Beyond Heat Stress: Intestinal Integrity Disruption and Mechanism-Based Intervention Strategies

 , ,
, ,
Abstract
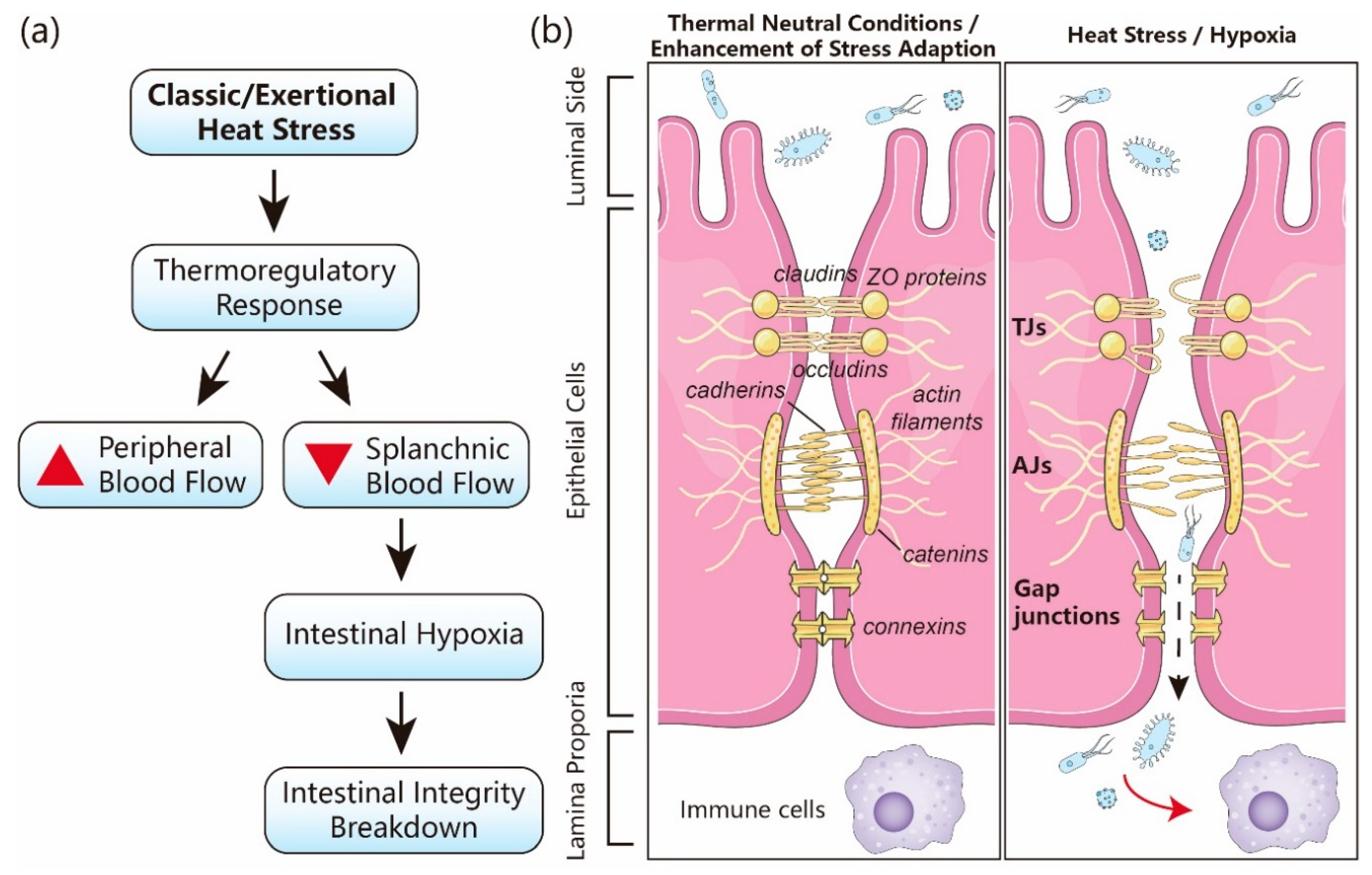
1. Introduction
2. Stress Adaptation Signaling Pathways
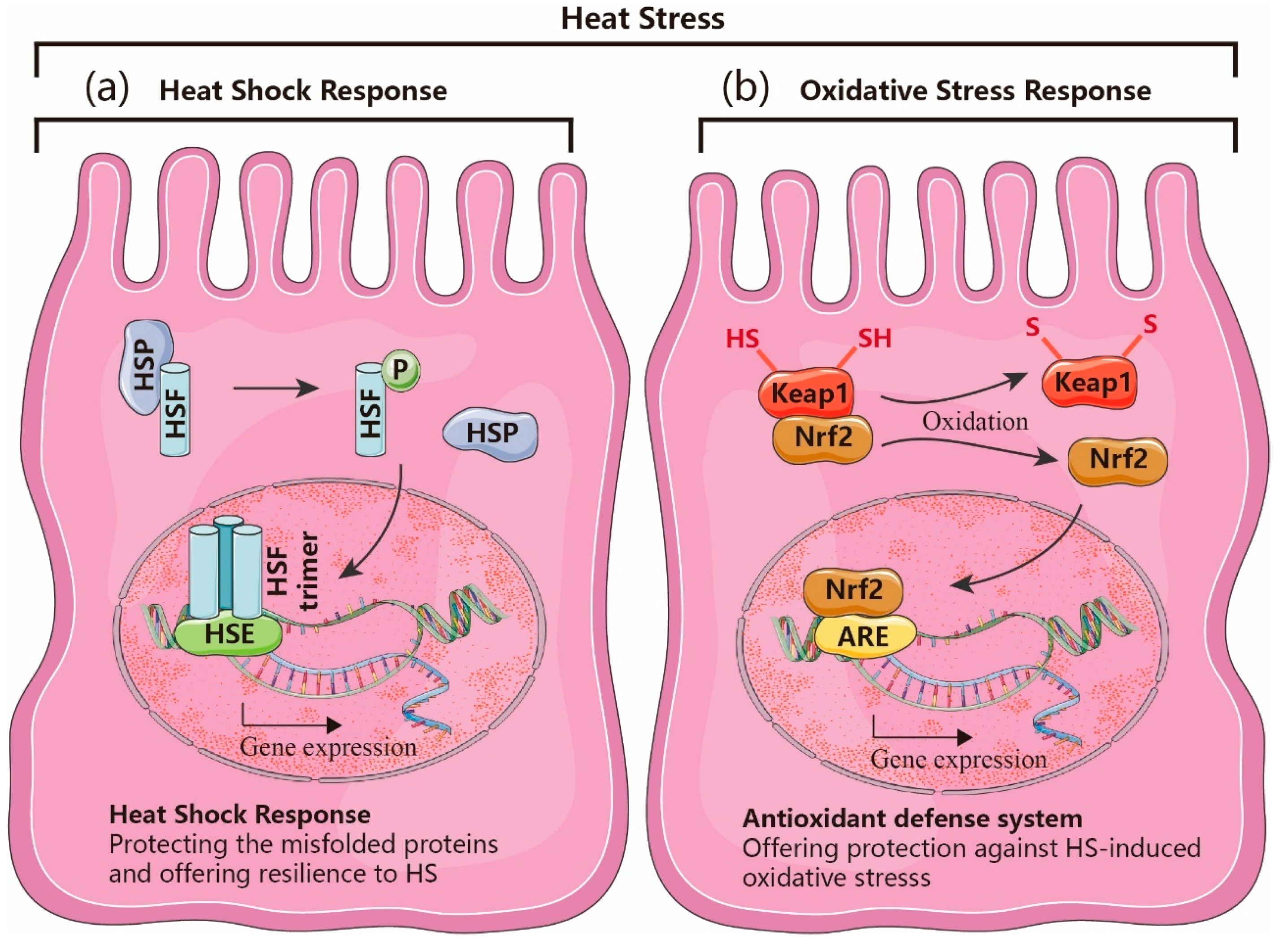
2.1. Heat Shock Response (HSR)
2.2. HSR and Oxidative Stress Response
2.3. Resilience Pathways and Intestinal Barrier Integrity
2.4. Resilience Pathways and Immune System
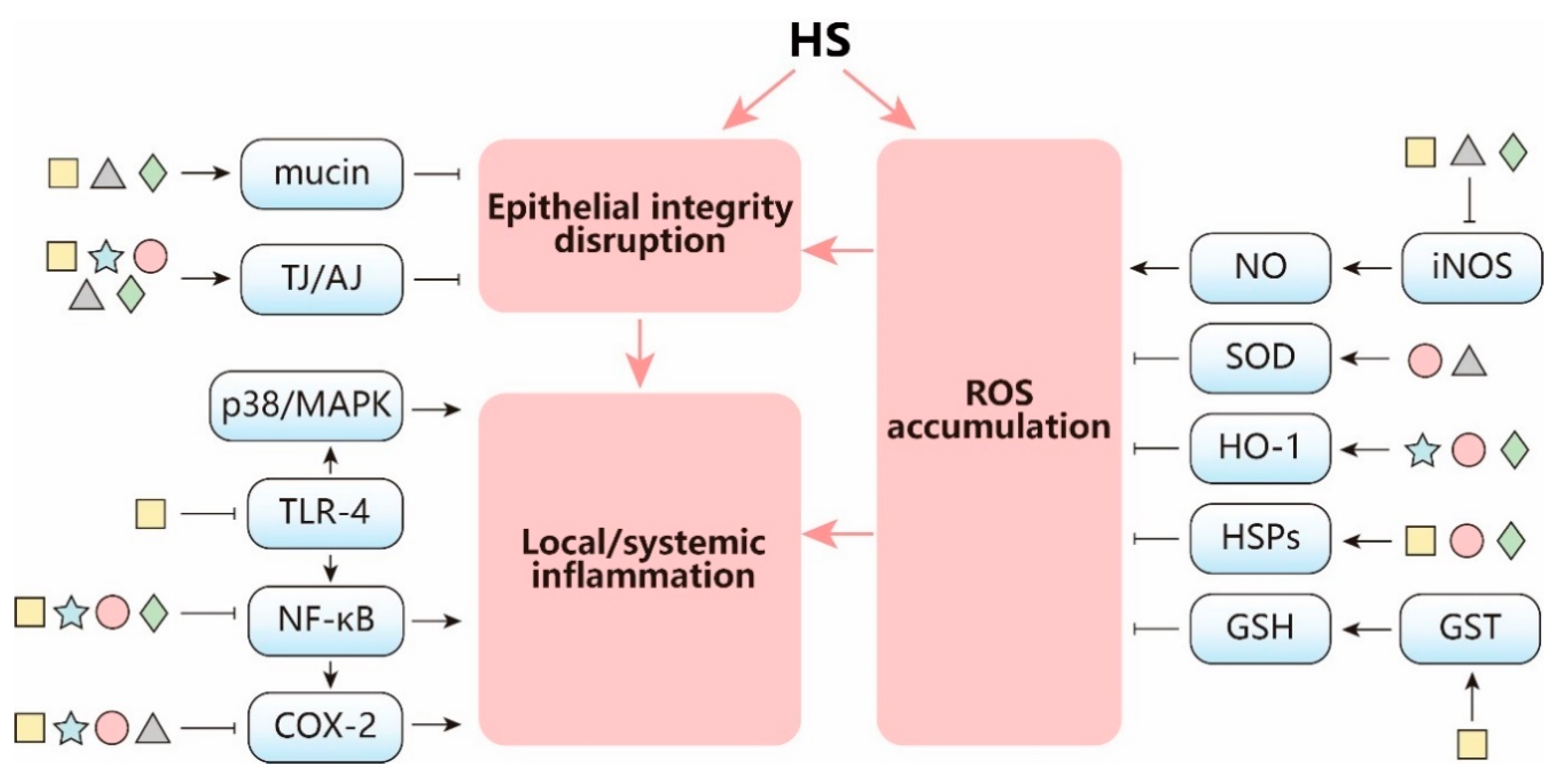
3. Intervention Strategies against HS
3.1. Microbiota Modulation
3.1.1. Probiotics
3.1.2. Prebiotics
3.2. Antioxidants
3.2.1. α-Lipoic Acid (ALA), a Fatty Acid with Antioxidant Properties
3.2.2. Resveratrol, a Plant Polyphenol Compound
3.3. Polyunsaturated Fatty Acids (PUFA)
3.4. Amino Acids
3.4.1. Arginine
- 1.
- The NO synthesis pathway: Arginine, as a precursor of NO production, stimulates the enzyme NO synthase (NOS) isoforms to facilitate the synthesis and bioavailability of NO [211]. The constitutive form of NOS (cNOS), which includes endothelial NOS (eNOS) and neuronal NOS (nNOS), generates relatively small amounts of NO, while iNOS produces a quantitatively larger amount of NO and is expressed in cells of the immune system as well as in intestinal epithelial cells [186,212,213].
- 2.
- Mammalian target of rapamycin (mTOR) pathway: Maintaining the intestinal epithelial function by arginine can also be related to activation of the mTOR pathway [196,218]. Arginine induces the downstream mTOR pathway by phosphorylation and activation of the protein synthesis regulator 70-kDa ribosomal protein S6 kinase (p70S6k) [219]. Activation of p70S6k by arginine increases protein synthesis, proliferation, and migration in disease conditions that induce intestinal epithelial injury [200]. For instance, oral administration of arginine in a porcine model of enteritis augments intestinal protein synthesis and attenuates intestinal permeability via mTOR signaling and p70S6k activation [218].
- 3.
- Arginase pathway: Metabolism of arginine via the arginase pathway results in the production of ornithine and polyamine, which promote intestinal epithelial repair and restitution processes [196]. Polyamines are involved in the regulation of cell-cell interactions and E-cadherin expression, being critically important for the maintenance of intestinal epithelial integrity [220]. Additionally, polyamines are important stress-responsive molecules, which facilitate the activation of HSF1 to induce HSP expression [221,222].
3.4.2. Glutamine
4. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Furness, J.B.; Kunze, W.A.; Clerc, N. Nutrient tasting and signaling mechanisms in the gut. II. The intestine as a sensory organ: Neural, endocrine, and immune responses. Am. J. Physiol. 1999, 277, G922–G928. [Google Scholar] [PubMed]
- Lambert, G.P. Stress-induced gastrointestinal barrier dysfunction and its inflammatory effects. J. Anim. Sci. 2009, 87, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Peterson, L.W.; Artis, D. Intestinal epithelial cells: Regulators of barrier function and immune homeostasis. Nat. Rev. Immunol. 2014, 14, 141–153. [Google Scholar] [CrossRef]
- Bouchama, A.; Knochel, J.P. Heat stroke. N. Engl. J. Med. 2002, 346, 1978–1988. [Google Scholar] [CrossRef]
- Leon, L.R.; Helwig, B.G. Heat stroke: Role of the systemic inflammatory response. J. Appl. Physiol. 2010, 109, 1980–1988. [Google Scholar] [CrossRef]
- Epstein, Y.; Roberts, W.O. The pathopysiology of heat stroke: An integrative view of the final common pathway. Scand. J. Med. Sci. Sports 2011, 21, 742–748. [Google Scholar] [CrossRef]
- Littmann, A.E.; Shields, R.K. Whole body heat stress increases motor cortical excitability and skill acquisition in humans. Clin. Neurophysiol. 2016, 127, 1521–1529. [Google Scholar] [CrossRef]
- Fan, H.; Zhao, Y.; Zhu, J.-H.; Song, F.-C.; Ye, J.-H.; Wang, Z.-Y.; Le, J.-W. Thrombocytopenia as a predictor of severe acute kidney injury in patients with heat stroke. Ren. Fail. 2015, 37, 877–881. [Google Scholar] [CrossRef]
- Heneghan, H.M.; Nazirawan, F.; Dorcaratto, D.; Fiore, B.; Boylan, J.F.; Maguire, D.; Hoti, E. Extreme heatstroke causing fulminant hepatic failure requiring liver transplantation: A case report. Transplant. Proc. 2014, 46, 2430–2432. [Google Scholar] [CrossRef] [PubMed]
- Welc, S.S.; Clanton, T.L.; Dineen, S.M.; Leon, L.R. Heat stroke activates a stress-induced cytokine response in skeletal muscle. J. Appl. Physiol. 2013, 115, 1126–1137. [Google Scholar] [CrossRef] [PubMed]
- Lambert, G.P.; Gisolfi, C.V.; Berg, D.J.; Moseley, P.L.; Oberley, L.W.; Kregel, K.C. Selected contribution: Hyperthermia-induced intestinal permeability and the role of oxidative and nitrosative stress. J. Appl. Physiol. 2002, 92, 1750–1761. [Google Scholar] [CrossRef] [PubMed]
- Yamoto, M.; Lee, C.; Chusilp, S.; Yazaki, Y.; Alganabi, M.; Li, B.; Pierro, A. The role of autophagy in intestinal epithelial injury. Pediatr. Surg. Int. 2019, 35, 1389–1394. [Google Scholar] [CrossRef] [PubMed]
- Shah, Y.M. The role of hypoxia in intestinal inflammation. Mol. Cell. Pediatr. 2016, 3, 1. [Google Scholar] [CrossRef] [PubMed]
- Vissenaekens, H.; Grootaert, C.; Rajkovic, A.; Van De Wiele, T.; Calatayud, M. The response of five intestinal cell lines to anoxic conditions in vitro. Biol. Cell 2019, 111, 232–244. [Google Scholar] [CrossRef]
- Belhadj Slimen, I.; Najar, T.; Ghram, A.; Abdrrabba, M. Heat stress effects on livestock: Molecular, cellular and metabolic aspects, a review. J. Anim. Physiol. Anim. Nutr. 2016, 100, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Settar, P.; Yalçin, S.; Türkmut, L.; Ozkan, S.; Cahanar, A. Season by genotype interaction related to broiler growth rate and heat tolerance. Poult. Sci. 1999, 78, 1353–1358. [Google Scholar] [CrossRef] [PubMed]
- Deeb, N.; Lamont, S.J. Genetic architecture of growth and body composition in unique chicken populations. J. Hered. 2002, 93, 107–118. [Google Scholar] [CrossRef]
- Tsiouris, V.; Georgopoulou, I.; Batzios, C.; Pappaioannou, N.; Ducatelle, R.; Fortomaris, P. Heat stress as a predisposing factor for necrotic enteritis in broiler chicks. Avian Pathol. 2018, 47, 616–624. [Google Scholar] [CrossRef]
- Zhu, L.; Liao, R.; Wu, N.; Zhu, G.; Yang, C. Heat stress mediates changes in fecal microbiome and functional pathways of laying hens. Appl. Microbiol. Biotechnol. 2019, 103, 461–472. [Google Scholar] [CrossRef]
- Akerfelt, M.; Morimoto, R.I.; Sistonen, L. Heat shock factors: Integrators of cell stress, development and lifespan. Nat. Rev. Mol. Cell Biol. 2010, 11, 545–555. [Google Scholar] [CrossRef]
- Joly, A.-L.; Wettstein, G.; Mignot, G.; Ghiringhelli, F.; Garrido, C. Dual role of heat shock proteins as regulators of apoptosis and innate immunity. J. Innate Immun. 2010, 2, 238–247. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-J.; Ock, C.Y.; Kim, S.-J.; Hahm, K.-B. Heat shock protein: Hard worker or bad offender for gastric diseases. Int. J. Proteom. 2010, 2010, 259163. [Google Scholar] [CrossRef] [PubMed]
- Fulda, S.; Gorman, A.M.; Hori, O.; Samali, A. Cellular stress responses: Cell survival and cell death. Int. J. Cell Biol. 2010, 2010, 214074. [Google Scholar] [CrossRef] [PubMed]
- Akbarian, A.; Michiels, J.; Degroote, J.; Majdeddin, M.; Golian, A.; De Smet, S. Association between heat stress and oxidative stress in poultry; mitochondrial dysfunction and dietary interventions with phytochemicals. J. Anim. Sci. Biotechnol. 2016, 7, 37. [Google Scholar] [CrossRef]
- Superoxide radical production in chicken skeletal muscle induced by acute heat stress. Poult. Sci. 2005, 84, 307–314. [CrossRef]
- Christen, F.; Desrosiers, V.; Dupont-Cyr, B.A.; Vandenberg, G.W.; Le François, N.R.; Tardif, J.C.; Dufresne, F.; Lamarre, S.G.; Blier, P.U. Thermal tolerance and thermal sensitivity of heart mitochondria: Mitochondrial integrity and ROS production. Free Radic. Biol. Med. 2018, 116, 11–18. [Google Scholar] [CrossRef]
- King, M.A.; Clanton, T.L.; Laitano, O. Hyperthermia, dehydration and osmotic stress: Unconventional sources of exercise-induced reactive oxygen species. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 310, 105–114. [Google Scholar] [CrossRef]
- de Roos, B.; Duthie, G.G. Role of dietary pro-oxidants in the maintenance of health and resilience to oxidative stress. Mol. Nutr. Food Res. 2015, 59, 1229–1248. [Google Scholar] [CrossRef]
- Kovac, S.; Angelova, P.R.; Holmström, K.M.; Zhang, Y.; Dinkova-Kostova, A.T.; Abramov, A.Y. Nrf2 regulates ROS production by mitochondria and NADPH oxidase. Biochim. Biophys. Acta 2015, 1850, 794–801. [Google Scholar] [CrossRef]
- Ivan, M.; Kondo, K.; Yang, H.; Kim, W.; Valiando, J.; Ohh, M.; Salic, A.; Asara, J.M.; Lane, W.S.; Kaelin, J. HIFα targeted for VHL-mediated destruction by proline hydroxylation: Implications for O2 sensing. Science 2001, 292, 464–468. [Google Scholar] [CrossRef]
- Guzy, R.D.; Mack, M.M.; Schumacker, P.T. Mitochondrial complex III is required for hypoxia-induced ROS production and gene transcription in yeast. Antioxid. Redox Signal. 2007, 9, 1317–1328. [Google Scholar] [CrossRef] [PubMed]
- Chandel, N.S.; McClintock, D.S.; Feliciano, C.E.; Wood, T.M.; Melendez, J.A.; Rodriguez, A.M.; Schumacker, P.T. Reactive oxygen species generated at mitochondrial Complex III stabilize hypoxia-inducible factor-1α during hypoxia: A mechanism of O2 sensing. J. Biol. Chem. 2000, 275, 25130–25138. [Google Scholar] [CrossRef] [PubMed]
- Dokladny, K.; Zuhl, M.N.; Moseley, P.L. Intestinal epithelial barrier function and tight junction proteins with heat and exercise. J. Appl. Physiol. 2016, 120, 692–701. [Google Scholar] [CrossRef]
- Dokladny, K.; Ye, D.; Kennedy, J.C.; Moseley, P.L.; Ma, T.Y. Cellular and molecular mechanisms of heat stress-induced up-regulation of occludin protein expression: Regulatory role of heat shock factor-1. Am. J. Pathol. 2008, 172, 659–670. [Google Scholar] [CrossRef] [PubMed]
- Varasteh, S.; Braber, S.; Kraneveld, A.D.; Garssen, J.; Fink-Gremmels, J. l-Arginine supplementation prevents intestinal epithelial barrier breakdown under heat stress conditions by promoting nitric oxide synthesis. Nutr. Res. 2018, 57, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Zuhl, M.; Schneider, S.; Lanphere, K.; Conn, C.; Dokladny, K.; Moseley, P. Exercise regulation of intestinal tight junction proteins. Br. J. Sports Med. 2014, 48, 980–986. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.-C.; He, S.-H.; Zheng, P.-Y. Investigation into the signal transduction pathway via which heat stress impairs intestinal epithelial barrier function. J. Gastroenterol. Hepatol. 2007, 22, 1823–1831. [Google Scholar] [CrossRef] [PubMed]
- Tsapara, A.; Matter, K.; Balda, M.S. The heat-shock protein Apg-2 binds to the tight junction protein ZO-1 and regulates transcriptional activity of ZONAB. Mol. Biol. Cell 2006, 17, 1322–1330. [Google Scholar] [CrossRef]
- Bidmon-Fliegenschnee, B.; Lederhuber, H.C.; Csaicsich, D.; Pichler, J.; Herzog, R.; Memaran-Dadgar, N.; Huber, W.-D.; Aufricht, C.; Kratochwill, K. Overexpression of Hsp70 confers cytoprotection during gliadin exposure in Caco-2 cells. Pediatr. Res. 2015, 78, 358–364. [Google Scholar] [CrossRef]
- Jin, W.; Wang, H.; Ji, Y.; Hu, Q.; Yan, W.; Chen, G.; Yin, H. Increased intestinal inflammatory response and gut barrier dysfunction in Nrf2-deficient mice after traumatic brain injury. Cytokine 2008, 44, 135–140. [Google Scholar] [CrossRef]
- Singh, R.; Chandrashekharappa, S.; Bodduluri, S.R.; Baby, B.V.; Hegde, B.; Kotla, N.G.; Hiwale, A.A.; Saiyed, T.; Patel, P.; Vijay-Kumar, M.; et al. Enhancement of the gut barrier integrity by a microbial metabolite through the Nrf2 pathway. Nat. Commun. 2019, 10, 89. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Bao, Z.; Xu, X.; Chao, H.; Lin, C.; Li, Z.; Liu, Y.; Wang, X.; You, Y.; Liu, N.; et al. Extracellular signal-regulated kinase/nuclear factor-erythroid2-like2/heme oxygenase-1 pathway-mediated mitophagy alleviates traumatic brain injury-induced intestinal mucosa damage and epithelial barrier dysfunction. J. Neurotrauma 2017, 34, 2119–2131. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Hu, Y.; Fang, Y.; Djukic, Z.; Yamamoto, M.; Shaheen, N.J.; Orlando, R.C.; Chen, X. Nrf2 deficiency impairs the barrier function of mouse oesophageal epithelium. Gut 2014, 63, 711–719. [Google Scholar] [CrossRef]
- Samborski, P.; Grzymisławski, M. The role of HSP70 heat shock proteins in the pathogenesis and treatment of inflammatory bowel diseases. Adv. Clin. Exp. Med. 2015, 24, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Sziksz, E.; Veres, G.; Vannay, A.; Prókai, A.; Gál, K.; Onody, A.; Korponay-Szabó, I.R.; Reusz, G.; Szabó, A.; Tulassay, T.; et al. Increased heat shock protein 72 expression in celiac disease. J. Pediatr. Gastroenterol. Nutr. 2010, 51, 573–578. [Google Scholar] [CrossRef] [PubMed]
- Pritts, T.A.; Wang, Q.; Sun, X.; Moon, M.R.; Fischer, D.R.; Fischer, J.E.; Wong, H.R.; Hasselgren, P.O. Induction of the stress response in vivo decreases nuclear factor-kappa B activity in jejunal mucosa of endotoxemic mice. Arch. Surg. 2000, 135, 860–866. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Tu, Z.; Tang, D.; Zhang, H.; Liu, M.; Wang, K.; Calderwood, S.K.; Xiao, X. The inhibition of LPS-induced production of inflammatory cytokines by HSP70 involves inactivation of the NF-kappaB pathway but not the MAPK pathways. Shock 2006, 26, 277–284. [Google Scholar] [CrossRef]
- van Eden, W. Diet and the anti-inflammatory effect of heat shock proteins. Endocr. Metab. Immune Disord. Drug Targets 2015, 15, 31–36. [Google Scholar] [CrossRef]
- van Puijvelde, G.H.M.; van Es, T.; van Wanrooij, E.J.A.; Habets, K.L.L.; de Vos, P.; van der Zee, R.; van Eden, W.; van Berkel, T.J.C.; Kuiper, J. Induction of oral tolerance to HSP60 or an HSP60-peptide activates T cell regulation and reduces atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 2677–2683. [Google Scholar] [CrossRef]
- Maron, R.; Sukhova, G.; Faria, A.-M.; Hoffmann, E.; Mach, F.; Libby, P.; Weiner, H.L. Mucosal administration of heat shock protein-65 decreases atherosclerosis and inflammation in aortic arch of low-density lipoprotein receptor-deficient mice. Circulation 2002, 106, 1708–1715. [Google Scholar] [CrossRef]
- Hauet-Broere, F.; Wieten, L.; Guichelaar, T.; Berlo, S.; van der Zee, R.; Van Eden, W. Heat shock proteins induce T cell regulation of chronic inflammation. Ann. Rheum. Dis. 2006, 65, iii65–iii68. [Google Scholar] [CrossRef] [PubMed]
- Borges, T.J.; Wieten, L.; Van Herwijnen, M.J.C.; Broere, F.; Van derZee, R.; Bonorino, C.; Van Eden, W. The anti-inflammatory mechanisms of Hsp70. Front. Immunol. 2012, 3, 95. [Google Scholar] [CrossRef] [PubMed]
- Spierings, J.; van Eden, W. Heat shock proteins and their immunomodulatory role in inflammatory arthritis. Rheumatology 2017, 56, 198–208. [Google Scholar] [CrossRef] [PubMed]
- Morgan, M.J.; Liu, Z. Crosstalk of reactive oxygen species and NF-κB signaling. Cell Res. 2011, 21, 103–115. [Google Scholar] [CrossRef]
- Dayalan Naidu, S.; Kostov, R.V.; Dinkova-Kostova, A.T. Transcription factors Hsf1 and Nrf2 engage in crosstalk for cytoprotection. Trends Pharmacol. Sci. 2015, 36, 6–14. [Google Scholar] [CrossRef]
- Hensen, S.M.M.; Heldens, L.; Van Genesen, S.T.; Pruijn, G.J.M.; Lubsen, N.H. A delayed antioxidant response in heat-stressed cells expressing a non-DNA binding HSF1 mutant. Cell Stress Chaperones 2013, 18, 455–473. [Google Scholar] [CrossRef]
- Zagorski, J.W.; Turley, A.E.; Freeborn, R.A.; VanDenBerg, K.R.; Dover, H.E.; Kardell, B.R.; Liby, K.T.; Rockwell, C.E. Differential effects of the Nrf2 activators tBHQ and CDDO-Im on the early events of T cell activation. Biochem. Pharmacol. 2018, 147, 67–76. [Google Scholar] [CrossRef]
- Turley, A.E.; Zagorski, J.W.; Rockwell, C.E. The Nrf2 activator tBHQ inhibits T cell activation of primary human CD4 T cells. Cytokine 2015, 71, 289–295. [Google Scholar] [CrossRef]
- Awuh, J.A.; Haug, M.; Mildenberger, J.; Marstad, A.; Do, C.P.N.; Louet, C.; Stenvik, J.; Steigedal, M.; Damås, J.K.; Halaas, Ø.; et al. Keap1 regulates inflammatory signaling in Mycobacterium avium -infected human macrophages. Proc. Natl. Acad. Sci. USA 2015, 112, E4272–E4280. [Google Scholar] [CrossRef]
- Mach, N.; Fuster-Botella, D. Endurance exercise and gut microbiota: A review. J. Sport Health Sci. 2017, 6, 179–197. [Google Scholar] [CrossRef]
- Suzuki, K.; Harasawa, R.; Yoshitake, Y.; Mitsuoka, T. Effects of crowding and heat stress on intestinal flora, body weight gain, and feed efficiency of growing rats and chicks. Jpn. J. Vet. Sci. 1983, 45, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Johnson-Henry, K.C.; Abrahamsson, T.R.; Wu, R.Y.; Sherman, P.M. Probiotics, Prebiotics, and synbiotics for the prevention of necrotizing enterocolitis. Adv. Nutr. 2016, 7, 928–937. [Google Scholar] [CrossRef] [PubMed]
- Farag, M.R.; Alagawany, M. Physiological alterations of poultry to the high environmental temperature. J. Therm. Biol. 2018, 76, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Ribet, D.; Cossart, P. How bacterial pathogens colonize their hosts and invade deeper tissues. Microbes Infect. 2015, 17, 173–183. [Google Scholar] [CrossRef]
- Ducray, H.A.G.; Globa, L.; Pustovyy, O.; Vodyanoy, V.; Sorokulova, I. Prebiotics and probiotics maintain the intestinal barrier function. FASEB J. 2019, 33, 589.7. [Google Scholar]
- Shokryazdan, P.; Faseleh Jahromi, M.; Navidshad, B.; Liang, J.B. Effects of prebiotics on immune system and cytokine expression. Med. Microbiol. Immunol. 2017, 206, 1–9. [Google Scholar] [CrossRef]
- Quigley, E.M.M. Prebiotics and probiotics in digestive health. Clin. Gastroenterol. Hepatol. 2019, 17, 333–344. [Google Scholar] [CrossRef]
- Wegh, C.A.M.; Geerlings, S.Y.; Knol, J.; Roeselers, G.; Belzer, C. Postbiotics and their potential applications in early life nutrition and beyond. Int. J. Mol. Sci. 2019, 20, 4673. [Google Scholar] [CrossRef]
- Reid, G. Probiotics: Definition, scope and mechanisms of action. Best Pract. Res. Clin. Gastroenterol. 2016, 30, 17–25. [Google Scholar] [CrossRef]
- Shing, C.M.; Peake, J.M.; Lim, C.L.; Briskey, D.; Walsh, N.P.; Fortes, M.B.; Ahuja, K.D.K.; Vitetta, L. Effects of probiotics supplementation on gastrointestinal permeability, inflammation and exercise performance in the heat. Eur. J. Appl. Physiol. 2014, 114, 93–103. [Google Scholar] [CrossRef]
- Shukla, G.; Sidhu, R.K. Lactobacillus casei as a probiotic in malnourished Giardia lamblia -infected mice: A biochemical and histopathological study. Can. J. Microbiol. 2011, 57, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Rowland, I.R.; Capurso, L.; Collins, K.; Cummings, J.; Delzenne, N.; Goulet, O.; Guarner, F.; Marteau, P.; Meier, R. Current level of consensus on probiotic science-Report of an expert meeting—London, 23 November 2009. Gut Microbes 2010, 1, 436–439. [Google Scholar] [CrossRef] [PubMed]
- Rao, R.K.; Samak, G. Protection and restitution of gut barrier by probiotics: Nutritional and clinical implications. Curr. Nutr. Food Sci. 2013, 9, 99–107. [Google Scholar] [PubMed]
- Lamprecht, M.; Bogner, S.; Schippinger, G.; Steinbauer, K.; Fankhauser, F.; Hallstroem, S.; Schuetz, B.; Greilberger, J.F. Probiotic supplementation affects markers of intestinal barrier, oxidation, and inflammation in trained men; a randomized, double-blinded, placebo-controlled trial. J. Int. Soc. Sports Nutr. 2012, 9, 45. [Google Scholar] [CrossRef] [PubMed]
- Barnett, A.; Roy, N.; Cookson, A.; McNabb, W. Metabolism of caprine milk carbohydrates by probiotic bacteria and caco-2:HT29–MTX epithelial co-cultures and their impact on intestinal barrier integrity. Nutrients 2018, 10, 949. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, S.; Zaneb, H.; Yousaf, M.S.; Ijaz, A.; Sohail, M.U.; Muti, S.; Usman, M.M.; Ijaz, S.; Rehman, H. Effect of dietary supplementation of prebiotics and probiotics on intestinal microarchitecture in broilers reared under cyclic heat stress. J. Anim. Physiol. Anim. Nutr. 2013, 97, 68–73. [Google Scholar] [CrossRef]
- Song, J.; Xiao, K.; Ke, Y.L.; Jiao, L.F.; Hu, C.H.; Diao, Q.Y.; Shi, B.; Zou, X.T. Effect of a probiotic mixture on intestinal microflora, morphology, and barrier integrity of broilers subjected to heat stress. Poulty Sci. 2014, 93, 581–588. [Google Scholar] [CrossRef]
- Lee, N.K.; Kim, W.S.; Paik, H.D. Bacillus strains as human probiotics: Characterization, safety, microbiome, and probiotic carrier. Food Sci. Biotechnol. 2019, 28, 1297–1305. [Google Scholar] [CrossRef]
- Rajput, I.R.; Li, L.; Xin, X.; Wu, B.; Juan, Z.; Cui, Z.; Yu, D.; Li, W. Effect of Saccharomyces boulardii and Bacillus subtilis B10 on intestinal ultrastructure modulation and mucosal immunity development mechanism in broiler chickens. Poult. Sci. 2013, 92, 956–965. [Google Scholar] [CrossRef]
- Moore, T.; Globa, L.; Pustovyy, O.; Vodyanoy, V.; Sorokulova, I. Oral administration of Bacillus subtilis strain BSB3 can prevent heat stress-related adverse effects in rats. J. Appl. Microbiol. 2014, 117, 1463–1471. [Google Scholar] [CrossRef]
- Zyrek, A.A.; Cichon, C.; Helms, S.; Enders, C.; Sonnenborn, U.; Schmidt, M.A. Molecular mechanisms underlying the probiotic effects of Escherichia coli Nissle 1917 involve ZO-2 and PKCzeta redistribution resulting in tight junction and epithelial barrier repair. Cell. Microbiol. 2007, 9, 804–816. [Google Scholar] [CrossRef] [PubMed]
- Hummel, S.; Veltman, K.; Cichon, C.; Sonnenborn, U.; Schmidt, M.A. Differential targeting of the E-Cadherin/β-Catenin complex by gram-positive probiotic lactobacilli improves epithelial barrier function. Appl. Environ. Microbiol. 2012, 78, 1140–1147. [Google Scholar] [CrossRef] [PubMed]
- Segawa, S.; Fujiya, M.; Konishi, H.; Ueno, N.; Kobayashi, N.; Shigyo, T.; Kohgo, Y. Probiotic-derived polyphosphate enhances the epithelial barrier function and maintains intestinal homeostasis through integrin-p38 MAPK pathway. PLoS ONE 2011, 6, e23278. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, K.; Barrett, K.E.; Silvia, C.R.-L. Probiotic inhibition of the entry of enteroinvasive E. coli into, human intestinal epithelial cells involves both Rho-dependent and -independent pathways. Gastroenterology 2003, 124, A106. [Google Scholar] [CrossRef]
- Deng, W.; Dong, X.F.; Tong, J.M.; Zhang, Q. The probiotic Bacillus licheniformis ameliorates heat stress-induced impairment of egg production, gut morphology, and intestinal mucosal immunity in laying hens. Poult. Sci. 2012, 91, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Giorgetti, G.; Brandimarte, G.; Fabiocchi, F.; Ricci, S.; Flamini, P.; Sandri, G.; Trotta, M.C.; Elisei, W.; Penna, A.; Lecca, P.G.; et al. Interactions between innate immunity, microbiota, and probiotics. J. Immunol. Res. 2015, 2015, 501361. [Google Scholar] [CrossRef] [PubMed]
- Viladomiu, M.; Hontecillas, R.; Yuan, L.; Lu, P.; Bassaganya-Riera, J. Nutritional protective mechanisms against gut inflammation. J. Nutr. Biochem. 2013, 24, 929–939. [Google Scholar] [CrossRef]
- Zakostelska, Z.; Kverka, M.; Klimesova, K.; Rossmann, P.; Mrazek, J.; Kopecny, J.; Hornova, M.; Srutkova, D.; Hudcovic, T.; Ridl, J.; et al. Lysate of probiotic Lactobacillus casei DN-114 001 ameliorates colitis by strengthening the gut barrier function and changing the gut microenvironment. PLoS ONE 2011, 6, e27961. [Google Scholar] [CrossRef]
- Jeon, S.G.; Kayama, H.; Ueda, Y.; Takahashi, T.; Asahara, T.; Tsuji, H.; Tsuji, N.M.; Kiyono, H.; Ma, J.S.; Kusu, T.; et al. Probiotic Bifidobacterium breve induces IL-10-producing Tr1 cells in the colon. PLoS Pathog. 2012, 8, e1002714. [Google Scholar] [CrossRef]
- Hardy, H.; Harris, J.; Lyon, E.; Beal, J.; Foey, A. Probiotics, prebiotics and immunomodulation of gut mucosal defences: Homeostasis and immunopathology. Nutrients 2013, 5, 1869–1912. [Google Scholar] [CrossRef]
- Yahfoufi, N.; Mallet, J.; Graham, E.; Matar, C. Role of probiotics and prebiotics in immunomodulation. Curr. Opin. Food Sci. 2018, 20, 82–91. [Google Scholar] [CrossRef]
- Remus, D.M.; van Kranenburg, R.; van Swam, I.I.; Taverne, N.; Bongers, R.S.; Wels, M.; Wells, J.M.; Bron, P.A.; Kleerebezem, M.; Van Kranenburg, R.; et al. Impact of 4 Lactobacillus plantarum capsular polysaccharide clusters on surface glycan composition and host cell signaling. Microb. Cell Fact. 2012, 11, 149. [Google Scholar] [CrossRef] [PubMed]
- van Baarlen, P.; Wells, J.M.; Kleerebezem, M. Regulation of intestinal homeostasis and immunity with probiotic lactobacilli. Trends Immunol. 2013, 34, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Valcheva, R.; Dieleman, L.A. Prebiotics: Definition and protective mechanisms. Best Pract. Res. Clin. Gastroenterol. 2016, 30, 27–37. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, M.; Guo, H.; Yan, J.; Chen, L.; Teng, W.; Ren, F.; Li, Y.; Wang, X.; Luo, J.; et al. Human milk oligosaccharides activate epidermal growth factor receptor and protect against hypoxia-induced injuries in the mouse intestinal epithelium and Caco2 cells. J. Nutr. 2020. [Google Scholar] [CrossRef]
- Holscher, H.D.; Faust, K.L.; Czerkies, L.A.; Litov, R.; Ziegler, E.E.; Lessin, H.; Hatch, T.; Sun, S.; Tappenden, K.A. Effects of prebiotic-containing infant formula on gastrointestinal tolerance and fecal microbiota in a randomized controlled trial. J. Parenter. Enter. Nutr. 2012, 36, 95S–105S. [Google Scholar] [CrossRef]
- Roberfroid, M.; Gibson, G.R.; Hoyles, L.; McCartney, A.L.; Rastall, R.; Rowland, I.; Wolvers, D.; Watzl, B.; Szajewska, H.; Stahl, B.; et al. Prebiotic effects: Metabolic and health benefits. Br. J. Nutr. 2010, 104, S1–S63. [Google Scholar] [CrossRef]
- Jeurink, P.V.; van Esch, B.C.; Rijnierse, A.; Garssen, J.; Knippels, L.M. Mechanisms underlying immune effects of dietary oligosaccharides. Am. J. Clin. Nutr. 2013, 98, 572–577. [Google Scholar] [CrossRef]
- Bhatia, S.; Prabhu, P.N.; Benefiel, A.C.; Miller, M.J.; Chow, J.; Davis, S.R.; Gaskins, H.R. Galacto-oligosaccharides may directly enhance intestinal barrier function through the modulation of goblet cells. Mol. Nutr. Food Res. 2015, 59, 566–573. [Google Scholar] [CrossRef]
- Yang, J.; Martínez, I.; Walter, J.; Keshavarzian, A.; Rose, D.J. Invitro characterization of the impact of selected dietary fibers on fecal microbiota composition and short chain fatty acid production. Anaerobe 2013, 23, 74–81. [Google Scholar] [CrossRef]
- Hamer, H.M.; Jonkers, D.M.A.E.; Bast, A.; Vanhoutvin, S.A.L.W.; Fischer, M.A.J.G.; Kodde, A.; Troost, F.J.; Venema, K.; Brummer, R.J.M. Butyrate modulates oxidative stress in the colonic mucosa of healthy humans. Clin. Nutr. 2009, 28, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.M.W.; de Souza, R.; Kendall, C.W.C.; Emam, A.; Jenkins, D.J.A. Colonic health: Fermentation and short chain fatty acids. J. Clin. Gastroenterol. 2006, 40, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Slimen, I.B.; Najar, T.; Ghram, A.; Dabbebi, H.; Ben Mrad, M.; Abdrabbah, M. Reactive oxygen species, heat stress and oxidative-induced mitochondrial damage. A review. Int. J. Hyperth. 2014, 30, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.J.; Goldsworthy, S.M.; Barnes, A.A.; Eilert, M.M.; Tcheang, L.; Daniels, D.; Muir, A.I.; Wigglesworth, M.J.; Kinghorn, I.; Fraser, N.J.; et al. The Orphan G protein-coupled receptors GPR41 and GPR43 are activated by propionate and other short chain carboxylic acids. J. Biol. Chem. 2003, 278, 11312–11319. [Google Scholar] [CrossRef]
- Levy, M.; Blacher, E.; Elinav, E. Microbiome, metabolites and host immunity. Curr. Opin. Microbiol. 2017, 35, 8–15. [Google Scholar] [CrossRef]
- Song, J.; Jiao, L.F.; Xiao, K.; Luan, Z.S.; Hu, C.H.; Shi, B.; Zhan, X.A. Cello-oligosaccharide ameliorates heat stress-induced impairment of intestinal microflora, morphology and barrier integrity in broilers. Anim. Feed Sci. Technol. 2013, 185, 175–181. [Google Scholar] [CrossRef]
- Sohail, M.U.; Hume, M.E.; Byrd, J.A.; Nisbet, D.J.; Ijaz, A.; Sohail, A.; Shabbir, M.Z.; Rehman, H. Effect of supplementation of prebiotic mannan-oligosaccharides and probiotic mixture on growth performance of broilers subjected to chronic heat stress. Poult. Sci. 2012, 91, 2235–2240. [Google Scholar] [CrossRef]
- Baurhoo, B.; Phillip, L.; Ruiz-Feria, C.A. Effects of purified lignin and mannan oligosaccharides on intestinal integrity and microbial populations in the ceca and litter of broiler chickens. Poult. Sci. 2007, 86, 1070–1078. [Google Scholar] [CrossRef]
- Chichlowski, M.; De Lartigue, G.; German, J.B.; Raybould, H.E.; Mills, D.A. Bifidobacteria isolated from infants and cultured on human milk oligosaccharides affect intestinal epithelial function. J. Pediatr. Gastroenterol. Nutr. 2012, 55, 321–327. [Google Scholar] [CrossRef]
- Pan, X.; Wu, T.; Zhang, L.; Cai, L.; Song, Z. Influence of oligosaccharides on the growth and tolerance capacity of lactobacilli to simulated stress environment. Lett. Appl. Microbiol. 2009, 48, 362–367. [Google Scholar] [CrossRef]
- Yousef, M.; Pichyangkura, R.; Soodvilai, S.; Chatsudthipong, V.; Muanprasat, C. Chitosan oligosaccharide as potential therapy of inflammatory bowel disease: Therapeutic efficacy and possible mechanisms of action. Pharmacol. Res. 2012, 66, 66–79. [Google Scholar] [CrossRef] [PubMed]
- Varasteh, S.; Braber, S.; Akbari, P.; Garssen, J.; Fink-Gremmels, J. Differences in susceptibility to heat stress along the chicken intestine and the protective effects of galacto-oligosaccharides. PLoS ONE 2015, 10, e0138975. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Cai, D.; Cai, W.; Geng, S.; Chen, L.; Han, T. Protective effect of galactooligosaccharide-supplemented enteral nutrition on intestinal barrier function in rats with severe acute pancreatitis. Clin. Nutr. 2009, 28, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Folkerts, J.; Folkerts, G.; Maurer, M.; Braber, S. Microbiota-dependent and -independent effects of dietary fiber on human health. Br. J. Pharmacol. 2019. [Google Scholar] [CrossRef]
- Wu, R.Y.; Abdullah, M.; Määttänen, P.; Pilar, A.V.C.; Scruten, E.; Johnson-Henry, K.C.; Napper, S.; O’Brien, C.; Jones, N.L.; Sherman, P.M. Protein kinase C δ signaling is required for dietary prebiotic-induced strengthening of intestinal epithelial barrier function. Sci. Rep. 2017, 7, 40820. [Google Scholar] [CrossRef] [PubMed]
- Varasteh, S.; Braber, S.; Garssen, J.; Fink-Gremmels, J. Galacto-oligosaccharides exert a protective effect against heat stress in a Caco-2 cell model. J. Funct. Foods 2015, 16, 265–277. [Google Scholar] [CrossRef]
- Akbari, P.; Braber, S.; Alizadeh, A.; Verheijden, K.A.; Schoterman, M.H.; Kraneveld, A.D.; Garssen, J.; Fink-Gremmels, J. Galacto-oligosaccharides protect the intestinal barrier by maintaining the tight junction network and modulating the inflammatory responses after a challenge with the mycotoxin deoxynivalenol in human Caco-2 cell monolayers and B6C3F1 mice. J. Nutr. 2015, 145, 1604–1613. [Google Scholar] [CrossRef]
- Akbari, P.; Fink-Gremmels, J.; Willems, R.H.A.M.; Difilippo, E.; Schols, H.A.; Schoterman, M.H.C.; Garssen, J.; Braber, S. Characterizing microbiota-independent effects of oligosaccharides on intestinal epithelial cells: Insight into the role of structure and size. Eur. J. Nutr. 2017, 56, 1919–1930. [Google Scholar] [CrossRef]
- Muanprasat, C.; Wongkrasant, P.; Satitsri, S.; Moonwiriyakit, A.; Pongkorpsakol, P.; Mattaveewong, T.; Pichyangkura, R.; Chatsudthipong, V. Activation of AMPK by chitosan oligosaccharide in intestinal epithelial cells: Mechanism of action and potential applications in intestinal disorders. Biochem. Pharmacol. 2015, 96, 225–236. [Google Scholar] [CrossRef]
- Gribar, S.C.; Richardson, W.M.; Sodhi, C.P.; Hackam, D.J. No longer an innocent bystander: Epithelial toll-like receptor signaling in the development of mucosal inflammation. Mol. Med. 2008, 14, 645–659. [Google Scholar] [CrossRef]
- Peterson, C.Y.; Costantini, T.W.; Loomis, W.H.; Putnam, J.G.; Wolf, P.; Bansal, V.; Eliceiri, B.P.; Baird, A.; Coimbra, R. Toll-like receptor-4 mediates intestinal barrier breakdown after thermal injury. Surg. Infect. (Larchmt) 2010, 11, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Antoni, L.; Nuding, S.; Wehkamp, J.; Stange, E.F. Intestinal barrier in inflammatory bowel disease. World J. Gastroenterol. 2014, 20, 1165–1179. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, M.; Guo, H.; Yan, J.; Liu, F.; Chen, J.; Li, Y.; Ren, F. Human milk oligosaccharides protect against necrotizing enterocolitis by inhibiting intestinal damage via increasing the proliferation of crypt cells. Mol. Nutr. Food Res. 2019, 63, e1900262. [Google Scholar] [CrossRef] [PubMed]
- Mollen, K.P.; Anand, R.J.; Tsung, A.; Prince, J.M.; Levy, R.M.; Billiar, T.R. Emerging paradigm: Toll-like receptor 4-sentinel for the detection of tissue damage. Shock 2006, 26, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Breslin, J.W.; Wu, M.H.; Guo, M.; Reynoso, R.; Yuan, S.Y. Toll-like receptor 4 contributes to microvascular inflammation and barrier dysfunction in thermal injury. Shock 2008, 29, 349–355. [Google Scholar] [CrossRef]
- Frye, R.; Frye, R.; Slattery, J.; MacFabe, D. The significance of the enteric microbiome on the development of childhood disease: A review of prebiotic and probiotic therapies in disorders of childhood. Clin. Med. Insights Pediatr. 2016. [Google Scholar] [CrossRef]
- Ishikawa, H.; Matsumoto, S.; Ohashi, Y.; Imaoka, A.; Setoyama, H.; Umesaki, Y.; Tanaka, R.; Otani, T. Beneficial effects of probiotic Bifidobacterium and galacto-oligosaccharide in patients with ulcerative colitis: A randomized controlled study. Digestion 2011, 84, 128–133. [Google Scholar] [CrossRef]
- Videla, S.; Vilaseca, J.; Antolin, M.; Garcia-Lafuente, A.; Guarner, F.; Crespo, E.; Casalots, J.; Salas, A.; Malagelada, J.R. Dietary inulin improves distal colitis induced by dextran sodium sulfate in the rat. Am. J. Gastroenterol. 2001, 96, 1486–1493. [Google Scholar] [CrossRef]
- de Kivit, S.; Saeland, E.; Kraneveld, A.D.; van de Kant, H.J.G.; Schouten, B.; van Esch, B.C.A.M.; Knol, J.; Sprikkelman, A.B.; van der Aa, L.B.; Knippels, L.M.J.; et al. Galectin-9 induced by dietary synbiotics is involved in suppression of allergic symptoms in mice and humans. Allergy 2012, 67, 343–352. [Google Scholar] [CrossRef]
- Jantscher-Krenn, E.; Zherebtsov, M.; Nissan, C.; Goth, K.; Guner, Y.S.; Naidu, N.; Choudhury, B.; Grishin, A.V.; Ford, H.R.; Bode, L. The human milk oligosaccharide disialyllacto-N-tetraose prevents necrotising enterocolitis in neonatal rats. Gut 2012, 61, 1417–1425. [Google Scholar] [CrossRef]
- Damaskos, D.; Kolios, G. Probiotics and prebiotics in inflammatory bowel disease: Microflora “on the scope”. Br. J. Clin. Pharmacol. 2008, 65, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Nam, K.-S.; Kim, M.-K.; Shon, Y.-H. Inhibition of proinflammatory cytokine-induced invasiveness of HT-29 cells by chitosan oligosaccharide. J. Microbiol. Biotechnol. 2007, 17, 2042–2045. [Google Scholar] [PubMed]
- Nam, K.-S.; Kim, M.-K.; Shon, Y.-H. Chemopreventive effect of chitosan oligosaccharide against colon carcinogenesis. J. Microbiol. Biotechnol. 2007, 17, 1546–1549. [Google Scholar] [PubMed]
- Verheijden, K.A.T.; Akbari, P.; Willemsen, L.E.M.; Kraneveld, A.D.; Folkerts, G.; Garssen, J.; Fink-Gremmels, J.; Braber, S. Inflammation-induced expression of the alarmin interleukin 33 can be suppressed by galacto-oligosaccharides. Int. Arch. Allergy Immunol. 2015, 167, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Faseleh Jahromi, M.; Wesam Altaher, Y.; Shokryazdan, P.; Ebrahimi, R.; Ebrahimi, M.; Idrus, Z.; Tufarelli, V.; Liang, J.B. Dietary supplementation of a mixture of Lactobacillus strains enhances performance of broiler chickens raised under heat stress conditions. Int. J. Biometeorol. 2016, 60, 1099–1110. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.Y.; Li, B.; Koike, Y.; Määttänen, P.; Miyake, H.; Cadete, M.; Johnson-Henry, K.C.; Botts, S.R.; Lee, C.; Abrahamsson, T.R.; et al. Human milk oligosaccharides increase mucin expression in experimental necrotizing enterocolitis. Mol. Nutr. Food Res. 2019, 63, 1800658. [Google Scholar] [CrossRef] [PubMed]
- Cherbut, C.; Michel, C.; Lecannu, G. The prebiotic characteristics of fructooligosaccharides are necessary for reduction of TNBS-induced colitis in rats. J. Nutr. 2003, 133, 21–27. [Google Scholar] [CrossRef]
- Wen, Z.; Liu, W.; Li, X.; Chen, W.; Liu, Z.; Wen, J.; Liu, Z. A protective role of the NRF2-Keap1 pathway in maintaining intestinal barrier function. Oxid. Med. Cell. Longev. 2019, 2019, 1759149. [Google Scholar] [CrossRef]
- Di Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ROS and RNS sources in physiological and pathological conditions. Oxid. Med. Cell. Longev. 2016, 2016, 1245049. [Google Scholar] [CrossRef]
- Blanco, A.; Blanco, G.; Blanco, A.; Blanco, G. Chapter 10—Antioxidants. In Medical Biochemistry; Academic Press: New York, NY, USA, 2017; pp. 205–214. ISBN 9780128035504. [Google Scholar]
- Li, J.; Liu, Y.; Duan, P.; Yu, R.; Gu, Z.; Li, L.; Liu, Z.; Su, L. NF-κB regulates HSF1 and c-Jun activation in heat stress-induced intestinal epithelial cell apoptosis. Mol. Med. Rep. 2018, 17, 3388–3396. [Google Scholar] [CrossRef]
- Yu, J.; Liu, F.; Yin, P.; Zhao, H.; Luan, W.; Hou, X.; Zhong, Y.; Jia, D.; Zan, J.; Ma, W.; et al. Involvement of oxidative stress and mitogen-activated protein kinase signaling pathways in heat stress-induced injury in the rat small intestine. Stress 2013, 16, 99–113. [Google Scholar] [CrossRef] [PubMed]
- Shila, S.; Subathra, M.; Devi, M.A.; Panneerselvam, C. Arsenic intoxication-induced reduction of glutathione level and of the activity of related enzymes in rat brain regions: Reversal by DL-alpha-lipoic acid. Arch. Toxicol. 2005, 79, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Rochette, L.; Ghibu, S.; Richard, C.; Zeller, M.; Cottin, Y.; Vergely, C. Direct and indirect antioxidant properties of α-lipoic acid and therapeutic potential. Mol. Nutr. Food Res. 2013, 57, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, P.P.; Jena, G.B. Role of α-lipoic acid in dextran sulfate sodium-induced ulcerative colitis in mice: Studies on inflammation, oxidative stress, DNA damage and fibrosis. Food Chem. Toxicol. 2013, 59, 339–355. [Google Scholar] [CrossRef] [PubMed]
- Fan, P.; Tan, Y.; Jin, K.; Lin, C.; Xia, S.; Han, B.; Zhang, F.; Wu, L.; Ma, X. Supplemental lipoic acid relieves post-weaning diarrhoea by decreasing intestinal permeability in rats. J. Anim. Physiol. Anim. Nutr. 2017, 101, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Varasteh, S.; Fink-Gremmels, J.; Garssen, J.; Braber, S.; Nl, S.B. α-Lipoic acid prevents the intestinal epithelial monolayer damage under heat stress conditions: Model experiments in Caco-2 cells. Eur. J. Nutr. 2018, 57, 1577–1589. [Google Scholar] [CrossRef]
- Ma, X.; He, P.; Sun, P.; Han, P. Lipoic acid: An immunomodulator that attenuates glycinin-induced anaphylactic reactions in a rat model. J. Agric. Food Chem. 2010, 58, 5086–5092. [Google Scholar] [CrossRef]
- Rao, R.K.; Basuroy, S.; Rao, V.U.; Karnaky, K.J., Jr.; Gupta, A. Tyrosine phosphorylation and dissociation of occludin-ZO-1 and E-cadherin-β-catenin complexes from the cytoskeleton by oxidative stress. Biochem. J. 2002, 368, 471–481. [Google Scholar] [CrossRef]
- Koriyama, Y.; Nakayama, Y.; Matsugo, S.; Kato, S. Protective effect of lipoic acid against oxidative stress is mediated by Keap1/Nrf2-dependent heme oxygenase-1 induction in the RGC-5 cell line. Brain Res. 2013, 1499, 145–157. [Google Scholar] [CrossRef]
- Durante, W. Protective role of heme oxygenase-1 against inflammation in atherosclerosis. Front. Biosci. 2011, 16, 2372–2388. [Google Scholar] [CrossRef]
- Park, S.J.; Lee, K.S.; Lee, S.J.; Kim, S.R.; Park, S.Y.; Jeon, M.S.; Lee, H.B.; Lee, Y.C. L-2-Oxothiazolidine-4-carboxylic acid or α-lipoic acid attenuates airway remodeling: Involvement of nuclear factor-κB (NF-κB), nuclear factor erythroid 2p45-related factor-2 (Nrf2), and hypoxia-inducible factor (HIF). Int. J. Mol. Sci. 2012, 13, 7915–7937. [Google Scholar] [CrossRef] [PubMed]
- Shay, K.P.; Moreau, R.F.; Smith, E.J.; Smith, A.R.; Hagen, T.M. Alpha-lipoic acid as a dietary supplement: Molecular mechanisms and therapeutic potential. Biochim. Biophys. Acta 2009, 1790, 1149–1160. [Google Scholar] [CrossRef] [PubMed]
- El-Gowelli, H.M.; Saad, E.I.; Abdel-Galil, A.G.A.; Ibrahim, E.R. Co-administration of α-lipoic acid and cyclosporine aggravates colon ulceration of acetic acid-induced ulcerative colitis via facilitation of NO/COX-2/miR-210 cascade. Toxicol. Appl. Pharmacol. 2015, 288, 300–312. [Google Scholar] [CrossRef] [PubMed]
- Sahin, K.; Orhan, C.; Smith, M.O.; Sahin, N. Molecular targets of dietary phytochemicals for the alleviation of heat stress in poultry. Worlds. Poult. Sci. J. 2013, 69, 113–124. [Google Scholar] [CrossRef]
- Zhuang, H.; Kim, Y.S.; Koehler, R.C.; Doré, S. Potential mechanism by which resveratrol, a red wine constituent, protects neurons. Ann. N. Y. Acad. Sci. 2003, 993, 276–286. [Google Scholar] [CrossRef]
- Liu, L.L.; He, J.H.; Xie, H.B.; Yang, Y.S.; Li, J.C.; Zou, Y. Resveratrol induces antioxidant and heat shock protein mRNA expression in response to heat stress in black-boned chickens. Poult. Sci. 2014, 93, 54–62. [Google Scholar] [CrossRef]
- Cao, S.; Shen, Z.; Wang, C.; Zhang, Q.; Hong, Q.; He, Y.; Hu, C. Resveratrol improves intestinal barrier function, alleviates mitochondrial dysfunction and induces mitophagy in diquat challenged piglets. Food Funct. 2019, 10, 344–354. [Google Scholar] [CrossRef]
- Putics, A.; Végh, E.M.; Csermely, P.; Soti, C. Resveratrol induces the heat-shock response and protects human cells from severe heat stress. Antioxid. Redox Signal. 2008, 10, 65–75. [Google Scholar] [CrossRef]
- Burkitt, M.J.; Duncan, J. Effects of trans-resveratrol on copper-dependent hydroxyl-radical formation and DNA damage: Evidence for hydroxyl-radical scavenging and a novel, glutathione-sparing mechanism of action. Arch. Biochem. Biophys. 2000, 381, 253–263. [Google Scholar] [CrossRef]
- Wang, N.; Han, Q.; Wang, G.; Ma, W.P.; Wang, J.; Wu, W.X.; Guo, Y.; Liu, L.; Jiang, X.Y.; Xie, X.L.; et al. Resveratrol protects oxidative stress-induced intestinal epithelial barrier dysfunction by upregulating heme oxygenase-1 expression. Dig. Dis. Sci. 2016, 61, 2522–2534. [Google Scholar] [CrossRef]
- Liu, L.; Fu, C.; Yan, M.; Xie, H.; Li, S.; Yu, Q.; He, S.; He, J. Resveratrol modulates intestinal morphology and HSP70/90, NF-κB and EGF expression in the jejunal mucosa of black-boned chickens on exposure to circular heat stress. Food Funct. 2016, 7, 1329–1338. [Google Scholar] [CrossRef] [PubMed]
- Ling, K.H.; Wan, M.L.Y.; El-Nezami, H.; Wang, M. Protective capacity of resveratrol, a natural polyphenolic compound, against deoxynivalenol-induced intestinal barrier dysfunction and bacterial translocation. Chem. Res. Toxicol. 2016, 9, 823–833. [Google Scholar] [CrossRef] [PubMed]
- Mayangsari, Y.; Suzuki, T. Resveratrol ameliorates intestinal barrier defects and inflammation in colitic mice and intestinal cells. J. Agric. Food Chem. 2018, 66, 12666–12674. [Google Scholar] [CrossRef] [PubMed]
- Jo, H.A.; Hwang, D.; Kim, J.K.; Lim, Y.H. Oxyresveratrol improves tight junction integrity through the PKC and MAPK signaling pathways in Caco-2 cells. Food Chem. Toxicol. 2017, 108, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Yoo, C.-G.; Lee, S.; Lee, C.-T.; Kim, Y.W.; Han, S.K.; Shim, Y.-S. Anti-inflammatory effect of heat shock protein induction is related to stabilization of IB through preventing IB kinase activation in respiratory epithelial cells. J. Immunol. 2000, 164, 5416–5423. [Google Scholar] [CrossRef] [PubMed]
- Rossi, A.; Coccia, M.; Trotta, E.; Angelini, M.; Santoro, M.G. Regulation of cyclooxygenase-2 expression by heat: A novel aspect of heat shock factor 1 function in human cells. PLoS ONE 2012, 7, e31304. [Google Scholar] [CrossRef]
- Sengottuvelan, M.; Deeptha, K.; Nalini, N. Influence of dietary resveratrol on early and late molecular markers of 1,2-dimethylhydrazine-induced colon carcinogenesis. Nutrition 2009, 25, 1169–1176. [Google Scholar] [CrossRef]
- Sergent, T.; Piront, N.; Meurice, J.; Toussaint, O.; Schneider, Y.J. Anti-inflammatory effects of dietary phenolic compounds in an in vitro model of inflamed human intestinal epithelium. Chem. Biol. Interact. 2010, 188, 659–667. [Google Scholar] [CrossRef]
- Szajewska, H.; Horvath, A.; Koletzko, B. Effect of n-3 long-chain polyunsaturated fatty acid supplementation of women with low-risk pregnancies on pregnancy outcomes and growth measures at birth: A meta-analysis of randomized controlled trials. Am. J. Clin. Nutr. 2006, 83, 1337–1344. [Google Scholar] [CrossRef]
- Yates, C.M.; Calder, P.C.; Ed Rainger, G. Pharmacology and therapeutics of omega-3 polyunsaturated fatty acids in chronic inflammatory disease. Pharmacol. Ther. 2014, 141, 272–282. [Google Scholar] [CrossRef]
- Giordano, E.; Visioli, F. Long-chain omega 3 fatty acids: Molecular bases of potential antioxidant actions. Prostaglandins Leukot. Essent. Fat. Acids 2014, 90, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Visioli, F.; Giordano, E.; Nicod, N.M.; Dávalos, A. Molecular targets of omega 3 and conjugated linoleic fatty acids—“Micromanaging” cellular response. Front. Physiol. 2012, 3, 42. [Google Scholar] [CrossRef] [PubMed]
- Brahmbhatt, V.; Oliveira, M.; Briand, M.; Perrisseau, G.; Bastic Schmid, V.; Destaillats, F.; Pace-Asciak, C.; Benyacoub, J.; Bosco, N. Protective effects of dietary EPA and DHA on ischemia-reperfusion-induced intestinal stress. J. Nutr. Biochem. 2013, 24, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Xiao, G.; Tang, L.; Yuan, F.; Zhu, W.; Zhang, S.; Liu, Z.; Geng, Y.; Qiu, X.; Zhang, Y.; Su, L. Eicosapentaenoic acid enhances heat stress-impaired intestinal epithelial barrier function in Caco-2 cells. PLoS ONE 2013, 8, e73571. [Google Scholar] [CrossRef] [PubMed]
- Beguin, P.; Errachid, A.; Larondelle, Y.; Schneider, Y.J. Effect of polyunsaturated fatty acids on tight junctions in a model of the human intestinal epithelium under normal and inflammatory conditions. Food Funct. 2013, 4, 923–931. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Wang, C.; Chin, Y.; Chen, X.; Gao, Y.; Yuan, S.; Xue, C.; Wang, Y.; Tang, Q. DHA-phospholipids (DHA-PL) and EPA-phospholipids (EPA-PL) prevent intestinal dysfunction induced by chronic stress. Food Funct. 2019, 10, 277–288. [Google Scholar] [CrossRef]
- Grimble, R.F. Basics in clinical nutrition: Immunonutrition—Nutrients which influence immunity: Effect and mechanism of action. e-SPEN Eur. e-J. Clin. Nutr. Metab. 2009, 1, e10–e13. [Google Scholar] [CrossRef]
- Zhao, J.; Shi, P.; Sun, Y.; Sun, J.; Dong, J.-N.; Wang, H.-G.; Zuo, L.-G.; Gong, J.-F.; Li, Y.; Gu, L.-L.; et al. DHA protects against experimental colitis in IL-10-deficient mice associated with the modulation of intestinal epithelial barrier function. Br. J. Nutr. 2015, 114, 181–188. [Google Scholar] [CrossRef]
- Calviello, G.; Di Nicuolo, F.; Gragnoli, S.; Piccioni, E.; Serini, S.; Maggiano, N.; Tringali, G.; Navarra, P.; Ranelletti, F.O.; Palozza, P. n-3 PUFAs reduce VEGF expression in human colon cancer cells modulating the COX-2/PGE 2 induced ERK-1 and -2 and HIF-1α induction pathway. Carcinogenesis 2004, 25, 2303–2310. [Google Scholar] [CrossRef]
- Narayanan, B.A.; Narayanan, N.K.; Simi, B.; Reddy, B.S. Modulation of inducible nitric oxide synthase and related proinflammatory genes by the omega-3 fatty acid docosahexaenoic acid in human colon cancer cells. Cancer Res. 2003, 63, 972–979. [Google Scholar]
- De Carlo, F.; Witte, T.R.; Hardman, W.E.; Claudio, P.P. Omega-3 eicosapentaenoic acid decreases CD133 colon cancer stem-like cell marker expression while increasing sensitivity to chemotherapy. PLoS ONE 2013, 8, e69760. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, W.; Sun, Y.; Han, F.; Hu, C.-A.A.; Wu, Z. Amino acid deprivation disrupts barrier function and induces protective autophagy in intestinal porcine epithelial cells. Amino Acids 2015, 47, 2177–2184. [Google Scholar] [CrossRef] [PubMed]
- Genton, L.; Cani, P.D.; Schrenzel, J. Alterations of gut barrier and gut microbiota in food restriction, food deprivation and protein-energy wasting. Clin. Nutr. 2015, 34, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Brake, J.; Balnave, D.; Dibner, J.J. Optimum dietary arginine:lysine ratio for broiler chickens is altered during heat stress in association with changes in intestinal uptake and dietary sodium chloride. Br. Poult. Sci. 1998, 39, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.W.; Qiao, S.Y.; Li, D.F. Amino acids and gut function. Amino Acids 2009, 37, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Morales, A.; Hernández, L.; Buenabad, L.; Avelar, E.; Bernal, H.; Baumgard, L.H.; Cervantes, M. Effect of heat stress on the endogenous intestinal loss of amino acids in growing pigs. J. Anim. Sci. 2016, 94, 165–172. [Google Scholar] [CrossRef]
- Wang, B.; Wu, Z.; Ji, Y.; Sun, K.; Dai, Z.; Wu, G. L-Glutamine enhances tight junction integrity by activating CaMK kinase 2–AMP-activated protein kinase signaling in intestinal porcine epithelial cells. J. Nutr. 2016, 146, 501–508. [Google Scholar] [CrossRef]
- Marc Rhoads, J.; Wu, G. Glutamine, arginine, and leucine signaling in the intestine. Amino Acids 2009, 37, 111–122. [Google Scholar] [CrossRef]
- Grishin, A.; Bowling, J.; Bell, B.; Wang, J.; Ford, H.R. Roles of nitric oxide and intestinal microbiota in the pathogenesis of necrotizing enterocolitis. J. Pediatr. Surg. 2016, 51, 13–17. [Google Scholar] [CrossRef]
- Chen, Y.-C.; Liu, Y.-C.; Yen, D.H.-T.; Wang, L.-M.; Huang, C.-I.; Lee, C.-H.; Lin, M.-T. L-Arginine causes amelioration of cerebrovascular dysfunction and brain inflammation during experimental heatstroke. Shock 2007, 29, 212–216. [Google Scholar] [CrossRef]
- Chatterjee, S.; Premachandran, S.; Sharma, D.; Bagewadikar, R.S.; Poduval, T.B. Therapeutic treatment with L-arginine rescues mice from heat stroke-induced death: Physiological and molecular mechanisms. Shock 2005, 24, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-K.; Chang, C.-P.; Chiu, W.-T.; Lin, M.-T. Prevention and repair of circulatory shock and cerebral ischemia/injury by various agents in experimental heatstroke. Curr. Med. Chem. 2006, 13, 3145–3154. [Google Scholar] [CrossRef] [PubMed]
- Sukhotnik, I.; Helou, H.; Mogilner, J.; Lurie, M.; Bernsteyn, A.; Coran, A.G.; Shiloni, E. Oral arginine improves intestinal recovery following ischemia-reperfusion injury in rat. Pediatr. Surg. Int. 2005, 21, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.; Yin, Y.; Kong, X.; Li, P.; Li, X.; Gao, H.; Li, X.; Huang, R.; Wu, G. L-Arginine stimulates proliferation and prevents endotoxin-induced death of intestinal cells. Amino Acids 2010, 38, 1227–1235. [Google Scholar] [CrossRef] [PubMed]
- Rhoads, J.M.; Chen, W.; Gookin, J.; Wu, G.Y.; Fu, Q.; Blikslager, T.; Rippe, R.A.; Argenzio, R.A.; Cance, W.G.; Weaver, E.M.; et al. Arginine stimulates intestinal cell migration through a focal adhesion kinase dependent mechanism. Gut 2004, 53, 514–522. [Google Scholar] [CrossRef] [PubMed]
- Sharma, J.N.; Al-Omran, A.; Parvathy, S.S. Role of nitric oxide in inflammatory diseases. Inflammopharmacology 2007, 15, 252–259. [Google Scholar] [CrossRef]
- Kozar, R.A.; Verner-Cole, E.; Schultz, S.G.; Sato, N.; Bick, R.J.; Desoignie, R.; Poindexter, B.J.; Moore, F.A. The immune-enhancing enteral agents arginine and glutamine differentially modulate gut barrier function following mesenteric ischemia/reperfusion. J. Trauma 2004, 57, 1150–1156. [Google Scholar] [CrossRef]
- Morris, S.M. Enzymes of arginine metabolism. J. Nutr. 2004, 134, 2743–2747. [Google Scholar] [CrossRef]
- Rhoads, J.M.; Liu, Y.; Niu, X.; Surendran, S.; Wu, G. Arginine stimulates cdx2-transformed intestinal epithelial cell migration via a mechanism requiring both nitric oxide and phosphorylation of p70 S6 kinase. J. Nutr. 2008, 138, 1652–1657. [Google Scholar] [CrossRef]
- Inoue, T.; Kwon, A.; Oda, M.; Kaibori, M.; Kamiyama, Y.; Nishizawa, M.; Ito, S.; Okumura, T. Hypoxia and heat inhibit inducible nitric oxide synthase gene expression by different mechanisms in rat hepatocytes. Hepatology 2000, 32, 1037–1044. [Google Scholar] [CrossRef]
- Costa, K.A.; Soares, A.D.; Wanner, S.P.; Santos, R.D.; Fernandes, S.O.; Martins, F.D.; Nicoli, J.R.; Coimbra, C.C.; Cardoso, V.N. L-Arginine supplementation prevents increases in intestinal permeability and bacterial translocation in male swiss mice subjected to physical exercise under environmental heat stress. J. Nutr. 2014, 144, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Lacerda, A.C.R.; Marubayashi, U.; Coimbra, C.C. Nitric oxide pathway is an important modulator of heat loss in rats during exercise. Brain Res. Bull. 2005, 67, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Quirino, I.E.; Cardoso, V.N.; Santos, R.D.; Evangelista, W.P.; Arantes, R.M.; Fiúza, J.A.; Glória, M.B.; Alvarez-Leite, J.I.; Batista, M.A.; Correia, M.I.T.D. The role of L-arginine and inducible nitric oxide synthase in intestinal permeability and bacterial translocation. J. Parenter. Enter. Nutr. 2013, 37, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Bailey, S.J.; Winyard, P.G.; Vanhatalo, A.; Blackwell, J.R.; DiMenna, F.J.; Wilkerson, D.P.; Jones, A.M. Acute L-arginine supplementation reduces the O2 cost of moderate-intensity exercise and enhances high-intensity exercise tolerance. J. Appl. Physiol. 2010, 109, 1394–1403. [Google Scholar] [CrossRef] [PubMed]
- Vanhatalo, A.; Fulford, J.; Bailey, S.J.; Blackwell, J.R.; Winyard, P.G.; Jones, A.M. Dietary nitrate reduces muscle metabolic perturbation and improves exercise tolerance in hypoxia. J. Physiol. 2011, 589, 5517–5528. [Google Scholar] [CrossRef] [PubMed]
- Coburn, L.A.; Gong, X.; Singh, K.; Asim, M.; Scull, B.P.; Allaman, M.M.; Williams, C.S.; Rosen, M.J.; Washington, M.K.; Barry, D.P.; et al. L-arginine supplementation improves responses to injury and inflammation in dextran sulfate sodium colitis. PLoS ONE 2012, 7, e33546. [Google Scholar] [CrossRef] [PubMed]
- Chapman, J.C.; Liu, Y.; Zhu, L.; Rhoads, J.M. Arginine and citrulline protect intestinal cell monolayer tight junctions from hypoxia-induced injury in piglets. Pediatr. Res. 2012, 72, 576–582. [Google Scholar] [CrossRef]
- Tanaka, A.; Mizoguchi, H.; Kunikata, T.; Miyazawa, T.; Takeuchi, K. Protection by constitutively formed nitric oxide of intestinal damage induced by indomethacin in rats. J. Physiol. Paris 2001, 95, 35–41. [Google Scholar] [CrossRef]
- Viana, M.; Santos, R.G.; Generoso, S.V.; Arantes, R.M.; Correia, M.I.; Cardoso, V.N. Pretreatment with arginine preserves intestinal barrier integrity and reduces bacterial translocation in mice. Nutrition 2010, 26, 218–223. [Google Scholar] [CrossRef]
- Morris, S.M. Arginine metabolism: Boundaries of our knowledge. J. Nutr. 2007, 137, 1602–1609. [Google Scholar] [CrossRef]
- Singer, I.I.; Kawka, D.W.; Scott, S.; Weidner, J.R.; Mumford, R.A.; Riehl, T.E.; Stenson, W.F. Expression of inducible nitric oxide synthase and nitrotyrosine in colonic epithelium in inflammatory bowel disease. Gastroenterology 1996, 111, 871–885. [Google Scholar] [CrossRef]
- Marcinkiewicz, E.W.A.; Opicki, S.C.H.; Marcinkiewicz, J. Nitric oxide—A pro-inflammatory and anti-inflammatory mediator. Cent. Eur. J. Immunol. 2003, 28, 74–78. [Google Scholar]
- Katsube, T.; Tsuji, H.; Onoda, M. Nitric oxide attenuates hydrogen peroxide-induced barrier disruption and protein tyrosine phosphorylation in monolayers of intestinal epithelial cell. Biochim. Biophys. Acta 2007, 1773, 794–803. [Google Scholar] [CrossRef] [PubMed]
- Sheth, P.; Seth, A.; Atkinson, K.J.; Gheyi, T.; Kale, G.; Giorgianni, F.; Desiderio, D.M.; Li, C.; Naren, A.; Rao, R. Acetaldehyde dissociates the PTP1B-E-cadherin-beta-catenin complex in Caco-2 cell monolayers by a phosphorylation-dependent mechanism. Biochem. J. 2007, 402, 291–300. [Google Scholar] [CrossRef]
- Maher, P.A.; Pasquale, E.B. Heat shock induces protein tyrosine phosphorylation in cultured cells. J. Cell Biol. 1989, 108, 2029–2035. [Google Scholar] [CrossRef]
- Gookin, J.L.; Rhoads, J.M.; Argenzio, R.A. Inducible nitric oxide synthase mediates early epithelial repair of porcine ileum. Am. J. Physiol. Gastrointest. Liver Physiol. 2002, 283, 157–168. [Google Scholar] [CrossRef]
- Corl, B.A.; Odle, J.; Niu, X.; Moeser, A.J.; Gatlin, L.A.; Phillips, O.T.; Blikslager, A.T.; Rhoads, J.M. Arginine activates intestinal p70(S6k) and protein synthesis in piglet rotavirus enteritis. J. Nutr. 2008, 138, 24–29. [Google Scholar] [CrossRef]
- Ban, H.; Shigemitsu, K.; Yamatsuji, T.; Haisa, M.; Nakajo, T.; Takaoka, M.; Nobuhisa, T.; Gunduz, M.; Tanaka, N.; Naomoto, Y. Arginine and Leucine regulate p70 S6 kinase and 4E-BP1 in intestinal epithelial cells. Int. J. Mol. Med. 2004, 13, 537–543. [Google Scholar] [CrossRef]
- Wang, J.Y. Polyamines regulate expression of E-cadherin and play an important role in control of intestinal epithelial barrier function. Inflammopharmacology 2005, 13, 91–101. [Google Scholar] [CrossRef]
- Iwashita, Y.; Sakiyama, T.; Musch, M.W.; Ropeleski, M.J.; Tsubouchi, H.; Chang, E.B. Polyamines mediate glutamine-dependent induction of the intestinal epithelial heat shock response. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 301, 181–187. [Google Scholar] [CrossRef]
- Rhee, H.J.; Kim, E.J.; Lee, J.K. Physiological polyamines: Simple primordial stress molecules. J. Cell. Mol. Med. 2007, 11, 685–703. [Google Scholar] [CrossRef] [PubMed]
- Viana, M.L.; Dos Santos, R.D.; Generoso, S.D.V.; Nicoli, J.R.; Martins, F.D.S.; Nogueira-Machado, J.A.; Arantes, R.M.; Correia, M.I.; Cardoso, V.N. The role of l-arginine-nitric oxide pathway in bacterial translocation. Amino Acids 2013, 45, 1089–1096. [Google Scholar] [CrossRef] [PubMed]
- Connelly, L.; Palacios-Callender, M.; Ameixa, C.; Moncada, S.; Hobbs, A.J. Biphasic regulation of NF-kappa B activity underlies the pro- and anti-inflammatory actions of nitric oxide. J. Immunol. 2001, 166, 3873–3881. [Google Scholar] [CrossRef] [PubMed]
- Yi, G.F.; Allee, G.L.; Knight, C.D.; Dibner, J.J. Impact of glutamine and Oasis hatchling supplement on growth performance, small intestinal morphology, and immune response of broilers vaccinated and challenged with Eimeria maxima. Poult. Sci. 2005, 84, 283–293. [Google Scholar] [CrossRef]
- Wu, G. Functional amino acids in growth, reproduction, and health. Adv. Nutr. 2010, 1, 31–37. [Google Scholar] [CrossRef]
- Windmueller, H.G.; Spaeth, A.E. Identification of ketone bodies and glutamine as the major respiratory fuels in vivo for postabsorptive rat small intestine. J. Biol. Chem. 1978, 253, 69–76. [Google Scholar]
- Daniele, B.; Perrone, F.; Gallo, C.; Pignata, S.; De Martino, S.; De Vivo, R.; Barletta, E.; Tambaro, R.; Abbiati, R.; D’Agostino, L. Oral glutamine in the prevention of fluorouracil induced intestinal toxicity: A double blind, placebo controlled, randomised trial. Gut 2001, 48, 28–33. [Google Scholar] [CrossRef]
- Dai, S.F.; Gao, F.; Xu, X.L.; Zhang, W.H.; Song, S.X.; Zhou, G.H. Effects of dietary glutamine and gamma-aminobutyric acid on meat colour, pH, composition, and water-holding characteristic in broilers under cyclic heat stress. Br. Poult. Sci. 2012, 53, 471–481. [Google Scholar] [CrossRef]
- Ulluwishewa, D.; Anderson, R.C.; McNabb, W.C.; Moughan, P.J.; Wells, J.M.; Roy, N.C. Regulation of tight junction permeability by intestinal bacteria and dietary components. J. Nutr. 2011, 141, 769–776. [Google Scholar] [CrossRef]
- Li, P.; Yin, Y.; Li, D.; Kim, S.W.; Wu, G. Review Article: Amino acids and immune function. Br. J. Nutr. 2007, 98, 237–252. [Google Scholar] [CrossRef]
- Wischmeyer, P.E.; Musch, M.W.; Madonna, M.B.; Thisted, R.; Chang, E.B. Glutamine protects intestinal epithelial cells: Role of inducible HSP70. Am. J. Physiol. 1997, 272, G879–G884. [Google Scholar] [CrossRef] [PubMed]
- Ropeleski, M.J.; Riehm, J.; Baer, K.A.; Musch, M.W.; Chang, E.B. Anti-apoptotic effects of L-glutamine-mediated transcriptional modulation of the heat shock protein 72 during heat shock. Gastroenterology 2005, 129, 170–184. [Google Scholar] [CrossRef] [PubMed]
- Phanvijhitsiri, K.; Musch, M.W.; Ropeleski, M.J.; Chang, E.B. Heat induction of heat shock protein 25 requires cellular glutamine in intestinal epithelial cells. Am. J. Physiol. Cell Physiol. 2006, 291, C290–C299. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Souba, W.W.; Croce, C.M.; Verne, G.N. MicroRNA-29a regulates intestinal membrane permeability in patients with irritable bowel syndrome. Gut 2010, 59, 775–784. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, K.K.; Shukla, P.K.; Mir, H.; Manda, B.; Gangwar, R.; Yadav, N.; McMullen, M.; Nagy, L.E.; Rao, R.K. Glutamine supplementation attenuates ethanol-induced disruption of apical junctional complexes in colonic epithelium and ameliorates gut barrier dysfunction and fatty liver in mice. J. Nutr. Biochem. 2016, 27, 16–26. [Google Scholar] [CrossRef]
- Bertrand, J.; Ghouzali, I.; Guérin, C.; Bôle-Feysot, C.; Gouteux, M.; Déchelotte, P.; Ducrotté, P.; Coëffier, M. Glutamine restores tight junction protein Claudin-1 expression in colonic mucosa of patients with diarrhea-predominant irritable bowel syndrome. J. Parenter. Enter. Nutr. 2016, 40, 1170–1176. [Google Scholar] [CrossRef]
- Shakeri, M.; Zulkifli, I.; Soleimani, A.F.; O’Reilly, E.L.; Eckersall, P.D.; Anna, A.A.; Kumari, S.; Abdullah, F.F.J. Environment, well-being, and behavior: Response to dietary supplementation of L-glutamine and L-glutamate in broiler chickens reared at different stocking densities under hot, humid tropical conditions. Poult. Sci. 2014, 93, 2700–2708. [Google Scholar] [CrossRef]
- Wu, G.; Meier, S.A.; Knabe, D.A. Dietary glutamine supplementation prevents jejunal atrophy in weaned pigs. J. Nutr. 1996, 126, 2578–2584. [Google Scholar] [CrossRef]
- Li, N.; Lewis, P.; Samuelson, D.; Liboni, K.; Neu, J. Glutamine regulates Caco-2 cell tight junction proteins. Am. J. Physiol. Liver Physiol. 2004, 287, G726–G733. [Google Scholar] [CrossRef]
- Li, N.; DeMarco, V.G.; West, C.M.; Neu, J. Glutamine supports recovery from loss of transepithelial resistance and increase of permeability induced by media change in Caco-2 cells. J. Nutr. Biochem. 2003, 14, 401–408. [Google Scholar] [CrossRef]
- Gu, X.H.; Hao, Y.; Wang, X.L. Overexpression of heat shock protein 70 and its relationship to intestine under acute heat stress in broilers: 2. Intestinal oxidative stress. Poult. Sci. 2012, 91, 790–799. [Google Scholar] [CrossRef] [PubMed]
- Zuhl, M.N.; Lanphere, K.R.; Kravitz, L.; Mermier, C.M.; Schneider, S.; Dokladny, K.; Moseley, P.L. Effects of oral glutamine supplementation on exercise-induced gastrointestinal permeability and tight junction protein expression. J. Appl. Physiol. 2014, 116, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Akagi, R.; Ohno, M.; Matsubara, K.; Fujimoto, M.; Nakai, A.; Inouye, S. Glutamine protects intestinal barrier function of colon epithelial cells from ethanol by modulating Hsp70 expression. Pharmacology 2013, 91, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Giriş, M.; Erbil, Y.; Öztezcan, S.; Olgaç, V.; Barbaros, U.; Deveci, U.; Kirgiz, B.; Uysal, M.; Toker, G.A. The effect of heme oxygenase-1 induction by glutamine on radiation-induced intestinal damage: The effect of heme oxygenase-1 on radiation enteritis. Am. J. Surg. 2006, 191, 503–509. [Google Scholar] [CrossRef]
- Seth, A.; Basuroy, S.; Sheth, P.; Rao, R.K. L-Glutamine ameliorates acetaldehyde-induced increase in paracellular permeability in Caco-2 cell monolayer. Am. J. Physiol. Liver Physiol. 2004, 287, G510–G517. [Google Scholar] [CrossRef]
- Li, N.; Neu, J. Glutamine deprivation alters intestinal tight junctions via a PI3-K/Akt mediated pathway in Caco-2 cells. J. Nutr. 2009, 139, 710–714. [Google Scholar] [CrossRef]
- Beutheu, S.; Ghouzali, I.; Galas, L.; Déchelotte, P.; Coëffier, M. Glutamine and arginine improve permeability and tight junction protein expression in methotrexate-treated Caco-2 cells. Clin. Nutr. 2013, 32, 863–869. [Google Scholar] [CrossRef]
- Coëffier, M.; Marion, R.; Ducrotté, P.; Déchelotte, P. Modulating effect of glutamine on IL-1β-induced cytokine production by human gut. Clin. Nutr. 2003, 22, 407–413. [Google Scholar] [CrossRef]
- Li, N.; Liboni, K.; Fang, M.Z.; Samuelson, D.; Lewis, P.; Patel, R.; Neu, J. Glutamine decreases lipopolysaccharide-induced intestinal inflammation in infant rats. Am. J. Physiol. Liver Physiol. 2004, 286, G914–G921. [Google Scholar] [CrossRef]
- Cöeffier, M.; Marion-Letellier, R.; Déchelotte, P. Potential for amino acids supplementation during inflammatory bowel diseases. Inflamm. Bowel Dis. 2010, 16, 518–524. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, P.; Deng, G.; Liu, X.; Lu, D. Improvements of immune status, intestinal integrity and gain performance in the early-weaned calves parenterally supplemented with l-alanyl-l-glutamine dipeptide. Vet. Immunol. Immunopathol. 2012, 145, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.J.; Liu, N.; Wu, X.H.; Wang, G.Y.; Lin, L. Glutamine alleviates heat stress-induced impairment of intestinal morphology, intestinal inflammatory response, and barrier integrity in broilers. Poult. Sci. 2018, 97, 2675–2683. [Google Scholar] [CrossRef] [PubMed]
- Lesueur, C.; Bôle-Feysot, C.; Bekri, S.; Husson, A.; Lavoinne, A.; Brasse-Lagnel, C. Glutamine induces nuclear degradation of the NF-κB p65 subunit in Caco-2/TC7 cells. Biochimie 2012, 94, 806–815. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, A.; Snead, C.; Yetik-Anacak, G.; Antonova, G.; Zeng, J.; Catravas, J.D. Heat shock protein 90 inhibitors attenuate LPS-induced endothelial hyperpermeability. Am. J. Physiol. Lung Cell. Mol. Physiol. 2008, 294, L755–L763. [Google Scholar] [CrossRef]
- Fanelli, M.A.; Montt-Guevara, M.; Diblasi, A.M.; Gago, F.E.; Tello, O.; Cuello-Carrión, F.D.; Callegari, E.; Bausero, M.A.; Ciocca, D.R. P-Cadherin and β-catenin are useful prognostic markers in breast cancer patients; β-catenin interacts with heat shock protein Hsp27. Cell Stress Chaperones 2008, 13, 207–220. [Google Scholar] [CrossRef] [PubMed]
- Shinde, T.; Perera, A.P.; Vemuri, R.; Gondalia, S.V.; Karpe, A.V.; Beale, D.J.; Shastri, S.; Southam, B.; Eri, R.; Stanley, R. Synbiotic supplementation containing whole plant sugar cane Fibre and probiotic spores potentiates protective synergistic effects in mouse model of IBD. Nutrients 2019, 11, 818. [Google Scholar] [CrossRef]
- Salberg, S.; Yamakawa, G.; Christensen, J.; Kolb, B.; Mychasiuk, R. Assessment of a nutritional supplement containing resveratrol, prebiotic fiber, and omega-3 fatty acids for the prevention and treatment of mild traumatic brain injury in rats. Neuroscience 2017, 365, 146–157. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Name | Integrity | Immunomodulation | Other Effect(s) | Reference | |
|---|---|---|---|---|---|
| Pro-biotics | Lactobacillus species | TEER↑ | IL-10↑ | Integrin-p38 MAPK activation↑ | [77,82,83,84,88,89,135] |
| Intestinal permeability↓ | IL-27↑ | HSP expression↑ | |||
| ZO-1↑ | IL-1↓ | Antioxidative capacity↑ | |||
| occludin↑ | IL-6↓ | Nutrient transporters↑ | |||
| E-cadherin↑ | TNF-α↓ | ||||
| claudin-2↑ | NF-κB activation↓ | ||||
| Bifidobacterium species | claudin-3↑ | Corticosterone↓ | Mucin genes transcription and protein production↑ | [77,88,89] | |
| Morphological damage↓ | IgA secreting cells↑ | ||||
| β-catenin↑ | Intraepithelial lymphocytes↓ | ||||
| Bacillus species | [79,80,85,88,89] | ||||
| E. coli Nissle | ZO-2 dissociation↓ | - | [81,88,89] | ||
| Streptococcus thermophiles | occludin delocalization↓ | - | [84,88,89] | ||
| HMO | ZO-1↑ | IL-10↑ | Mucus production↑ | [109,123,130,136] | |
| occludin↑ | TLR-4↓ | HIF-1α↓ | |||
| JAM-A↑ | NF-κB translocation↓ | Cleaved caspase-3↓ | |||
| Crypt proliferation↑ | p38 MAPK activation↓ | EGFR activation↑ | |||
| Intestinal permeability↓ | |||||
| Pre-biotics | GOS | TEER↑ | IL-6 mRNA↓ | HSP expression↓ | [99,100,102,103,119] |
| Intestinal permeability↓ | IL-8 mRNA↓ | Populations of probiotics↑ | |||
| occludin↑ | TLR-4↓ | HO-1 expression↓ | |||
| claudin-3↑ | IL-33↓ | ||||
| E-cadherin↑ | CXCL-8↓ | ||||
| CXCL-1↓ | |||||
| CXCL-2↓ | |||||
| MOS | Intestinal permeability↓ | - | Goblet cells↑ | [106,107,108] | |
| permeability↓ | Populations of probiotics↑ | ||||
| Villus height↑ | E. coli load↑ | ||||
| COS | Intestinal permeability↓ | - | - | [106,107] | |
| Morphological damage↓ | |||||
| FOS | TEER↑ | Colonic SCFA concentration↑ | [110,115,137] | ||
| Intestinal permeability↓ | - | Mucosal damage↓ | |||
| occludin↑ | |||||
| ZO-1↑ | |||||
| Chitosan oligosaccharides | TEER↑ | IL-6↓ | GST↑ | [111,119,132,133] | |
| Epithelial degeneration↓ | TNF-α↓ | ||||
| TJ proteins redistribution and distortion↓ | COX-2 activation↓ | ||||
| iNOS↓ | |||||
| NO production↓ | |||||
| NF-κB translocation↓ | |||||
| Compound | Integrity | Immunomodulation | Other Effect(s) | Reference |
|---|---|---|---|---|
| α-lipoic acid | Intestinal permeability↓ | COX-2 activation↓ | Epithelial proliferation↑ | [144,145,146,147,148,150,151,152,153,154] |
| ZO-1↑ | IL-17↓ | HSP70 expression↑ | ||
| occludin↑ | IL-6↓ | HO-1 activation↑ | ||
| E-cadherin↑ | TNF-α↓ | |||
| Morphological damage↓ | IκB activation↑ | |||
| Resveratrol | ZO-1↑ | IL-6 mRNA↓ | MDA↓ | [155,156,160,161,162,163,165,168,169] |
| occludin↑ | IL-1β mRNA↓ | SOD↑ | ||
| TEER↑ | PTGS1 mRNA↓ | GSH↓ | ||
| Intestinal permeability↓ | COX-2 activation↓ | ROS↓ | ||
| claudin-1↑ | NF-κB activation↓ | HO-1 activation↑ | ||
| claudin-4↑ | HSP70↑ | |||
| Crypt depth↓ | HSP90↑ | |||
| Villus height↑ |
| Compound | Integrity | Immunomodulation | Other Effect(s) | Reference |
|---|---|---|---|---|
| EPA and DHA | TEER↑ | Acute inflammation↓ | Mucosal damage↓ | [172,174,175,176,177,179,180,181,182] |
| Intestinal permeability↓ | IL-1β↓ | ROS production↓ | ||
| occludin↑ | IL-6↓ | SOD↑ | ||
| ZO-1↑ | IL-17↓ | CAT↑ | ||
| E-cadherin↑ | TNF-α↓ | Total nitrate/nitrite ratio↓ | ||
| TJ proteins redistribution and distortion↓ | INF-γ↓ | Microbiota composition restore↑ | ||
| COX-2 activation↓ | MUC-2 gene↑ | |||
| iNOS↓ | Cytokeratin gene↑ | |||
| cGMP↓ |
| Compound | Integrity | Immunomodulation | Other Effect(s) | Reference |
|---|---|---|---|---|
| Arginine | Intestinal permeability↓ | iNOS↑ | Intestinal necrosis↓ | [35,207,208,209,218,223,224] |
| TEER↑ | Intestinal s-IgA↑ | Mucus production and fluid secretion↑ | ||
| ZO-1↑ | NF-κB activation↓ | |||
| E-cadherin↑ | Pro-inflammatory cytokines↓ | |||
| pro-inflammatory chemokines↓ | ||||
| Villus height↑ | ||||
| Glutamine | Intestinal permeability↓ | NF-κB activation↑ | Mucus production↑ | [188,235,236,237,238,239,240,241,242,243,245,246,249,250,251,252,253,254] |
| Villus atrophy↓ | CD2+ and CD4+ lymphocytes↑ | HSP70 expression↑ | ||
| CD4+/CD8+↑ | HSF-1 expression↑ | |||
| occludin↑ | Serum IgA and IgG↑ | HO-1 expression↑ | ||
| claudin-1↑ | Intestinal mucosal s-IgA↑ | Cell viability and antioxidant capacity↑ | ||
| claudin-4↑ | TNF-α↓ | Hyperthermia↓ | ||
| JAM-A↑ | D-lactate↓ | Diarrhea occurrence↓ | ||
| ZO-1, ZO-2 and ZO-3↑ | DAO activity↓ | |||
| E-cadherin↑ | sICAM-1↓ | |||
| β-catenin↑ | IL-6↓ | |||
| IL-8↓ | ||||
| IL-10↑ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lian, P.; Braber, S.; Garssen, J.; Wichers, H.J.; Folkerts, G.; Fink-Gremmels, J.; Varasteh, S. Beyond Heat Stress: Intestinal Integrity Disruption and Mechanism-Based Intervention Strategies. Nutrients 2020, 12, 734. https://doi.org/10.3390/nu12030734
Lian P, Braber S, Garssen J, Wichers HJ, Folkerts G, Fink-Gremmels J, Varasteh S. Beyond Heat Stress: Intestinal Integrity Disruption and Mechanism-Based Intervention Strategies. Nutrients. 2020; 12(3):734. https://doi.org/10.3390/nu12030734
Chicago/Turabian StyleLian, Puqiao, Saskia Braber, Johan Garssen, Harry J. Wichers, Gert Folkerts, Johanna Fink-Gremmels, and Soheil Varasteh. 2020. "Beyond Heat Stress: Intestinal Integrity Disruption and Mechanism-Based Intervention Strategies" Nutrients 12, no. 3: 734. https://doi.org/10.3390/nu12030734
APA StyleLian, P., Braber, S., Garssen, J., Wichers, H. J., Folkerts, G., Fink-Gremmels, J., & Varasteh, S. (2020). Beyond Heat Stress: Intestinal Integrity Disruption and Mechanism-Based Intervention Strategies. Nutrients, 12(3), 734. https://doi.org/10.3390/nu12030734