Pathogens from Sticking to the Host. Adv. Food Nutr. Res. 2008, 55, 101–161. [Google Scholar]
- Rosenberg, M.; Bayer, E.A.; Delarea, J.; Rosenberg, E. Role of Thin Fimbriae in Adherence and Growth of Acinetobacter calcoaceticus RAG-1 on Hexadecane. Appl. Environ. Microbiol. 1982, 44, 929–937. [Google Scholar] [CrossRef]
- Duguid, J.P.; Old, D.C. Adhesive Properties of Enterobacteriaceae BT. In Bacterial Adherence; Beachey, E.H., Ed.; Springer: Dordrecht, The Netherladns, 1980; pp. 185–217. ISBN 978-94-009-5863-0. [Google Scholar]
- Coppa, G.V.; Zampini, L.; Galeazzi, T.; Facinelli, B.; Ferrante, L.; Capretti, R.; Orazio, G. Human Milk Oligosaccharides Inhibit the Adhesion to Caco-2 Cells of Diarrheal Pathogens: Escherichia coli, Vibrio cholerae, and Salmonella fyris. Pediatr. Res. 2006, 59. [Google Scholar] [CrossRef]
- Newburg, D.S. Do the binding properties of oligosaccharides in milk protect human infants from gastrointestinal bacteria? J. Nutr. 1997, 127, 980S–984S. [Google Scholar] [CrossRef]
- Lin, A.E.; Autran, C.A.; Szyszka, A.; Escajadillo, T.; Huang, M.; Godula, K.; Prudden, A.R.; Boons, G.; Lewis, A.L.; Doran, K.S.; et al. Human milk oligosaccharides inhibit growth of group B. J. Biol. Chem. 2017, 292, 11243–11249. [Google Scholar] [CrossRef]
- Andreas, N.J.; Al-Khalidi, A.; Jaiteh, M.; Clarke, E.; Hyde, M.J.; Modi, N.; Holmes, E.; Kampmann, B.; Mehring Le Doare, K. Role of human milk oligosaccharides in Group B Streptococcus colonisation. Clin. Transl. Immunol. 2016, 5, e99. [Google Scholar] [CrossRef]
- Ackerman, D.L.; Doster, R.S.; Weitkamp, J.; Aronoff, D.M.; Gaddy, J.A.; Townsend, S.D. Human Milk Oligosaccharides Exhibit Antimicrobial and Antibio film Properties against Group B Streptococcus. ACS Infect. Dis. 2017. [Google Scholar] [CrossRef]
- Patel, S.; Goyal, A. Functional Oligosaccharides: Production, Properties and Applications. World J. Microbiol. Biotechnol. 2011, 27. [Google Scholar] [CrossRef]
- Smilowitz, J.T.; Lebrilla, C.B.; Mills, D.A.; German, J.B.; Freeman, S.L. Breast Milk Oligosaccharides: Structure-Function Relationships in the Neonate. Annu. Rev. Nutr. 2014, 34, 143–169. [Google Scholar] [CrossRef]
- Wicinski, M.; Sawicka, E.; Gebalski, J.; Kubiak, K.; Malinowski, B. Human Milk Oligosaccharides: Health Benefits, Potential Applications in Infant Formulas, and Pharmacology. Nutrients 2020, 12, 266. [Google Scholar] [CrossRef] [PubMed]
- Bode, L. Human milk oligosaccharides: Every baby needs a sugar mama. Glycobiology 2012, 22, 1147–1162. [Google Scholar] [CrossRef] [PubMed]
- Craft, K.M.; Townsend, S.D. Mother Knows Best: Deciphering the Antibacterial Properties of Human Milk Oligosaccharides. Acc. Chem. Res. 2019, 52, 760–768. [Google Scholar] [CrossRef]
- Martín-Sosa, S.; Martín, M.-J.; Hueso, P. The sialylated fraction of milk oligosaccharides is partially responsible for binding to enterotoxigenic and uropathogenic Escherichia coli human strains. J. Nutr. 2002, 132, 3067–3072. [Google Scholar] [CrossRef]
- Coppa, G.V.; Gabrielli, O.; Giorgi, P.; Catassi, C.; Montanari, M.P.; Varaldo, P.E.; Nichols, B.L. Preliminary study of breastfeeding and bacterial adhesion to uroepithelial cells. Lancet 1990, 335, 569–571. [Google Scholar] [CrossRef]
- Andersson, B.; Porras, O.; Hanson, L.A.; Lagergfird, T.; Svanborg-Eden, C. Inhibition of Attachment of Streptococcus pneumoniae and Haemophilus influenzae by Human Milk and Receptor Oligosaccharides. J. Infect. Dis. 1986, 153, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.A.N.; Clare, S.; Goulding, D.; Arasteh, J.M.; Stares, M.D.; Browne, H.P.; Keane, J.A.; Page, A.J.; Kumasaka, N.; Kane, L.; et al. Epithelial IL-22RA1-mediated fucosylation promotes intestinal colonization resistance to an opportunistic pathogen. Cell Host Microbe 2014, 16, 504–516. [Google Scholar] [CrossRef]
- Asahara, T.; Takahashi, A.; Yuki, N.; Kaji, R.; Takahashi, T.; Nomoto, K. Protective effect of a synbiotic against multidrug-resistant Acinetobacter baumannii in a murine infection model. Antimicrob. Agents Chemother. 2016, 60, 3041–3050. [Google Scholar] [CrossRef]
- Goto, Y.; Obata, T.; Kunisawa, J.; Sato, S.; Ivanov, I.I.; Lamichhane, A.; Takeyama, N.; Kamioka, M.; Sakamoto, M.; Matsuki, T.; et al. Innate lymphoid cells regulate intestinal epithelial cell glycosylation. Science 2014, 345. [Google Scholar] [CrossRef] [PubMed]
- Newburg, D.S.; Ruiz-Palacios, G.M.; Morrow, A.L. Human Milk Glycans Protect Infants Against Enteric Pathogens. Annu. Rev. Nutr. 2005, 25, 37–58. [Google Scholar] [CrossRef] [PubMed]
- Cravioto, A.; Tello, A.; Villafán, H.; Ruiz, J.; del Vedovo, S.; Neeser, J.-R. Inhibition of Localized Adhesion of Enteropathogenic Escherichia coli to HEp-2 Cells by Immunoglobulin and Oligosaccharide Fractions of Human Colostrum and Breast Milk. J. Infect. Dis. 1991, 163, 1247–1255. [Google Scholar] [CrossRef] [PubMed]
- Hakkarainen, J.; Toivanen, M.; Leinonen, A.; Frängsmyr, L.; Strömberg, N.; Lapinjoki, S.; Nassif, X.; Tikkanen-Kaukanen, C. Human and Bovine Milk Oligosaccharides Inhibit Neisseria meningitidis Pili Attachment In Vitro. J. Nutr. 2005, 135, 2445–2448. [Google Scholar] [CrossRef]
- Kobata, A. Structures and application of oligosaccharides in human milk. Proc. Jpn. Acad. Ser. Phys. Biol. Sci. 2010, 86, 731–747. [Google Scholar] [CrossRef]
- Craft, K.M.; Gaddy, J.A.; Townsend, S.D. Human Milk Oligosaccharides (HMOs) Sensitize Group B Streptococcus to Clindamycin, Erythromycin, Gentamicin, and Minocycline on a Strain Specific Basis. ACS Chem. Biol. 2018, 13, 2020–2026. [Google Scholar] [CrossRef]
- Hsuchen, C.C.; Feingold, D.S. The Mechanism of Polymyxin B Action and Selectivity toward Biologic Membranes. Biochemistry 1973, 12, 2105–2111. [Google Scholar] [CrossRef]
- Lin, A.E.; Autran, C.A.; Espanola, S.D.; Bode, L.; Nizet, V. Human milk oligosaccharides protect bladder epithelial cells against uropathogenic escherichia coli invasion and cytotoxicity. J. Infect. Dis. 2014, 209, 389–398. [Google Scholar] [CrossRef]
- Turner, C.E. Paxillin and focal adhesion signalling. Nat. Cell Biol. 2000, 2, 231–236. [Google Scholar] [CrossRef]
- Schumacher, G.; Bendas, G.; Stahl, B.; Beermann, C. Human milk oligosaccharides affect P-selectin binding capacities: In vitro investigation. Nutrition 2006, 22, 620–627. [Google Scholar] [CrossRef]
- Craft, K.M.; Thomas, H.C.; Townsend, S.D. Interrogation of Human Milk Oligosaccharide Fucosylation Patterns for Antimicrobial and Antibiofilm Trends in Group B Streptococcus. ACS Infect. Dis. 2018, 4, 1755–1765. [Google Scholar] [CrossRef] [PubMed]
- Ackerman, D.L.; Craft, K.M.; Doster, R.S.; Weitkamp, J.H.; Aronoff, D.M.; Gaddy, J.A.; Townsend, S.D. Antimicrobial and Antibiofilm Activity of Human Milk Oligosaccharides against Streptococcus agalactiae, Staphylococcus aureus, and Acinetobacter baumannii. ACS Infect. Dis. 2018, 4, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Craft, K.M.; Thomas, H.C.; Townsend, S.D. Sialylated variants of lacto-: N-tetraose exhibit antimicrobial activity against Group B Streptococcus. Org. Biomol. Chem. 2019, 17, 1893–1900. [Google Scholar] [CrossRef] [PubMed]
- Bode, L.; Jantscher-Krenn, E. Structure-function relationships of human milk oligosaccharides. Adv. Nutr. 2012, 3, 383S–391S. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Palacios, G.M.; Cervantes, L.E.; Ramos, P.; Chavez-Munguia, B.; Newburg, D.S. Campylobacter jejuni binds intestinal H(O) antigen (Fuc alpha 1, 2Gal beta 1, 4GlcNAc), and fucosyloligosaccharides of human milk inhibit its binding and infection. J. Biol. Chem. 2003, 278, 14112–14120. [Google Scholar] [CrossRef] [PubMed]
- Weichert, S.; Jennewein, S.; Hufner, E.; Weiss, C.; Borkowski, J.; Putze, J.; Schroten, H. Bioengineered 2’-fucosyllactose and 3-fucosyllactose inhibit the adhesion of Pseudomonas aeruginosa and enteric pathogens to human intestinal and respiratory cell lines. Nutr. Res. 2013, 33, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Obermeier, S.; Rudloff, S.; Pohlentz, G.; Lentze, M.J.; Kunz, C. Secretion of 13C-labelled oligosaccharides into human milk and infant’s urine after an oral [13C]galactose load. Isotopes Environ. Health Stud. 1999, 35, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Craft, K.M.; Townsend, S.D. 1-Amino-2’-fucosyllactose inhibits biofilm formation by Streptococcus agalactiae. J. Antibiot. Tokyo 2019, 72, 507–512. [Google Scholar] [CrossRef]
- Thurl, S.; Munzert, M.; Boehm, G.; Matthews, C.; Stahl, B. Systematic review of the concentrations of oligosaccharides in human milk. Nutr. Rev. 2017, 75, 920–933. [Google Scholar] [CrossRef]
- Ninonuevo, M.R.; Park, Y.; Yin, H.; Zhang, J.; Ward, R.E.; Clowers, B.H.; German, J.B.; Freeman, S.L.; Killeen, K.; Grimm, R.; et al. A strategy for annotating the human milk glycome. J. Agric. Food Chem. 2006, 54, 7471–7480. [Google Scholar] [CrossRef]
- Marotta, M.; Ryan, J.T.; Hickey, R.M. The predominant milk oligosaccharide 6’-sialyllactose reduces the internalisation of Pseudomonas aeruginosa in human pneumocytes. J. Funct. Foods 2014, 6, 367–373. [Google Scholar] [CrossRef]
- Lee, J.M.; Oh, S.Y.; Johnston, T.V.; Ku, S.; Ji, G.E. Biocatalysis of fucodian in undaria pinnatifida sporophyll using bifidobacterium longum rd47 for production of prebiotic fucosylated oligosaccharide. Mar. Drugs 2019, 17, 117. [Google Scholar] [CrossRef] [PubMed]
- Fitton, J.H. Therapies from fucoidan; multifunctional marine polymers. Mar. Drugs 2011, 9, 1731–1760. [Google Scholar] [CrossRef] [PubMed]
- Hanisch, F.G.; Hansman, G.S.; Morozov, V.; Kunz, C.; Schroten, H. Avidity of -fucose on human milk oligosaccharides and blood group–unrelated oligo/polyfucoses is essential for potent norovirus-binding targets. J. Biol. Chem. 2018, 293, 11955–11965. [Google Scholar] [CrossRef]
- Newburg, D.S.; Ruiz-Palacios, G.M.; Altaye, M.; Chaturvedi, P.; Meinzen-Derr, J.; de Lourdes Guerrero, M.; Morrow, A.L. Innate protection conferred by fucosylated oligosaccharides of human milk against diarrhea in breastfed infants. Glycobiology 2004, 14, 253–263. [Google Scholar] [CrossRef]
- Rehm, B.H.A. Bacterial polymers: Biosynthesis, modifications and applications. Nat. Rev. Microbiol. 2010, 8, 578. [Google Scholar] [CrossRef]
- Lattner, D.; Flemming, H.-C.; Mayer, C. 13C-NMR study of the interaction of bacterial alginate with bivalent cations. Int. J. Biol. Macromol. 2003, 33, 81–88. [Google Scholar] [CrossRef]
- Skjåk-Bræk, G.; Grasdalen, H.; Larsen, B. Monomer sequence and acetylation pattern in some bacterial alginates. Carbohydr. Res. 1986, 154, 239–250. [Google Scholar] [CrossRef]
- Remminghorst, U.; Rehm, B.H.A. Bacterial alginates: From biosynthesis to applications. Biotechnol. Lett. 2006, 28, 1701–1712. [Google Scholar] [CrossRef]
- Schurks, N.; Wingender, J.; Flemming, H.-C.; Mayer, C. Monomer composition and sequence of alginates from Pseudomonas aeruginosa. Int. J. Biol. Macromol. 2002, 30, 105–111. [Google Scholar] [CrossRef]
- Overhage, J.; Lewenza, S.; Marr, A.K.; Hancock, R.E.W. Identification of genes involved in swarming motility using a Pseudomonas aeruginosa PAO1 mini-Tn5-lux mutant library. J. Bacteriol. 2007, 189, 2164–2169. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Tøndervik, A.; Sletta, H.; Klinkenberg, G.; Emanuel, C.; Onsøyen, E.; Myrvold, R. Overcoming Drug Resistance with Alginate Oligosaccharides Able To Potentiate the Action of Selected Antibiotics. Antimicrob. Agents Chemother. 2012, 56, 5134–5141. [Google Scholar] [CrossRef] [PubMed]
- Powell, L.C.; Sowedan, A.; Khan, S.; Wright, C.J.; Onsøyen, E.; Myrvold, R.; Hill, K.E.; Thomas, D.W. The effect of alginate oligosaccharides on the mechanical properties of Gram-negative biofilms. Biofouling 2013, 7014. [Google Scholar] [CrossRef] [PubMed]
- Powell, L.C.; Pritchard, M.F.; Emanuel, C.; Onsøyen, E.; Rye, P.D.; Wright, C.J.; Hill, K.E.; Thomas, D.W. Original Research: A Nanoscale Characterization of the Interaction of a Novel Alginate Oligomer with the Cell Surface and Motility of Pseudomonas aeruginosa. Am. J. Respir. Cell Mol. Biol. 2014, 50, 483–492. [Google Scholar] [CrossRef]
- Pritchard, M.F.; Powell, L.C.; Jack, A.A.; Powell, K.; Beck, K.; Florance, H.; Forton, J.; Rye, P.D.; Dessen, A.; Hill, K.E.; et al. A Low-Molecular-Weight Alginate Oligosaccharide Disrupts Pseudomonal Microcolony Formation and Enhances Antibiotic Effectiveness. Antimicrob. Agents Chemother. 2017, 61, 1–14. [Google Scholar] [CrossRef]
- Pritchard, M.F.; Powell, L.C.; Khan, S.; Griffiths, P.C.; Mansour, O.T.; Schweins, R.; Beck, K.; Buurma, N.J.; Dempsey, C.E.; Wright, C.J.; et al. The antimicrobial effects of the alginate oligomer OligoG CF-5/20 are independent of direct bacterial cell membrane disruption. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- Overhage, J.; Bains, M.; Brazas, M.D.; Hancock, R.E.W. Swarming of Pseudomonas aeruginosa is a complex adaptation leading to increased production of virulence factors and antibiotic resistance. J. Bacteriol. 2008, 190, 2671–2679. [Google Scholar] [CrossRef]
- Powell, L.C.; Pritchard, M.F.; Ferguson, E.L.; Powell, K.A.; Patel, S.U.; Rye, P.D.; Sakellakou, S.-M.; Buurma, N.J.; Brilliant, C.D.; Copping, J.M.; et al. Targeted disruption of the extracellular polymeric network of Pseudomonas aeruginosa biofilms by alginate oligosaccharides. NPJ Biofilms Microbiomes 2018, 4. [Google Scholar] [CrossRef]
- Roberts, J.L.; Khan, S.; Emanuel, C.; Powell, L.C.; Pritchard, M.F.; Onsoyen, E.; Myrvold, R.; Thomas, D.W.; Hill, K.E. An in vitro study of alginate oligomer therapies on oral biofilms. J. Dent. 2013, 41, 892–899. [Google Scholar] [CrossRef]
- Korstgens, V.; Flemming, H.C.; Wingender, J.; Borchard, W. Influence of calcium ions on the mechanical properties of a model biofilm of mucoid Pseudomonas aeruginosa. Water Sci. Technol. 2001, 43, 49–57. [Google Scholar] [CrossRef]
- Aslam, S.N.; Newman, M.-A.; Erbs, G.; Morrissey, K.L.; Chinchilla, D.; Boller, T.; Jensen, T.T.; De Castro, C.; Ierano, T.; Molinaro, A.; et al. Bacterial polysaccharides suppress induced innate immunity by calcium chelation. Curr. Biol. 2008, 18, 1078–1083. [Google Scholar] [CrossRef] [PubMed]
- Pier, G.B.; Coleman, F.; Grout, M.; Franklin, M.; Ohman, D.E. Role of Alginate O Acetylation in Resistance of Mucoid Pseudomonas aeruginosa to Opsonic Phagocytosis. Infect. Immun. 2001, 69, 1895–1901. [Google Scholar] [CrossRef] [PubMed]
- Nivens, D.E.; Ohman, D.E.; Williams, J.; Franklin, M.J. Role of alginate and its O acetylation in formation of Pseudomonas aeruginosa microcolonies and biofilms. J. Bacteriol. 2001, 183, 1047–1057. [Google Scholar] [CrossRef] [PubMed]
- Sarkisova, S.; Patrauchan, M.A.; Berglund, D.; Nivens, D.E.; Franklin, M.J. Calcium-induced virulence factors associated with the extracellular matrix of mucoid Pseudomonas aeruginosa biofilms. J. Bacteriol. 2005, 187, 4327–4337. [Google Scholar] [CrossRef] [PubMed]
- Banin, E.; Vasil, M.L.; Greenberg, E.P. Iron and Pseudomonas aeruginosa biofilm formation. Proc. Natl. Acad. Sci. USA 2005, 102, 11076–11081. [Google Scholar] [CrossRef]
- Wiens, J.R.; Vasil, A.I.; Schurr, M.J.; Vasil, M.L. Iron-regulated expression of alginate production, mucoid phenotype, and biofilm formation by Pseudomonas aeruginosa. MBio 2014, 5, e01010–e1013. [Google Scholar] [CrossRef]
- Falkeborg, M.; Cheong, L.-Z.; Gianfico, C.; Sztukiel, K.M.; Kristensen, K.; Glasius, M.; Xu, X.; Guo, Z. Alginate oligosaccharides: Enzymatic preparation and antioxidant property evaluation. Food Chem. 2014, 164, 185–194. [Google Scholar] [CrossRef]
- Jiang, Q.; Chen, J.; Yang, C.; Yin, Y.; Yao, K.; Song, D. Quorum Sensing: A Prospective Therapeutic Target for Bacterial Diseases. Biomed. Res. Int. 2019, 2019. [Google Scholar] [CrossRef]
- Williams, P.; Cámara, M. Quorum sensing and environmental adaptation in Pseudomonas aeruginosa: A tale of regulatory networks and multifunctional signal molecules. Curr. Opin. Microbiol. 2009, 12, 182–191. [Google Scholar] [CrossRef]
- Allesen-Holm, M.; Barken, K.B.; Yang, L.; Klausen, M.; Webb, J.S.; Kjelleberg, S.; Molin, S.; Givskov, M.; Tolker-Nielsen, T. A characterization of DNA release in Pseudomonas aeruginosa cultures and biofilms. Mol. Microbiol. 2006, 59, 1114–1128. [Google Scholar] [CrossRef]
- Yang, L.; Barken, K.B.; Skindersoe, M.E.; Christensen, A.B.; Givskov, M.; Tolker-Nielsen, T. Effects of iron on DNA release and biofilm development by Pseudomonas aeruginosa. Microbiology 2007, 153, 1318–1328. [Google Scholar] [CrossRef] [PubMed]
- Whitchurch, C.B.; Tolker-Nielsen, T.; Ragas, P.C.; Mattick, J.S. Extracellular DNA required for bacterial biofilm formation. Science 2002, 295, 1487. [Google Scholar] [CrossRef]
- Jack, A.A.; Khan, S.; Powell, L.C.; Pritchard, M.F.; Beck, K.; Sadh, H.; Sutton, L.; Cavaliere, A.; Florance, H.; Rye, P.D.; et al. Alginate oligosaccharide-induced modification of the lasI-lasR and rhlI-rhlR quorum-sensing systems in pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [PubMed]
- Phillips, P.L.; Schultz, G.S. Molecular Mechanisms of Biofilm Infection: Biofilm Virulence Factors. Adv. Wound Care 2012, 1, 109–114. [Google Scholar] [CrossRef]
- Das, T.; Manefield, M. Pyocyanin Promotes Extracellular DNA Release in Pseudomonas aeruginosa. PLoS ONE 2012, 7, e46718. [Google Scholar] [CrossRef]
- Elkins, J.G.; Hassett, D.J.; Stewart, P.S.; Schweizer, H.P.; McDermott, T.R. Protective role of catalase in Pseudomonas aeruginosa biofilm resistance to hydrogen peroxide. Appl. Environ. Microbiol. 1999, 65, 4594–4600. [Google Scholar] [CrossRef]
- He, X.; Hwang, H.; Aker, W.G.; Wang, P.; Lin, Y.; Jiang, X.; He, X.; Sim, C.O.S. Synergistic combination of marine oligosaccharides and azithromycin against Pseudomonas aeruginosa. Microbiol. Res. 2014, 169, 759–767. [Google Scholar] [CrossRef]
- Jollès, P.; Muzzarelli, R.A.A. Chitin and Chitinases; Birkhäuser Verlag: Basel, Switzerland, 1999; ISBN 9783764358150. [Google Scholar]
- Liaqat, F.; Eltem, R. Chitooligosaccharides and their biological activities: A comprehensive review. Carbohydr. Polym. 2018, 184, 243–259. [Google Scholar] [CrossRef]
- Lodhi, G.; Kim, Y.-S.; Hwang, J.-W.; Kim, S.-K.; Jeon, Y.-J.; Je, J.-Y.; Ahn, C.-B.; Moon, S.-H.; Jeon, B.-T.; Park, P.-J. Chitooligosaccharide and its derivatives: Preparation and biological applications. Biomed. Res. Int. 2014, 2014, 654913. [Google Scholar] [CrossRef]
- Mourya, V.K.; Inamdar, N.N.; Choudhari, Y.M. Chitooligosaccharides: Synthesis, characterization and applications. Polym. Sci. Ser. A 2011, 53, 583–612. [Google Scholar] [CrossRef]
- Kyoon, H.; Young, N.; Ho, S.; Meyers, S.P. Antibacterial activity of chitosans and chitosan oligomers with different molecular weights. Int. J. Food Microbiol. 2002, 74, 65–72. [Google Scholar]
- Zheng, L.-Y.; Zhu, J.-F. Study on antimicrobial activity of chitosan with different molecular weights. Carbohydr. Polym. 2003, 54, 527–530. [Google Scholar] [CrossRef]
- Fernandes, J.C.; Tavaria, F.K.; Soares, J.C.; Ramos, O.S.; João Monteiro, M.; Pintado, M.E.; Xavier Malcata, F. Antimicrobial effects of chitosans and chitooligosaccharides, upon Staphylococcus aureus and Escherichia coli, in food model systems. Food Microbiol. 2008, 25, 922–928. [Google Scholar] [CrossRef]
- Kim, S.-K.; Rajapakse, N. Enzymatic production and biological activities of chitosan oligosaccharides (COS): A review. Carbohydr. Polym. 2005, 62, 357–368. [Google Scholar] [CrossRef]
- Kurita, K.; Kaji, Y.; Mori, T.; Nishiyama, Y. Enzymatic degradation of β-chitin: Susceptibility and the influence of deacetylation. Carbohydr. Polym. 2000, 42, 19–21. [Google Scholar] [CrossRef]
- Riegman, N.; Kusters, R.; Van Veggel, H.; Bergmans, H.; Van Bergen en Henegouwen, P.; Hacker, J.; Van Die, I. F1C fimbriae of a uropathogenic Escherichia coli strain: Genetic and functional organization of the foc gene cluster and identification of minor subunits. J. Bacteriol. 1990, 172, 1114–1120. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.K.; Park, P.J.; Yang, H.P.; Han, S.S. Subacute toxicity of chitosan oligosaccharide in Sprague-Dawley rats. Arzneimittelforschung 2001, 51, 769–774. [Google Scholar] [CrossRef]
- VandeVord, P.J.; Matthew, H.W.T.; DeSilva, S.P.; Mayton, L.; Wu, B.; Wooley, P.H. Evaluation of the biocompatibility of a chitosan scaffold in mice. J. Biomed. Mater. Res. 2002, 59, 585–590. [Google Scholar] [CrossRef]
- Allan, C.R.; Hadwiger, L.A. The fungicidal effect of chitosan on fungi of varying cell wall composition. Exp. Mycol. 1979, 3, 285–287. [Google Scholar] [CrossRef]
- Choi, B.; Kim, K.; Yoo, Y.; Oh, S. In vitro antimicrobial activity of a chitooligosaccharide mixture against Actinobacillus actinomycetemcomitans and Streptococcus mutans. Int. J. Antimicrob. Agents 2001, 18, 553–557. [Google Scholar] [CrossRef]
- Sanchez, A.; Mengibar, M.; Rivera-Rodriguez, G.; Moerchbacher, B.; Acosta, N.; Heras, A. The effect of preparation processes on the physicochemical characteristics and antibacterial activity of chitooligosaccharides. Carbohydr. Polym. 2017, 157, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, J.C.; Tavaria, F.K.; Fonseca, S.C.; Ramos, O.S.; Pintado, M.E.; Malcata, F.X. In vitro screening for anti-microbial activity of chitosans and chitooligosaccharides, aiming at potential uses in functional textiles. J. Microbiol. Biotechnol. 2010, 20, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Jeon, Y.; Park, P.; Kim, S. Antimicrobial effect of chitooligosaccharides produced by bioreactor. Carbohydr. Polym. 2001, 44, 71–76. [Google Scholar] [CrossRef]
- Helander, I.M.; Latva-Kala, K.; Lounatmaa, K. Permeabilizing action of polyethyleneimine on Salmonella typhimurium involves disruption of the outer membrane and interactions with lipopolysaccharide. Microbiology 1998, 144, 385–390. [Google Scholar] [CrossRef]
- Demeneix, B.; Behr, J.B.T.-A. Polyethylenimine (PEI). In Non-Viral Vectors for Gene Therapy, 2nd ed.; Academic Press: San Diego, CA, USA, 2005; Volume 53, pp. 215–230. ISBN 0120176548. [Google Scholar]
- Vancha, A.R.; Govindaraju, S.; Parsa, K.V.L.; Jasti, M.; González-García, M.; Ballestero, R.P. Use of polyethyleneimine polymer in cell culture as attachment factor and lipofection enhancer. BMC Biotechnol. 2004, 4, 23. [Google Scholar] [CrossRef] [PubMed]
- Helander, I.M.; Nurmiaho-Lassila, E.L.; Ahvenainen, R.; Rhoades, J.; Roller, S. Chitosan disrupts the barrier properties of the outer membrane of gram-negative bacteria. Int. J. Food Microbiol. 2001, 71, 235–244. [Google Scholar] [CrossRef]
- Young, D.H.; Kohle, H.; Kauss, H. Effect of Chitosan on Membrane Permeability of Suspension-Cultured Glycine max and Phaseolus vulgaris Cells. Plant Physiol. 1982, 70, 1449–1454. [Google Scholar] [CrossRef]
- Young, D.H.; Kauss, H. Release of Calcium from Suspension-Cultured Glycine max Cells by Chitosan, Other Polycations, and Polyamines in Relation to Effects on Membrane Permeability. Plant Physiol. 1983, 73, 698–702. [Google Scholar] [CrossRef]
- Kumar, A.B.V.; Varadaraj, M.C.; Gowda, L.R.; Tharanathan, R.N. Characterization of chito-oligosaccharides prepared by chitosanolysis with the aid of papain and Pronase, and their bactericidal action against Bacillus cereus and Escherichia coli. Biochem. J. 2005, 175, 167–175. [Google Scholar]
- Moon, J.-S.; Kim, H.-K.; Koo, H.C.; Joo, Y.-S.; Nam, H.; Park, Y.H.; Kang, M.-I. The antibacterial and immunostimulative effect of chitosan-oligosaccharides against infection by Staphylococcus aureus isolated from bovine mastitis. Appl. Microbiol. Biotechnol. 2007, 75, 989–998. [Google Scholar] [CrossRef]
- Benhabiles, M.S.; Salah, R.; Lounici, H.; Drouiche, N.; Goosen, M.F.A.; Mameri, N. Food Hydrocolloids Antibacterial activity of chitin, chitosan and its oligomers prepared from shrimp shell waste. Food Hydrocoll. 2012, 29, 48–56. [Google Scholar] [CrossRef]
- Quintero-villegas, M.I.; Aam, B.B.; Rupnow, J.; Sørlie, M.; Eijsink, V.G.H.; Hutkins, R.W. Adherence Inhibition of Enteropathogenic Escherichia coli by Chitooligosaccharides with Speci fi c Degrees of Acetylation and Polymerization. Agric. Food Chem. 2013, 61, 2748–2754. [Google Scholar] [CrossRef] [PubMed]
- Rhoades, J.; Gibson, G.; Formentin, K.; Beer, M.; Rastall, R. Inhibition of the adhesion of enteropathogenic Escherichia coli strains to HT-29 cells in culture by chito-oligosaccharides. Carbohydr. Polym. 2006, 64, 57–59. [Google Scholar] [CrossRef]
- Hadwiger, L.A.; Beckman, J.M. Chitosan as a Component of Pea-Fusarium solani Interactions. Plant Physiol. 1980, 66, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-F.; Feng, X.-Q.; Yang, S.; Wang, T.-P.; Su, Z.-X. Effects of molecular weight and concentration of chitosan on antifungal activity against aspergillus niger. Iran. Polym. J. Engl. Ed. 2008, 17, 843–852. [Google Scholar]
- Sebti, I.; Martial-Gros, A.; Carnet-Pantiez, A.; Grelier, S.; Coma, V. Chitosan Polymer as Bioactive Coating and Film against Aspergillus niger Contamination. J. Food Sci. 2005, 70, M100–M104. [Google Scholar] [CrossRef]
- Wagoner, W.; Loschke, D.C.; Hadwiger, L.A. Two-dimensional electrophoretic analysis of in vivo and in vitro synthesis of proteins in peas inoculated with compatible and incompatible Fusarium solani. Physiol. Plant Pathol. 1982, 99–107. [Google Scholar] [CrossRef]
- Liu, X.F.; Lin Guan, Y.; Zhi Yang, D.; Li, Z.; De Yao, K. Antibacterial action of chitosan and carboxymethylated chitosan. J. Appl. Polym. Sci. 2001, 79, 1324–1335. [Google Scholar]
- Raafat, D.; Sahl, H.-G. Chitosan and its antimicrobial potential—A critical literature survey. Microb. Biotechnol. 2009, 2, 186–201. [Google Scholar] [CrossRef]
- Nikaido, H. Multidrug resistance in bacteria. Annu. Rev. Biochem. 2009, 78, 119–146. [Google Scholar] [CrossRef]
- Tin, S.; Sing Lim, C.; Sakharkar, M.; Ramaji Sakharkar, K. Synergistic Combinations of Chitosans and Antibiotics in Staphylococcus Aureus; Bentham Science Publishers: Sharjah, UAE, 2010; Volume 7. [Google Scholar]
- Tin, S.; Sakharkar, K.R.; Lim, C.S.; Sakharkar, M.K. Activity of Chitosans in combination with antibiotics in Pseudomonas aeruginosa. Int. J. Biol. Sci. 2009, 5, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Jeon, Y.; Kim, S. Production of chitooligosaccharides using an ultrafiltration membrane reactor and their antibacterial activity. Carbohydr. Polym. 2000, 41, 133–141. [Google Scholar] [CrossRef]
- Lee, B.C.; Kim, S.M.; Choi, S.H.; Kim, K.Y.; Kim, T.S. In vitro and in vivo antimicrobial activity of water-soluble chitosan oligosaccharides against Vibrio vulnificus. Int. J. Mol. Med. 2009, 327–333. [Google Scholar] [CrossRef]
- Laokuldilok, T.; Potivas, T.; Kanha, N.; Surawang, S.; Seesuriyachan, P.; Wangtueai, S.; Phimolsiripol, Y.; Regenstein, J.M. Physicochemical, antioxidant, and antimicrobial properties of chitooligosaccharides produced using three different enzyme treatments. Food Biosci. 2017, 18, 28–33. [Google Scholar] [CrossRef]
- Roberfroid, M.B. Functional foods: Concepts and application to inulin and oligofructose. Br. J. Nutr. 2002, 87, S139–S143. [Google Scholar] [CrossRef] [PubMed]
- Al-Ghazzewi, F.H.; Tester, R.F. Inhibition of the adhesion of Escherichia coli to human epithelial cells by carbohydrates. Bioact. Carbohydr. Diet. Fibre 2014, 4, 1–5. [Google Scholar] [CrossRef]
- Kim, J.K.; Shin, E.C.; Park, H.G. Fructooligosaccharides decreased the ability of probiotic Escherichia coli Nissle 1917 to adhere to co-cultures of human intestinal cell lines. J. Korean Soc. Appl. Biol. Chem. 2015, 58, 45–52. [Google Scholar] [CrossRef]
- Ortega-González, M.; Sánchez De Medina, F.; Molina-Santiago, C.; López-Posadas, R.; Pacheco, D.; Krell, T.; Martínez-Augustin, O.; Abdelali, D. Fructooligosacharides reduce Pseudomonas aeruginosa PAO1 pathogenicity through distinct mechanisms. PLoS ONE 2014, 9, e85772. [Google Scholar] [CrossRef]
- Intanon, M.; Arreola, S.L.; Pham, N.H.; Kneifel, W.; Haltrich, D.; Nguyen, T.H. Nature and biosynthesis of galacto-oligosaccharides related to oligosaccharides in human breast milk. FEMS Microbiol. Lett. 2014, 353, 89–97. [Google Scholar] [CrossRef]
- Rastall, R.A. Functional oligosaccharides: Application and manufacture. Annu. Rev. Food Sci. Technol. 2010, 1, 305–339. [Google Scholar] [CrossRef]
- Coulier, L.; Timmermans, J.; Bas, R.; Van Den Dool, R.; Haaksman, I.; Klarenbeek, B.; Slaghek, T.; Van Dongen, W. In-depth characterization of prebiotic galacto-oligosaccharides by a combination of analytical techniques. J. Agric. Food Chem. 2009, 57, 8488–8495. [Google Scholar] [CrossRef] [PubMed]
- Petri, W.A., Jr.; Smith, R.D.; Schlesinger, P.H.; Murphy, C.F.; Ravdin, J.I. Isolation of the galactose-binding lectin that mediates the in vitro adherence of Entamoeba histolytica. J. Clin. Investig. 1987, 80, 1238–1244. [Google Scholar] [CrossRef] [PubMed]
- Petri, W.A., Jr.; Haque, R.; Mann, B.J. The bittersweet interface of parasite and host: Lectin-carbohydrate interactions during human invasion by the parasite Entamoeba histolytica. Annu. Rev. Microbiol. 2002, 56, 39–64. [Google Scholar] [CrossRef]
- Shoaf, K.; Mulvey, G.L.; Armstrong, G.D.; Hutkins, R.W. Prebiotic Galactooligosaccharides Reduce Adherence of Enteropathogenic Escherichia coli to Tissue Culture Cells. Infect. Immun. 2006, 74, 6920–6928. [Google Scholar] [CrossRef]
- Searle, L.E.J.; Cooley, W.A.; Jones, G.; Nunez, A.; Crudgington, B.; Weyer, U.; Dugdale, A.H.; Tzortzis, G.; Collins, J.W.; Woodward, M.J.; et al. Purified galactooligosaccharide, derived from a mixture produced by the enzymic activity of Bifidobacterium bifidum, reduces Salmonella enterica serovar Typhimurium adhesion and invasion in vitro and in vivo. J. Med. Microbiol. 2010, 1428–1439. [Google Scholar] [CrossRef]
- Kittana, H.; Quintero-Villegas, M.I.; Bindels, L.B.; Gomes-Neto, J.C.; Schmaltz, R.J.; Segura Munoz, R.R.; Cody, L.A.; Moxley, R.A.; Hostetter, J.; Hutkins, R.W.; et al. Galactooligosaccharide supplementation provides protection against citrobacter rodentium-induced colitis without limiting pathogen burden. Microbiology UK 2018, 164, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Ebersbach, T.; Bo, J.; Bergstro, A.; Hutkins, R.W.; Rask, T. Xylo-oligosaccharides inhibit pathogen adhesion to enterocytes in vitro. Res. Microbiol. 2012, 163, 22–27. [Google Scholar] [CrossRef]
- Quintero, M.; Maldonado, M.; Perez-Munoz, M.; Jimenez, R.; Fangman, T.; Rupnow, J.; Wittke, A.; Russell, M.; Hutkins, R. Adherence inhibition of Cronobacter sakazakii to intestinal epithelial cells by prebiotic oligosaccharides. Curr. Microbiol. 2011, 62, 1448–1454. [Google Scholar] [CrossRef]
- Mange, J.; Stephan, R.; Borel, N.; Wild, P.; Kim, K.S.; Pospischil, A.; Lehner, A. Adhesive properties of Enterobacter sakazakii to human epithelial and brain microvascular endothelial cells. BMC Microbiol. 2006, 10. [Google Scholar] [CrossRef]
- Muanprasat, C.; Chatsudthipong, V. Cholera: Pathophysiology and emerging therapeutic targets. Future Med. Chem. 2013, 5, 781–798. [Google Scholar] [CrossRef]
- Lanne, B.; Ångstrom, J.; Schierbeck, B. Binding of Cholera Toxin B-Subunits to Derivatives of the Natural Ganglioside Receptor, GM1. J. Biochem. 1999, 126, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Wallenfels, K.; Malhotra, O.P. Galactosidases. Adv. Carbohydr. Chem. 1962, 16, 239–298. [Google Scholar]
- Holmgren, J. Actions of cholera toxin and the prevention and treatment of cholera. Nature 1981, 292, 413–417. [Google Scholar] [CrossRef]
- Haksar, D.; Pieters, R.J. A hybrid polymer to target blood group dependence of cholera toxin. Org. Biomol. Chem. 2020, 18, 52–55. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, H.R.; De Slegte, J.; Gibson, G.R.; Rastall, R.A. Galactooligosaccharides (GOS) inhibit vibrio cholerae toxin binding to its GM1 receptor. J. Agric. Food Chem. 2009, 57, 3113–3119. [Google Scholar] [CrossRef] [PubMed]
- Ommen, D.D.Z.; Pukin, A.V.; Fu, O.; Quarles, L.H.C.; Van, U.; Janssens, H.M.; Beekman, M.; Pieters, R.J. Functional Characterization of Cholera Toxin Inhibitors Using Human Intestinal Organoids. J. Med. Chem. 2016, 59, 6968–6972. [Google Scholar] [CrossRef]
- Ofek, I.; Beachey, E.H. Mannose binding and epithelial cell adherence of Escherichia coli. Infect. Immun. 1978, 22, 247–254. [Google Scholar] [CrossRef]
- Ofek, I.; Mirelman, D.; Sharon, N. Adherence of Escherichia coli to human mucosal cells mediated by mannose receptors. Nature 1977, 265, 623–625. [Google Scholar] [CrossRef]
- Firon, N.; Ofek, I.; Sharon, N. Carbohydrate specificity of the surface lectins of Escherichia coli, Klebsiella pneumoniae, and Salmonella typhimurium. Carbohydr. Res. 1983, 120, 235–249. [Google Scholar] [CrossRef]
- Liu, X.-Y.; Wang, Q.; Cui, S.W.; Liu, H.-Z. A new isolation method of β-d-glucans from spent yeast Saccharomyces cerevisiae. Food Hydrocoll. 2008, 22, 239–247. [Google Scholar] [CrossRef]
- Jones, C.H.; Pinkner, J.S.; Roth, R.; Heuser, J.; Nicholes, A.V.; Abraham, S.N.; Hultgren, S.J. FimH adhesin of type 1 pili is assembled into a fibrillar tip structure in the Enterobacteriaceae. Proc. Natl. Acad. Sci. USA 1995, 92, 2081–2085. [Google Scholar] [CrossRef] [PubMed]
- Krogfelt, K.A.; Bergmans, H.; Klemm, P. Direct evidence that the FimH protein is the mannose-specific adhesin of Escherichia coli type 1 fimbriae. Infect. Immun. 1990, 58, 1995–1998. [Google Scholar] [CrossRef] [PubMed]
- Firon, N.; Ofek, I.; Sharon, N. Carbohydrate-binding sites of the mannose-specific fimbrial lectins of enterobacteria. Infect. Immun. 1984, 43, 1088–1090. [Google Scholar] [CrossRef] [PubMed]
- Firon, N.; Ashkenazi, S.; Mirelman, D.; Ofek, I.; Sharon, N. Aromatic alpha-glycosides of mannose are powerful inhibitors of the adherence of type 1 fimbriated Escherichia coli to yeast and intestinal epithelial cells. Infect. Immun. 1987, 55, 472–476. [Google Scholar] [CrossRef]
- Lindhorst, T.K.; Kieburg, C.; Krallmann-Wenzel, U. Inhibition of the type 1 fimbriae-mediated adhesion of Escherichia coli to erythrocytes by multiantennary alpha-mannosyl clusters: The effect of multivalency. Glycoconj. J. 1998, 15, 605–613. [Google Scholar] [CrossRef]
- Bouckaert, J.; Mackenzie, J.; De Paz, J.L.; Chipwaza, B.; Choudhury, D.; Zavialov, A.; Mannerstedt, K.; Anderson, J.; Piérard, D.; Wyns, L.; et al. The affinity of the FimH fimbrial adhesin is receptor-driven and quasi-independent of Escherichia coli pathotypes. Mol. Microbiol. 2006, 61, 1556–1568. [Google Scholar] [CrossRef]
- Rosen, D.A.; Pinkner, J.S.; Walker, J.N.; Elam, J.S.; Jones, J.M.; Hultgren, S.J. Molecular variations in Klebsiella pneumoniae and Escherichia coli FimH affect function and pathogenesis in the urinary tract. Infect. Immun. 2008, 76, 3346–3356. [Google Scholar] [CrossRef]
- Sokurenko, E.V.; Chesnokova, V.; Dykhuizen, D.E.; Ofek, I.; Wu, X.R.; Krogfelt, K.A.; Struve, C.; Schembri, M.A.; Hasty, D.L. Pathogenic adaptation of Escherichia coli by natural variation of the FimH adhesin. Proc. Natl. Acad. Sci. USA 1998, 95, 8922–8926. [Google Scholar] [CrossRef]
- Ramirez-Hernandez, A.; Rupnow, J.; Hutkins, R.W. Adherence reduction of campylobacter jejuni and campylobacter coli strains to HEp-2 cells by mannan oligosaccharides and a high-molecular-weight component of cranberry extract. J. Food Prot. 2015, 78, 1496–1505. [Google Scholar] [CrossRef]
- Ganner, A.; Schatzmayr, G. Capability of yeast derivatives to adhere enteropathogenic bacteria and to modulate cells of the innate immune system. Appl. Microbiol. Biotechnol. 2012, 95, 289–297. [Google Scholar] [CrossRef]
- Ganner, A.; Stoiber, C.; Uhlik, J.T.; Dohnal, I.; Schatzmayr, G. Quantitative evaluation of E. coli F4 and Salmonella Typhimurium binding capacity of yeast derivatives. AMB Express 2013, 3, 62. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Qiao, Y.; Peng, Q.; Gao, L.; Shi, B. Inhibitory effects of YCW and MOS from Saccharomyces cerevisiae on Escherichia coli and Salmonella pullorum adhesion to Caco-2 cells. Front. Biol. Beijing 2017, 12, 370–375. [Google Scholar] [CrossRef]
- Firon, N.; Ofek, I.; Sharon, N. Interaction of mannose-containing oligosaccharides with the fimbrial lectin of Escherichia coli. Biochem. Biophys. Res. Commun. 1982, 105, 1426–1432. [Google Scholar] [CrossRef]
- Elbein, A.D.; Rock, L.; Examiner, P.; Saucier, S.E. Inhibition of Bacterial Binding by High-Mannose Oligosacchardes. USA Patent 5,939,279, 17 August 1999. [Google Scholar]
- Mualikrishna, G.; Tharanathan, R.N. Characterization of pectic polysaccharides from pulse husks. Food Chem. 1994, 50, 87–89. [Google Scholar] [CrossRef]
- Thakur, B.R.; Singh, R.K.; Handa, A.K. Chemistry and uses of pectin—A review. Crit. Rev. Food Sci. Nutr. 1997, 37, 47–73. [Google Scholar] [CrossRef]
- May, C.D. Industrial pectins: Sources, production and applications. Carbohydr. Polym. 1990, 12, 79–99. [Google Scholar] [CrossRef]
- Morris, G.A.; Foster, T.J.; Harding, S.E. The effect of the degree of esterification on the hydrodynamic properties of citrus pectin. Food Hydrocoll. 2000, 14, 227–235. [Google Scholar] [CrossRef]
- Mandalari, G.; Bennett, R.N.; Kirby, A.R.; Lo Curto, R.B.; Bisignano, G.; Waldron, K.W.; Faulds, C.B. Enzymatic Hydrolysis of Flavonoids and Pectic Oligosaccharides from Bergamot (Citrus bergamia Risso) Peel. J. Agric. Food Chem. 2006, 54, 8307–8313. [Google Scholar] [CrossRef]
- Olano-Martin, E.; Mountzouris, K.C.; Gibson, G.R.; Rastall, R.A. Continuous Production of Pectic Oligosaccharides in an Enzyme Membrane Reactor. J. Food Sci. 2001, 66, 966–971. [Google Scholar] [CrossRef]
- Nangia-Makker, P.; Conklin, J.; Hogan, V.; Raz, A. Carbohydrate-binding proteins in cancer, and their ligands as therapeutic agents. Trends Mol. Med. 2002, 8, 187–192. [Google Scholar] [CrossRef]
- Li, P.; Xia, J.; Nie, Z.; Shan, Y. Pectic oligosaccharides hydrolyzed from orange peel by fungal multi-enzyme complexes and their prebiotic and antibacterial potentials. LWT Food Sci. Technol. 2016, 69, 203–210. [Google Scholar] [CrossRef]
- Ganan, M.; Collins, M.; Rastall, R.; Hotchkiss, A.T.; Chau, H.K.; Carrascosa, A.V.; Martinez-Rodriguez, A.J. Inhibition by pectic oligosaccharides of the invasion of undifferentiated and differentiated Caco-2 cells by Campylobacter jejuni. Int. J. Food Microbiol. 2010, 137, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, T.; Zhu, R.; Wang, N.; Song, Y.; Wang, S.; Guo, M. Antibacterial Action of Haw Pectic Oligosaccharides. Int. J. Food Prop. 2013, 16. [Google Scholar] [CrossRef]
- Pristov, J.B.; Jovanović, S.V.; Mitrović, A.; Spasojević, I. UV-irradiation provokes generation of superoxide on cell wall polygalacturonic acid. Physiol. Plant. 2013, 148, 574–581. [Google Scholar] [CrossRef]
- Tazawa, K.; Hanmyo, K.; Namikawa, H.; Mizumoto, J.; Oida, N.; Ohkami, H.; Saito, T.; Okamoto, M.; Yamashita, I.; Komatu, K.; et al. Inhibitory effects of apple pectin in colon carcinogenesis—Evaluation of the radical scavenging activity of pectic oligosaccharide. Biotherapy 1999, 13, 510–512. [Google Scholar]
- Marx, J.L. Oxygen free radicals linked to many diseases. Science 1987, 235, 529–531. [Google Scholar] [CrossRef]
- Zheng, R.L.; Lesko, S.A.; Ts’o, P.O. DNA damage induced in mammalian cells by active oxygen species. Sci. Sin. Ser. B Chem. Biol. Agric. Med. Earth Sci. Chung Kuo K Hsueh Yuan Chu Pan 1988, 31, 676–686. [Google Scholar]
- Slemmer, J.E.; Shacka, J.J.; Sweeney, M.I.; Weber, J.T. Antioxidants and free radical scavengers for the treatment of stroke, traumatic brain injury and aging. Curr. Med. Chem. 2008, 15, 404–414. [Google Scholar]
- Martinov, J.; Krstić, M.; Spasić, S.; Miletić, S.; Stefanović-Kojić, J.; Nikolić-Kokić, A.; Blagojević, D.; Spasojević, I.; Spasić, M.B. Apple pectin-derived oligosaccharides produce carbon dioxide radical anion in Fenton reaction and prevent growth of Escherichia coli and Staphylococcus aureus. Food Res. Int. 2017, 100, 132–136. [Google Scholar] [CrossRef]
- Olano-Martin, E.; Williams, M.R.; Gibson, G.R.; Rastall, R.A. Pectins and pectic-oligosaccharides inhibit Escherichia coli O157:H7 Shiga toxin as directed towards the human colonic cell line HT29. FEMS Microbiol. Lett. 2003, 218, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Rhoades, J.; Manderson, K.; Wells, A.; Hotchkiss, A.; Gibson, G.; Formentin, K.; Beer, M.; Rastall, R.A. Oligosaccharide-Mediated Inhibition of the Adhesion of Pathogenic Escherichia coli Strains to Human Gut Epithelial Cells In Vitro. J. Food Prot. 2008, 71, 2272–2277. [Google Scholar] [CrossRef] [PubMed]
- Guggenbichler, J.P.; De Bettignies-Dutz, A.; Meissner, P.; Schellmoser, S.; Jurenitsch, J. Acidic oligosaccharides from natural sources block adherence of Escherichia coli on uroepithelial cells. Pharm. Pharmacol. Lett. 1997, 7, 35–38. [Google Scholar]
- Di, R.; Vakkalanka, M.S.; Onumpai, C.; Chau, H.K.; White, A.; Rastall, R.A.; Yam, K.; Hotchkiss, A.T., Jr. Pectic oligosaccharide structure-function relationships: Prebiotics, inhibitors of Escherichia coli O157:H7 adhesion and reduction of Shiga toxin cytotoxicity in HT29 cells. Food Chem. 2017, 227, 245–254. [Google Scholar] [CrossRef]
- Lee, J.-H.; Shim, J.S.; Lee, J.S.; Kim, M.-K.; Chung, M.-S.; Kim, K.H. Pectin-like acidic polysaccharide from Panax ginseng with selective antiadhesive activity against pathogenic bacteria. Carbohydr. Res. 2006, 341, 1154–1163. [Google Scholar] [CrossRef]
- Cohen, A.; Hannigan, G.E.; Williams, B.R.; Lingwood, C.A. Roles of globotriosyl- and galabiosylceramide in verotoxin binding and high affinity interferon receptor. J. Biol. Chem. 1987, 262, 17088–17091. [Google Scholar]
- Lingwood, C.A.; Law, H.; Richardson, S.; Petric, M.; Brunton, J.L.; De Grandis, S.; Karmali, M. Glycolipid binding of purified and recombinant Escherichia coli produced verotoxin in vitro. J. Biol. Chem. 1987, 262, 8834–8839. [Google Scholar]
- Bergan, J.; Dyve Lingelem, A.B.; Simm, R.; Skotland, T.; Sandvig, K. Shiga toxins. Toxicon 2012, 60, 1085–1107. [Google Scholar] [CrossRef]
- Melton-Celsa, A.R. Shiga Toxin (Stx) Classification, Structure, and Function. Microbiol. Spectr. 2014, 2. [Google Scholar] [CrossRef]
- Keusch, G.T.; Jacewicz, M.; Mobassaleh, M.; Donohue-Rolfe, A. Shiga Toxin: Intestinal Cell Receptors and Pathophysiology of Enterotoxic Effects. Rev. Infect. Dis. 1991, 13, S304–S310. [Google Scholar] [CrossRef]
- Acheson, D.W.; Moore, R.; De Breucker, S.; Lincicome, L.; Jacewicz, M.; Skutelsky, E.; Keusch, G.T. Translocation of Shiga toxin across polarized intestinal cells in tissue culture. Infect. Immun. 1996, 64, 3294–3300. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Feizi, T. The neoglycolipid (NGL) technology-based microarrays and future prospects. FEBS Lett. 2018, 592, 3976–3991. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Chai, W. Mucin O-glycan microarrays. Curr. Opin. Struct. Biol. 2019, 56, 187–197. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HMO Characteristics (Source) | [HMO] | Strains Used | Observed Effects | References |
|---|---|---|---|---|
| Breast milk collected from first and fourth week of lactation | 1:2 dilution | Gram-positive: Streptococcus pneumoniae Gram-negative: Haemophilus influenzae | Anti-adhesive effects against Haemophilus influenzae (HMWF) and Streptococcus pneumoniae (all HMOs) | [23] |
| Breast milk from healthy women collected 30 days after delivery | 6 mg/mL | Gram-negative: UPEC | Anti-adhesive effects of neutral fractions (high > low Mw) | [28] |
| Colostrum (d1–4), transitional (d12–17) and mature (d28–32) breast milk from healthy women | 25–1200 μg/well (50 μL) | Gram-negative: ETEC (CFA/I, CFA/II fimbriae), UPEC (P, P-like fimbriae) | Inhibition of hemagglutination by desialylated fraction associated with binding to P-fimbriae | [27] |
| Pooled transitional breast milk samples | 20 g/L | Gram-negative: Neisseria meningitidis | Inhibition of binding to pili by acidic HMO fraction | [35] |
| HMO fractions and modified HMO fractions from pooled human milk | 1–2 g/L | Isolated, immobilized P-selectin | Interference acidic HMO fraction and P-selectin. Neutral HMOs show no interference. | [41] |
| Colostrum (different fractions) collected four days after delivery | 1, 5, and 10 mg/mL | Gram-negative: EPEC, Vibrio cholerae, Salmonella fyris | Anti-adhesive effects against Salmonella fyris (acidic, neutral, LMw), and Vibrio cholerae (neutral, hMw) | [17] |
| HMOs isolated from pooled human milk | 15 mg/mL | Gram-negative: UPEC | Inhibition of bacterial invasion but no anti-adhesive effects, protection | [39] |
| Breast milk from healthy women collected between 3 days and 3 months postnatal | 5 mg/mL | Gram-positive: GBS | Up to 40% growth inhibition | [21] |
| Breast milk from healthy women collected between 3 days and 3 months postnatal | 5 mg/mL | Gram-positive: GBS (CNCTC, GB590, GB2) | 8–32× MIC reduction with antibiotics in combination with HMOs | [42] |
| Breast milk from healthy women collected between 3 days and 3 months postnatal | 5 mg/mL | Gram-positive: GBS, Staphylococcus aureus Gram-negative: Acinetobacter baumannii | GBS and Staphylococcus aureus biofilm inhibition, no antimicrobial effect | [43] |
| Breast milk from healthy women collected between 3 days and 3 months postnatal | 5 mg/mL | Gram-negative: GBS (GB590, GB2) | HMO mixture more effective inhibition of pathogen growth and viability reduction than isolated oligosaccharides | [44] |
| HMO Characteristics | [HMO] | Strains Used | Observed Effects | References |
|---|---|---|---|---|
| 2-FL, 3-FL, 3-SL, 6-SL | 2-FL = 2.5 mg/mL 3-FL = 0.5 mg/mL 3′-SL = 0.1 mg/mL 6′-SL = 0.3 mg/mL | EPEC, Vibrio cholerae, Salmonella fyris | Anti-adhesive effect of 6-SL and 3-FL against Escherichia coli and Salmonella fyris | [17] |
| Synthesized 2-FL, 3-FL | 10 mg/mL | Gram-negative: Campylobacter jejuni, EPEC, Salmonella enterica, Pseudomonas aeruginosa | Differential anti-adhesive effect | [47] |
| 3-SL and 6-SL | 2 μg/mL–1 mg/mL | Gram-negative: Pseudomonas aeruginosa | Dose-dependent inhibition by 6-SL of pneumocyte invasion (lung) | [52] |
| 3-SL, 6-SL, LNT, LSTa, LSTc, DSLNT | 5 mg/mL | Gram-positive: GBS | Inhibition of biofilm production and growth by larger sialylated oligosaccharides | [44] |
| 2-FL, 3-FL, DFL, LNT, LNnT, LNFP I, LNFP II, LNFP III, LNT II, para-LNnH, LNnH | 5 mg/mL | Gram-positive: GBS | Strain-specific antimicrobial activity, no biofilm inhibition, fucose not involved in antimicrobial function | [42] |
| 2-FL1-N-2-FL | - | Gram-positive: GBS | No antimicrobial or antibiofilm activity of 2-FL Antimicrobial/antibiofilm activity due to cationic moiety of 1-N-2-FL | [49] |
| AOS Characteristics | [AOS] | Strains Used | Observed Effects | References |
|---|---|---|---|---|
| GC: 90–95% (OligoG CF 5/20), 46%, 0% MW: 2.6 kDa | 2%, 6%, 10% | Gram-negative:Pseudomonas aeruginosa, Proteus mirabilis, Escherichia coli | Inhibition of motility and biofilm formation, antibiotic synergy | [63] |
| OligoG CF-5/20 | 2%, 6%, 10% | Gram-negative:Pseudomonas aeruginosa, Acinetobacterbaumannii | Structural interference biofilm formation, antibiotic synergy | [64] |
| OligoG CF-5/20 | 2%, 6%, 10% | Gram-negative:Porphyromonas gingivalis Gram-positive:Streptococcus mutans | Triclosan synergy | [70] |
| OligoG CF-5/20 | 0.2%–10% | Gram-negative:Pseudomonas aeruginosa, Burkholderia spp. | Inhibition of pathogenic cell motility | [65] |
| Alginate-derived oligosaccharides M/G: 2.28 MW: 300 kDa | 2 mg/mL | Gram-negative:Pseudomonas aeruginosa | Antibiotic synergy, anti-biofilm effect, decrease in virulence, increase in susceptibility to H2O2 of pathogen | [88] |
| OligoG CF-5/20 | 5–100 mg/mL | Gram-negative:Pseudomonas aeruginosa Gram-positive:Streptococcus mutans | Cellular aggregation of S. mutans and P. aeruginosa and binding of OligoG CF-5/20 to P. aeruginosa. Anti-microbial effects are not related to structural alterations in LPS or cell permeability | [67] |
| OligoG CF-5/20 | 2% | Gram-negative:Pseudomonas aeruginosa | Antibiotic synergy | [66] |
| OligoG CF-5/20 | - | Gram-negative:Pseudomonas aeruginosa | Inhibition of QS-signaling | [84] |
| OligoG CF-5/20 | 0.5%, 2%, 6% | Gram-negative:Pseudomonas aeruginosa | Disruptive effect on biofilm formation, established biofilm | [69] |
| COS Characteristics + Source | [COS] | Strains Used | Observed Effects | References |
|---|---|---|---|---|
| Chitosan oligosaccharides-from chitosan with DA 89%, DP 3–6 (805) | 0.01–0.5% | Gram-negative:Escherichia coli | Antibacterial activity (anti-growth) and 0.5% completely inhibited the growth of E. coli | [126] |
| Chito-oligosaccharides DA 8.5%, Mw 2–30 kDa | 0.10% | Gram-negative:Aggregatibacter actinomycetemcomitans Gram-positive:Streptococcus mutans | Pathogenic membrane disruption | [102] |
| Chito-oligosaccharides DA 11% Mw <10, <5, <1 kDa | 0.1–1% | Gram-negative:Escherichia coli, Salmonella typhimurium, Pseudomonas aeruginosa Gram-positive:Streptococcus mutans, Micrococcus luteus, Staphylococcus aureus, Staphylococcus epidermidis, Bacillus subtilis | Higher anti-microbial (anti-growth) effect high-Mw COS | [105] |
| Chitosan Mw 5, 8 kDa | 0.01–0.5% | Gram-negative:Escherichia coli | mRNA transcription inhibition | [121] |
| Chitosans (Mw = 1671, 1106, 746, 470, 224, and 28 kDa) Chitosan oligomers (Mw = 22, 10, 7, 4, 2, and 1 kDa) | 1% | Gram-negative:Escherichia coli, Pseudomonas fluorescens, Salmonella typhimurium, Vibrio parahaemolyticusGram-positive:Listeria monocytogenes, Bacillus megaterium, Bacillus cereus, Staphylococcus aureus | Higher anti-microbial effect (anti-growth) of chitosan compared to COS Chitosan showed stronger bactericidal effects for gram-positive bacteria than gram-negative bacteria | [93] |
| Chitosan Mw < 5 kDa | 0.25–1% | Gram-negative:Escherichia coli Gram-positive:Staphylococcus aureus | E. coli growth inhibition (lower Mw more effective) S. aureus growth inhibition (higher Mw more effective) | [94] |
| Chito-oligosaccharides DA 35.2–37.8% DP 1–6 | 0.1–0.5% | Gram-negative:Escherichia coli Gram-positive:Bacillus cereus | Growth inhibition and cell membrane disruption | [112] |
| Chito-oligosaccharides DA 3%, DP ~4 | 0.25–2.5% | Gram-negative:VTEC, EPEC, Desulfovibrio desulfuricans | Selective anti-adhesion properties | [116] |
| Chitosan oligosaccharides DA 98.8%, DP 1–16 | 0.0001–0.5% | Gram-positive:Staphylococcus aureus | Cell membrane lysis | [113] |
| Chito-oligosaccharides DA 15–20%, MW <5, <3 kDa | 1–5% | Gram-negative:Escherichia coli Gram-positive:Staphylococcus aureus | Antimicrobial effect (anti-growth) on Escherichia coli, but not Staphylococcus aureus | [95] |
| Chitosan DA > 90% | 0.0004–6.7% | Gram-negative:Pseudomonas aeruginosa (MDR) | Synergy with sulfamethoxazole treatment (anti-growth effect) | [125] |
| Chitosan oligosaccharides-Mw= 10,000 Da and 1000 Da)-from chitosan with DA 90–95% | 0.5–10 mg/mL | Gram-negative:Vibrio vulnificus | Higher antimicrobial effect (anti-growth) of water-soluble COS with high molecular weight | [127] |
| Chitosan DA >90% | 0.0004–6.7% | Gram-possitive:Staphylococcus aureus (MDR) | Synergy with several AB treatments (anti-growth effect) | [124] |
| Chitosans DA 80–85% Mw = 628, 591 and 107 kDa Chito-oligosaccharides DA 80–85% Mw = <5 and <3 kDa | 0.5% | Gram-negative:Escherichia coli, Pseudomonas aeruginosa, Klebsiella pneumoniae Gram-positive:Staphylococcus aureus, Staphylococcus epidermidis | Higher antimicrobial effect (anti-growth) of the 3 chitosans | [104] |
| Chitin (DA 35, Mw 388 Da) Chitosan (DA 80, Mw 12 Da) COS | 0.003–0.1% | Gram-negative:Escherichia coli, Pseudomonas aeruginosa, Salmonella typhimurium, Vibrio cholerae, Shigella dysenteriae, Enterobacter agglomerans, Prevotella melaninogenica, Bacteroides fragilis Gram-positive:Staphylococcus aureus, Bacillus subtilis, Bacillus cereus | Higher antimicrobial effect (anti-growth) COS compared to biopolymers | [114] |
| Chito-oligosaccharides DA ~65%, DP 3–5 | 2% | Gram-negative:Pseudomonas aeruginosa | Anti-growth, anti-biofilm functionalities and synergy with azithromycin | [88] |
| Chito-oligosaccharides DA 9–14%, DP <5–30 | 1–10% | Gram-negative:Escherichia coli Gram-positive:Listeria monocytogenes | High antimicrobial effect (anti-growth) with high DP | [103] |
| Chito-oligosaccharides-from chitosan with DD 80 and 90%-from chitosan with Mw = 5.1, 14.3 and 41.1 kDa | 0.002–0.064% | Gram-negative:Escherichia coli, Salmonella typhimurium, Salmonella enteritidis | High antimicrobial effect (anti-growth) with low DP, potent ferrous chelating activity at low DP | [128] |
| GOS Characteristics | [GOS] | Strains Used | Observed Effects | References |
|---|---|---|---|---|
| DP 3–7 | 0–32 mg/mL | Gram-negative: EPEC | Anti-adhesive effect | [138] |
| DP 3–6 | 1.56–100 mg/mL | Gram-negative: Vibrio cholerae | Anti-Ctx | [149] |
| DP 1–4 | 2.5 mg/mL | Gram-negative: Salmonella typhimurium | Anti-adhesive and anti-invasive effect | [139] |
| DP 2–6 | 20 mg/mL | Gram-positive: Listeria monocytogenes | No anti-adhesive and anti-growth effect | [141] |
| DP 3–6 | 16 mg/mL | Gram-negative: Cronobacter sakazakii | Anti-adhesive effect | [142] |
| - | 10–50 mg/mL | Gram-negative: Citrobacter rodentium | Anti-adhesive effect | [140] |
| MOS Characteristics | [MOS] | Strains Used | Observed Effects | References |
|---|---|---|---|---|
| DP 2–6 | 0.1–0.5 mM | Gram-negative:Escherichia coli | Anti-adhesive effect | [167] |
| DP 9 | 25 μM | Gram-negative:Enterobacter cloacae | Anti-adhesive effect | [168] |
| DP 3 MOS | 0.13 M–087 M | Gram-negative:Escherichia coli | Affinity for FimH mannose > MOS | [160] |
| Partially purified yeast MOS and soluble supernatant fraction of MOS | 10–50 mg/mL | Gram-negative:Campylobacter jejuni, Campylobacter coli | Anti-adhesive effects | [163] |
| Yeast MOS | 6 mg/mL | Gram-negative:Escherichia coli, Salmonella pullorum | Anti-adhesive effect (less effective than yeast cell walls) | [166] |
| POS Characteristics + Source | [POS] | Strains Used | Observed Effects | References |
|---|---|---|---|---|
| Mw 1–4 kDa Citrus (high methylation) Apple (low methylation) | 10 mg/mL | Gram-negative:Escherichia coli O157:H7 Shiga toxin | Inhibition of host cell infiltration of Stx | [185] |
| Mw 1–12 kDa Panax ginseng | 0.01–0.5 mg/mL | Gram-negative:Aggregatibacter actinomycetemcomitans Gram-positive:Staphylococcus aureus | Anti-adhesive effect | [189] |
| DP 2–3 Orange peel | 2.5 mg/mL | Gram-negative: EPEC, VTEC, Desulfovibrio desulfuricans | Anti-adhesive effect | [186] |
| GalA:Rhamnose 1:1 Albedo of orange peel | 0.05–2.5 mg/mL | Gram-negative:Campylobacter jejuni | Inhibited Caco-2 cell invasion | [177] |
| 0.2–6 kDa 93.6% Uronic acid Haw | 1–10 mg/mL | Gram-negative:Escherichia coli | Antimicrobial activity dependent on concentration and low pH | [178] |
| DP 6–19 Orange peel | 1–100 mg/mL | Gram-negative:Escherichia coli Gram-positive:Staphylococcus aureus, Bacillus subtilis | Antimicrobial activity-low Mw more effective | [176] |
| Apple, citrus, polygalacturonic acid | 0.1 mg/mL | Gram-negative:Escherichia coli Gram-positive:Staphylococcus aureus | Growth inhibition, potentially through CO2 radical production | [184] |
| Mw 9–73 kDa Orange peel | 0.005–5 mg/mL | Gram-negative: Shigatoxigenic Escherichia coli | Anti-adhesive effect, direct interaction with Stx | [188] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Asadpoor, M.; Peeters, C.; Henricks, P.A.J.; Varasteh, S.; Pieters, R.J.; Folkerts, G.; Braber, S. Anti-Pathogenic Functions of Non-Digestible Oligosaccharides In Vitro. Nutrients 2020, 12, 1789. https://doi.org/10.3390/nu12061789
Asadpoor M, Peeters C, Henricks PAJ, Varasteh S, Pieters RJ, Folkerts G, Braber S. Anti-Pathogenic Functions of Non-Digestible Oligosaccharides In Vitro. Nutrients. 2020; 12(6):1789. https://doi.org/10.3390/nu12061789
Chicago/Turabian StyleAsadpoor, Mostafa, Casper Peeters, Paul A. J. Henricks, Soheil Varasteh, Roland J. Pieters, Gert Folkerts, and Saskia Braber. 2020. "Anti-Pathogenic Functions of Non-Digestible Oligosaccharides In Vitro" Nutrients 12, no. 6: 1789. https://doi.org/10.3390/nu12061789
APA StyleAsadpoor, M., Peeters, C., Henricks, P. A. J., Varasteh, S., Pieters, R. J., Folkerts, G., & Braber, S. (2020). Anti-Pathogenic Functions of Non-Digestible Oligosaccharides In Vitro. Nutrients, 12(6), 1789. https://doi.org/10.3390/nu12061789